Cytological and Proteomic Analysis of Wheat Pollen Abortion Induced by Chemical Hybridization Agent

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
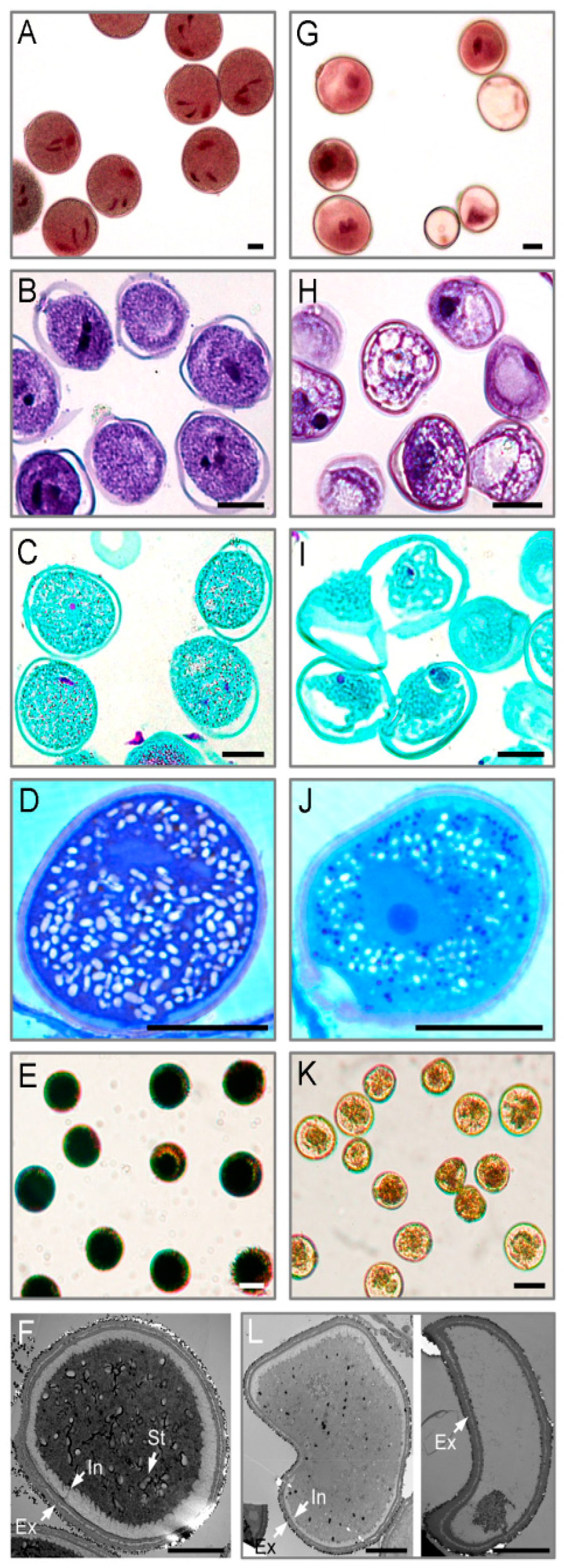
2.1. Cytological Changes in Pollen Abortion Induced by CHA-SQ-1 in Wheat
2.2. Pollen Grain Proteomic Analysis of CHA-SQ-1-Treated Wheat Plants
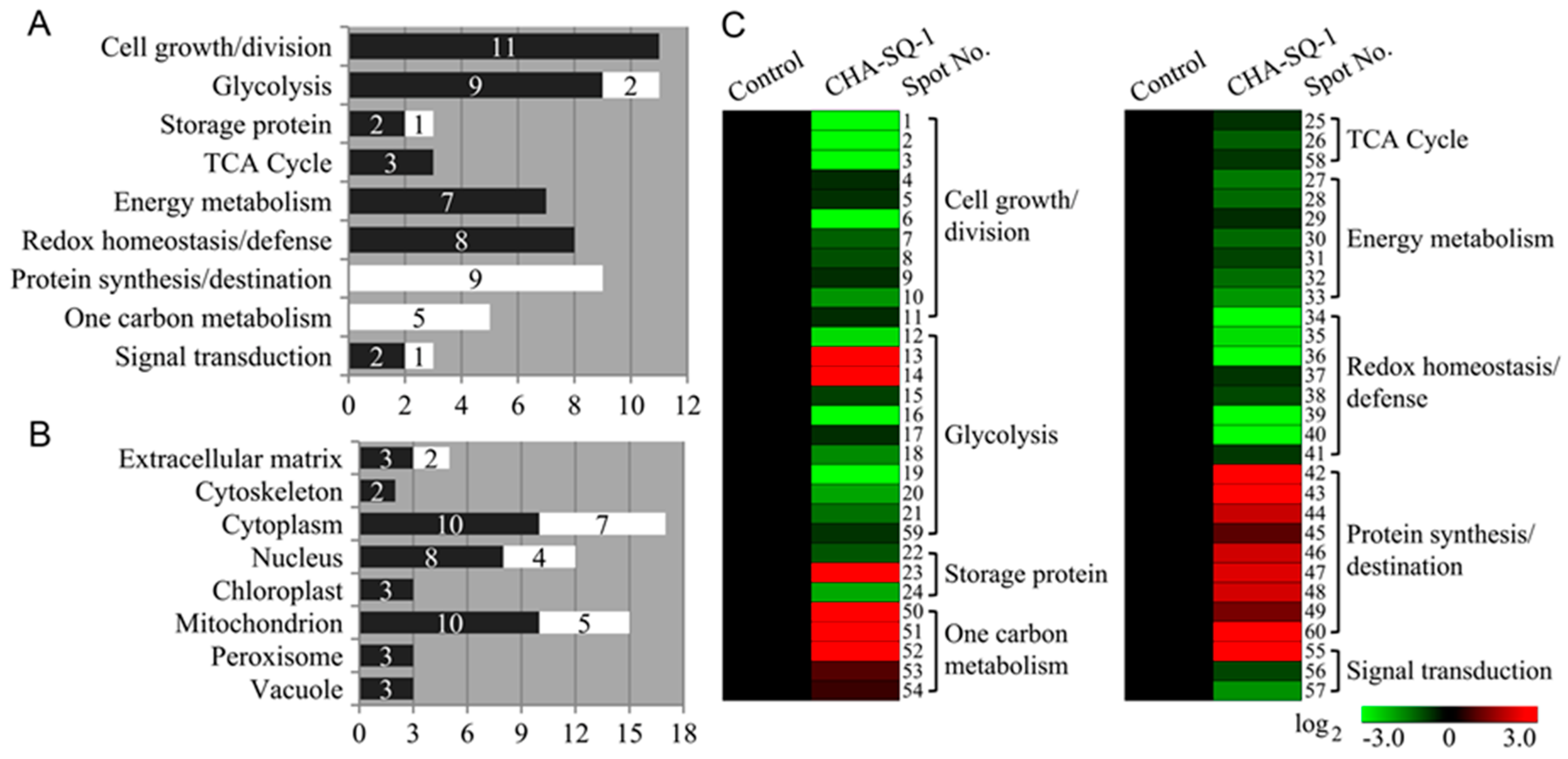
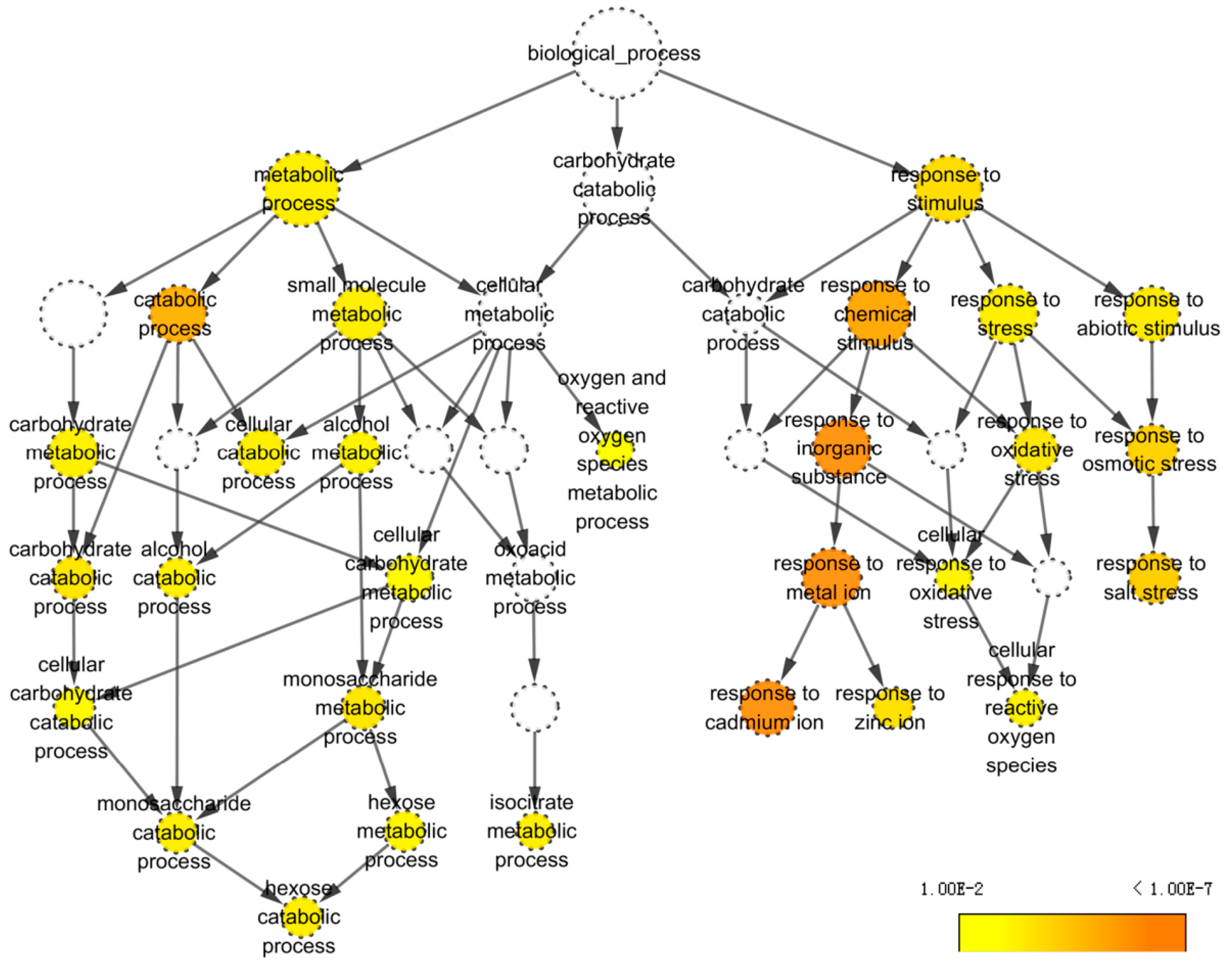
2.3. Identification and Classification of DAPs
2.4. Protein-Protein Interactions Network in CHA-SQ-1-Induced Pollen Abortion
3. Discussion
3.1. CHA-SQ-1 Induced Complete Pollen Abortion
3.2. Proteins Involved in Cell Growth and Division
3.3. Proteins Involved in Carbohydrate and Energy Metabolism
3.4. Stress Response Related Proteins
3.5. Proteins Related to Synthesis, Folding, and Proteolysis
3.6. Other Proteins
4. Materials and Methods
4.1. Plant Material and Treatment
4.2. Sample Fixation and Infiltration
4.3. Light Microscopic Observation
4.4. Electron Microscopic Observation
4.5. Fluorescence Microscopic Observation
4.6. Protein Extraction
4.7. Two-Dimensional Gel Electrophoresis
4.8. Image Analysis
4.9. MS Analysis and Database Search
4.10. Bioinformatic Analysis of Identified Proteins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 2-DE | Two-dimensional gel electrophoresis |
| CBB | Coomassie Brilliant Blue |
| CHA | Chemical hybridizing agent |
| DAPI | 4′, 6-diamidino-2-phenylindole |
| DAPs | Differential abundant proteins |
| DMSO | Dimethyl sulfoxide |
| FDA | Fluorescein diacetate |
| HXK | Hexokinase |
| mtETC | Mitochondrial electron transport chain |
| mtPDC | Mitochondria pyruvate dehydrogenase complex |
| PAS | Periodic acid-Schiff |
| PGK | 3-phosphoglycerate kinase |
| PGM | Phosphoglucomutase |
| pI | Isoelectric point |
| ROS | Reactive oxygen species |
| SPP | Sucrose phosphate phosphatase |
| SPS | Sucrose phosphate synthase |
| TEM | Transmission electron microscopy |
| TIM | Triosephosphat-isomerase |
| UGP | UTP-glucose-1-phosphate uridylyltransferase |
| VIN | Vacuolar invertase |
References
- Shi, J.-X.; Cui, M.-H.; Yang, L.; Kim, Y.J.; Zhang, D.-B. Genetic and biochemical mechanisms of pollen wall development. Trends Plant Sci. 2015, 20, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-P.; Zhang, G.-S.; Song, Q.-L.; Zhang, Y.-X.; Li, Z.; Guo, J.-L.; Niu, N.; Ma, S.-C.; Wang, J.-W. Abnormal development of tapetum and microspores induced by chemical hybridization agent SQ-1 in wheat. PLoS ONE 2015, 10, e0119557. [Google Scholar] [CrossRef]
- Wang, S.-P.; Zhang, G.-S.; Song, Q.-L.; Zhang, Y.-X.; Li, Y.; Chen, Z.; Niu, N.; Ma, S.-C.; Wang, J.-W. Programmed cell death, antioxidant response and oxidative stress in wheat flag leaves induced by chemical hybridization agent SQ-1. J. Integr. Agr. 2016, 15, 76–86. [Google Scholar] [CrossRef]
- Wang, S.-P.; Zhang, Y.-X.; Song, Q.-L.; Fang, Z.-W.; Chen, Z.; Zhang, Y.-M.; Zhang, L.-L.; Zhang, L.; Niu, N.; Ma, S.-C.; et al. Mitochondrial dysfunction causes oxidative stress and tapetal apoptosis in chemical hybridization reagent-induced male sterility in wheat. Front. Plant Sci. 2018, 8, 2217. [Google Scholar] [CrossRef] [PubMed]
- Ba, Q.-S.; Zhang, G.-S.; Wang, J.-S.; Che, H.-X.; Liu, H.-Z.; Niu, N.; Ma, S.-C.; Wang, J.-W. Relationship between metabolism of reactive oxygen species and chemically induced male sterility in wheat (Triticum aestivum L.). Can. J. Plant Sci. 2013, 93, 675–681. [Google Scholar] [CrossRef]
- Ba, Q.-S.; Zhang, G.-S.; Che, H.X.; Liu, H.-Z.; Ng, T.B.; Zhang, L.; Wang, J.-S.; Sheng, Y.; Niu, N.; Ma, S.-C.; et al. Aliphatic metabolism during anther development interfered by chemical hybridizing agent in wheat. Crop Sci. 2014, 54, 1458–1467. [Google Scholar] [CrossRef]
- Ba, Q.-S.; Zhang, G.-S.; Wang, J.-S.; Niu, N.; Ma, S.-C.; Wang, J.-W. Gene expression and DNA methylation alterations in chemically induced male sterility anthers in wheat (Triticum aestivum L.). Acta Physiol. Plant 2014, 36, 503–512. [Google Scholar] [CrossRef]
- Nakamura, Y.; Teo, N.Z.; Shui, G.; Chua, C.H.; Cheong, W.F.; Parameswaran, S.; Koizumi, R.; Ohta, H.; Wenk, M.R.; Ito, T. Transcriptomic and lipidomic profiles of glycerolipids during Arabidopsis flower development. New Phytol. 2014, 203, 310–322. [Google Scholar] [CrossRef]
- Rutley, N.; Twell, D. A decade of pollen transcriptomics. Plant Reprod. 2015, 28, 73–89. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.-D.; Liu, D.; Lv, X.-M.; Wang, Y.; Xun, Z.-L.; Liu, Z.-X.; Li, F.-L.; Lu, H. The cysteine protease CEP1, a key executor involved in tapetal programmed cell death, regulates pollen development in Arabidopsis. Plant Cell 2014, 26, 2939–2961. [Google Scholar] [CrossRef]
- Sumiyoshi, M.; Inamura, T.; Nakamura, A.; Aohara, T.; Ishii, T.; Satoh, S.; Iwai, H. UDP-arabinopyranose mutase 3 is required for pollen wall morphogenesis in rice (Oryza sativa). Plant Cell Physiol. 2015, 56, 232–241. [Google Scholar] [CrossRef]
- Tan, C.; Liu, Z.Y.; Huang, S.N.; Feng, H. Mapping of the male sterile mutant gene ftms in Brassica rapa L. ssp. pekinensis via BSR-Seq combined with whole-genome resequencing. Theor. Appl. Genet. 2019, 132, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Li, L.; Chen, T.; Chong, K.; Xue, Y.; Wang, T. Proteomic analyses of Oryza sativa mature pollen reveal novel proteins associated with pollen germination and tube growth. Proteomics 2006, 6, 2504–2529. [Google Scholar] [CrossRef]
- Sheoran, I.S.; Pedersen, E.J.; Ross, A.R.; Sawhney, V.K. Dynamics of protein expression during pollen germination in canola (Brassica napus). Planta 2009, 230, 779–793. [Google Scholar] [CrossRef]
- Kim, M.H.; Kim, Y.S.; Park, S.K.; Shin, D.I.; Park, H.S.; Chung, I.K. A genotype-specific pollen gene associated with self-incompatibility in Lycopersicon peruvianum. Mol. Cells 2003, 16, 260–265. [Google Scholar] [PubMed]
- Keller, M.; Hu, Y.-J.; Mesihovic, A.; Fragkostefanakis, S.; Schleiff, E.; Simm, S. Alternative splicing in tomato pollen in response to heat stress. DNA Res. 2017, 24, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Jegadeesan, S.; Chaturvedi, P.; Ghatak, A.; Pressman, E.; Meir, S.; Faigenboim, A.; Rutley, N.; Beery, A.; Harel, A.; Weckwerth, W.; et al. Proteomics of heat-stress and ethylene-mediated thermotolerance mechanisms in tomato pollen grains. Front. Plant Sci. 2018, 9, 1558. [Google Scholar] [CrossRef]
- Barton, D.A.; Cantrill, L.C.; Law, A.M.; Phillips, C.G.; Sutton, B.G.; Overall, R.L. Chilling to zero degrees disrupts pollen formation but not meiotic microtubule arrays in Triticum aestivum L. Plant Cell Environ. 2014, 37, 2781–2794. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, S.; Ogasawara, K.; Fujino, K.; Sato, Y.; Kishima, Y. Low temperature-responsive changes in the anther transcriptome’s repeat sequences are indicative of stress sensitivity and pollen sterility in rice strains. Plant Physiol. 2014, 164, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Yang, H.-X.; Wei, Z.; Ma, H.; Ge, X.C. Rice male development under drought stress: Phenotypic changes and stage-dependent transcriptomic reprogramming. Mol. Plant 2013, 6, 1630–1645. [Google Scholar] [CrossRef]
- Koonjul, P.K.; Minhas, J.S.; Nunes, C.; Sheoran, I.S.; Saini, H.S. Selective transcriptional down-regulation of anther invertases precedes the failure of pollen development in water-stressed wheat. J. Exp. Bot. 2005, 56, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Abiko, M.; Akibayashi, K.; Sakata, T.; Kimura, M.; Kihara, M.; Itoh, K.; Asamizu, E.; Sato, S.; Takahashi, H.; Higashitani, A. High-temperature induction of male sterility during barley (Hordeum vulgare L.) anther development is mediated by transcriptional inhibition. Sex. Plant Reprod. 2005, 18, 91–100. [Google Scholar] [CrossRef]
- Liao, J.-L.; Zhou, H.-W.; Zhang, H.-Y.; Zhong, P.-A.; Huang, Y.-J. Comparative proteomic analysis of differentially expressed proteins in the early milky stage of rice grains during high temperature stress. J. Exp. Bot. 2014, 65, 655–671. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Y.; Qiu, L.; Song, Q.-L.; Wang, S.-P.; Wang, Y.-J.; Ge, Y.-H. Root proteomics reveals the effects of wood vinegar on wheat growth and subsequent tolerance to drought stress. Int. J. Mol. Sci. 2019, 20, 942. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Lau, V.; Song, R.; Ierullo, M.; Esteban, E.; Wu, Y.; Sivieng, T.; Nahal-Bose, H.K.; Gaudinier, A.; Pasha, A.; et al. Proteome-wide, structure-based prediction of protein-protein interactions/new molecular interactions viewer. Plant Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Fotovat, R.; Alikhani, M.; Valizadeh, M.; Mirzaei, M.; Salekdeh, G.H. A proteomics approach to discover drought tolerance proteins in wheat pollen grain at meiosis stage. Protein Peptide Lett. 2017, 24, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Krishnan, P.; Nayak, M.; Ramakrishnan, B. High temperature stress effects on pollens of rice (Oryza sativa L.) genotypes. Environ. Exp. Bot. 2014, 101, 36–46. [Google Scholar] [CrossRef]
- Zhu, W.; Ma, S.; Zhang, G.; Liu, H.; Ba, Q.; Li, Z.; Song, Y.; Zhang, P.; Niu, N.; Wang, J. Carbohydrate metabolism and gene regulation during anther development disturbed by chemical hybridizing agent in wheat. Crop Sci. 2015, 55, 868–876. [Google Scholar] [CrossRef]
- Zhu, Q.-D.; Song, Y.-L.; Zhang, G.-S.; Ju, L.; Zhang, J.-G.; Yu, Y.-G.; Niu, N.; Wang, J.-W.; Ma, S.-C. De Novo assembly and transcriptome analysis of wheat with male sterility induced by the chemical hybridizing agent SQ-1. PLoS ONE 2015, 10, e0123556. [Google Scholar] [CrossRef]
- Song, Q.-L.; Wang, S.-P.; Zhang, G.-S.; Li, Y.; Li, Z.; Guo, J.-L.; Niu, N.; Wang, J.-W.; Ma, S.-C. Comparative proteomic analysis of a membrane-enriched fraction from flag leaves reveals responses to chemical hybridization agent SQ-1 in wheat. Front. Plant Sci. 2015, 6, 669. [Google Scholar] [CrossRef]
- Liu, H.-Z.; Zhang, G.-S.; Wang, J.-S.; Li, J.-J.; Song, Y.-L.; Qiao, L.; Niu, N.; Wang, J.-W.; Ma, S.-C.; Li, L.-L. Chemical hybridizing agent SQ-1-induced male sterility in Triticum aestivum L.: A comparative analysis of the anther proteome. BMC Plant Biol. 2018, 18, 7. [Google Scholar] [CrossRef]
- Wang, S.-P.; Zhang, G.-S.; Zhang, Y.-X.; Song, Q.-L.; Chen, Z.; Wang, J.-S.; Guo, J.-L.; Niu, N.; Wang, J.-W.; Ma, S.-C. Comparative studies of mitochondrial proteomics reveal an intimate protein network of male sterility in wheat (Triticum aestivum L.). J. Exp. Bot. 2015, 66, 6191–6203. [Google Scholar] [CrossRef]
- Peng, Z.; Wang, M.; Li, F.; Lv, H.; Li, C.; Xia, G. A proteomic study of the response to salinity and drought stress in an introgression strain of bread wheat. Mol. Cell Proteom. 2009, 8, 2676–2686. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.-L.; Wang, J.-W.; Zhang, G.-S.; Zhao, X.-L.; Zhang, P.-F.; Niu, N.; Ma, S.-C. Isolation and characterization of a wheat F8-1 homolog required for physiological male sterility induced by a chemical hybridizing agent (CHA) SQ-1. Euphytica 2015, 205, 707–720. [Google Scholar] [CrossRef]
- Dai, S.; Wang, T.; Yan, X.; Chen, S. Proteomics of pollen development and germination. J. Proteome Res. 2007, 6, 4556–4563. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-P.; Nayak, S.; Koch, K.; Ming, R. Carbon partitioning in sugarcane (Saccharum species). Front. Plant Sci. 2013, 4, 201. [Google Scholar] [CrossRef] [PubMed]
- Oliver, S.N.; Dennis, E.S.; Rudy, D. ABA regulates apoplastic sugar transport and is a potential signal for cold-induced pollen sterility in rice. Plant Cell Physiol. 2007, 48, 1319–1330. [Google Scholar] [CrossRef]
- Tiessen, A.; Padilla-Chacon, D. Subcellular compartmentation of sugar signaling: Links among carbon cellular status, route of sucrolysis, sink-source allocation, and metabolic partitioning. Front. Plant Sci. 2013, 3, 306. [Google Scholar] [CrossRef]
- Wierenga, R.-K.; Kapetaniou, E.-G.; Venkatesan, R. Triosephosphate isomerase: A highly evolved biocatalyst. Cell. Mol. Life Sci. 2010, 67, 3961–3982. [Google Scholar] [CrossRef]
- Oliver, S.N.; Dongen, J.T.V.; Alfred, S.C.; Mamun, E.A.; Zhao, X.C.; Saini, H.S.; Fernandes, S.F.; Blanchard, C.L.; Sutton, B.G.; Geigenberger, P.; et al. Cold-induced repression of the rice anther-specific cell wall invertase gene OSINV4 is correlated with sucrose accumulation and pollen sterility. Plant Cell Environ. 2010, 28, 1534–1551. [Google Scholar] [CrossRef]
- Zhu, X.-L.; Liang, W.-Q.; Cui, X.; Chen, M.-J.; Yin, C.-S.; Luo, Z.-J.; Zhu, J.-Y.; Lucas, W.J.; Wang, Z.-Y.; Zhang, D.-B. Brassinosteroids promote development of rice pollen grains and seeds by triggering expression of Carbon Starved Anther, a MYB domain protein. Plant J. 2015, 82, 570–581. [Google Scholar] [CrossRef]
- Signorelli, S.; Tarkowski, L.P.; Van den Ende, W.; Bassham, D.C. Linking autophagy to abiotic and biotic stress responses. Trends Plant Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, P.; Doerfler, H.; Jegadeesan, S.; Ghatak, A.; Pressman, E.; Castillejo, M.A.; Wienkoop, S.; Egelhofer, V.; Firon, N.; Weckwerth, W. Heat-Treatment-Responsive proteins in different developmental stages of tomato pollen detected by targeted mass accuracy precursor alignment (tMAPA). J. Proteome Res. 2015, 14, 4463–4471. [Google Scholar] [CrossRef]
- Chaturvedi, P.; Ghatak, A.; Weckwerth, W. Pollen proteomics: From stress physiology to developmental priming. Plant Reprod. 2016, 29, 119–132. [Google Scholar] [CrossRef]
- Kuntumalla, S.; Braisted, J.C.; Huang, S.T.; Parmar, P.P.; Clark, D.J.; Alami, H.; Zhang, Q.-S.; Donohue-Rolfe, A.; Tzipori, S.; Fleischmann, R.D.; et al. Comparison of two label-free global quantitation methods, APEX and 2D gel electrophoresis, applied to the Shigella dysenteriae proteome. Proteome Sci. 2009, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-Q.; Zhang, X.-L.; Zhang, J.; Fan, W.; Lu, M.-Z.; Hu, J.-J. Proteomic analysis and identification of possible allergenic proteins in mature pollen of Populus tomentosa. Int. J. Mol. Sci. 2018, 19, 250. [Google Scholar] [CrossRef]
- Konyar, S.T.; Dane, F. Cytochemistry of pollen development in Campsis radicans (L.) Seem. (Bignoniaceae). Plant Syst. Evol. 2013, 299, 87–95. [Google Scholar] [CrossRef]
- Tian, Q.-Q.; Lu, C.M.; Li, X.; Fang, X.-W. Low temperature treatments of rice (Oryza sativa L.) anthers changes polysaccharide and protein composition of the anther walls and increases pollen fertility and callus induction. Plant Cell Tissue Organ Culture 2015, 120, 89–98. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, L.-S.; Fan, W.; Zhang, X.-L.; Jia, H.-X.; Li, Y.; Yin, Y.-F.; Hu, J.-J.; Lu, M.-Z. Proteomic analysis and candidate allergenic proteins in Populus deltoides CL. “2KEN8” mature pollen. Front. Plant Sci. 2015, 6, 548. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Zhang, Y.; Fang, Z.; Zhang, Y.; Song, Q.; Hou, Z.; Sun, K.; Song, Y.; Li, Y.; Ma, D.; et al. Cytological and Proteomic Analysis of Wheat Pollen Abortion Induced by Chemical Hybridization Agent. Int. J. Mol. Sci. 2019, 20, 1615. https://doi.org/10.3390/ijms20071615
Wang S, Zhang Y, Fang Z, Zhang Y, Song Q, Hou Z, Sun K, Song Y, Li Y, Ma D, et al. Cytological and Proteomic Analysis of Wheat Pollen Abortion Induced by Chemical Hybridization Agent. International Journal of Molecular Sciences. 2019; 20(7):1615. https://doi.org/10.3390/ijms20071615
Chicago/Turabian StyleWang, Shuping, Yingxin Zhang, Zhengwu Fang, Yamin Zhang, Qilu Song, Zehao Hou, Kunkun Sun, Yulong Song, Ying Li, Dongfang Ma, and et al. 2019. "Cytological and Proteomic Analysis of Wheat Pollen Abortion Induced by Chemical Hybridization Agent" International Journal of Molecular Sciences 20, no. 7: 1615. https://doi.org/10.3390/ijms20071615
APA StyleWang, S., Zhang, Y., Fang, Z., Zhang, Y., Song, Q., Hou, Z., Sun, K., Song, Y., Li, Y., Ma, D., Liu, Y., Zhu, Z., Niu, N., Wang, J., Ma, S., & Zhang, G. (2019). Cytological and Proteomic Analysis of Wheat Pollen Abortion Induced by Chemical Hybridization Agent. International Journal of Molecular Sciences, 20(7), 1615. https://doi.org/10.3390/ijms20071615