Effects of Chemotherapy and Radiotherapy on Spermatogenesis: The Role of Testicular Immunology


Abstract
:1. Introduction and Immunological Tolerance of the Testis
2. Chemotherapy
3. Radiotherapy
4. Conclusion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ASA | antisperm antibody |
| BTB | blood–testis barrier |
| FSH | follicle-stimulating hormone |
| MCP1 | macrophage chemotactic protein 1 |
| ROS | reactive oxygen species |
| Sat2 | spermidine/spermine N1-acetyltransferase 2 |
| TBI | total body irradiation |
| TLR | Toll-like receptor |
| TNF | tumor necrosis factor |
| ZO-1 | zonula occludens-1 |
References
- Head, J.R.; Billingham, R.E. Immune privilege in the testis. II. Evaluation of potential local factors. Transplantation 1982, 40, 269–295. [Google Scholar] [CrossRef]
- Pollanen, P.; Maddocks, S. Macrophages, lymphocytes and MHC II antigen in the ram and rat testis. J. Reprod. Fertil. 1988, 82, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, C.R.; Law, L.; Magnuson, J.A.; Griswold, M.D.; Magnuson, N.S. Suppression of lymphocyte proliferation by cultured Sertoli cells. J. Reprod. Immunol. 1988, 14, 27–40. [Google Scholar] [CrossRef]
- Itoh, M.; Terayama, H.; Naito, M.; Ogawa, Y.; Tainosho, S. Tissue microcircumstances for leukocytic infiltration into the testis and epididymis in mice. J. Reprod. Immunol. 2005, 67, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, S.; Meinhardt, A. The macrophages in testis function. J. Reprod. Immunol. 2017, 119, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Grossman, C. Interactions between the gonadal steroid and the immune system. Science 1985, 227, 157. [Google Scholar] [CrossRef]
- Russell, L.; Brinster, R. Ultrastructural observations of spermatogenesis following transplantation or rat testis cells into mouse seminiferous tubules. J. Androl. 1996, 17, 615–627. [Google Scholar]
- Bellgrau, D.; Gold, D.; Selawry, H.; Moore, J.; Franzusoff, A.; Duke, R.C. A role for CD95 ligand in preventing graft rejection. Nature 1995, 377, 630–632. [Google Scholar] [CrossRef]
- D’Alessio, A.; Riccioli, A.; Lauretti, P.; Padula, F.; Muciaccia, B.; De Cesaris, P. Testicular FasL is expressed by sperm cell. Proc. Natl. Acad. Sci. USA 2001, 98, 3316–3321. [Google Scholar] [CrossRef]
- O’Connell, J.; O’Sullivan, G.; Collins, K.; Shanahan, F. The Fas counter attack: Fas mediated T cell killing by colon cancer cells expressing Fas-ligand. J. Exp. Med. 1996, 184, 1075–1082. [Google Scholar] [CrossRef]
- Shimamoto, K.; Yamamoto, Y.; Sofikitis, N.; Miyagawa, I. Influence of anterior chamber eye-cells interferon-r and caspase inhibitor on the cellular immune reactions following transplantation of spermatogonia. J. Urol. 1999, 161, 345. [Google Scholar] [CrossRef]
- Watanabe, M.; Kashiwakura, Y.; Kusumi, N.; Tamayose, K.; Nasu, Y.; Nagai, A. Adeno-associated virus-mediated human IL-10 gene transfer suppresses the development of experimental autoimmune orchites. Gene Ther. 2005, 12, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Hedger, M.P.; Meinhardt, A. Cytokines and the immune-testicular axis. J. Reprod. Immunol. 2003, 58, 1–26. [Google Scholar] [CrossRef]
- Jacobo, P.; Guazzone, V.A.; Theas, M.S.; Lustig, L. Testicular autoimmunity. Autoimmun. Rev. 2011, 10, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Naito, M.; Terayama, H.; Hirai, S.; Qu, N.; Lustig, L.; Itoh, M. Experimental autoimmune orchitis as a model of immunological male infertility. Med. Mol. Morphol. 2012, 45, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Fijak, M.; Pilatz, A.; Hedger, M.P.; Nicolas, N.; Bhushan, S.; Michel, V.; Tung, K.S.K.; Schuppe, H.C.; Meinhardt, A. Infectious, inflammatory and ‘autoimmune’ male factor infertility: How do rodent models inform clinical practice? Hum. Reprod. Update 2018, 24, 416–441. [Google Scholar] [CrossRef] [PubMed]
- Meistirch, M.L. Effects of chemotherapy and radiotherapy on spermatogenesis in human. Fertil. Steril. 2013, 100, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Da Cunha, M.F.; Meistrich, M.L.; Fuller, L.M.; Cundiff, J.H.; Hagemeister, F.B.; Velasquez, W.S. Recovery of spermatogenesis after treatment for Hodgkin’s disease: Limiting dose of MOPP chemotherapy. J. Clin. Oncol. 1984, 2, 571–577. [Google Scholar] [CrossRef]
- Watson, A.R.; Rance, C.P.; Bain, J. Long term effects of cyclophosphamide on testicular function. Br. Med. J. Clin. Res. Ed. 1985, 291, 1457–1460. [Google Scholar] [CrossRef]
- Meistrich, M.L.; Chawla, S.P.; da Cunha, M.F.; Johnson, S.L.; Plager, C.; Papadopoulos, N.E. Recovery of sperm production after chemotherapy for osteosarcoma. Cancer 1989, 63, 2115–2123. [Google Scholar] [CrossRef] [Green Version]
- Pryzant, R.M.; Meistrich, M.L.; Wilson, G.; Brown, B.; McLaughlin, P. Long-term reduction in sperm count after chemotherapy with and without radiation therapy for non-Hodgkin’s lymphomas. J. Clin. Oncol. 1993, 11, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Rivkees, S.A.; Crawford, J.D. The relationship of gonaldal activity and chemotherapy-induced gonadal damage. JAMA 1988, 259, 2123–2125. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.E.; Hawley, J.; Levy, W.; Gooley, T.; Buckner, C.D.; Deeg, H.J.; Doney, K.; Storb, R.; Sullivan, K.; Witherspoon, R.; et al. Pregnancies following high-dose cyclophosphamide with or without high-dose busulfan or total-body irradiation and bone marrow transplantation. Blood 1996, 87, 3045–3052. [Google Scholar] [PubMed]
- Buggia, I.; Locatelli, F.; Regazzi, M.B.; Zecca, M. Busulfan. Ann. Pharmacother. 1994, 28, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Galaup, A.; Paci, A. Pharmacology of dimethanesulfonate alkylating agents: Busulfan and treosulfan. Expert Opin. Drug Metab. Toxicol. 2013, 9, 333–347. [Google Scholar] [CrossRef] [PubMed]
- De Rooij, D.G.; Kramer, M.F. The effect of three alkylating agents on the seminiferous epithelium of rodents. I. Depletory effect. Virchows. Arch. B. Cell. Pathol. 1970, 4, 267–275. [Google Scholar] [PubMed]
- Shinohara, T.; Avarbock, M.R.; Brinster, R.L. Beta 1- and alpha 6-integrin are surface markers on mouse spermatogonial stem cells. Proc. Natl. Acad. Sci. USA 1999, 96, 5504–5509. [Google Scholar] [CrossRef]
- Ogawa, T. Spermatogonial transplantation: The principle and possible application. J. Mol. Med. 2001, 79, 368–374. [Google Scholar] [CrossRef]
- Perez-Crespo, M.; Pericuesta, E.; Perez-Cerezales, S.; Arenas, M.I.; Lobo, M.O.; Diaz-Gil, J.J. Effect of liver growth factor on both testicular regeneration and recovery of spermatogenesis in busulfan-treated mice. Reprod. Biol. Endocrinol. 2011, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Zohni, K.; Zhang, X.; Tan, S.L.; Chan, P.; Nagano, M.C. The efficiency of male fertility restoration is dependent on the recovery kinetics of spermatogonial stem cells after cytotoxic treatment with busulfan in mice. Hum. Reprod. 2012, 27, 44–53. [Google Scholar] [CrossRef]
- Gutierrez, K.; Glanzner, W.G.; Chemeris, R.O.; Rigo, M.L.; Comim, F.V.; Bordignon, V.; Gonçalves, P.B. Gonadotoxic effects of busulfan in two strains of mice. Reprod. Toxicol. 2016, 59, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Arechaga, J.M.; Avarbock, M.R.; Brinster, R.L. Transplantation of testis germinal cells into mouse seminiferous tubules. Int. J. Dev. Biol. 1997, 41, 111–122. [Google Scholar] [PubMed]
- Ogawa, T.; Ohmura, M.; Yumura, Y.; Sawada, H.; Kubota, Y. Expansion of murine spermatogonial stem cells through serial transplantation. Biol. Reprod. 2003, 68, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.X.; Short, R.V. Male germ cell transplantation in rats: Apparent synchronization of spermatogenesis between host and donor seminiferous epithelia. Int. J. Androl. 1995, 18, 326–330. [Google Scholar] [CrossRef]
- Zhang, Z.; Renfree, M.B.; Short, R.V. Successful intra- and interspecific male germ cell transplantation in the rat. Biol. Reprod. 2003, 68, 961–967. [Google Scholar] [CrossRef]
- Honaramooz, A.; Behboodi, E.; Hausler, C.L.; Blash, S.; Ayres, S.; Azuma, C.; Echelard, Y.; Dobrinski, I. Depletion of endogenous germ cells in male pigs and goats in preparation for germ cell transplantation. J. Androl. 2005, 26, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Bao, J.; Kong, Q.; Bai, Y.; Luo, F.; Zhou, S.; Wu, Y.; Huang, J. Effective production of recipient male pigs for spermatogonial stem cell transplantation by intratesticular injection with busulfan. Theriogenology 2017, 89, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Hur, T.Y.; Lee, S.H.; Ock, S.A.; Song, H.; Park, H.J.; Lee, R.; Sung, S.H.; Jhun, H.; Lee, W.Y. Dose-dependent effects of busulfan on dog testes in preparation for spermatogonial stem cell transplantation. Lab. Anim. Res. 2017, 33, 264–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vahdati, A.; Fathi, A.; Hajihoseini, M.; Aliborzi, G.; Hosseini, E. The regenerative effect of bone marrow-derived stem cells in spermatogenesis of infertile hamster. World J. Plast. Surg. 2017, 6, 18–25. [Google Scholar] [PubMed]
- Hermann, B.P.; Sukhwani, M.; Lin, C.C.; Sheng, Y.; Tomko, J.; Rodriguez, M.; Shuttleworth, J.J.; McFarland, D.; Hobbs, R.M.; Pandolfi, P.P.; et al. Characterization, cryopreservation, and ablation of spermatogonial stem cells in adult rhesus macaques. Stem Cells 2007, 25, 2330–2338. [Google Scholar] [CrossRef]
- Ghadimi, F.; Shakeri, M.; Zhandi, M.; Zaghari, M.; Piryaei, A.; Moslehifar, P.; Rajabinejad, A. Different approaches to establish infertile rooster. Anim. Reprod. Sci. 2017, 186, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Bhartiya, D.; Sriraman, K.; Mallick, A. Underlying mechanisms that restore spermatogenesis on transplanting healthy niche cells in busulphan treated mouse testis. Stem Cell Rev. Rep. 2016, 12, 682–697. [Google Scholar] [CrossRef] [PubMed]
- Bhartiya, D.; Anand, S. Effects of oncotherapy on testicular stem cells and niche. Mol. Hum. Reprod. 2017, 23, 654–655. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Ok, D.W.; Kwon, D.N.; Chung, J.I.; Kim, H.C.; Yeo, S.M.; Kim, T.; Seo, H.G.; Kim, J.H. Murine male germ cell apoptosis induced by busulfan treatment correlates with loss of c-kit-expression in a Fas/FasL- and p53-independent manner. FEBS Lett. 2004, 575, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; He, X.; Zhuang, M.; Niu, B.; Wu, C.; Mu, H.; Tang, F.; Cui, Y.; Liu, W.; Zhao, B.; et al. Melatonin ameliorates busulfan-induced spermatogonial stem cell oxidative apoptosis in mouse testes. Antioxid. Redox Signal. 2018, 28, 385–400. [Google Scholar] [CrossRef] [PubMed]
- Xian, Y.; Wu, M.; Liu, Y.; Hao, J.; Wu, Y.; Liao, X.; Li, G. Increased Sat2 expression is associated with busulfan-induced testicular Sertoli cell injury. Toxicol. In Vitro 2017, 43, 47–57. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, T.; Deng, T.; Xiong, W.; Lui, P.; Li, N.; Chen, Y.; Han, D. Damaged spermatogenic cells induce inflammatory gene expression in mouse Sertoli cells through the activation of Toll-like receptors 2 and 4. Mol. Cell. Endocrinol. 2013, 365, 162–173. [Google Scholar] [CrossRef]
- Qu, N.; Kuramasu, M.; Hirayanagi, Y.; Nagahori, K.; Hayashi, S.; Ogawa, Y.; Terayama, H.; Suyama, K.; Naito, M.; Sakabe, K.; et al. Gosha-Jinki-Gan recovers spermatogenesis in mice with busulfan-induced aspermatogenesis. Int. J. Mol. Sci. 2018, 19, 2606. [Google Scholar] [CrossRef]
- Piccinini, A.M.; Midwood, K.S. DAMPening inflammation by modulating TLR signalling. Mediat. Inflamm. 2010, 2010, 672395. [Google Scholar] [CrossRef]
- Yu, L.; Wang, L.; Chen, S. Endogenous toll-like receptor ligands and their biological significance. J. Cell. Mol. Med. 2010, 14, 2592–2603. [Google Scholar] [CrossRef] [Green Version]
- Bhushan, S.; Tchatalbachev, S.; Lu, Y.; Frohlich, S.; Fijak, M.; Vijayan, V.; Chakraborty, T.; Meinhardt, A. Differential activation of inflammatory pathways in testicular macrophages provides a rationale for their subdued inflammatory capacity. J. Immunol. 2015, 194, 5455–5464. [Google Scholar] [CrossRef] [PubMed]
- Sinisi, A.A.; D’Apuzzo, A.; Pasquali, D.; Venditto, T.; Esposito, D.; Pisano, G.; De Bellis, A.; Ventre, I.; Papparelia, A.; Perrone, L.; et al. Antisperm antibodies in prepubertal boys treated with chemotherapy for malignant or non-malignant diseases and in boys with genital tract abnormalities. Int. J. Androl. 1997, 20, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Song, H.; Kwon, D.N.; Cho, S.K.; Kang, S.J.; Yoe, S.M.; Kim, H.C.; Lee, H.T.; Park, C.; Kim, J.H. Significant IgG-immunoreactivity of the spermatogonia of the germ cell-depleted testis after busulfan treatment. Anim. Reprod. Sci. 2006, 91, 317–335. [Google Scholar] [CrossRef] [PubMed]
- Qu, N.; Naito, M.; Li, J.; Terayama, H.; Hirai, S.; Itoh, M. Xenogeneic and endogenous spermatogenesis following transplantation of rat germ cells into testes of immunocompetent mice. Reprod. Fertil. Dev. 2012, 24, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Hirayanagi, Y.; Qu, N.; Hirai, S.; Naito, M.; Terayama, H.; Hayashi, S.; Hatayama, N.; Kuramasu, M.; Ogawa, Y.; Itoh, M. Busulfan pretreatment for transplantation of rat spermatogonia differentially affects immune and reproductive systems in male recipient mice. Anat. Sci. Int. 2015, 90, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Kuban, D.A.; Meistrich, M.L.; Cheung, R.; Hussey, D.H. Chapter 27—The Testicle. In Radiation Oncology, 9th ed.; Cox, J.D., Ang, K.K., Eds.; Mosby Elsevier: Philadelphia, PA, USA, 2010; pp. 653–675. [Google Scholar]
- Abuelhija, M.; Weng, C.C.; Shetty, G.; Meistrich, M.L. Rat models of post-irradiation recovery of spermatogenesis: Interstrain differences. Andrology 2013, 1, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Osterberg, E.C.; Ramasamy, R.; Masson, P.; Brannigan, R.E. Current practices in fertility preservation male cancer patients. Urol. Ann. 2014, 6, 13–17. [Google Scholar] [CrossRef]
- Howell, S.J.; Shalet, S.M. Spermaotgenesis after cancer treatment: Damage and recovery. J. Natl. Cancer Inst. Monogr. 2005, 34, 12–17. [Google Scholar] [CrossRef]
- Centola, G.M.; Keller, J.W.; Henzler, M.; Rubin, P. Effect of low-dose testicular irradiation on sperm count and fertility in patients with testicular seminoma. J. Androl. 1994, 15, 608–613. [Google Scholar]
- Kinsella, T.J.; Trivette, G.; Rowland, J.; Sorace, R.; Miller, R.; Fraass, B. Long-term follow-up of testicular function following radiation therapy for early-stage Hodgkin’s disease. J. Clin. Oncol. 1989, 7, 718–724. [Google Scholar] [CrossRef]
- Vergouwen, R.P.; Huiskamp, R.; Bas, R.J.; Roepers-Gajadien, H.L.; de Jong, F.H.; van Eerdenburg, F.J.; Davids, J.A.; de Rooij, D.G. Radiosensitivity of testicular cells in the prepubertal mouse. Radiat. Res. 1994, 139, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Adhikari, J.S.; Rizvi, M.A.; Chaudhury, N.K. Radioprotective potential of melatonin against Co γ-ray-induced testicular injury in male C57BL/6 mice. J. Biomed. Sci. 2015, 22, 61. [Google Scholar] [CrossRef] [PubMed]
- Cordelli, E.; Fresegna, A.M.; Leter, G.; Eleuteri, P.; Spano, M.; Villani, P. Evaluation of DNA damage in different stages of mouse spermatogenesis after testicular X irradiation. Radiat. Res. 2003, 160, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Ergur, B.U.; Kiray, M.; Pekcetin, C.; Bagriyanik, H.A.; Erbil, G. Protective effect of erythropoietin pretreatment in testicular ischmiareperfusion injury in rats. J. Pediatr. Surg. 2008, 43, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Xu, B.; Yang, M.; Li, Z.; Zhang, Y.; Jiang, D. Protection by hydrogen against gamma ray-induced testicular damage in rats. Basic Clin. Pharmacol. Toxicol. 2013, 112, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Pino-Lataillade, G.; Velez de la Calle, J.F.; Viguier-Martinez, M.C. Influence of germ cells upon Sertoli cells during continuous low-dose rate gamma-irradiation of adult rats. Mol. Cell. Endocrinol. 1988, 58, 51–63. [Google Scholar] [CrossRef]
- Hasegawa, M.; Wilson, G.; Russell, L.D. Radiation-induced cell death in the mouse testis: Relationship to apoptosis. Radiat. Res. 1997, 147, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Park, J.S.; Kim, Y.J. Differential expression of Prx I and II in mouse testis and their up-regulation by radiation. Biochem. Biophys. Res. Commun. 2002, 296, 337–342. [Google Scholar] [CrossRef]
- Ma, Y.; Nie, H.; Sheng, C.; Chen, H.; Wang, B.; Liu, T.; Shao, J.; He, X.; Zhang, T.; Zheng, C.; et al. Roles of oxidative stress in synchrotron radiation X-ray-induced testicular damage of rodents. Int. J. Physiol. Pathophysiol. Pharmacol. 2012, 4, 108–114. [Google Scholar] [PubMed]
- Aktoz, T.; Caloglu, M.; Uurut-Caloglu, V.; Yalcin, O.; Aydogdu, N.; Vurlu, D.; Arda, E.; Inci, O. Histopathological and biochemical comparisons of the protective effects of amifostine and L-carnitine against radiation-induced acute testicular toxicity in rats. Andrologia 2017, 49, e12754. [Google Scholar] [CrossRef]
- Silva, A.M.; Correia, S.; Casalta-Lopes, J.E.; Mamede, A.C.; Cavaco, J.E.; Botelho, M.F.; Socorro, S.; Maia, C.J. The protective effect of regucalcin against radiation-induced damage in testicular cells. Life Sci. 2016, 164, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Nagahori, K.; Qu, N.; Kuramasu, M.; Hirayanagi, Y.; Hayashi, S.; Ogawa, Y.; Hatayama, N.; Terayama, H.; Suyama, K.; et al. The traditional Japanese medicine Goshajinkigan makes a recovery from irradiation-induced aspermatogenesis in mice. 2019; manuscript under review. [Google Scholar]
- Vergouwen, R.P.; Huiskamp, R.; Bas, R.J. Radiosensitivity of testicular cells in the fetal mouse. Radiat. Res. 1995, 141, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Guitton, N.; Brouazin-Jousseaume, V.; Dupaix, A.; Jegou, B.; Chenal, C. Radiation effect on rat Sertoli cell function in vitro and in vivo. Int. J. Radiat. Biol. 1999, 75, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, A.V.; Almeida, F.R.; Weng, C.C.; Shetty, G.; Meistrich, M.L.; Chiarini-Garcia, H. Spermatogonial behavior in rats during radiation-induced arrest and recovery after hormone suppression. Reproduction 2013, 146, 363–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kangasniemi, M.; Huhtaniemi, I.; Meistrich, M.L. Failure of spermatogenesis to recover despite the presence of a spermatogonia in the irradiated LBNF1 rat. Biol. Reprod. 1996, 54, 1200–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuttlesworth, G.A.; de Rooij, D.G.; Huhtaniemi, I.; Reissmann, T.; Russell, L.D.; Shetty, G.; Meistrich, M.L. Enhancement of a spermatogonial proliferation and differentiation in irradiated rats by gonadotropin-releasing hormone antagonist administration. Endocrinology 2000, 141, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Monsees, T.K.; Franz, M.; Gebhardt, S.; Winterstein, U.; Schill, W.B.; Hayatpour, J. Sertoli cells as a target for reproductive hazards. Andrologia 2000, 32, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Defamie, N.; Mograbi, B.; Roger, C.; Cronier, L.; Malassine, A.; Brucker-Davis, F.; Fenichel, P.; Segretain, D.; Pointis, G. Disruption of gap junctional intercellular communication by lindane is associated with aberrant localization of connexin43 and zonula occludens-1 in 42GPA9 Sertoli cells. Carcinogenesis 2001, 22, 1537–1542. [Google Scholar] [CrossRef] [Green Version]
- Fiorini, C.; Tilloy-Ellul, A.; Chevalier, S.; Charuel, C.; Pointis, G. Sertoli cell junctional proteins as early targets for different classes of reproductive toxicants. Reprod. Toxicol. 2004, 18, 413–421. [Google Scholar] [CrossRef]
- Wang, X.W.; Ding, G.R.; Shi, C.H.; Zeng, L.H.; Liu, J.Y.; Li, J.; Zhao, T.; Chen, Y.B.; Guo, G.Z. Mechanisms involved in the blood-testis barrier increased permeability induced by EMP. Toxicology 2010, 276, 58–63. [Google Scholar] [CrossRef]
- Son, Y.; Heo, K.; Bae, M.J.; Lee, C.G.; Cho, W.S.; Kim, S.D.; Yang, K.; Shin, I.S.; Lee, M.Y.; Kim, J.S. Injury to the blood-testis barrier after low-dose-rate chronic radiation exposure in mice. Radiat. Prot. Dosim. 2015, 167, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liu, T.; Shao, J.; Sheng, C.; Hong, Y.; Ying, W.; Xia, W. Antioxidant protects blood-testis barrier against synchrotron radiation X-ray-induced disruption. Spermatogenesis 2015, 5, e1009313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinhardt, A.; Hedger, M.P. Immunological, paracrine and endocrine aspects of testicular immune privilege. Mol. Cell. Endocrinol. 2011, 335, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.Y.; Mruk, D.D. The blood-testis barrier and its implications for male contraception. Pharmacol. Rev. 2012, 64, 16–64. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Furuse, M.; Fujimoto, K.; Tsukita, S. Claudin multigene family encoding four-transmembrane domain protein components of tight junction strands. Proc. Natl. Acad. Sci. USA 1999, 96, 511–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, K.; Sasaki, H.; Fujimoto, K.; Furuse, M.; Tsukita, S. Claudin-11/OSP-based tight junction of myelin sheaths in brain and Sertoli cells in testis. J. Cell Biol. 1999, 145, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Furuse, M.; Hirase, T.; Itoh, M.; Nagafuchi, A.; Yonemura, S.; Tsukita, S.; Tsukita, S. Occludin: A novel integral membrane protein localizing at tight junction. J. Cell Biol. 1993, 123, 1777–1788. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, B.R.; Siliciano, J.D.; Mooseker, M.S.; Goodenough, D.A. Identification of ZO-1: A high molecular weight polypeptide associated with the tight junction (zonula occludens) in a variety of epithelia. J. Cell Biol. 1986, 103, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Fanning, A.S.; Jameson, B.J.; Jesaitis, L.A.; Anderson, J.M. The tight junction protein ZO-1 establishes a link between the transmembrane protein occluding and the actin cytoskeleton. J. Biol. Chem. 1998, 273, 29745–29753. [Google Scholar] [CrossRef]
- Gow, A.; Southwood, C.M.; Li, J.S.; Pariali, M.; Riordan, G.P.; Brodie, S.E.; Danias, J.; Bronstein, J.M.; Kachar, B.; Lazzarini, R.A. CNS myelin and Sertoli cell tight junction strands are absent in Osp/claudin-11 null mice. Cell 1999, 99, 649–659. [Google Scholar] [CrossRef]
- Mazaud-Guittot, S.; Meugnier, E.; Pesenti, S.; Wu, X.; Vidal, H.; Gow, A.; Le Magueresse-Battistoni, B. Claudin 11 deficiency in mice results in loss of the Sertoli cell epithelial phenotype in the testis. Biol. Reprod. 2010, 82, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Saitou, M.; Furuse, M.; Sasaki, H.; Schulzke, J.D.; Fromm, M.; Takano, H.; Noda, T.; Tsukita, S. Complex phenotype of mice lacking occluding, a component of tight junction strands. Mol. Biol. Cell. 2000, 11, 4131–4142. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.H. Changes in the blood-testis barrier of the guinea-pig in relation to histological damage following iso-immunization with testis. J. Reprod. Fertil. 1970, 22, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Nistal, M.; Riestra, M.L.; Paniagua, R. Focal orchitis in undescended testes: Discussion of pathogenetic mechanisms of tubular atrophy. Arch. Pathol. Lab. Med. 2002, 126, 64–69. [Google Scholar] [PubMed]
- Naito, M.; Itoh, M. Patterns of infiltration of lymphocyte into the testis under normal and pathological conditions in mice. Am. J. Reprod. Immunol. 2008, 59, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Q.; Cheng, C.Y. A seamless trespass: Germ cell migration across the seminiferous epithelium during spermatogenesis. J. Cell. Biol. 2007, 178, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Shetty, G.; Meistrich, M.L. Hormonal approaches to preservation and restoration of male fertility after cancer treatment. J. Natl. Cancer Inst. Monogr. 2005, 2005, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Meistrich, M.L.; Shetty, G. Inhibition of spermatogonial differentiation by testosterone. J. Androl. 2003, 24, 135–148. [Google Scholar] [CrossRef] [PubMed]



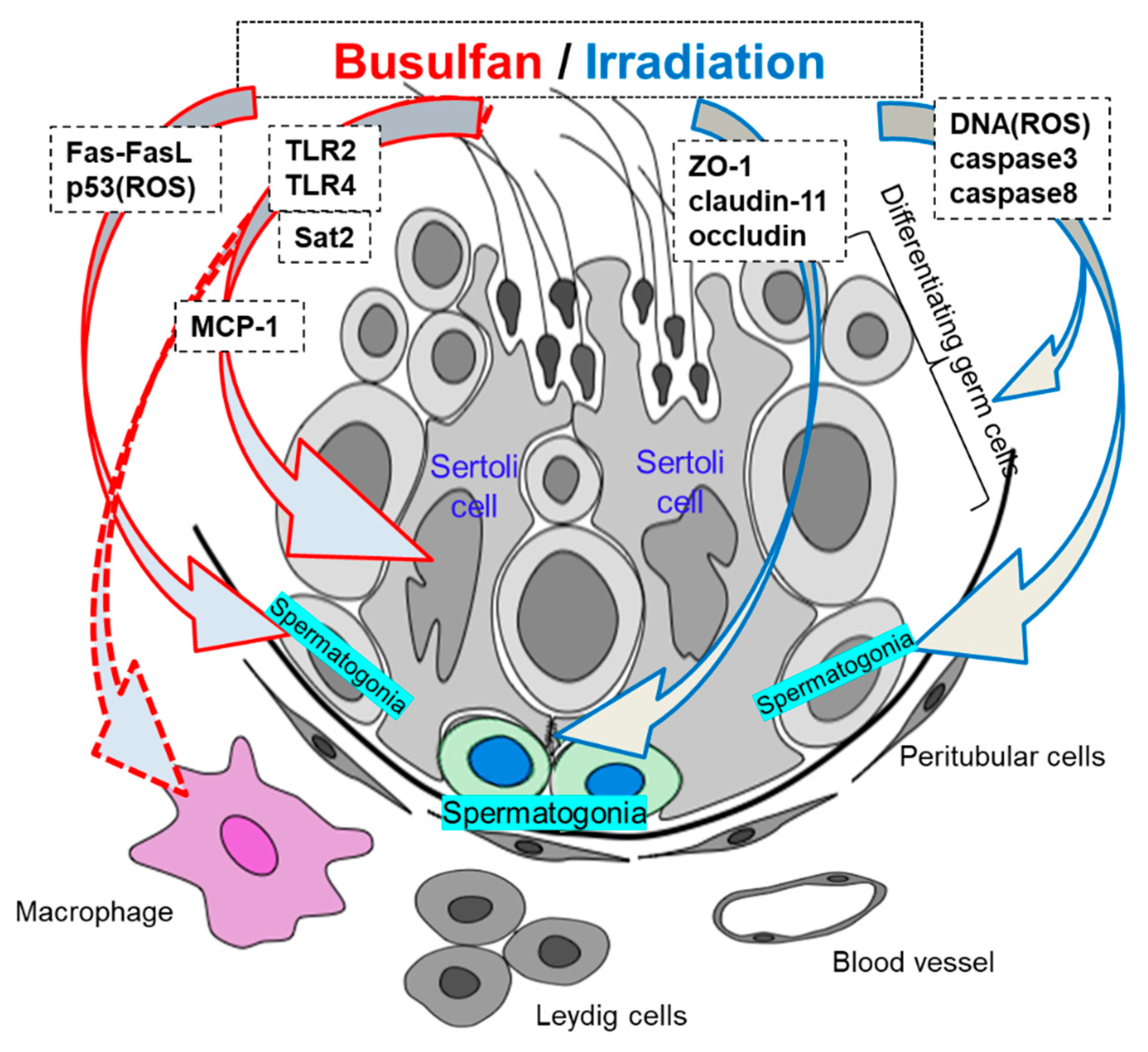{kind=link}
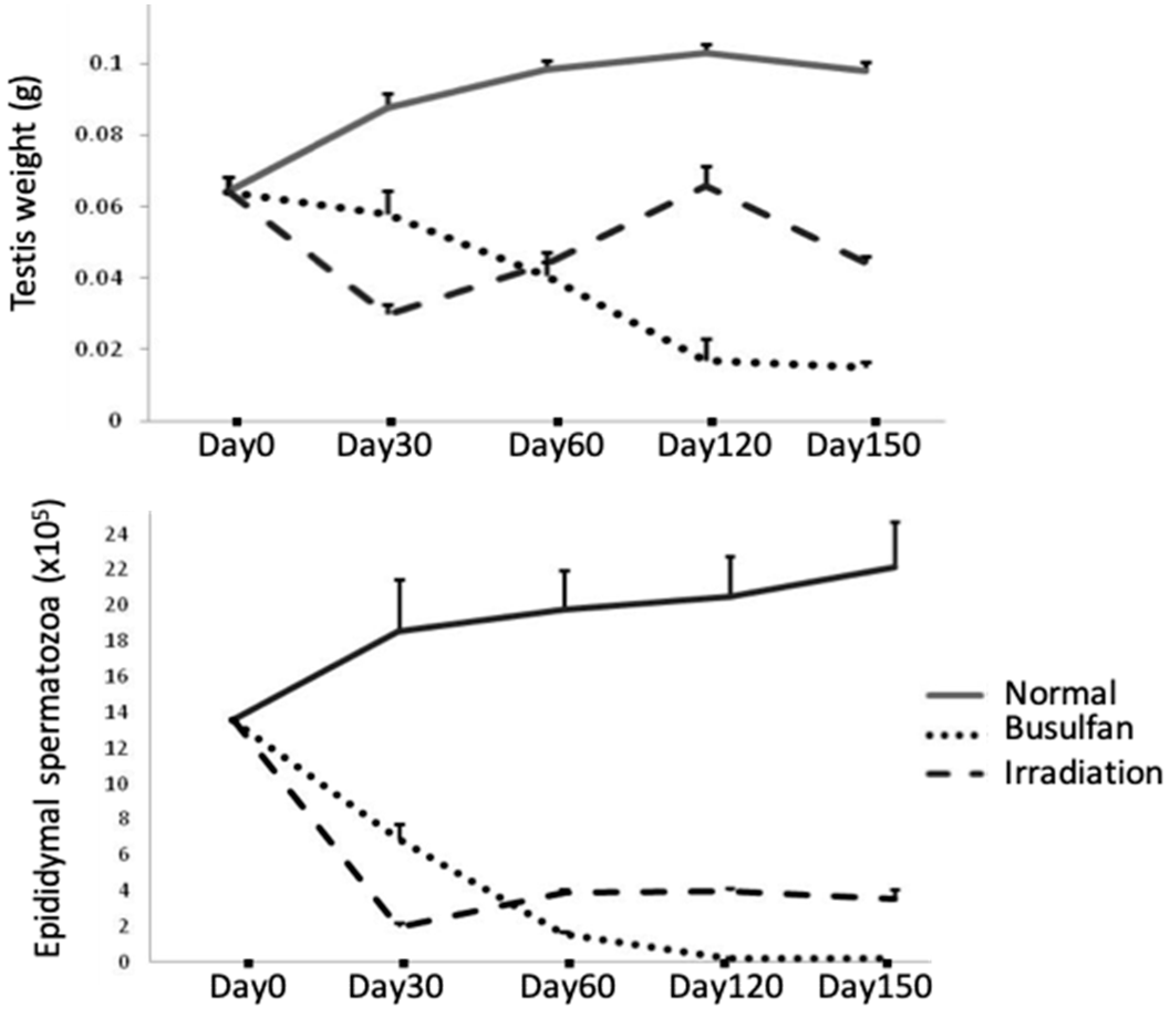{kind=link}
| Immunosuppressive Factors in Normal Testis | Local Function | Testicular Immunology Consequences after Busulfan-Treatment | Testicular Immunology Consequences after Irradiation-Treatment | Testicular Immunology Consequences in Autoimmune Orchitis | ||
|---|---|---|---|---|---|---|
| Germ cells | transforming growth factor β | Leydig cell steroidogenesis ↓ | ||||
| Fas ligand | apoptosis of Fas-bearing lymphocyte | ↑ or (-) | ||||
| interferon-γ | Leydig cell steroidogenesis ↓ | |||||
| tumor necrosis factor α | Leydig cell steroidogenesis ↓ or ↑ | |||||
| Fas ↑ caspase3-8 ↑ | Fas ↑ | caspase 3–8 ↑ | ||||
| Fas ↑ | apoptosis through oxidative stress | Bax ↑ | caspase 9 ↑ | |||
| p53-ROS ↑ caspase3 ↑ | DNA damage | |||||
| Sertoli cells | activin | mitogenesis of lymphocytes ↓ | ||||
| inhibin | mitogenesis of lymphocytes ↑ | |||||
| interleukin-6 | meiotic DNA synthesis of germ cell | ↑ ? | ↑ | |||
| Fas ligand | apoptosis of Fas-bearing lymphocyte | |||||
| transforming growth factor β | inhibin secretion ↑ | |||||
| TNFα↑ MCP-1 ↑ TLR2,4 ↑ | ZO-1, occludin, claudin-11 ↓ | occludin, claudin-11 ↓ | ||||
| Sat2 ↑ | ||||||
| Leydig cells | testosterone | ↓ | ↓ | ↓ | ||
| protein S | Leydig cell steroidogenesis ↓ | |||||
| insulin-like growth factor-1 | testosterone secretion ↑ | |||||
| Fas ligand | apoptosis of Fas-bearing germ cell | |||||
| interleukin-10 | immune privilege | |||||
| transforming growth factor β | contractility of myoid cell | |||||
| Leydig cell apoptosis | ||||||
| Testicular | interleukin-10 | inhibition of T cell-mediated immune response response | ||||
| macrophages | interferon-γ | Fas ligand expression by Sertoli cell ↑ | ↑ | |||
| interleukin-6 | radioprotection of germ cell by Sertoli cell | ↑ ? | ↑ | |||
| tumor necrosis factor α | Fas ligand expression by Sertoli cell ↓ or ↑ | ↑ | ↑ | |||
| macrophage infiltration (+) | macrophage infiltration (-) | macrophage infiltration (+) | ||||
| Others | ASA? | ASA (+) | T cells·B cells infiltration (+) | |||
| ASA (+) | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, N.; Itoh, M.; Sakabe, K. Effects of Chemotherapy and Radiotherapy on Spermatogenesis: The Role of Testicular Immunology. Int. J. Mol. Sci. 2019, 20, 957. https://doi.org/10.3390/ijms20040957
Qu N, Itoh M, Sakabe K. Effects of Chemotherapy and Radiotherapy on Spermatogenesis: The Role of Testicular Immunology. International Journal of Molecular Sciences. 2019; 20(4):957. https://doi.org/10.3390/ijms20040957
Chicago/Turabian StyleQu, Ning, Masahiro Itoh, and Kou Sakabe. 2019. "Effects of Chemotherapy and Radiotherapy on Spermatogenesis: The Role of Testicular Immunology" International Journal of Molecular Sciences 20, no. 4: 957. https://doi.org/10.3390/ijms20040957
APA StyleQu, N., Itoh, M., & Sakabe, K. (2019). Effects of Chemotherapy and Radiotherapy on Spermatogenesis: The Role of Testicular Immunology. International Journal of Molecular Sciences, 20(4), 957. https://doi.org/10.3390/ijms20040957