Human Heart Explant-Derived Extracellular Vesicles: Characterization and Effects on the In Vitro Recellularization of Decellularized Heart Valves

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Explant Characterization: Cells Derived from Different Heart Regions Are Similar and Heterogeneous
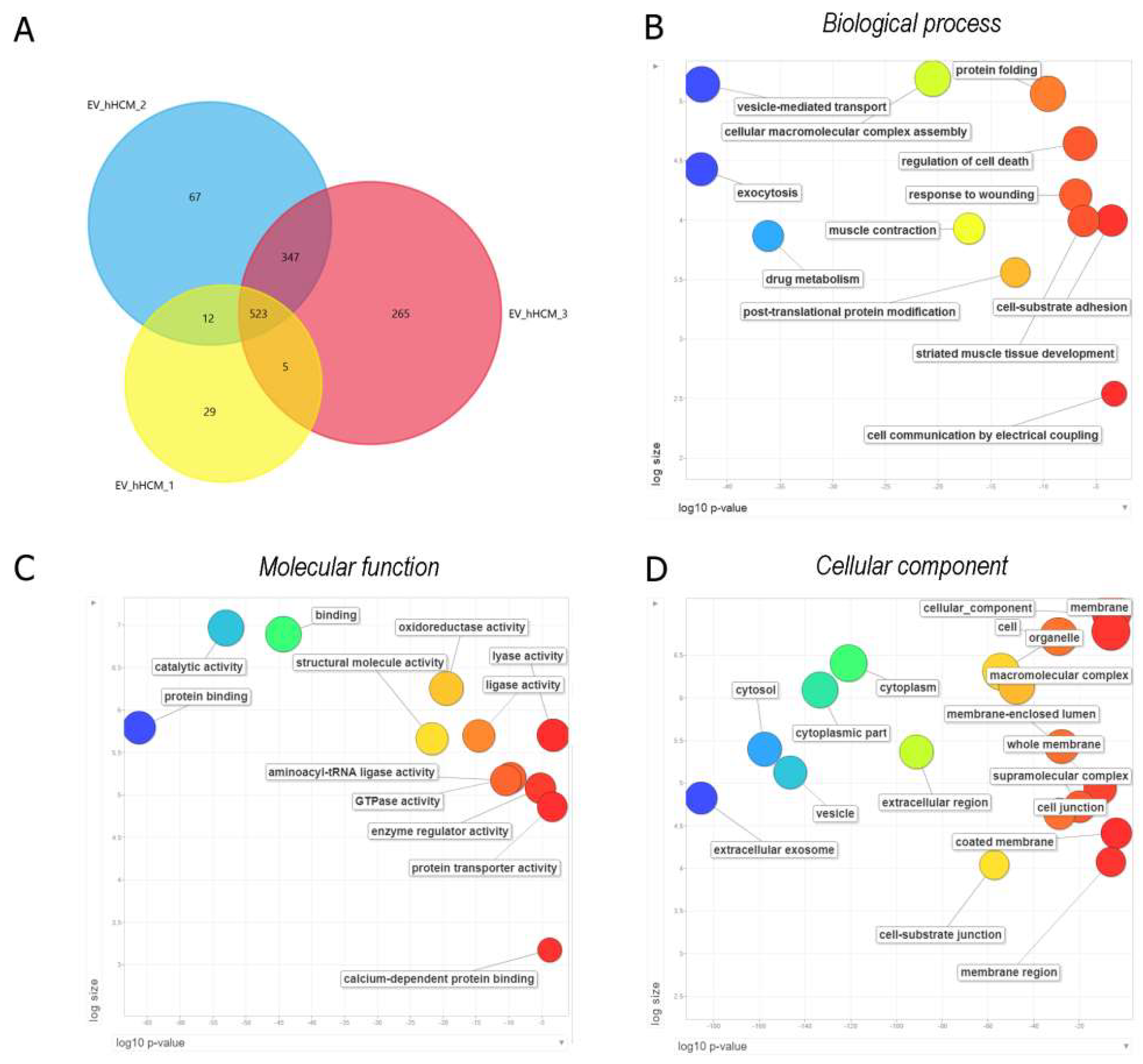
2.2. Characterization and Prospective Functional Analysis of Human Heart-Derived Extracellular Vesicles (hH-EVs)
2.3. hH-EVs Stimulate Directional Cell migration and Decrease the Adhesion Capabilities of Adipose-Derived Stem Cells (ADSCs)
2.4. hH-EVs Stimulate Proliferation and the in Vitro Angiogenesis of Human Umbilical Vein Endothelial Cells (HUVECs)
2.5. Effect of Left Ventricular Endocardium Extracellular Vesicles (LVE-EVs) on Leaflet Scaffold Recellularization
3. Discussion
4. Materials and Methods
4.1. Ethics Statements
4.2. The Culture and Characterization of the Explants
4.3. Isolation of Heart-Derived Extracellular Vesicles
4.4. hH-EV Characterization by Nano-Tracking Analysis, Transmission Electron Microscopy (TEM) and Mass Spectrometry (MS) Analysis
4.5. ADSC and HUVEC Cultures
4.6. Cell Proliferation Assay
4.7. Cell Adhesion Assay
4.8. Scratch Wound Assay
4.9. In Vitro Angiogenesis Assay
4.10. Decellularization of Porcine Heart Valves
4.11. In Vitro Porcine Valve Leaflet Scaffold Recellularization
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EVs | Extracellular vesicles |
| hH-EVs | Human heart-derived extracellular vesicles |
| NTA | Nanoparticle Tracking Analysis |
| TEM | Transmission Electron Microscopy |
| MS | Mass spectrometer |
| GO | Gene ontology |
| CRCS | Cardiac resident stromal cell |
| miRNA | microRNA |
| DHV | Decellularized heart valves |
| LVE | Left ventricular endocardium |
| LVM | Left ventricular myocardium |
| RVE | Right ventricular endocardium |
| RVM | Right ventricular myocardium |
| AUM | Right auricle myocardium |
| AUE | Right auricle endocardium |
| MTL | Mitral valve leaflet |
| FDR | False discovery Rate |
| ADSC | Human adipose-derived stem cells |
| HUVEC | Human umbilical vein endothelial cells |
| EdU | 5-ethynyl-2′-deoxyuridine |
| ITGA1 | Integrin, alpha I |
| ILK | Integrin-linked kinase |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| EBM-2 | Endothelial Cell Growth Medium |
| iPS | Induced pluripotent stem cells |
| LVE-EV | Left ventricular endocardium-derived extracellular vesicles |
| ITGA1 | Integrin, alpha 1 |
| ITGA5 | Integrin, alpha V |
| ITGB1 | Integrin, beta I |
| VTN | Vitronectin |
| CTNNB1 | Catenin (cadherin-associated protein), beta 1 |
| CDH13 | Cadherin 13 |
| CDCs | Cardiosphere-derived cells |
| HSP-90 | Heat shock protein 90 kDa alpha |
References
- Gerbin, K.A.; Murry, C.E. The winding road to regenerating the human heart. Cardiovasc. Pathol. 2015, 24, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, A.R.; Ilinykh, A.; Ivey, M.J.; Kuwabara, J.T.; D’antoni, M.L.; Debuque, R.; Chandran, A.; Wang, L.; Arora, K.; Rosenthal, N.A.; et al. Revisiting cardiac cellular composition. Circ. Res. 2016, 118, 400–409. [Google Scholar] [CrossRef]
- De Aguiar, A.M.; Kuligovski, C.; da Costa, M.T.B.A.; Stimamiglio, M.A.; Rebelatto, C.L.K.; Senegaglia, A.C.; Brofman, P.R.S.; Dallagiovanna, B.; Goldenberg, S.; Correa, A. Alkaline phosphatase-positive cells isolated from human hearts have mesenchymal stem cell characteristics. Stem Cell Discov. 2011, 01, 71–80. [Google Scholar] [CrossRef]
- Reus, T.L.; Robert, A.W.; da Costa, M.B.A.; de Aguiar, A.M.; Stimamiglio, M.A. Secretome from resident cardiac stromal cells stimulates proliferation, cardiomyogenesis and angiogenesis of progenitor cells. Int. J. Cardiol. 2016, 221, 396–403. [Google Scholar] [CrossRef]
- Jackson, R.; Mount, S.; Ye, B.; Mayfield, A.E.; Chan, V.; Boodhwani, M.; Davies, R.A.; Haddad, H.; Davis, D.R. Isolation of human explant derived cardiac stem cells from cryopreserved heart tissue. PLoS ONE 2017, 12, e0176000. [Google Scholar] [CrossRef] [PubMed]
- Marbán, E.; Cingolani, E. Heart to heart: Cardiospheres for myocardial regeneration. Heart Rhythm 2012, 9, 1727–1731. [Google Scholar]
- Smith, R.R.; Barile, L.; Cho, C.H.; Leppo, M.K.; Hare, J.M.; Messina, E.; Giacomello, A.; Abraham, R.; Marbán, E. Regenerative Potential of Cardiosphere-Derived Cells Expanded From Percutaneous Endomyocardial Biopsy Specimens. Circulation 2007, 115, 896–908. [Google Scholar] [Green Version]
- Urbanelli, L.; Magini, A.; Buratta, S.; Brozzi, A.; Sagini, K.; Polchi, A.; Tancini, B.; Emiliani, C. Signaling pathways in exosomes biogenesis, secretion and fate. Genes 2013, 4, 152–170. [Google Scholar] [CrossRef] [PubMed]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Chargaff, E.; West, R. The Biological Significance of the thromboplastic protein of blood. J. Biol. Chem. 1946, 166, 189–197. [Google Scholar]
- Dignat-George, F.; Boulanger, C.M. The many faces of endothelial microparticles. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 27–33. [Google Scholar] [CrossRef]
- Hutcheson, J.D.; Aikawa, E. Extracellular vesicles in cardiovascular homeostasis ans disease. Curr. Opin. Cardiol. 2018, 33, 290–297. [Google Scholar]
- Alibhai, F.J.; Tobin, S.W.; Yeganeh, A.; Weisel, R.D.; Li, R.-K. Emerging roles of extracellular vesicles in cardiac repair and rejuvenation. Am. J. Physiol. Hear. Circ. Physiol. 2018, 315, H733–H744. [Google Scholar] [CrossRef]
- El Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular vesicles: Biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef]
- Lv, L.-L.; Wu, W.-J.; Feng, Y.; Li, Z.-L.; Tang, T.-T.; Liu, B.-C. Therapeutic application of extracellular vesicles in kidney disease: Promises and challenges. J. Cell. Mol. Med. 2017, 22, 728–737. [Google Scholar] [CrossRef]
- Xie, H.; Wang, Z.; Zhang, L.; Lei, Q.; Zhao, A.; Wang, H.; Li, Q.; Cao, Y.; Zhang, W.J.; Chen, Z. Extracellular Vesicle-functionalized Decalcified Bone Matrix Scaffolds with Enhanced Pro-angiogenic and Pro-bone Regeneration Activities. Sci. Rep. 2017, 7, 45622. [Google Scholar] [CrossRef] [Green Version]
- Namiri, M.; Ashtiani, M.K.; Mashinchian, O.; Hasani-Sadrabadi, M.M.; Mahmoudi, M.; Aghdami, N.; Baharvand, H. Engineering natural heart valves: Possibilities and challenges. J. Tissue Eng. Regen. Med. 2016, 4, 524–531. [Google Scholar] [CrossRef]
- Della Barbera, M.; Valente, M.; Basso, C.; Thiene, G. Morphologic studies of cell endogenous repopulation in decellularized aortic and pulmonary homografts implanted in sheep. Cardiovasc. Pathol. 2015, 24, 102–109. [Google Scholar] [CrossRef]
- Cheung, D.Y.; Duan, B.; Butcher, J.T. Current progress in tissue engineering of heart valves: Multiscale problems, multiscale solutions. Expert Opin. Biol. Ther. 2015, 15, 1155–1172. [Google Scholar] [CrossRef]
- Weinhausr, A.J.; Roberts, K.P. Anatomy of the Human Heart; Springer: Berlin/Heidelberg, Germany, 2005; Chapter 4. [Google Scholar]
- Bergmann, O.; Zdunek, S.; Felker, A.; Salehpour, M.; Alkass, K.; Bernard, S.; Sjostrom, S.L.; Szewczykowska, M.; Jackowska, T.; Remedios, C.D.; et al. Dynamics of Cell Generation and Turnover in the Human Heart. Cell 2015, 161, 1566–1575. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Pu, W.T. Recounting cardiac cellular composition. Circ. Res. 2017, 118, 368–370. [Google Scholar] [CrossRef]
- Davis, D.R.; Zhang, Y.; Smith, R.R.; Cheng, K.; Terrovitis, J.; Malliaras, K.; Li, T.; White, A.; Makkar, R.; Marba, E. Validation of the Cardiosphere Method to Culture Cardiac Progenitor Cells from Myocardial Tissue. PLoS ONE 2009, 4, e7195. [Google Scholar] [CrossRef]
- Borosch, S.; Dahmen, E.; Beckers, C.; Stoppe, C.; Buhl, E.M.; Denecke, B.; Goetzenich, A.; Kraemer, S. Characterization of extracellular vesicles derived from cardiac cells in an in vitro model of preconditioning. J. Extracell. Vesicles 2017, 6, 1390391. [Google Scholar] [CrossRef]
- Yarana, C.; Carroll, D.; Chen, J.; Chaiswing, L.; Zhao, Y.; Noel, T.; Alstott, M.; Bae, Y.; Dressler, E.V.; Moscow, J.A.; et al. Extracellular Vesicles Released by Cardiomyocytes in a Doxorubicin-Induced Cardiac Injury Mouse Model Contain Protein Biomarkers of Early Cardiac Injury. Clin. Cancer Res. 2018, 24, 1644–1653. [Google Scholar] [CrossRef]
- Barile, L.; Lionetti, V.; Cervio, E.; Matteucci, M.; Gherghiceanu, M.; Popescu, L.M.; Torre, T.; Siclari, F.; Moccetti, T.; Vassalli, G. Extracellular vesicles fromhuman cardiac progenitor cells inhibit cardiomyocyte apoptosis and improve cardiac function aftermyocardial infarction. Cardiovasc. Res. 2014, 103, 530–541. [Google Scholar] [CrossRef]
- Barile, L.; Milano, G.; Vassalli, G. Beneficial effects of exosomes secreted by cardiac-derived progenitor cells and other cell types in myocardial ischemia. Stem Cell Investig. 2017, 4, 93. [Google Scholar] [CrossRef]
- Liu, B.; Lee, B.W.; Nakanishi, K.; Villasante, A.; Williamson, R.; Metz, J.; Kim, J.; Kanai, M.; Bi, L.; Brown, K.; et al. Cardiac recovery via extended cell-free delivery of extracellular vesicles secreted by cardiomyocytes derived from induced pluripotent stem cells. Nat. Biomed. Eng. 2018, 2, 293–303. [Google Scholar] [CrossRef]
- Veevers-Lowe, J.; Ball, S.G.; Shuttleworth, A.; Kielty, C.M. Mesenchymal stem cell migration is regulated by fibronectin through α5β1-integrin-mediated activation of PDGFR-β and potentiation of growth factor signals. J. Cell Sci. 2011, 124, 1288–1300. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Tian, H.; Li, H.; Ge, C.; Zhao, F.; Yao, M.; Li, J. Derivate isocorydine (d-ICD) suppresses migration and invasion of hepatocellular carcinoma cell by downregulating ITGA1 expression. Int. J. Mol. Sci. 2017, 18, 514. [Google Scholar] [CrossRef]
- Zheng, Q.M.; Chen, X.Y.; Bao, Q.F.; Yu, J.; Chen, L.H. ILK enhances migration and invasion abilities of human endometrial stromal cells by facilitating the epithelial–mesenchymal transition. Gynecol. Endocrinol. 2018, 34, 1091–1096. [Google Scholar] [CrossRef]
- Zu, L.; Ren, C.; Pan, B.; Zhou, B.; Zhou, E.; Niu, C.; Wang, X.; Zhao, M.; Gao, W.; Guo, L.; et al. Endothelial microparticles after antihypertensive and lipid-lowering therapy inhibit the adhesion of monocytes to endothelial cells. Int. J. Cardiol. 2016, 202, 756–759. [Google Scholar] [CrossRef]
- Jansen, F.; Yang, X.; Baumann, K.; Przybilla, D.; Schmitz, T.; Flender, A.; Paul, K.; Alhusseiny, A.; Nickenig, G.; Werner, N. Endothelial microparticles reduce ICAM-1 expression in a microRNA-222-dependent mechanism Cell culture and EMP generation. J. Cell. Mol. Med. 2015, 19, 2202–2214. [Google Scholar] [CrossRef]
- Garcia, N.A.; Ontoria-Oviedo, I.; González-King, H.; Diez-Juan, A.; Sepúlveda, P. Glucose starvation in cardiomyocytes enhances exosome secretion and promotes angiogenesis in endothelial cells. PLoS ONE 2015, 10, e0138849. [Google Scholar] [CrossRef]
- Todorova, D.; Simoncini, S.; Lacroix, R.; Sabatier, F.; Dignat-George, F. Extracellular vesicles in angiogenesis. Circ. Res. 2017, 120, 1658–1673. [Google Scholar] [CrossRef]
- Taraboletti, G.; D’Ascenzo, S.; Borsotti, P.; Giavazzi, R.; Pavan, A.; Dolo, V. Shedding of the matrix metalloproteinases MMP-2, MMP-9, and MT1-MMP as membrane vesicle-associated components by endothelial cells. Am. J. Pathol. 2002, 160, 673–680. [Google Scholar] [CrossRef]
- Crabbé, A.; Liu, Y.; Sarker, S.F.; Bonenfant, N.R.; Barrila, J.; Borg, Z.D.; Lee, J.J.; Weiss, D.J.; Nickerson, C.A. Recellularization of Decellularized Lung Scaffolds Is Enhanced by Dynamic Suspension Culture. PLoS ONE 2015, 10, e0126846. [Google Scholar] [CrossRef]
- Williams, C.; Budina, E.; Stoppel, W.L.; Sullivan, K.E.; Emani, S.; Emani, S.M.; Black, L.D. Cardiac extracellular matrix–fibrin hybrid scaffolds with tunable properties for cardiovascular tissue engineering. Acta Biomater. 2015, 14, 84–95. [Google Scholar] [CrossRef]
- Lee, J.S.; Cho, A.N.; Jin, Y.; Kim, J.; Kim, S.; Cho, S.W. Bio-artificial tongue with tongue extracellular matrix and primary taste cells. Biomaterials 2018, 151, 24–37. [Google Scholar] [CrossRef]
- Heuschkel, M.A.; Leitolis, A.; Roderjan, J.G.; Suss, P.H.; Luzia, C.A.O.; da Costa, F.D.A.; Correa, A.; Stimamiglio, M.A. In vitro evaluation of bovine pericardium after a soft decellularization approach for use in tissue engineering. Xenotransplantation 2018, e12464. [Google Scholar] [CrossRef]
- Costa, F.; Dohmen, P.; Vieira, E.; Lopes, S.V.; Colatusso, C.; Pereira, E.W.L.; Matsuda, C.N.; Cauduro, S. Ross Operation with decellularized pulmonary allografts: Medium-term results. Rev. Bras. Cir. Cardiovasc. 2007, 22, 454–462. [Google Scholar] [CrossRef]
- Namiri, M.; Ashtiani, M.K.; Abbasalizadeh, S.; Mazidi, Z.; Mahmoudi, E.; Nikeghbalian, S.; Aghdami, N.; Baharvand, H. Improving the biological function of decellularized heart valves through integration of protein tethering and three-dimensional cell seeding in a bioreactor. J. Tissue Eng. Regen. Med. 2018, 12, e1865–e1879. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Wang, H.; Zhou, J.; Li, H.; Liu, J.; Wang, Z.; Chen, A.; Zhao, Q. The Effect of Heparin-VEGF Multilayer on the Biocompatibility of Decellularized Aortic Valve with Platelet and Endothelial Progenitor Cells. PLoS ONE 2013, 8, e54622. [Google Scholar] [CrossRef] [PubMed]
- Jordan, J.E.; Williams, J.K.; Lee, S.; Raghavan, D.; Atala, A.; Yoo, J.J. Bioengineered self-seeding heart valves. J. Thorac. Cardiovasc. Surg. 2012, 143, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, D.; Hoerstrup, S.P. Tissue engineered heart valves based on human cells. Swiss Med. Wkly. 2007, 137 (Suppl. 155), 618. [Google Scholar]
- Cebotari, S.; Mertsching, H.; Kallenbach, K.; Kostin, S.; Repin, O.; Batrinac, A.; Kleczka, C.; Ciubotaru, A.; Haverich, A. Construction of autologous human heart valves based on an acellular allograft matrix. Circulation 2002, 106 (Suppl. 1), I63–I68. [Google Scholar] [PubMed]
- Angulski, A.B.B.; Capriglione, L.G.; Batista, M.; Marcon, B.H.; Senegaglia, A.C.; Stimamiglio, M.A.; Correa, A. The Protein Content of Extracellular Vesicles Derived from Expanded Human Umbilical Cord Blood-Derived CD133+ and Human Bone Marrow-Derived Mesenchymal Stem Cells Partially Explains Why both Sources are Advantageous for Regenerative Medicine. Stem Cell Rev. Rep. 2017, 13, 244–257. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Markers | LVE | RVE | AUE | LVM | RVM | AUM | MTL |
|---|---|---|---|---|---|---|---|
| CD90 | 22 ± 8.3 | 17.15 ± 3.0 | 19.9 ± 6.0 | 27.1 ± 9.0 | 55.95 ± 4.5 | 31.2 ± 2.4 | 36.5 ± 20.4 |
| CD105 | 92.05 ± 7.3 | 91.90 ± 7.4 | 84.65 ± 13.2 | 65.75 ± 17.0 | 94.75 ± 2.9 | 98.4 ± 0.2 | 98.65 ± 1.3 |
| CD 73 | 97.7 ± 2.0 | 99.20 ± 0.3 | 91.15 ± 7.8 | 76.05 ± 23.4 | 95.4 ± 4.0 | 99.25 ± 0.5 | 97.6 ± 1.3 |
| CD146 | 3.93 ± 3.5 | 1.25 ± 1.1 | 1.21 ± 0.6 | 0.91 ± 0.6 | 2.67 ± 0.8 | 1.35 ± 0.1 | 1.56 ± 0.2 |
| CD140b | 23.15 ± 10.5 | 32.80 ± 13.1 | 7.34 ± 1.0 | 53.60 ± 39.5 | 92.65 ± 1.1 | 67.05 ± 1.9 | 9.07 ± 1.9 |
| CD117 | 6.20 ± 1.8 | 8.10 ± 2.3 | 11.4 ± 7.9 | 11.15 ± 1.1 | 24.95 ± 19.6 | 8.28 ± 3.6 | 5.09 ± 2.3 |
| CD31 | 3.16 ± 1.4 | 2.13 ± 0.7 | 1.07 ± 0.4 | 1.71 ± 0.5 | 1.29 ± 0.3 | 0.96 ± 0.0 | 1.77 ± 0.1 |
| DDR2 | 30.95 ± 24.4 | 22.65 ± 0.2 | 3.68 ± 2.3 | 4.98 ± 2.9 | 13.36 ± 13.4 | 17.15 ± 7.2 | 29.78 ± 22.5 |
| Subject | Gender | Age | Weight | Race | LVM | LVE | RVM | RVE | AUM | AUE | MTL |
|---|---|---|---|---|---|---|---|---|---|---|---|
| A | M | 48 | 67 | White | / | + | + | / | + | / | + |
| C | M | 51 | 80 | Black | + | + | + | + | + | + | / |
| D | F | 50 | 60 | Black | / | + | / | / | + | + | + |
| E | M | 19 | 60 | Mix of Races | + | / | / | + | / | + | / |
| F | M | 42 | 115 | White | / | / | / | + | / | + | + |
| G | F | 56 | 56 | Mix of Races | + | / | + | / | + | / | / |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leitolis, A.; Suss, P.H.; Roderjan, J.G.; Angulski, A.B.B.; da Costa, F.D.A.; Stimamiglio, M.A.; Correa, A. Human Heart Explant-Derived Extracellular Vesicles: Characterization and Effects on the In Vitro Recellularization of Decellularized Heart Valves. Int. J. Mol. Sci. 2019, 20, 1279. https://doi.org/10.3390/ijms20061279
Leitolis A, Suss PH, Roderjan JG, Angulski ABB, da Costa FDA, Stimamiglio MA, Correa A. Human Heart Explant-Derived Extracellular Vesicles: Characterization and Effects on the In Vitro Recellularization of Decellularized Heart Valves. International Journal of Molecular Sciences. 2019; 20(6):1279. https://doi.org/10.3390/ijms20061279
Chicago/Turabian StyleLeitolis, Amanda, Paula Hansen Suss, João Gabriel Roderjan, Addeli Bez Batti Angulski, Francisco Diniz Affonso da Costa, Marco Augusto Stimamiglio, and Alejandro Correa. 2019. "Human Heart Explant-Derived Extracellular Vesicles: Characterization and Effects on the In Vitro Recellularization of Decellularized Heart Valves" International Journal of Molecular Sciences 20, no. 6: 1279. https://doi.org/10.3390/ijms20061279
APA StyleLeitolis, A., Suss, P. H., Roderjan, J. G., Angulski, A. B. B., da Costa, F. D. A., Stimamiglio, M. A., & Correa, A. (2019). Human Heart Explant-Derived Extracellular Vesicles: Characterization and Effects on the In Vitro Recellularization of Decellularized Heart Valves. International Journal of Molecular Sciences, 20(6), 1279. https://doi.org/10.3390/ijms20061279