Role of Histamine H3 Receptor Antagonists on Intraocular Pressure Reduction in Rabbit Models of Transient Ocular Hypertension and Glaucoma

 , and
, and
Abstract
:
1. Introduction
2. Results
2.1. Histamine Receptor Protein and mRNA Expression in the Eye of New Zealand White Rabbits
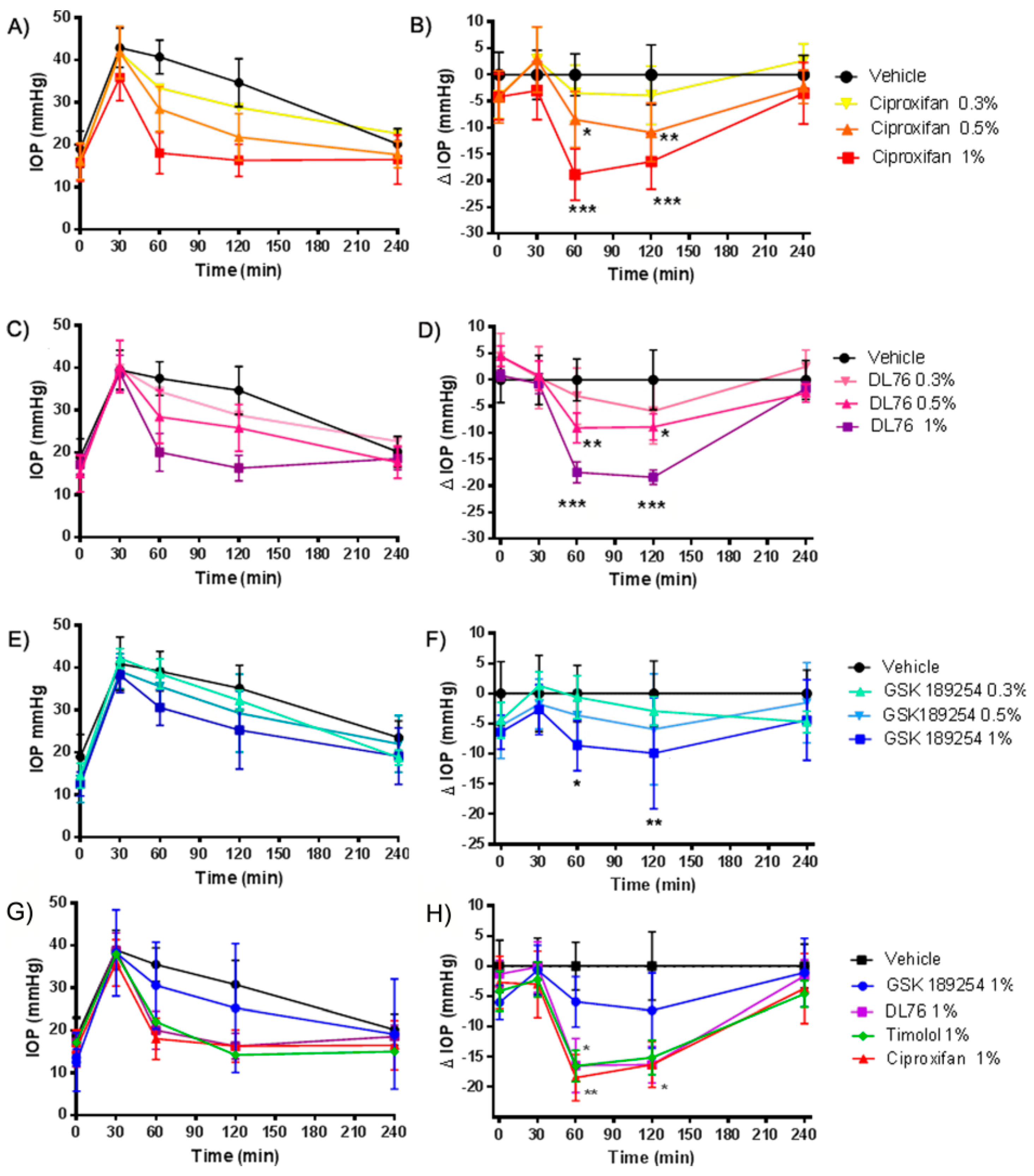
2.2. Pharmacological Studies of H3R Antagonists in Transient Ocular Hypertension Model
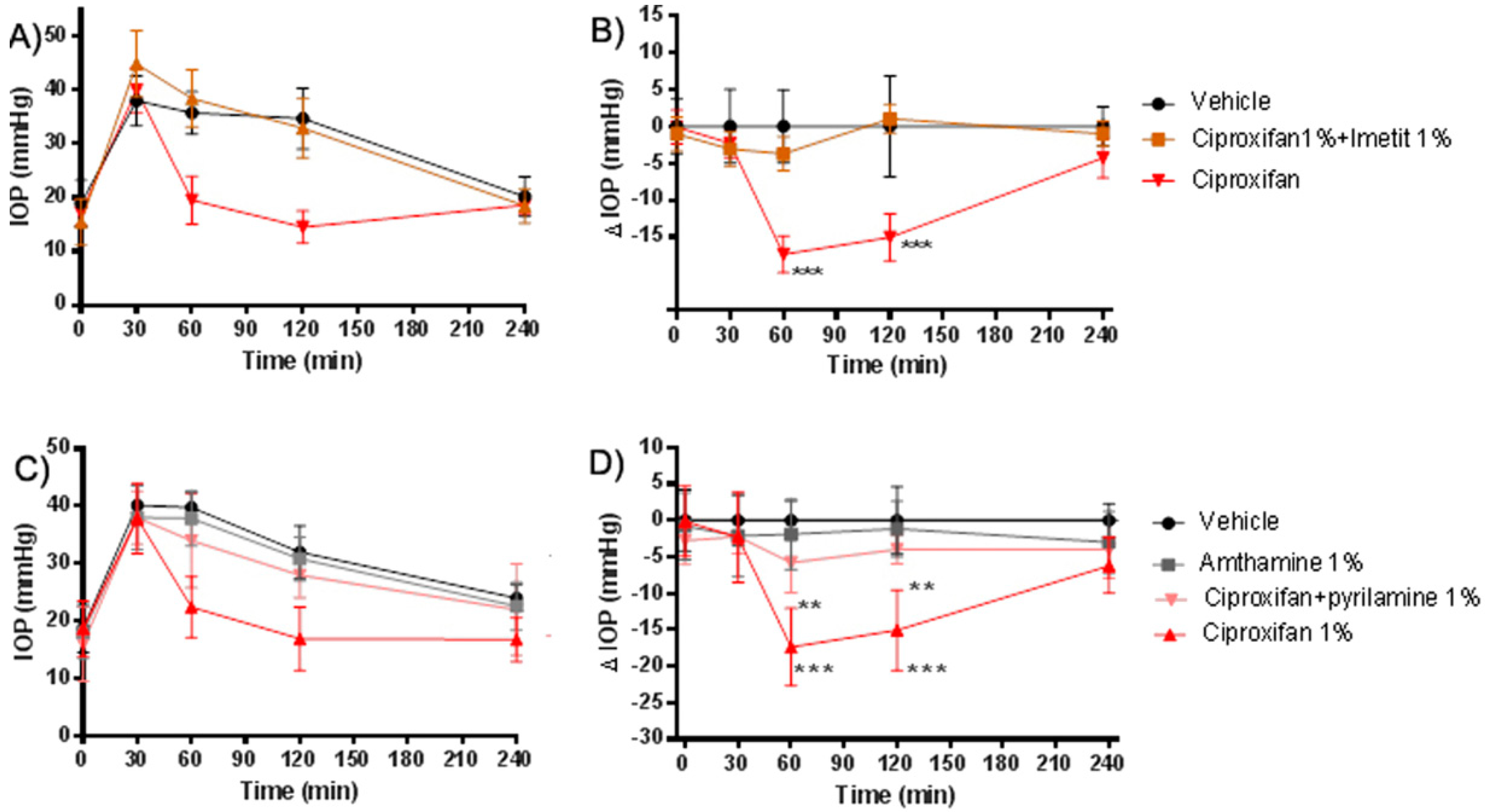
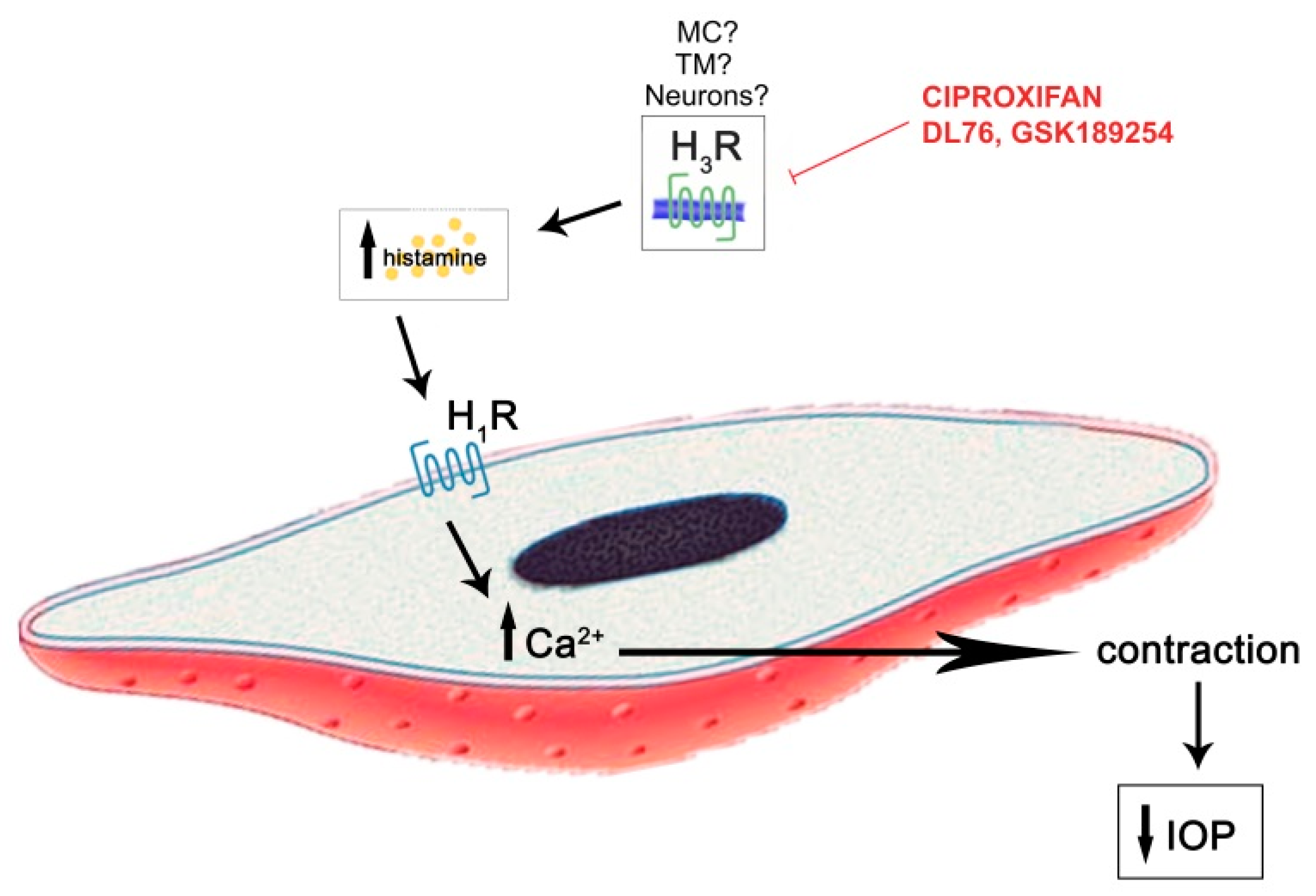
2.3. Evaluation of Specificity of H3R Antagonistic Action
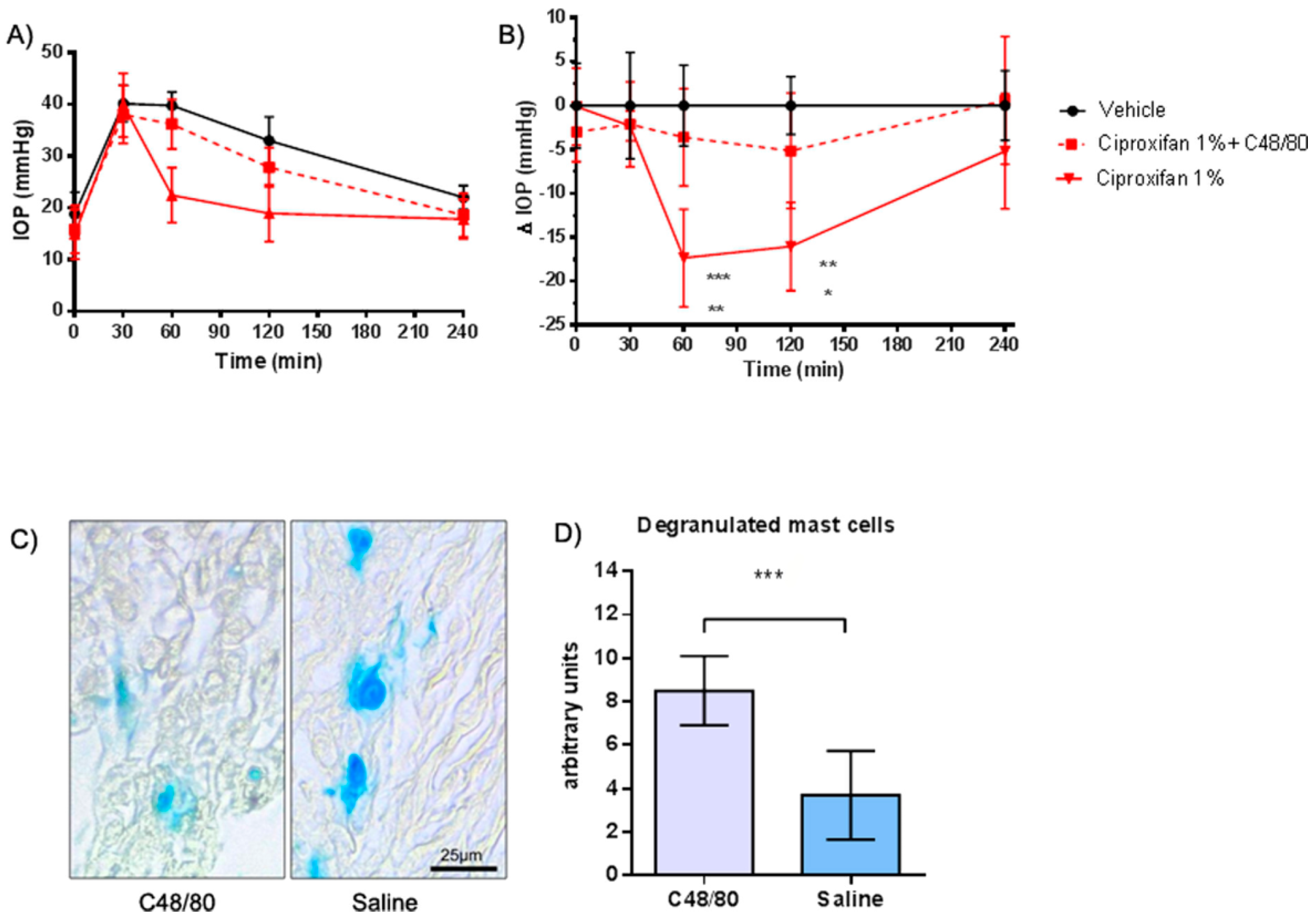
2.4. Evaluation of the IOP Profile after Choroidal Mast Cell Granule Depletion by Compound 48/80 Pre-Treatment
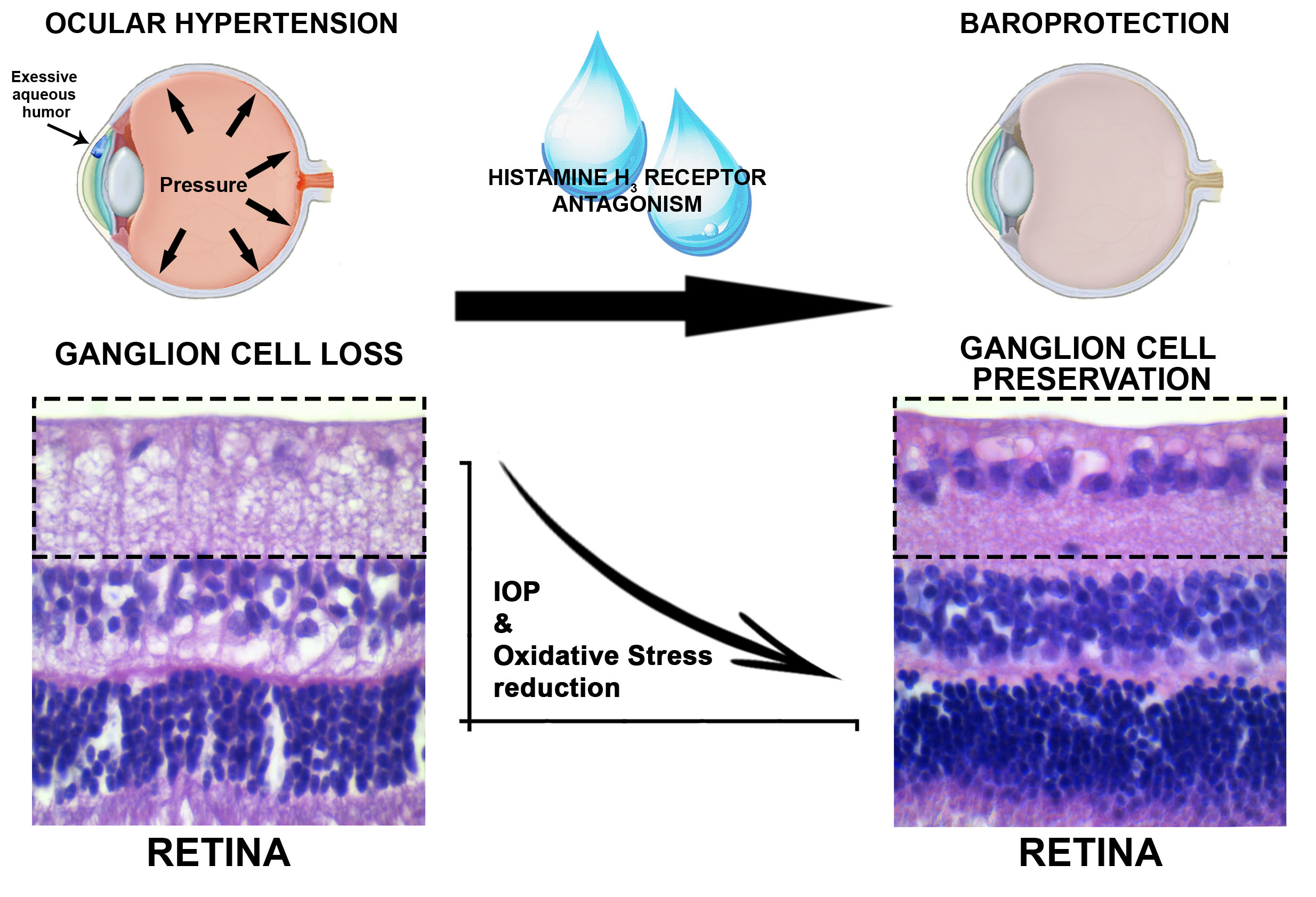
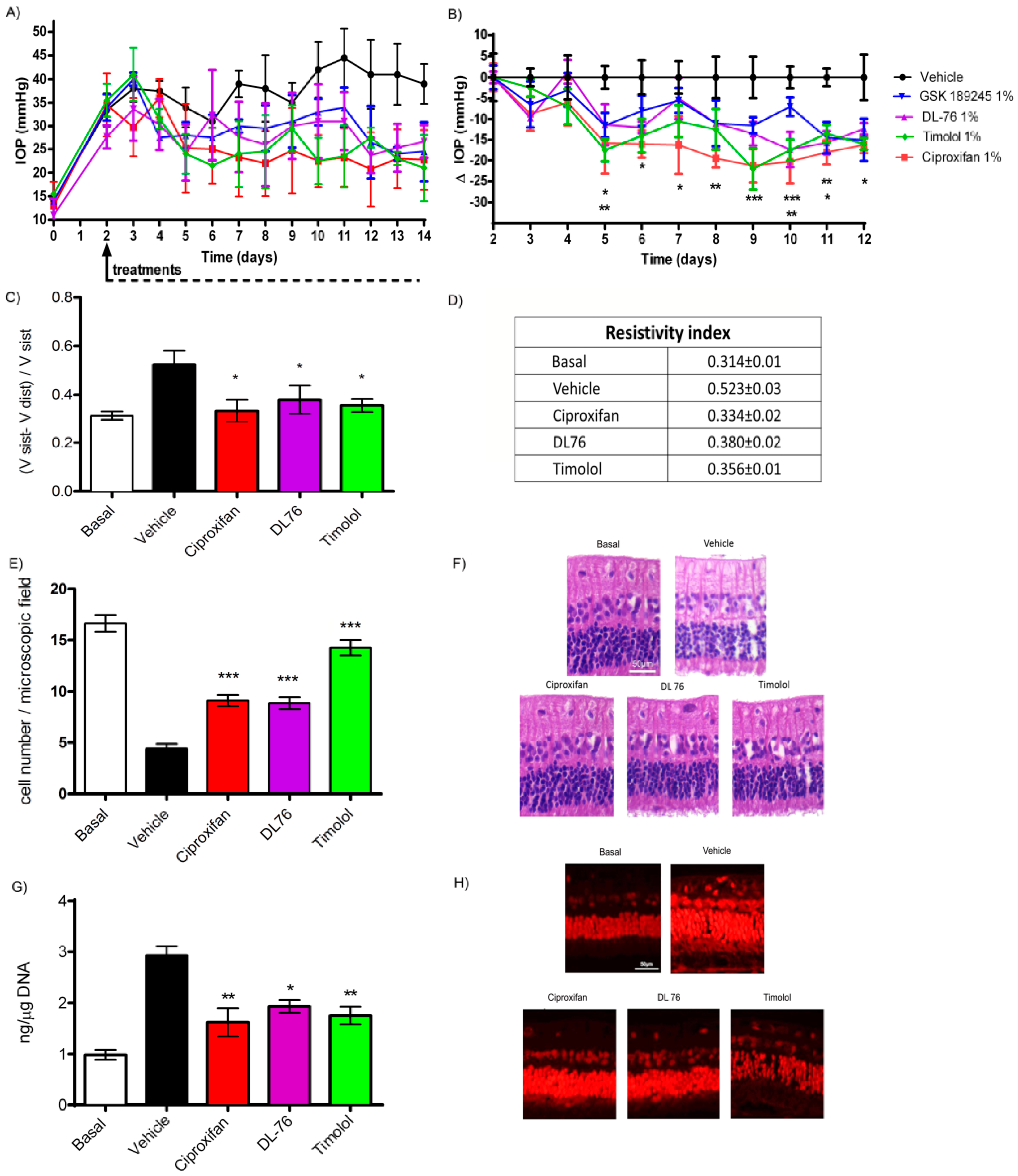
2.5. Effects of Histamine H3R Antagonists on IOP, Ocular Vascular Dynamics, Retinal Ganglion Cell Conservation and Oxidative Stress in Glaucoma Model
3. Discussion
4. Materials and Methods
4.1. Animal Models of Glaucoma
4.2. Rabbit Models of Ocular Hypertension and Glaucoma
4.3. IOP Measurements
4.4. Compounds Used in the Experiments
4.5. Pharmacological Evaluation of H3R Antagonists on IOP Reduction in Transient OHT Model: Dose/Effect and Specificity
4.6. Pharmacological Evaluation of H3R Antagonists on IOP Reduction and Baroprotection in Glaucoma Model
4.7. Sample Collection
4.8. Western Blot Analysis of Histamine Receptors
4.9. Semi-Quantitative RT-PCR of Histamine Receptors
4.10. Immunofluorescence of Ciliary Bodies and Retinae
4.11. Determination of 8-Hydroxy-2′-deoxyguanosine
4.12. Fluorescent Dye Dihydroethidium (DHE) Staining
4.13. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| RGC | retinal ganglion cells |
| OHT | ocular hypertension |
| IOP | intraocular pressure |
| DHE | dihydroethidium |
| TM | trabecular meshwork |
| HR | histamine receptor |
| MC | mast cells |
| 8OHdG | 8-hydroxy-2′-deoxy-guanosine |
| DAPI | 4′,6′-diamidino-2-phenylindole |
| RI | resistivity index |
| CDI | color doppler imaging |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
- Quigley, H.A.; Broman, A.T. The number of people with glaucoma worldwide in 2010 and 2020. Br. J. Ophthalmol. 2006, 90, 262–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Huang, C.; Zhang, H.; Wu, R. Autophagy in glaucoma: Crosstalk with apoptosis and its implications. Brain Res. Bull. 2015, 117, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Varma, R.; Lee, P.P.; Goldberg, I.; Kotak, S. An assessment of the health and economic burdens of glaucoma. Am. J. Ophthalmol. 2011, 152, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Matlach, J.; Bender, S.; König, J.; Binder, H.; Pfeiffer, N.; Hoffmann, E.M. Investigation of intraocular pressure fluctuation as a risk factor of glaucoma progression. Clin. Ophthalmol. 2018, 13, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Nau, C.B.; Malihi, M.; McLaren, J.W.; Hodge, D.O.; Sit, A.J. Circadian variation of aqueous humor dynamics in older healthy adults. Invest. Ophthalmol. Vis. Sci. 2013, 54, 7623–7629. [Google Scholar] [PubMed]
- Smith, S.D.; Gregory, D.S. A circadian rhythm of aqueous flow underlies the circadian rhythm of IOP in NZW rabbits. Invest. Ophthalmol. Vis. Sci. 1989, 30, 775–778. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-S.; Sergeeva, O.A.; Haas, H.L. Histamine H3 Receptors and Sleep-Wake Regulation. J. Pharmacol. Exp. Ther. 2011, 336, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Gastinger, M.J.; Tian, N.; Horvath, T.; Marshak, D.W. Retinopetal Axons in Mammals: Emphasis on Histamine and Serotonin. Curr. Eye Res. 2006, 31, 655–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markwardt, K.L.; Magnino, P.E.; Pang, I.H. Histamine induced contraction of human ciliary muscle cells. Exp. Eye Res. 1997, 64, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Caprioli, J.; Coleman, A.L. Intraocular Pressure Fluctuation. Ophthalmology 2008, 115, 1123.e3–1129.e3. [Google Scholar] [CrossRef] [PubMed]
- Marshall, I. Characterization and distribution of histamine H1- and H2-receptors in precapillary vessels. J. Cardiovasc. Pharmacol. 1984, 6 (Suppl. 4), S587–S597. [Google Scholar] [CrossRef] [PubMed]
- Gastinger, M.J.; Barber, A.J.; Vardi, N.; Marshak, D.W. Histamine receptors in mammalian retinas. J. Comp. Neurol. 2006, 495, 658–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masini, E.; Mannaioni, P.F.; Pistelli, A.; Salvemini, D.; Vane, J. Impairment of the L-arginine-nitric oxide pathway in mast cells from spontaneously hypertensive rats. Biochem. Biophys. Res. Commun. 1991, 177, 1178–1182. [Google Scholar] [CrossRef]
- Cacabelos, R. Histaminergic system: Neuroendocrine function of brain histamine. Methods Find. Exp. Clin. Pharmacol. 1990, 12, 341–376. [Google Scholar] [CrossRef] [PubMed]
- Cacabelos, R.; Torrellas, C.; Fernández-Novoa, L.; López-Muñoz, F. Histamine and Immune Biomarkers in CNS Disorders. Mediators Inflamm. 2016, 2016, 1924603. [Google Scholar] [CrossRef] [PubMed]
- Markwardt, K.L.; Magnino, P.E.; Pang, I.H. Effect of histamine on phosphoinositide turnover and intracellular calcium in human ciliary muscle cells. Exp. Eye Res. 1996, 62, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Bouhenni, R.A.; Dunmire, J.; Sewell, A.; Edward, D.P. Animal Models of Glaucoma. J. Biomed. Biotechnol 2012, 2012, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Chen, Z.; Song, J. A study of experimental carbomer glaucoma and other experimental glaucoma in rabbits. Zhonghua. Yan Ke Za Zhi. 2002, 38, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Fabrizi, F.; Mincione, F.; Somma, T.; Scozzafava, G.; Galassi, F.; Masini, E.; Impagnatiello, F.; Supuran, C.T. A new approach to antiglaucoma drugs: Carbonic anhydrase inhibitors with or without NO donating moieties. Mechanism of action and preliminary pharmacology. J. Enzyme Inhib. Med. Chem. 2012, 27, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Yu, L.; Zhuang, Q.X.; Peng, S.Y.; Zhu, J.N.; Wang, J.J. Postsynaptic mechanisms underlying the excitatory action of histamine on medial vestibular nucleus neurons in rats. Br. J. Pharmacol. 2013, 170, 156–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morini, G.; Becchi, G.; Shenton, F.C.; Chazot, P.L.; Grandi, D. Histamine H3 and H4 receptors are expressed on distinct endocrine cell types in the rat fundic mucosa. Inflamm. Res. 2008, 57 (Suppl. 1), S57–S58. [Google Scholar] [CrossRef] [PubMed]
- Veglia, E.; Grange, C.; Pini, A.; Moggio, A.; Lanzi, C.; Camussi, G.; Chazot, P.L.; Rosa, A.C. Histamine receptor expression in human renal tubules: A comparative pharmacological evaluation. Inflamm. Res. 2015, 64. [Google Scholar] [CrossRef] [PubMed]
- Sorkhabi, R.; Ghorbanihaghjo, A.; Javadzadeh, A.; Rashtchizadeh, N.; Moharrery, M. Oxidative DNA damage and total antioxidant status in glaucoma patients. Mol. Vis. 2011, 17, 41–46. [Google Scholar] [PubMed]
- Grant, W.M. Further studies on facility of flow through the trabecular meshwork. AMA. Arch. Ophthalmol. 1958, 60, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Flammer, J.; Orgül, S.; Costa, V.P.; Orzalesi, N.; Krieglstein, G.K.; Serra, L.M.; Renard, J.P.; Stefánsson, E. The impact of ocular blood flow in glaucoma. Prog. Retin. Eye Res. 2002, 21, 359–393. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orihashi, M.; Shima, Y.; Tsuneki, H.; Kimura, I. Potent reduction of intraocular pressure by nipradilol plus latanoprost in ocular hypertensive rabbits. Biol. Pharm. Bull. 2005, 28, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Galassi, F.; Masini, E.; Giambene, B.; Fabrizi, F.; Uliva, C.; Bolla, M.; Ongini, E. A topical nitric oxide-releasing dexamethasone derivative: Effects on intraocular pressure and ocular haemodynamics in a rabbit glaucoma model. Br. J. Ophthalmol. 2006, 90, 1414–1419. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Sinha, G.; Sharma, R.; Nayak, B.; Patil, B.; Kashyap, B.; Shameer, A.; Dada, T. Agreement between diurnal variations of intraocular pressure by Tono-Pen and Goldmann applanation tonometer in patients on topical anti-glaucoma medication. Int. Ophthalmol. 2016, 36, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Goldmann, H.; Schmidt, T. Applanation tonometry. Ophthalmologica 1957, 134, 221–242. [Google Scholar] [CrossRef] [PubMed]
- Szafarz, M.; Kryczyk, A.; Łażewska, D.; Kieć-Kononowicz, K.; Wyska, E. Pharmacokinetics and tissue distribution of the new non-imidazole histamine H3 receptor antagonist 1-[3-(4-tert-butylphenoxy) propyl]piperidine in rats. Xenobiotica 2015, 45, 912–920. [Google Scholar] [CrossRef] [PubMed]
- Łażewska, D.; Kaleta, M.; Schwed, J.S.; Karcz, T.; Mogilski, S.; Latacz, G.; Olejarz, A.; Siwek, A.; Kubacka, M.; Lubelska, A.; et al. Biphenyloxy-alkyl-piperidine and azepane derivatives as histamine H3 receptor ligands. Bioorg. Med. Chem. 2017, 25, 5341–5354. [Google Scholar] [CrossRef] [PubMed]
- Łażewska, D.; Ligneau, X.; Schwartz, J.-C.; Schunack, W.; Stark, H.; Kieć-Kononowicz, K. Ether derivatives of 3-piperidinopropan-1-ol as non-imidazole histamine H3 receptor antagonists. Bioorg. Med. Chem. 2006, 14, 3522–3529. [Google Scholar] [CrossRef] [PubMed]
- Szafarz, M.; Szymura-Oleksiak, J.; Lazewska, D.; Kiec-Kononowicz, K. LC-MS-MS Method for the Analysis of New Non-Imidazole Histamine H(3) Receptor Antagonist 1-[3-(4-tert-Butylphenoxy)propyl]piperidine in Rat Serum-Application to Pharmacokinetic Studies. Chromatographia 2011, 73, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Panula, P.; Chazot, P.L.; Cowart, M.; Gutzmer, R.; Leurs, R.; Liu, W.L.S.; Stark, H.; Thurmond, R.L.; Haas, H.L. International Union of Basic and Clinical Pharmacology. XCVIII. Histamine Receptors. Pharmacol. Rev. 2015, 67, 601–655. [Google Scholar] [CrossRef] [PubMed]
- Wilhelmus, K.R. The Draize eye test. Surv. Ophthalmol. 2001, 45, 493–515. [Google Scholar] [CrossRef]
- Lodovici, M.; Casalini, C.; Cariaggi, R.; Michelucci, L.; Dolara, P. Levels of 8-hydroxydeoxyguanosine as a marker of DNA damage in human leukocytes. Free Radic. Biol. Med. 2000, 28, 13–17. [Google Scholar] [CrossRef]






| Name | Primer Sequence (5′-3′) | TM (°C) | Name | Primer Sequence (5′-3′) | TM (°C) | Product Length (bp) |
|---|---|---|---|---|---|---|
| r H1R F a | AGCATGGAACGTCCAGTAGT | 58.73 | r H1R R | TTGCCCTCACACATCCTGTC | 59.96 | 109 |
| r H2R F b | GGATACCGCGACTACGAACC | 60.32 | r H2R R | ATCGTGGGAAAGCTGACACG | 60.67 | 94 |
| r H3R F c | TCACTGGAGAAGCGCATGAA | 59.68 | r H3R R | GAGCCCAAAGATGCTCACGA | 60.39 | 114 |
| r H4R F d | GTAGGAAACGCGGTGGTCAT | 60.39 | r H4R R | TGAGCCAAAACAGGCAGACT | 59.82 | 184 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lanzi, C.; Lucarini, L.; Durante, M.; Sgambellone, S.; Pini, A.; Catarinicchia, S.; Łażewska, D.; Kieć-Kononowicz, K.; Stark, H.; Masini, E. Role of Histamine H3 Receptor Antagonists on Intraocular Pressure Reduction in Rabbit Models of Transient Ocular Hypertension and Glaucoma. Int. J. Mol. Sci. 2019, 20, 981. https://doi.org/10.3390/ijms20040981
Lanzi C, Lucarini L, Durante M, Sgambellone S, Pini A, Catarinicchia S, Łażewska D, Kieć-Kononowicz K, Stark H, Masini E. Role of Histamine H3 Receptor Antagonists on Intraocular Pressure Reduction in Rabbit Models of Transient Ocular Hypertension and Glaucoma. International Journal of Molecular Sciences. 2019; 20(4):981. https://doi.org/10.3390/ijms20040981
Chicago/Turabian StyleLanzi, Cecilia, Laura Lucarini, Mariaconcetta Durante, Silvia Sgambellone, Alessandro Pini, Stefano Catarinicchia, Dorota Łażewska, Katarzyna Kieć-Kononowicz, Holger Stark, and Emanuela Masini. 2019. "Role of Histamine H3 Receptor Antagonists on Intraocular Pressure Reduction in Rabbit Models of Transient Ocular Hypertension and Glaucoma" International Journal of Molecular Sciences 20, no. 4: 981. https://doi.org/10.3390/ijms20040981
APA StyleLanzi, C., Lucarini, L., Durante, M., Sgambellone, S., Pini, A., Catarinicchia, S., Łażewska, D., Kieć-Kononowicz, K., Stark, H., & Masini, E. (2019). Role of Histamine H3 Receptor Antagonists on Intraocular Pressure Reduction in Rabbit Models of Transient Ocular Hypertension and Glaucoma. International Journal of Molecular Sciences, 20(4), 981. https://doi.org/10.3390/ijms20040981