ATP-Binding Cassette (ABC) Transporter Genes Involved in Pyrethroid Resistance in the Malaria Vector Anopheles sinensis: Genome-Wide Identification, Characteristics, Phylogenetics, and Expression Profile

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
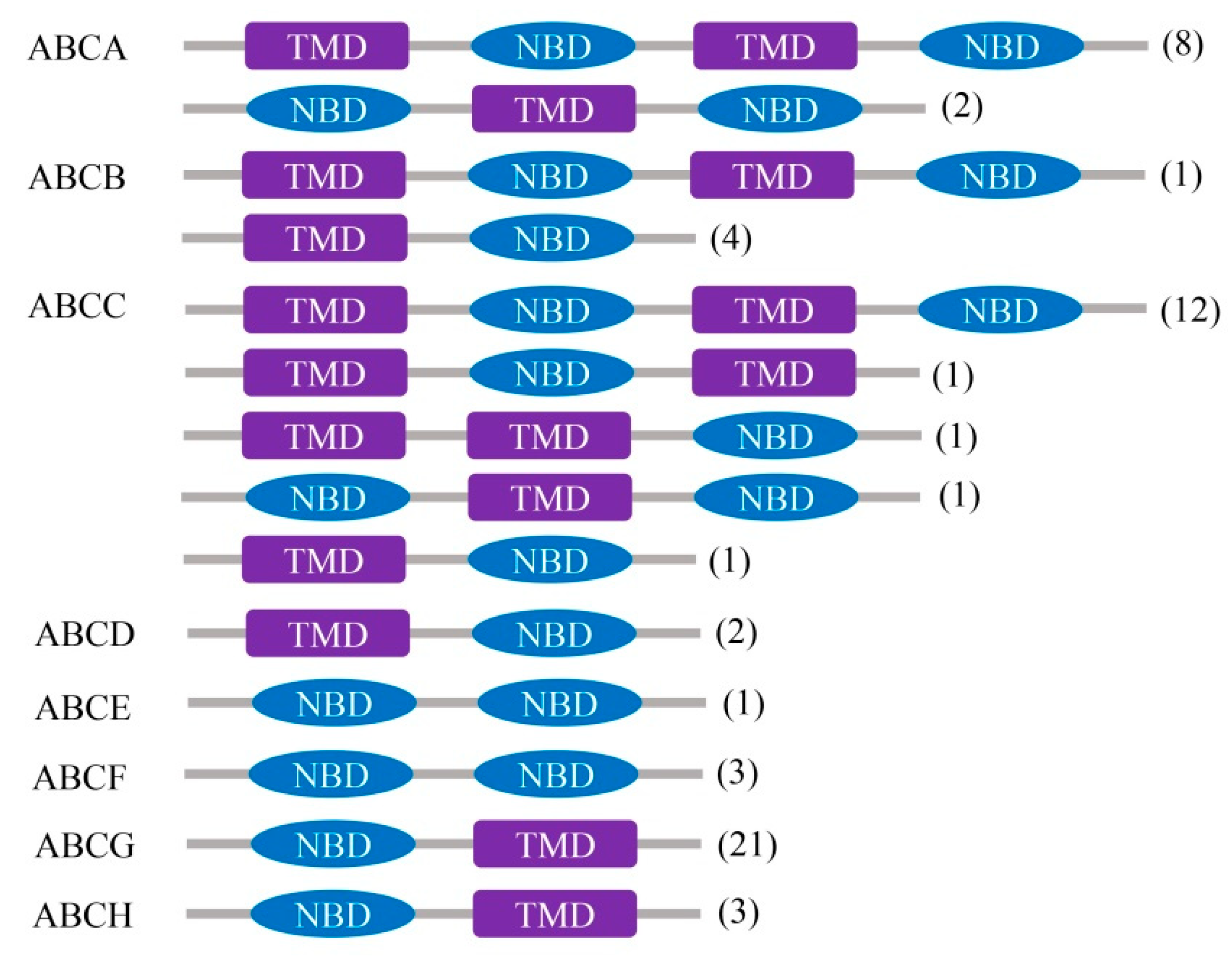
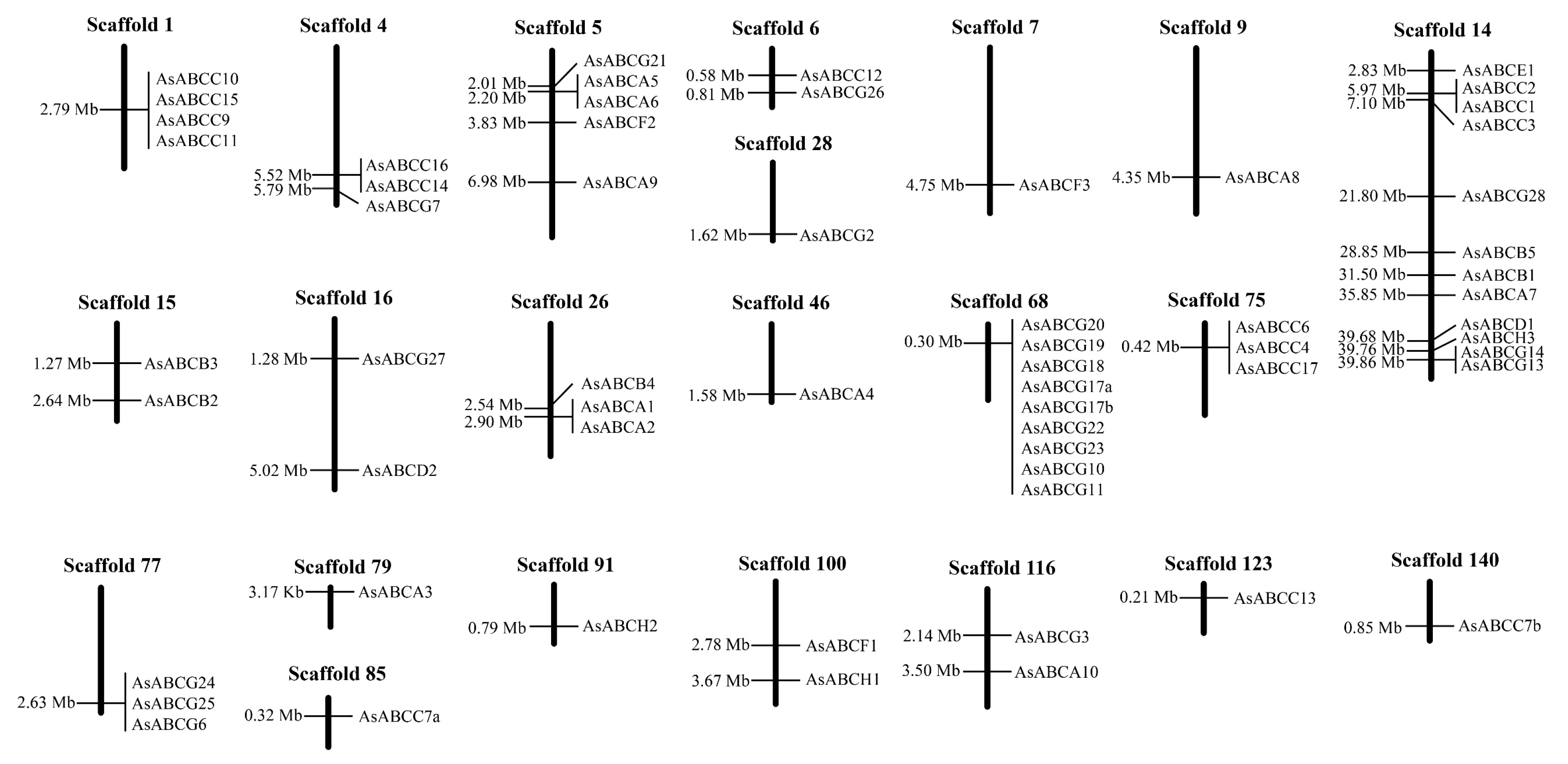
2.1. Diversity and Genomic Location of the A. sinensis ABC Transporter Genes
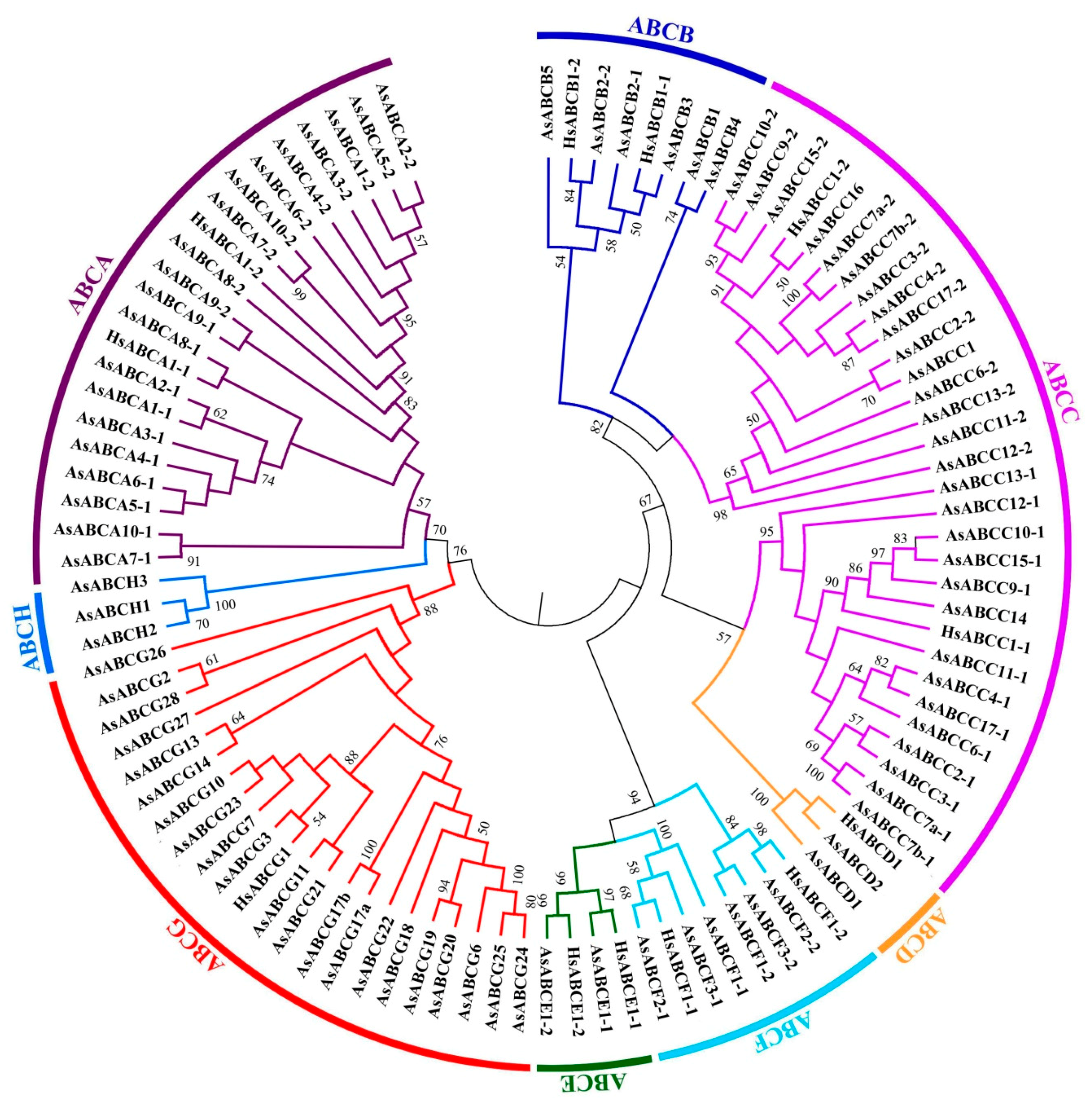
2.2. Phylogenetics and Characteristics of the A. sinensis ABC Transporter Genes
2.3. Ka/Ks of the Ortholog Genes in A. sinensis and A. gambiae
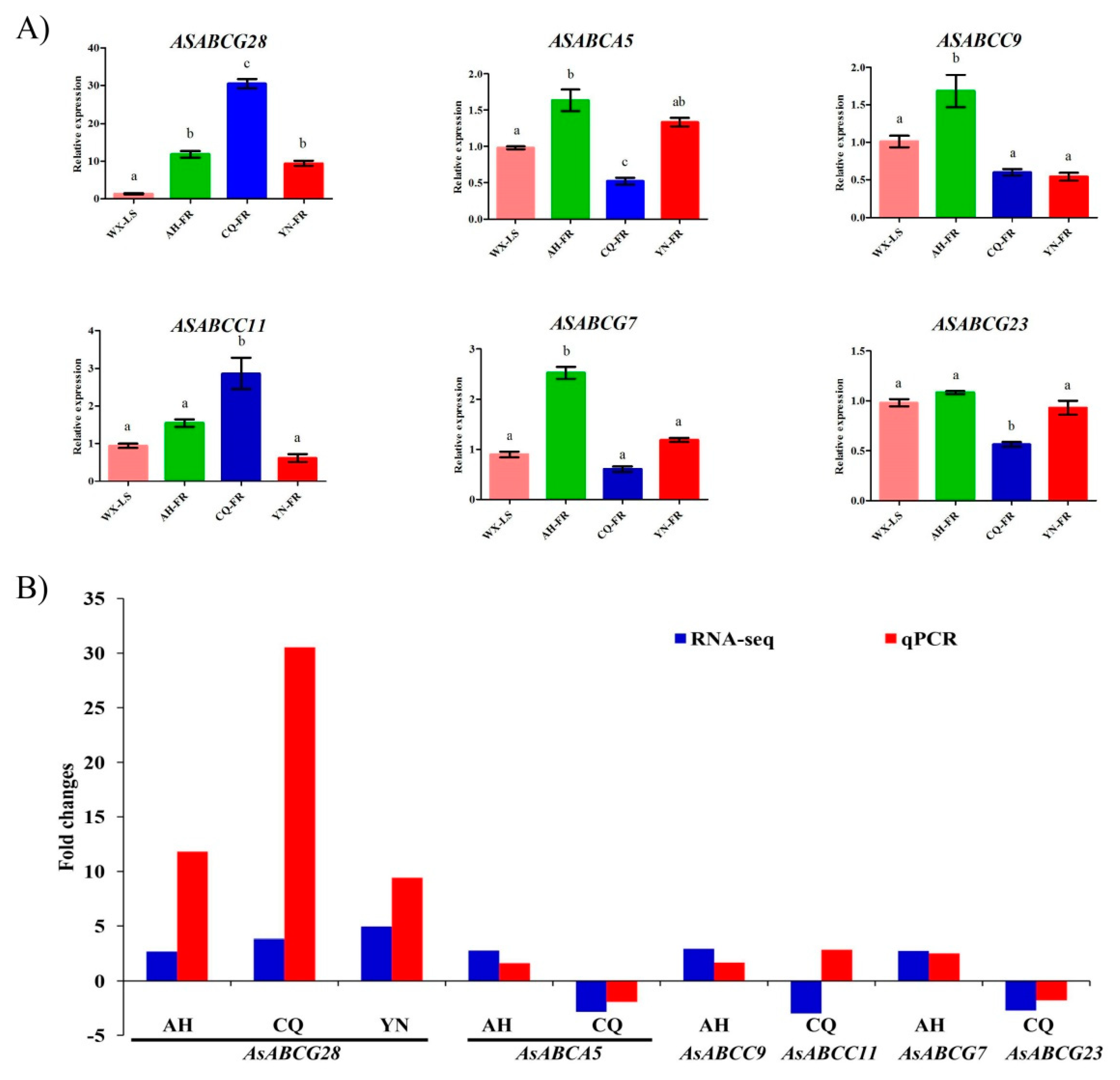
2.4. RNA-seq Expression Profile and qPCR Verification of ABC Transporters Associated with Pyrethorid Resistance
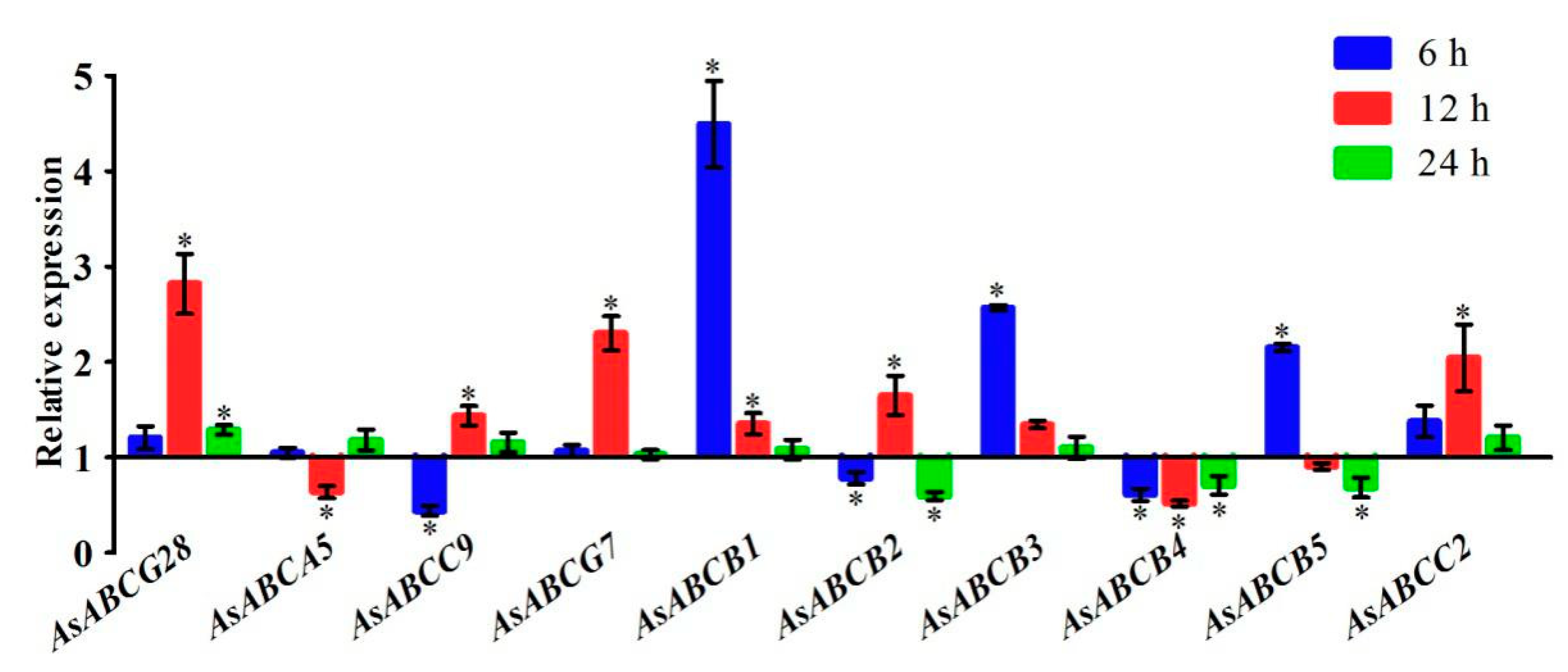
2.5. ABC Transporters Expression Subject to Deltamethrin Treatment
3. Materials and Methods
3.1. Mosquito Samples and Sequence Data
3.2. Identification and Characteristcs Analyses of A. sinensis ABC Transporter Genes
3.3. Phylogenetic Analysis of A. sinensis ABC Transporter Genes
3.4. Estimation of Synonymous Substitutions and Nonsynonymous Substitutions
3.5. RNA-seq Analysis and qPCR Verification of ABC Transporters Associated with Pyrethroid Resistance
3.6. Expression Changes of Pyrethroid Resistance-Relative ABC Transporter Genes Subject to Deltamethrin Treatment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dean, M.; Rzhetsky, A.; Allikmets, R. The human ATP-Binding cassette (ABC) transporter superfamily. Genome Res. 2001, 11, 1156–1166. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.F. ABC transporters: From microorganisms to man. Annu. Rev. Cell Biol. 1992, 8, 67–113. [Google Scholar] [CrossRef] [PubMed]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC transporters: The power to change. Nat. Rev. Mol. Cell Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dermauw, W.; Leeuwen, T.V. The ABC gene family in arthropods: Comparative genomics and role in insecticide transport and resistance. Insect Biochem. Mol. Biol. 2014, 45, 89–110. [Google Scholar] [CrossRef] [Green Version]
- Strauss, A.S.; Wang, D.; Stock, M.; René, R.G.; Groth, M.; Boland, W.; Burse, A. Tissue-specific transcript profiling for ABC transporters in the sequestering larvae of the phytophagous leaf beetle Chrysomela populi. PLoS ONE 2014, 9, e98637. [Google Scholar] [CrossRef] [PubMed]
- Hollenstein, K.; Dawson, R.J.; Locher, K.P. Structure and mechanism of ABC transporter proteins. Curr. Opin. Struct. Biol. 2007, 17, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhou, S.; Tian, L.; Guo, E.; Luan, Y.; Zhang, J.; Li, S. Genome-wide identification and characterization of ATP-binding cassette transporters in the silkworm, Bombyx mori. BMC Genom. 2001, 12, 476–491. [Google Scholar] [CrossRef]
- Broehan, G.; Kroeger, T.; Lorenzen, M.; Merzendorfer, H. Functional analysis of the ATP-binding cassette (ABC) transporter gene family of Tribolium castaneum. BMC Genom. 2013, 14, 6–25. [Google Scholar] [CrossRef]
- WHO. Pesticides and Their Aapplication for the Control of Vectors and Pests of Public Health Importance; World Health Organization: Geneva, Switzerland, 2006. [Google Scholar]
- Phillips, R.S. Current status of malaria and potential for control. Clin. Microbiol. Rev. 2001, 14, 208–226. [Google Scholar] [CrossRef]
- Hemingway, J.; Field, L.; Vontas, J. An overview of insecticide resistance. Science 2002, 298, 96–97. [Google Scholar] [CrossRef]
- Ranson, H.; N’guessan, R.; Lines, J.; Moiroux, N.; Nkuni, Z.; Corbel, V. Pyrethroid resistance in african anopheline mosquitoes: What are the implications for malaria control? Trends Parasitol. 2011, 27, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Hemingway, J.; Hawkes, N.J.; Mccarroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Mol. 2004, 34, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.M.; Xu, B.Y.; Si, F.L.; Li, J.J.; Yan, Z.T.; Yan, Z.W.; He, X.; Chen, B. Identification of carboxylesterase genes associated with pyrethroid resistance in the malaria vector Anopheles sinensis (Diptera: Culicidae): Carboxylesterase genes in Anopheles sinensis. Pest Manag. Sci. 2018, 74, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.W.; He, Z.B.; Yan, Z.T.; Si, F.L.; Zhou, Y.; Chen, B. Genome-wide and expression-profiling analyses suggest the main cytochrome P450 genes related to pyrethroid resistance in the malaria vector, Anopheles sinensis (Diptera Culicidae). Pest Manag. Sci. 2018, 74, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Bariami, V.; Jones, C.M.; Poupardin, R.; Ranson, H. Gene amplification, ABC transporters and cytochrome P450s: Unraveling the molecular basis of pyrethroid resistance in the dengue vector, Aedes aegypti. PLoS Negl. Trop. Dis. 2012, 6, e1692. [Google Scholar] [CrossRef] [PubMed]
- Bonizzoni, M.; Afrane, Y.; Dunn, W.A.; Atieli, F.K.; Zhou, G.; Zhong, D.; Li, J.; Githeko, A.; Yan, G. Comparative transcriptome analyses of deltamethrin resistant and -susceptible Anopheles gambiae mosquitoes from Kenya by RNASeq. PLoS ONE 2012, 7, e44607. [Google Scholar] [CrossRef]
- Epis, S.; Porretta, D.; Mastrantonio, V.; Comandatore, F.; Sassera, D.; Rossi, P.; Cafarchia, C.; Otranto, D.; Favia, G.; Genchi, C.; et al. ABC transporters are involved in defense against permethrin insecticide in the malaria vector Anopheles stephensi. Parasite Vector 2014, 7, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Epis, S.; Porretta, D.; Mastrantonio, V.; Urbanelli, S.; Sassera, D.; Marco, L.D.; Mereghetti, V.; Montagna, M.; Ricci, I.; Favia, G.; et al. Temporal dynamics of the ABC transporter response to insecticide treatment: Insights from the malaria vector Anopheles stephensi. Sci. Rep. 2014, 4, 7435–7439. [Google Scholar] [CrossRef] [PubMed]
- Mastrantonio, V.; Ferrari, M.; Epis, S.; Negri, A.; Scuccimarra, G.; Favia, G.; Porretta, D.; Urbanelli, S.; Bandi, C. Gene expression modulation of ABC transporter genes in response to permethrin in adults of the mosquito malaria vector Anopheles stephensi. Acta Trop. 2017, 171, 37–43. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, Y.J.; He, Z.B.; Li, W.S.; Si, F.L.; Tang, Y.; He, Q.Y.; Qiao, L.; Yan, Z.T.; Fu, W.B.; et al. De novo transcriptome sequencing and sequence analysis of the malaria vector Anopheles sinensis (Diptera: Culicidae). Parasite Vector 2014, 7, 314–325. [Google Scholar] [CrossRef]
- Qin, Q.; Li, Y.; Zhong, D.; Zhou, N.; Chang, X.; Cui, C.; Yan, L.; Chen, X.G. Insecticide resistance of Anopheles sinensis and An. vagus in Hainan Island, a malaria-endemic area of China. Parasites Vectors 2014, 7, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Mei, T.; Fu, W.B.; Li, B.; He, Z.B.; Chen, B. Comparative genomics of chemosensory protein genes (CSPs) in twenty-two mosquito species (Diptera: Culicidae): Identification, characterization, and evolution. PLoS ONE 2018, 13, e0190412. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.Q.; Qiao, L.; He, Q.Y.; Zhou, Y.; Ren, S.; Chen, B. Genome-wide identification, characterization and evolution of cuticular protein genes in the malaria vector Anopheles sinensis (Diptera: Culicidae). Curr. Opin. Insect Sci. 2018, 25, 739–750. [Google Scholar] [CrossRef]
- He, X.; He, Z.B.; Zhang, Y.J.; Zhou, Y.; Xian, P.J.; Qiao, L.; Chen, B. Genome-wide identification and characterization of odorant-binding proteins of genes in the malaria vector Anopheles sinensis (Diptera: Culicidae). Curr. Opin. Insect Sci. 2016, 23, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Si, F.L.; He, Z.B.; Chen, B. Genome-wide identification, characterization and classification of ionotropic glutamate receptor genes (iGluRs) in the malaria vector Anopheles sinensis (Diptera: Culicidae). Parasite Vector 2018, 11, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Roth, C.W.; Holm, I.; Graille, M.; Dehoux, P.; Rzhetsky, A.; Wincker, P.; Weissenbach, J.; Brey, P.T. Identification of the Anopheles gambiae ATP-binding cassette transporter superfamily genes. Mol. Cells 2003, 15, 150–158. [Google Scholar]
- Dutta, S. Genetic Regulation of Autophagic Cell Death in Drosophila melanogaster. Ph.D. Thesis, University of Maryland, Department Mol. Cell Bio., College Park, MD, USA, 2008; p. 130. [Google Scholar]
- Sturm, A.; Cunningham, P.; Dean, M. The ABC transporter gene family of Daphnia pulex. BMC Genom. 2009, 10, 170–187. [Google Scholar] [CrossRef] [PubMed]
- Decottignies, A.; Goffeau, A. Complete inventory of the yeast ABC proteins. Nat. Genet. 1997, 15, 137–145. [Google Scholar] [CrossRef]
- Sanchez-Fernandez, R.; Davies, T.G.E.; Coleman, J.O.D.; Rea, P.A. The Arabidopsis thaliana ABC Protein Superfamily, a Complete Inventory. J. Biol. Chem. 2001, 276, 30231–30244. [Google Scholar] [CrossRef] [Green Version]
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug resistance in cancer: Role of ATP-dependent transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar] [CrossRef]
- Mayer, F.; Mayer, N.; Chinn, L.; Pinsonneault, R.L.; Kroetz, D.; Bainton, R.J. Evolutionary conservation of vertebrate blood-brain barrier chemoprotective mechanisms in Drosophila. J. Neurosci. 2009, 29, 3538–3550. [Google Scholar] [CrossRef] [PubMed]
- Ricardo, S.; Lehmann, R. An ABC transporter controls export of a Drosophila germ cell attractant. Science 2009, 323, 943–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vache, C.; Camares, O.; Cardoso-Ferreira, M.C.; Dastugue, B.; Creveaux, I.; Vaury, C.; Bamdad, M. A potential genomic biomarker for the detection of polycyclic aromatic hydrocarbon pollutants: Multidrug resistance gene 49 in Drosophila melanogaster. Environ. Toxicol. Chem. 2007, 26, 1418–1424. [Google Scholar] [CrossRef]
- Buss, D.S.; Callaghan, A. Interaction of pesticides with p-glycoprotein and other ABC proteins: A survey of the possible importance to insecticide, herbicide and fungicide resistance. Pestic. Biochem. Physiol. 2008, 90, 141–153. [Google Scholar] [CrossRef]
- Hopper-Borge, E.; Xu, X.; Shen, T.; Shi, Z.; Chen, Z.S.; Kruh, G.D. Human multidrug resistance protein 7 (ABCC10) is a resistance factor for nucleoside analogues and epothilone B. Cancer Res. 2009, 69, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Yepiskoposyan, H.; Egli, D.; Fergestad, T.; Selvaraj, A.; Treiber, C.; Multhaup, G.; Georgiev, O.; Schaffner, W. Transcriptome response to heavy metal stress in Drosophila reveals a new zinc transporter that confers resistance to zinc. Nucleic Acids Res. 2006, 34, 4866–4877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, A. Peroxisomal ABC transporters. FEBS Lett. 2006, 580, 1139–1155. [Google Scholar] [Green Version]
- Kerr, I.D. Sequence analysis of twin ATP binding cassette proteins involved in translational control, antibiotic resistance, and ribonuclease L inhibition. Biochem. Biophys. Res. Commun. 2004, 315, 166–173. [Google Scholar] [CrossRef]
- Tian, Y.; Han, X.; Tian, D.L. The biological regulation of ABCE1. Iubmb Life 2012, 64, 795–800. [Google Scholar] [CrossRef] [Green Version]
- Paytubi, S.; Wang, X.; Lam, Y.W.; Izquierdo, L.; Hunter, M.J.; Jan, E.; Hundal, H.S.; Proud, C.G. ABC50 promotes translation initiation in mammalian cells. J. Biol. Chem. 2009, 284, 24061–24073. [Google Scholar] [CrossRef]
- Kovalchuk, A.; Driessen, A.J. Phylogenetic analysis of fungal ABC transporters. BMC Genom. 2010, 11, 177–197. [Google Scholar] [CrossRef] [PubMed]
- Verrier, P.J.; Bird, D.; Buria, B.; Dassa, E.; Forestier, C.; Geisler, M.; Klein, M.; Kolukisaoglu, U.; Lee, Y.; Martinoia, E. Plant ABC proteins a unified nomenclature and updated inventory. Trends Plant Sci. 2008, 13, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, D.T.; Grillo, S.L.; Kitos, R.J. Subcellular localization of the first three enzymes of the ommochrome synthetic pathway in Drosophila melanogaster. J. Exp. Zool. 1974, 188, 225–233. [Google Scholar] [CrossRef]
- Mackenzie, S.M.; Brooker, M.; Gill, T.R.; Cox, G.B.; Howells, A.J.; Ewart, G.D. Mutations in the white gene of Drosophila melanogaster affecting ABC transporters that determine eye colouration. Biochim. Biophys. Acta 1999, 1419, 173–185. [Google Scholar] [CrossRef] [Green Version]
- Komoto, N.; Quan, G.; Sezutsu, H.; Tamura, T. A single-base deletion in an ABC transporter gene causes white eyes, white eggs, and translucent larval skin in the silkworm w-3(oe) mutant. Insect Biochem. Mol. Biol. 2009, 39, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Pedra, J.H.F.; Mcintyre, L.M.; Scharf, M.E.; Pittendrigh, B.R. Genome-wide transcription profile of field- and laboratory-selected dichlorodiphenyltrichloroethane (DDT)-resistant Drosophila. Proc. Natl. Acad. Sci. USA 2004, 101, 7034–7039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, M.; Yue, Z.; He, W.; Yang, X.; Yang, G.; Xie, M.; Zhan, D.; Baxter, S.W.; Vasseur, L.; Gurr, G.M.; et al. A heterozygous moth genome provides insights into herbivory and detoxification. Nat. Genet. 2013, 45, 220–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, N.; Xie, W.; Jones, C.M.; Bass, C.; Jiao, X.; Yang, X.; Liu, B.; Li, R.; Zhang, Y. Transcriptome profiling of the whitefly Bemisia tabaci reveals stage-specific gene expression signatures for thiamethoxam resistance. Insect Mol. Biol. 2013, 22, 485–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, N.; Xie, W.; Yang, X.; Wang, S.; Wu, Q.; Li, R.; Pan, H.; Liu, B.; Shi, X.; Fang, Y.; et al. Transcriptomic and proteomic responses of sweetpotato whitefly, Bemisia tabaci, to thiamethoxam. PLoS ONE 2013, 8, e61820. [Google Scholar] [CrossRef]
- Jones, C.M.; Toé, H.K.; Sanou, A.; Namountougou, M.; Hughes, A.; Diabaté, A.; Dabiré, R.; Simard, F.; Ranson, H. Additional selection for insecticide resistance in urban malaria vectors: DDT resistance in Anopheles arabiensis from Bobo-Dioulasso, Burkina Faso. PLoS ONE 2012, 79, e45995. [Google Scholar] [CrossRef]
- Annilo, T.; Chen, Z.; Shulenin, S.; Costantino, J.; Thomas, L.; Lou, H.; Stefanov, S.; Dean, M.; Rzhetsky, A.; Allikmets, R. Evolution of the vertebrate ABC gene family: Analysis of gene birth and death. Genomics 2006, 88, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, A.X.; Jander, G.; Samaniego, H.; Ramsey, J.S.; Figueroa, C.C. Insecticide resistance mechanisms in the green peach aphid Myzus persicae (Hemiptera: Aphididae) I: A transcriptomic survey. PLoS ONE 2012, 7, e36366. [Google Scholar] [CrossRef]
- Tian, L.; Song, T.; He, R.; Zeng, Y.; Xie, W.; Wu, Q. Genome-wide analysis of atp-binding cassette (ABC) transporters in the sweetpotato whitefly, Bemisia tabaci. BMC Genom. 2017, 18, 330–345. [Google Scholar] [CrossRef] [PubMed]
- Pohl, P.C.; Klafke, G.M.; Carvalho, D.D.; Martins, J.R.; Daffre, S.; da Silva Vaz, I., Jr.; Masuda, A. ABC transporter efflux pumps: A defense mechanism against ivermectin in Rhipicephalus (Boophilus) microplus. Int. J. Parasitol. 2011, 41, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Figueira-Mansur, J.; Ferreira-Pereira, A.; Mansur, J.F.; Franco, T.A.; Alvarenga, E.S.; Sorgine, M.H.; Neves, B.C.; Melo, A.C.; Leal, W.S.; Masuda, H.; et al. Silencing of P-glycoprotein increases mortality in temephos-treated Aedes aegyptilarvae. Insect Mol. Biol. 2013, 22, 648–658. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Sun, Y.J.; Yang, L.; Huang, S.; Wu, Y.J. Avermectin induces P-glycoprotein expression in S2 cells via the calcium/calmodulin/NF-kB pathway. Chem. Biol. Interact. 2013, 203, 430–439. [Google Scholar] [CrossRef]
- Srinivas, R.; Shamsundar, G.S.; Jayalakshmi, S.K.; Sreermalu, K. Effect of insecticides and inhibitors on P-glycoprotein ATPase (M-type) activity of resistant pest Helicoverpa armigera. Curr. Sci. 2005, 88, 1449–1452. [Google Scholar]
- Simmons, J.; D’Souza, O.; Rheault, M.; Donly, C. Multidrug resistance protein gene expression in Trichoplusia ni caterpillars. Insect Mol. Biol. 2013, 22, 62–71. [Google Scholar] [CrossRef]
- Fossog, T.B.; Poupardin, R.; Costantini, C.; Awono-Ambene, P.; Wondji, C.S.; Ranson, H.; Antonio-Nkondjio, C. Resistance to DDT in an urban setting: Common mechanisms implicated in both M and S forms of Anopheles gambiae in the city of Yaoundé Cameroon. PLoS ONE 2013, 8, e61408. [Google Scholar] [CrossRef]
- Zhu, G.; Zhong, D.; Cao, J.; Zhou, H.; Li, J.; Liu, Y.; Bai, L.; Xu, S.; Wang, M.H.; Zhou, G.; et al. Transcriptome profiling of pyrethroid resistant and susceptible mosquitoes in the malaria vector Anopheles sinensis. BMC Genom. 2014, 15, 448–461. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- WHO. Test Procedures for Insecticide Resistance Monitoring in Malaria Vectors, Bio-Efficacy and Persistence of Insecticides on Treated Surfaces; World Health Organization: Geneva, Switzerland, 1998. [Google Scholar]
- Gao, Q.; Beebe, N.W.; Cooper, R.D. Molecular identification of the malaria vectors Anopheles anthropophagus and Anopheles sinensis (Diptera: Culicidae) in central China using polymerase chain reaction and appraisal of their position within the Hyrcanus group. J. Med. Entomol. 2004, 41, 5–11. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Lalitha, S. Primer premier 5. Biotech. Softw. Internet. Rep. 2000, 1, 270–272. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Anal. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Annotation ID | Genome Location a | Number of Exon | Gene Length (bp) (Exon + Intron) | Amino Acid Number (AA) | EST Support b |
|---|---|---|---|---|---|---|
| ABCA Subfamily (10 genes) | ||||||
| AsABCA1 | ASI10006665 | scaffold26:2898415-2904542(−) | 10 | 4839+1289 | 1613 | + |
| AsABCA2 | ASI10006666 | scaffold26:2907038-2912368(−) | 7 | 5304+3051 | 1768 | + |
| AsABCA3 | ASI10001787 | scaffold79:3171-8550(+) | 7 | 4956+424 | 1652 | + |
| AsABCA4 | ASI10005446 | scaffold46:1576155-1586398(−) | 7 | 4965+5279 | 1655 | + |
| AsABCA5 | ASI10011585 | scaffold5:2204987-2211927(−) | 8 | 4989+1952 | 1663 | + |
| AsABCA6 | ASI10011586 | scaffold5:2214289-2220237(−) | 6 | 4887+1062 | 1629 | + |
| AsABCA7 | ASI10017076 | scaffold14:35853362-35861993(−) | 7 | 4941+1825 | 1647 | + |
| AsABCA8 | ASI10008148 | scaffold9:4353984-4361438(+) | 9 | 5979+1476 | 1993 | + |
| AsABCA9 | ASI10012001 | scaffold5:6985656-6991716(−) | 14 | 5007+1054 | 1669 | + |
| AsABCA10 | ASI10008529 | scaffold116:3498475-3502806(−) | 5 | 3417+915 | 1139 | + |
| ABCB Subfamily (5 genes) | ||||||
| AsABCB1 | ASI10016633 | scaffold14:31503016-31505870(−) | 4 | 2547+308 | 849 | + |
| AsABCB2 | ASI10010010 | scaffold15:2635069-2641163(+) | 11 | 3882+2213 | 1294 | + |
| AsABCB3 | ASI10009929 | scaffold15:1271785-1274177(−) | 4 | 2181+212 | 727 | + |
| AsABCB4 | ASI10006641 | scaffold26:2544630-2550018(+) | 7 | 2280+3109 | 760 | + |
| AsABCB5 | ASI10016463 | scaffold14:28845118-28848788(−) | 9 | 1965+1706 | 655 | + |
| ABCC Subfamily (16 genes) | ||||||
| AsABCC1 | ASI10014633 | scaffold14:5983931-5989564(+) | 7 | 3534+2100 | 1178 | + |
| AsABCC2 | ASI10014632 | scaffold14:5970014-5980211(−) | 6 | 4140+6058 | 1380 | + |
| AsABCC3 | ASI10014742 | scaffold14:7100932-7109199(+) | 9 | 4347+3921 | 1449 | + |
| AsABCC4 | ASI10004082 | scaffold75:426548-436333(−) | 12 | 4404+5382 | 1468 | + |
| AsABCC6 | ASI10004081 | scaffold75:419144-426092(−) | 11 | 3972+2977 | 1324 | - |
| AsABCC7a | ASI10001723 | scaffold85:324079-329862(+) | 7 | 4236+1548 | 1412 | + |
| AsABCC7b | ASI10002723 | scaffold140:852869-858944(+) | 7 | 4236+1840 | 1412 | + |
| AsABCC9 | ASI10007114 | scaffold1:2805165-2810461(−) | 6 | 4512+785 | 1504 | + |
| AsABCC10 | ASI10007112 | scaffold1:2792623-2797620(−) | 7 | 4521+477 | 1507 | + |
| AsABCC11 | ASI10007115 | scaffold1:2812936-2817655(−) | 4 | 4212+508 | 1404 | + |
| AsABCC12 | ASI10003245 | scaffold6:579715-584317(−) | 3 | 4416+187 | 1472 | + |
| AsABCC13 | ASI10001014 | scaffold123:207484-225544(−) | 18 | 4017+14044 | 1339 | + |
| AsABCC14 | ASI10009681 | scaffold4:5523940-5529838(−) | 11 | 3453+2446 | 1151 | + |
| AsABCC15 | ASI10007113 | scaffold1:2799112-2804048(−) | 7 | 4515+422 | 1505 | + |
| AsABCC16 | ASI10009679 | scaffold4:5516323-5523342(−) | 9 | 2199+4821 | 733 | + |
| AsABCC17 | ASI10004083 | scaffold75:439279-443721(−) | 4 | 4203+240 | 1401 | + |
| ABCD Subfamily (2 genes) | ||||||
| AsABCD1 | ASI10017457 | scaffold14:39675247-39680568(−) | 6 | 2226+3096 | 742 | + |
| AsABCD2 | ASI10013565 | scaffold16:5022027-5030607(−) | 8 | 2085+6496 | 695 | + |
| ABCE Subfamily (1 gene) | ||||||
| AsABCE1 | ASI10014410 | scaffold14:2830364-2832333(+) | 3 | 1827+143 | 609 | + |
| ABCF Subfamily (3 genes) | ||||||
| AsABCF1 | ASI10006319 | scaffold100:2782009-2783929(−) | 2 | 1815+106 | 605 | + |
| AsABCF2 | ASI10011705 | scaffold5:3835256-3838673(−) | 2 | 2805+613 | 935 | + |
| AsABCF3 | ASI10009140 | scaffold7:4751204-4753620(+) | 4 | 2175+242 | 725 | + |
| ABCG Subfamily (21 genes) | ||||||
| AsABCG2 | ASI10004617 | scaffold28:1623903-1626178(−) | 5 | 1899+377 | 633 | + |
| AsABCG3 | ASI10008396 | scaffold116:2141408-2144367(+) | 6 | 2286+674 | 762 | + |
| AsABCG6 | ASI10005115 | scaffold77:2634493-2634706(−) | 6 | 1599+331 | 533 | + |
| AsABCG7 | ASI10009696 | scaffold4:5791835-5801142(−) | 7 | 2091+7217 | 697 | + |
| AsABCG10 | ASI10002998 | scaffold68:338877-350631(−) | 9 | 2073+9682 | 691 | + |
| AsABCG14 | ASI10017482 | scaffold14:39864271-39876401(−) | 8 | 1806+10325 | 602 | + |
| AsABCG13 | ASI10017484 | scaffold14:39904223-39909126(+) | 7 | 2297+2607 | 869 | + |
| AsABCG17a | ASI10002994 | scaffold68:308541-310459(−) | 3 | 1779+140 | 593 | + |
| AsABCG17b | ASI10002995 | scaffold68:315600-317518(−) | 3 | 1779+140 | 593 | + |
| AsABCG18 | ASI10002993 | scaffold68:304413-306415(−) | 4 | 1782+241 | 594 | + |
| AsABCG19 | ASI10002992 | scaffold68:299889-301953(−) | 5 | 1800+265 | 600 | + |
| AsABCG20 | ASI10002991 | scaffold68:295550-298589(−) | 5 | 1710+1330 | 570 | + |
| AsABCG21 | ASI10011562 | scaffold5:2012784-2021328(+) | 7 | 1839+6706 | 613 | + |
| AsABCG22 | ASI10002996 | scaffold68:318211-320277(−) | 5 | 1794+273 | 598 | + |
| AsABCG23 | ASI10002997 | scaffold68:321662-335687(−) | 7 | 2100+11926 | 700 | + |
| AsABCG11 | ASI10003001 | scaffold68:361453-367680(−) | 8 | 1995+4233 | 665 | + |
| AsABCG24 | ASI10005113 | scaffold77:2627164-2629127(−) | 6 | 1584+380 | 528 | - |
| AsABCG25 | ASI10005114 | scaffold77:2629922-2632124(−) | 7 | 1791+412 | 597 | - |
| AsABCG26 | ASI10003266 | scaffold6:809083-811434(−) | 7 | 1839+513 | 613 | + |
| AsABCG27 | ASI10014141 | scaffold16:12792427-12799169(−) | 5 | 1962+4781 | 654 | + |
| AsABCG28 | ASI10015886 | scaffold14:21804555-21808651(+) | 6 | 1833+17432 | 611 | + |
| ABCH Subfamily (3 genes) | ||||||
| AsABCH1 | ASI10006388 | scaffold100:3673129-3675938(−) | 7 | 2328+482 | 776 | + |
| AsABCH2 | ASI10003817 | scaffold91:793497-800786(+) | 11 | 2133+5157 | 711 | + |
| AsABCH3 | ASI10017471 | scaffold14:39764080-39771407(+) | 10 | 2172+5156 | 724 | + |
| ABC Subfamily | A. sinensis | A. gambiae | D. melanogaster | A. mellifera | B. Mori | T. castaneum | H. sapiens |
|---|---|---|---|---|---|---|---|
| ABCA | 10 | 9 | 10 | 3 | 6 | 10 | 13 |
| ABCB | 5 | 5 | 8 | 5 | 8 | 6 | 11 |
| ABCC | 16 | 13 | 14 | 9 | 15 | 35 | 12 |
| ABCD | 2 | 2 | 2 | 2 | 2 | 2 | 4 |
| ABCE | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| ABCF | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| ABCG | 21 | 16 | 15 | 15 | 13 | 13 | 5 |
| ABCH | 3 | 3 | 3 | 3 | 3 | 3 | 0 |
| Total | 61 | 52 | 56 | 41 | 51 | 73 | 48 |
| Gene Name | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Size (Base Pairs) |
|---|---|---|---|
| AsABCG28 | CAACCTGTACTCCACCACCC | TAACGACCAGACCGAGCAAC | 140 |
| AsABCA5 | GGGAGTTCAAGTGTCTCGGG | TGTACTGTTTGACGTCCGCT | 129 |
| AsABCC9 | TGCGTAGCCGGTTGACTATC | AATGAGACAGCTCCAGTGCC | 126 |
| AsABCG7 | ATCTGAACCTCGGCGGAAAC | CCCAGAACTGGTGGAACTCG | 198 |
| AsABCB1 | GTAAAACCGGCGAAGTGCTG | GCGTCATGGTGAGAAACACG | 177 |
| AsABCB2 | GTGAGATCCCGATGCAGGAG | TTCTGACCACCGGACAGTTG | 130 |
| AsABCB3 | TTCTCGGTTCGGTCATTCGG | ACACAAACTCTACGCCCTCG | 150 |
| AsABCB4 | GTTTCACGACACCATTCGGC | AACTTAAGGCCACGCTCTCC | 156 |
| AsABCB5 | ACGGGTTGAACATGGGTGAG | GATAGCACGGGCGATAGCAA | 176 |
| AsABCC2 | GGAGAGGTGCTGATCGATGG | CTGGGTAGTCCTCGAAGGGA | 142 |
| RPS7 | CGGAGAAGATGGCATGGGAGAT | ATAGTGAGCATAGGCCCGGTTA | 148 |
| RPL49 | GGAGCCGGTCGGTGATATGT | TTCCTTCTCGGTCGGCTTCG | 121 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Q.; Yan, Z.; Si, F.; Zhou, Y.; Fu, W.; Chen, B. ATP-Binding Cassette (ABC) Transporter Genes Involved in Pyrethroid Resistance in the Malaria Vector Anopheles sinensis: Genome-Wide Identification, Characteristics, Phylogenetics, and Expression Profile. Int. J. Mol. Sci. 2019, 20, 1409. https://doi.org/10.3390/ijms20061409
He Q, Yan Z, Si F, Zhou Y, Fu W, Chen B. ATP-Binding Cassette (ABC) Transporter Genes Involved in Pyrethroid Resistance in the Malaria Vector Anopheles sinensis: Genome-Wide Identification, Characteristics, Phylogenetics, and Expression Profile. International Journal of Molecular Sciences. 2019; 20(6):1409. https://doi.org/10.3390/ijms20061409
Chicago/Turabian StyleHe, Qiyi, Zhentian Yan, Fengling Si, Yong Zhou, Wenbo Fu, and Bin Chen. 2019. "ATP-Binding Cassette (ABC) Transporter Genes Involved in Pyrethroid Resistance in the Malaria Vector Anopheles sinensis: Genome-Wide Identification, Characteristics, Phylogenetics, and Expression Profile" International Journal of Molecular Sciences 20, no. 6: 1409. https://doi.org/10.3390/ijms20061409
APA StyleHe, Q., Yan, Z., Si, F., Zhou, Y., Fu, W., & Chen, B. (2019). ATP-Binding Cassette (ABC) Transporter Genes Involved in Pyrethroid Resistance in the Malaria Vector Anopheles sinensis: Genome-Wide Identification, Characteristics, Phylogenetics, and Expression Profile. International Journal of Molecular Sciences, 20(6), 1409. https://doi.org/10.3390/ijms20061409