Photobiomodulation Enhances the Angiogenic Effect of Mesenchymal Stem Cells to Mitigate Radiation-Induced Enteropathy

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
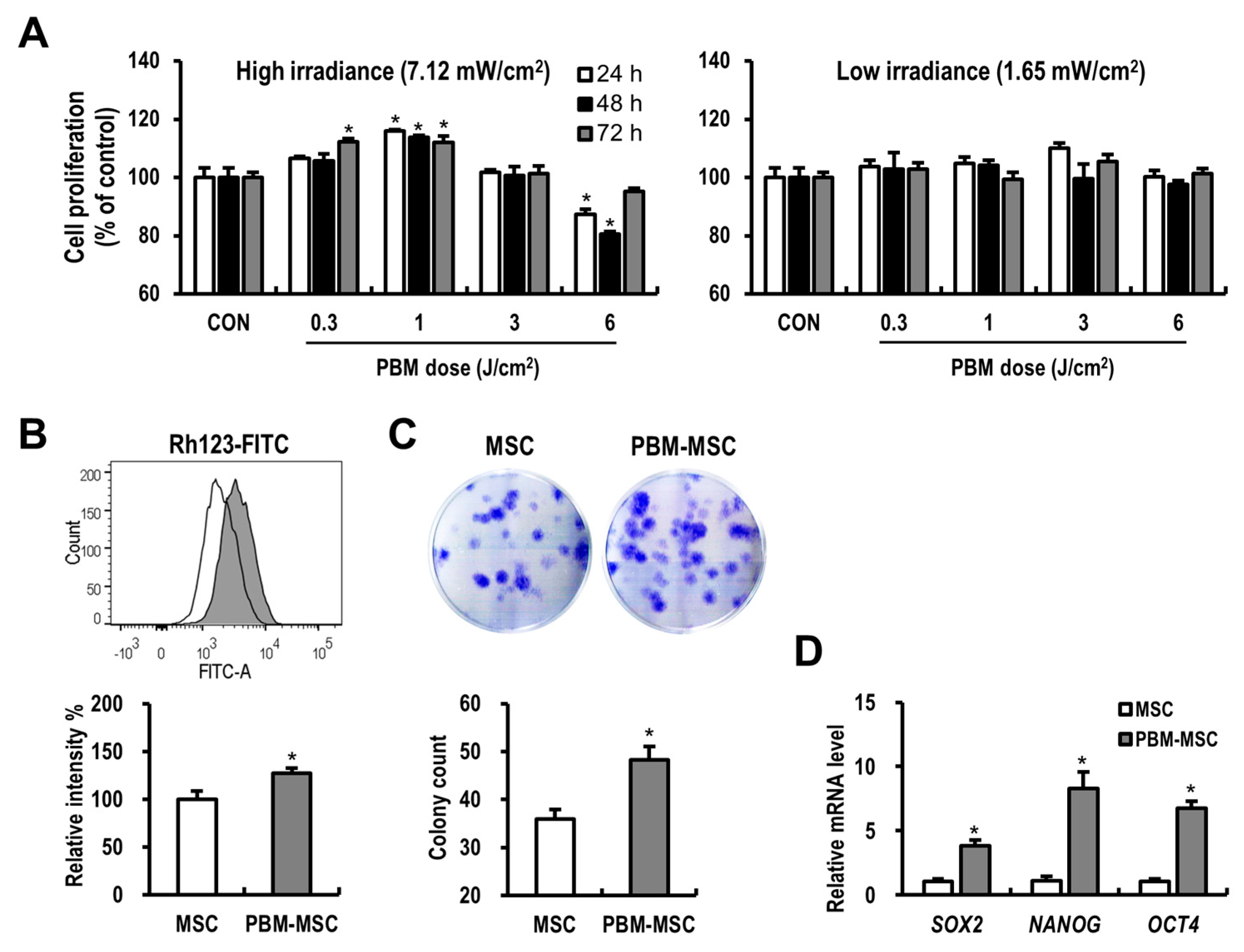
2.1. PBM Enhances the Proliferation and Self-Renewal of MSCs
2.2. PBM Maintains the Immunophenotype and Differentiation Potential of MSCs
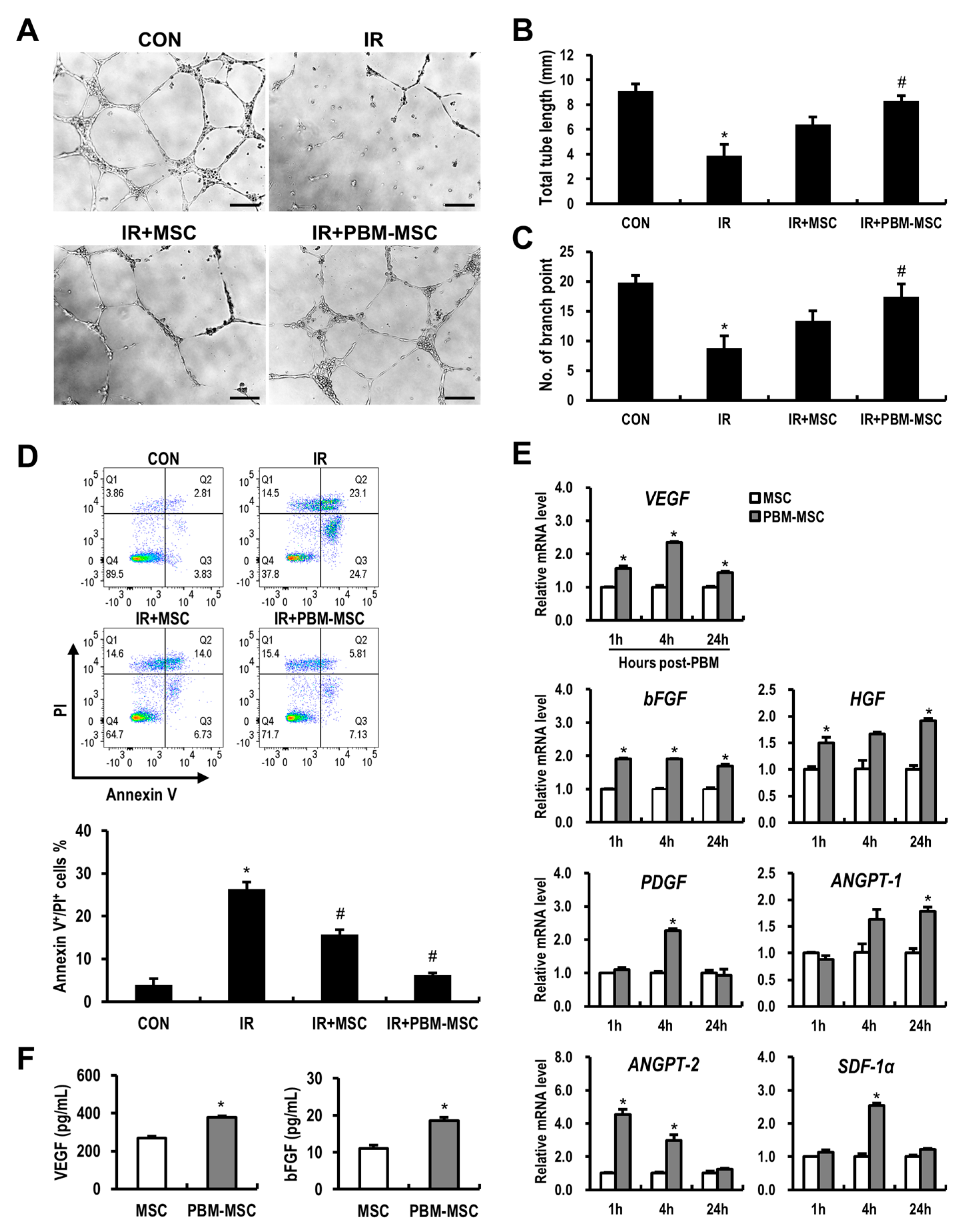
2.3. PBM Promotes the Angiogenic Capacity of MSCs to Attenuate Radiation-Induced Damage to Vascular Endothelial Cells
2.4. PBM Preconditioning Enhances the Therapeutic Efficacy of MSCs against Radiation-Induced Enteropathy
2.5. PBM-Preconditioned MSCs Attenuate Intestinal Barrier Damage and Inflammation During Radiation-Induced Enteropathy
2.6. PBM-Preconditioned MSCs Rescue Microvasculature Damage in the Irradiated Intestine
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. PBM Preconditioning
4.3. Cell Proliferation
4.4. Mitochondrial Membrane Potential
4.5. Colony-Forming Unit-Fibroblast (CFU-f) Clonogenic Assay
4.6. Immunophenotype of MSCs
4.7. Adipogenic and Osteogenic Differentiation of MSCs
4.8. RNA Extraction and Real-Time RT-PCR
4.9. Preparation of MSC-Conditioned Medium
4.10. HUVEC Apoptosis and Capillary-Like Tube Formation Assays
4.11. ELISA
4.12. Animals
4.13. Irradiation and MSC Administration
4.14. Histological Analysis
4.15. Bacterial Translocation
4.16. Flow Cytometric Analysis of Isolated Intestinal Stromal Cells
4.17. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ANGPT | angiopoietin |
| ANOVA | analysis of variance |
| APC | allophycocyanin |
| bFGF | basic fibroblast growth factor |
| BGLAP | bone gamma-carboxyglutamate protein |
| CD | cluster of differentiation |
| CFU-f | colony-forming unit-fibroblast |
| Cldn3 | claudin 3 |
| DMEM | Dulbecco’s modified Eagle medium |
| EBM-2 | endothelial cell basal medium-2 |
| EDTA | ethylenediaminetetraacetic acid |
| EGM-2 | endothelial cell growth medium-2 |
| FBS | fetal bovine serum |
| FITC | fluorescein isothiocyanate |
| HGF | hepatocyte growth factor |
| HLA-DR | human leukocyte antigen-DR isotype |
| HUVEC | human umbilical vein endothelial cell |
| H&E | hematoxylin and eosin |
| Il | interleukin |
| ISCT | International Society of Cellular therapy |
| KIRAMS | Korea Institute of Radiological and Medical Sciences |
| LPL | lipoprotein lipase |
| MEM-α | minimum essential medium-alpha |
| Mmp | matrix metallopeptidase |
| Mpo | myeloperoxidase |
| MSC | mesenchymal stem cell |
| MSC-CM | MSC-conditioned medium |
| NANOG | nanog homeobox |
| OCT4 | octamer-binding transcription factor 4 |
| PBM | photobiomodulation |
| PBS | phosphate-buffered saline |
| PDGF | platelet-derived growth factor |
| PE | phycoerythrin |
| PE-Cy7 | phycoerythrin-cyanine 7 |
| PI | propidium iodide |
| PPARγ | peroxisome proliferator-activated receptor gamma |
| Rh123 | rhodamine 123 |
| RT-PCR | reverse transcription-polymerase chain reaction |
| SDF-1α | stromal cell-derived factor 1 alpha |
| SEM | standard error of the mean |
| SOX2 | sex determining region Y-box 2 |
| SPF | specific pathogen-free |
| TJ | tight junction |
| VEGF | vascular endothelial growth factor |
References
- Mettler, F.A., Jr.; Voelz, G.L. Major radiation exposure—what to expect and how to respond. N. Engl. J. Med. 2002, 346, 1554–1561. [Google Scholar] [CrossRef] [PubMed]
- Hauer-Jensen, M.; Denham, J.W.; Andreyev, H.J. Radiation enteropathy—pathogenesis, treatment and prevention. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 470–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moussa, L.; Usunier, B.; Demarquay, C.; Benderitter, M.; Tamarat, R.; Semont, A.; Mathieu, N. Bowel Radiation Injury: Complexity of the Pathophysiology and Promises of Cell and Tissue Engineering. Cell Transplant. 2016, 25, 1723–1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalla, R.V.; Bowen, J.; Barasch, A.; Elting, L.; Epstein, J.; Keefe, D.M.; McGuire, D.B.; Migliorati, C.; Nicolatou-Galitis, O.; Peterson, D.E.; et al. MASCC/ISOO clinical practice guidelines for the management of mucositis secondary to cancer therapy. Cancer 2014, 120, 1453–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atwood, K.C.; Norman, A. On the Interpretation of Multi-Hit Survival Curves. Proc. Natl. Acad. Sci. USA 1949, 35, 696–709. [Google Scholar] [CrossRef] [PubMed]
- Paris, F.; Fuks, Z.; Kang, A.; Capodieci, P.; Juan, G.; Ehleiter, D.; Haimovitz-Friedman, A.; Cordon-Cardo, C.; Kolesnick, R. Endothelial apoptosis as the primary lesion initiating intestinal radiation damage in mice. Science 2001, 293, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Maj, J.G.; Paris, F.; Haimovitz-Friedman, A.; Venkatraman, E.; Kolesnick, R.; Fuks, Z. Microvascular function regulates intestinal crypt response to radiation. Cancer Res. 2003, 63, 4338–4341. [Google Scholar] [PubMed]
- Okunieff, P.; Mester, M.; Wang, J.; Maddox, T.; Gong, X.; Tang, D.; Coffee, M.; Ding, I. In vivo radioprotective effects of angiogenic growth factors on the small bowel of C3H mice. Radiat. Res. 1998, 150, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.H.; Kammerer, R.A.; Lee, H.J.; Yasunaga, K.; Kim, K.T.; Choi, H.H.; Kim, W.; Kim, S.H.; Park, S.K.; Lee, G.M.; et al. Designed angiopoietin-1 variant, COMP-Ang1, protects against radiation-induced endothelial cell apoptosis. Proc. Natl. Acad. Sci. USA 2004, 101, 5553–5558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotolo, J.; Stancevic, B.; Zhang, J.; Hua, G.; Fuller, J.; Yin, X.; Haimovitz-Friedman, A.; Kim, K.; Qian, M.; Cardo-Vila, M.; et al. Anti-ceramide antibody prevents the radiation gastrointestinal syndrome in mice. J. Clin. Investig. 2012, 122, 1786–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rannou, E.; Francois, A.; Toullec, A.; Guipaud, O.; Buard, V.; Tarlet, G.; Mintet, E.; Jaillet, C.; Iruela-Arispe, M.L.; Benderitter, M.; et al. In vivo evidence for an endothelium-dependent mechanism in radiation-induced normal tissue injury. Sci. Rep. 2015, 5, 15738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinnaird, T.; Stabile, E.; Burnett, M.S.; Shou, M.; Lee, C.W.; Barr, S.; Fuchs, S.; Epstein, S.E. Local delivery of marrow-derived stromal cells augments collateral perfusion through paracrine mechanisms. Circulation 2004, 109, 1543–1549. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tredget, E.E.; Wu, P.Y.; Wu, Y. Paracrine factors of mesenchymal stem cells recruit macrophages and endothelial lineage cells and enhance wound healing. PLoS ONE 2008, 3, e1886. [Google Scholar] [CrossRef] [PubMed]
- Timmers, L.; Lim, S.K.; Hoefer, I.E.; Arslan, F.; Lai, R.C.; van Oorschot, A.A.; Goumans, M.J.; Strijder, C.; Sze, S.K.; Choo, A.; et al. Human mesenchymal stem cell-conditioned medium improves cardiac function following myocardial infarction. Stem Cell Res. 2011, 6, 206–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linero, I.; Chaparro, O. Paracrine effect of mesenchymal stem cells derived from human adipose tissue in bone regeneration. PLoS ONE 2014, 9, e107001. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.D.; Sjoquist, P.O. Myocardial regeneration with stem cells: Pharmacological possibilities for efficacy enhancement. Pharmacol. Res. 2006, 53, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Baldari, S.; Di Rocco, G.; Piccoli, M.; Pozzobon, M.; Muraca, M.; Toietta, G. Challenges and Strategies for Improving the Regenerative Effects of Mesenchymal Stromal Cell-Based Therapies. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Anders, J.J.; Lanzafame, R.J.; Arany, P.R. Low-level light/laser therapy versus photobiomodulation therapy. Photomed. Laser Surg. 2015, 33, 183–184. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, A.P.; Paraguassu, G.M.; Silveira, N.T.; de Souza, J.; Cangussu, M.C.; dos Santos, J.N.; Pinheiro, A.L. Laser and LED phototherapies on angiogenesis. Lasers Med. Sci. 2013, 28, 981–987. [Google Scholar] [CrossRef] [PubMed]
- De Freitas, L.F.; Hamblin, M.R. Proposed Mechanisms of Photobiomodulation or Low-Level Light Therapy. IEEE J. Sel Top. Quantum Electron. 2016, 22. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.F.; Zhang, H.; Yuan, X.; Li, J.; Wei, Y.J.; Hu, S.S. In vitro effects of low-level laser irradiation for bone marrow mesenchymal stem cells: Proliferation, growth factors secretion and myogenic differentiation. Lasers Surg. Med. 2008, 40, 726–733. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, T.S.; Serra, A.J.; Manchini, M.T.; Bassaneze, V.; Krieger, J.E.; de Tarso Camillo de Carvalho, P.; Antunes, D.E.; Bocalini, D.S.; Ferreira Tucci, P.J.; Silva, J.A., Jr. Effects of low level laser therapy on attachment, proliferation, and gene expression of VEGF and VEGF receptor 2 of adipocyte-derived mesenchymal stem cells cultivated under nutritional deficiency. Lasers Med. Sci. 2015, 30, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.B. Mitochondrial membrane potential in living cells. Annu. Rev. Cell Biol. 1988, 4, 155–181. [Google Scholar] [CrossRef] [PubMed]
- Waselenko, J.K.; MacVittie, T.J.; Blakely, W.F.; Pesik, N.; Wiley, A.L.; Dickerson, W.E.; Tsu, H.; Confer, D.L.; Coleman, C.N.; Seed, T.; et al. Medical management of the acute radiation syndrome: Recommendations of the Strategic National Stockpile Radiation Working Group. Ann. Intern. Med. 2004, 140, 1037–1051. [Google Scholar] [CrossRef] [PubMed]
- Giris, M.; Erbil, Y.; Oztezcan, S.; Olgac, V.; Barbaros, U.; Deveci, U.; Kirgiz, B.; Uysal, M.; Toker, G.A. The effect of heme oxygenase-1 induction by glutamine on radiation-induced intestinal damage: The effect of heme oxygenase-1 on radiation enteritis. Am. J. Surg. 2006, 191, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Lee, J.; Park, S.; Myung, H.; Kang, J.; Kim, K.; Kim, H.; Jang, W.S.; Lee, S.J.; Shim, S.; et al. Pravastatin Attenuates Acute Radiation-Induced Enteropathy and Improves Epithelial Cell Function. Front. Pharmacol. 2018, 9, 1215. [Google Scholar] [CrossRef] [PubMed]
- Duff, S.E.; Li, C.; Garland, J.M.; Kumar, S. CD105 is important for angiogenesis: Evidence and potential applications. FASEB J. 2003, 17, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, R.M.; Capoccia, B.J.; Devine, S.M.; Dipersio, J.; Trinkaus, K.M.; Ingram, D.; Link, D.C. Angiogenic cells can be rapidly mobilized and efficiently harvested from the blood following treatment with AMD3100. Blood 2006, 108, 3662–3667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.Y.; Qu, Y.Q.; Wang, J.; Dong, L.H. The potential of mesenchymal stem cells in the management of radiation enteropathy. Cell Death Dis. 2015, 6, e1840. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Min, X.H.; Wang, Q.Y.; Leung, F.W.; Shi, L.; Zhou, Y.; Yu, T.; Wang, C.M.; An, G.; Sha, W.H.; et al. Pre-activation of mesenchymal stem cells with TNF-alpha, IL-1beta and nitric oxide enhances its paracrine effects on radiation-induced intestinal injury. Sci. Rep. 2015, 5, 8718. [Google Scholar] [CrossRef] [PubMed]
- Linard, C.; Strup-Perrot, C.; Lacave-Lapalun, J.V.; Benderitter, M. Flagellin preconditioning enhances the efficacy of mesenchymal stem cells in an irradiation-induced proctitis model. J. Leukoc. Biol. 2016, 100, 569–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moussa, L.; Pattappa, G.; Doix, B.; Benselama, S.L.; Demarquay, C.; Benderitter, M.; Semont, A.; Tamarat, R.; Guicheux, J.; Weiss, P.; et al. A biomaterial-assisted mesenchymal stromal cell therapy alleviates colonic radiation-induced damage. Biomaterials 2017, 115, 40–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Gong, J.F.; Zhang, W.; Zhu, W.M.; Li, J.S. Effects of transplanted bone marrow mesenchymal stem cells on the irradiated intestine of mice. J. Biomed. Sci. 2008, 15, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Baxter, G.D.; Liu, L.; Petrich, S.; Gisselman, A.S.; Chapple, C.; Anders, J.J.; Tumilty, S. Low level laser therapy (Photobiomodulation therapy) for breast cancer-related lymphedema: A systematic review. BMC Cancer 2017, 17, 833. [Google Scholar] [CrossRef] [PubMed]
- Chow, R.T.; Johnson, M.I.; Lopes-Martins, R.A.; Bjordal, J.M. Efficacy of low-level laser therapy in the management of neck pain: A systematic review and meta-analysis of randomised placebo or active-treatment controlled trials. Lancet 2009, 374, 1897–1908. [Google Scholar] [CrossRef]
- Huang, Z.; Chen, J.; Ma, J.; Shen, B.; Pei, F.; Kraus, V.B. Effectiveness of low-level laser therapy in patients with knee osteoarthritis: A systematic review and meta-analysis. Osteoarthr. Cartil. 2015, 23, 1437–1444. [Google Scholar] [CrossRef] [PubMed]
- Peplow, P.V.; Chung, T.Y.; Baxter, G.D. Laser photobiomodulation of proliferation of cells in culture: A review of human and animal studies. Photomed. Laser Surg. 2010, 28 (Suppl. 1), S3–S40. [Google Scholar] [CrossRef]
- Tucker, L.D.; Lu, Y.; Dong, Y.; Yang, L.; Li, Y.; Zhao, N.; Zhang, Q. Photobiomodulation Therapy Attenuates Hypoxic-Ischemic Injury in a Neonatal Rat Model. J. Mol. Neurosci. MN 2018, 65, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Muili, K.A.; Gopalakrishnan, S.; Meyer, S.L.; Eells, J.T.; Lyons, J.A. Amelioration of experimental autoimmune encephalomyelitis in C57BL/6 mice by photobiomodulation induced by 670 nm light. PLoS ONE 2012, 7, e30655. [Google Scholar] [CrossRef] [PubMed]
- Mvula, B.; Mathope, T.; Moore, T.; Abrahamse, H. The effect of low level laser irradiation on adult human adipose derived stem cells. Lasers Med. Sci. 2008, 23, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Li, W.T.; Leu, Y.C.; Wu, J.L. Red-light light-emitting diode irradiation increases the proliferation and osteogenic differentiation of rat bone marrow mesenchymal stem cells. Photomed. Laser Surg. 2010, 28 (Suppl. 1), S157–S165. [Google Scholar] [CrossRef]
- Tuby, H.; Maltz, L.; Oron, U. Induction of autologous mesenchymal stem cells in the bone marrow by low-level laser therapy has profound beneficial effects on the infarcted rat heart. Lasers Surg. Med. 2011, 43, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Zhu, R.; Wang, S.; Zhao, R.C. Low-Level Laser Effect on Proliferation, Migration, and Antiapoptosis of Mesenchymal Stem Cells. Stem Cells Dev. 2017, 26, 762–775. [Google Scholar] [CrossRef] [PubMed]
- Du, W.J.; Chi, Y.; Yang, Z.X.; Li, Z.J.; Cui, J.J.; Song, B.Q.; Li, X.; Yang, S.G.; Han, Z.B.; Han, Z.C. Heterogeneity of proangiogenic features in mesenchymal stem cells derived from bone marrow, adipose tissue, umbilical cord, and placenta. Stem Cell Res. Ther. 2016, 7, 163. [Google Scholar] [CrossRef] [PubMed]
- Nagata, M.; Iwasaki, K.; Akazawa, K.; Komaki, M.; Yokoyama, N.; Izumi, Y.; Morita, I. Conditioned Medium from Periodontal Ligament Stem Cells Enhances Periodontal Regeneration. Tissue Eng. Part A 2017, 23, 367–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, F.G.; Carvalho, M.M.; Panchalingam, K.M.; Rodrigues, A.J.; Mendes-Pinheiro, B.; Anjo, S.; Manadas, B.; Behie, L.A.; Sousa, N.; Salgado, A.J. Impact of the Secretome of Human Mesenchymal Stem Cells on Brain Structure and Animal Behavior in a Rat Model of Parkinson’s Disease. Stem Cells Transl. Med. 2017, 6, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Shi, M.; Li, L.; Liu, J.; Chen, B.; Chen, Y.; An, X.; Liu, S.; Luo, R.; Long, D.; et al. Mesenchymal stem cell-conditioned media ameliorate diabetic endothelial dysfunction by improving mitochondrial bioenergetics via the Sirt1/AMPK/PGC-1alpha pathway. Clin. Sci. 2016, 130, 2181–2198. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Pham, P.V.; Vu, N.B.; Pham, V.M.; Truong, N.H.; Pham, T.L.; Dang, L.T.; Nguyen, T.T.; Bui, A.N.; Phan, N.K. Good manufacturing practice-compliant isolation and culture of human umbilical cord blood-derived mesenchymal stem cells. J. Transl. Med. 2014, 12, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stzepourginski, I.; Eberl, G.; Peduto, L. An optimized protocol for isolating lymphoid stromal cells from the intestinal lamina propria. J. Immunol. Methods 2015, 421, 14–19. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wavelength | Irradiance (mW/cm2) | Duration (sec) | Fluence (J/cm2) |
|---|---|---|---|
| 633 nm | 1.65 | 182 606 1818 3636 | 0.3 1 3 6 |
| 7.12 | 42 140 421 843 | 0.3 1 3 6 |
| Species | Primer | Forward (5′–3′) | Reverse (5′–3′) | bp |
|---|---|---|---|---|
| Human | SOX2 | GCCCTGCAGTACAACTCCAT | GACTTGACCACCGAACCCAT | 128 |
| NANOG | AAGGCCTCAGCACCTACCTA | TGCACCAGGTCTGAGTGTTC | 181 | |
| OCT4 | GATGTGGTCCGAGTGTGGTT | AGCCTGGGGTACCAAAATGG | 160 | |
| PPARγ | CTAAAGAGCCTGCGAAAG | TGTCTGTCTCCGTCTTCTTG | 331 | |
| LPL | TCAACTGGATGGAGGAG | GGGGCTTCTGCATACTCAAA | 169 | |
| ALP | CAACAGGGTAGATTTCTCTTGG | GGTCAGATCCAGAATGTTCC | 135 | |
| BGLAP | GGCAGCGAGGTAGTGAAGAG | CAGCAGAGCGACACCCTAGAC | 195 | |
| VEGF | CCCACTGAGGAGTCCAACAT | TTTCTTGCGCTTTCGTTTTT | 186 | |
| bFGF | CGACCCTCACATCAAGCTACA | CGTTTCAGTGCCACATACCAA | 219 | |
| HGF | ATCAAATGTCAGCCCTGGAG | TCGATAACTCTCCCCATTGC | 207 | |
| ANGPT-1 | CATTCTTCGCTGCCATTCTG | GCACATTGCCCATGTTGAATC | 103 | |
| ANGPT-2 | ACTGTGTCCTCTTCCACCAC | GGATGTTTAGGGTCTTGCTTT | 132 | |
| PDGF | GCAAGACCAGGACGGTCATTT | GGCACTTGACACTGCTCGT | 135 | |
| SDF-1α | TCAGCCTGAGCTACAGATGC | CTTTAGCTTCGGGTCAATGC | 161 | |
| GAPDH | GGACTCATGACCACAGTCCATGCC | TCAGGGATGACCTTGCCCACAG | 152 | |
| Mouse | Il-1β | GGTCAAAGGTTTGGAAGCAG | TGTGAAATGCCACCTTTTGA | 94 |
| Mmp9 | GCCCTGGAACTCACACGACA | TTGGAAACTCACACGCCAGAAG | 85 | |
| Cldn3 | AAGCCGAATGGACAAAGAA | CTGGCAAGTAGCTGCAGTG | 72 | |
| β-actin | TCCCTGGAGAAGAGCTATGA | CGATAAAGGAAGGCTGGAA | 100 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.; Lee, J.; Jang, H.; Park, S.; Na, J.; Myung, J.K.; Kim, M.-J.; Jang, W.-S.; Lee, S.-J.; Kim, H.; et al. Photobiomodulation Enhances the Angiogenic Effect of Mesenchymal Stem Cells to Mitigate Radiation-Induced Enteropathy. Int. J. Mol. Sci. 2019, 20, 1131. https://doi.org/10.3390/ijms20051131
Kim K, Lee J, Jang H, Park S, Na J, Myung JK, Kim M-J, Jang W-S, Lee S-J, Kim H, et al. Photobiomodulation Enhances the Angiogenic Effect of Mesenchymal Stem Cells to Mitigate Radiation-Induced Enteropathy. International Journal of Molecular Sciences. 2019; 20(5):1131. https://doi.org/10.3390/ijms20051131
Chicago/Turabian StyleKim, Kyuchang, Janet Lee, Hyosun Jang, Sunhoo Park, Jiyoung Na, Jae Kyung Myung, Min-Jung Kim, Won-Suk Jang, Sun-Joo Lee, Hyewon Kim, and et al. 2019. "Photobiomodulation Enhances the Angiogenic Effect of Mesenchymal Stem Cells to Mitigate Radiation-Induced Enteropathy" International Journal of Molecular Sciences 20, no. 5: 1131. https://doi.org/10.3390/ijms20051131
APA StyleKim, K., Lee, J., Jang, H., Park, S., Na, J., Myung, J. K., Kim, M. -J., Jang, W. -S., Lee, S. -J., Kim, H., Myung, H., Kang, J., & Shim, S. (2019). Photobiomodulation Enhances the Angiogenic Effect of Mesenchymal Stem Cells to Mitigate Radiation-Induced Enteropathy. International Journal of Molecular Sciences, 20(5), 1131. https://doi.org/10.3390/ijms20051131