The Potential Role of iNOS in Ovarian Cancer Progression and Chemoresistance


Abstract
:1. Introduction
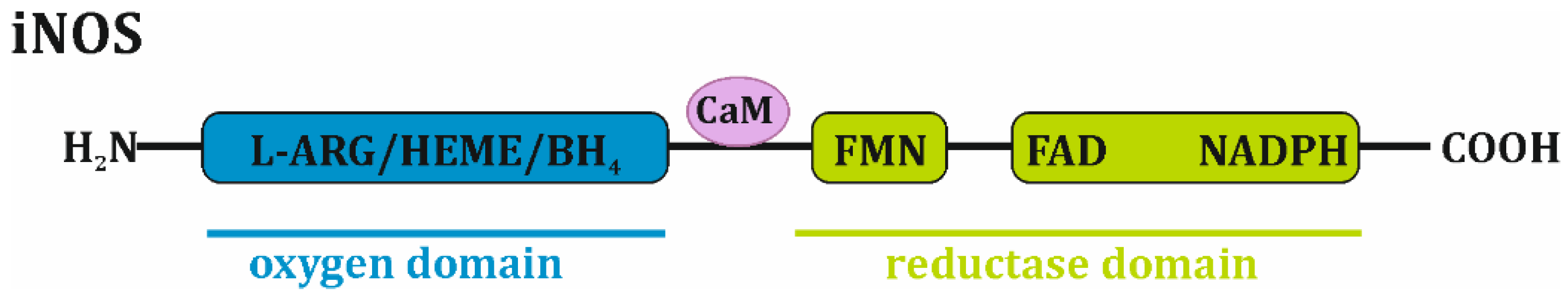
2. iNOS—Structure, Enzymatic Activity and Regulation in Normal Cells
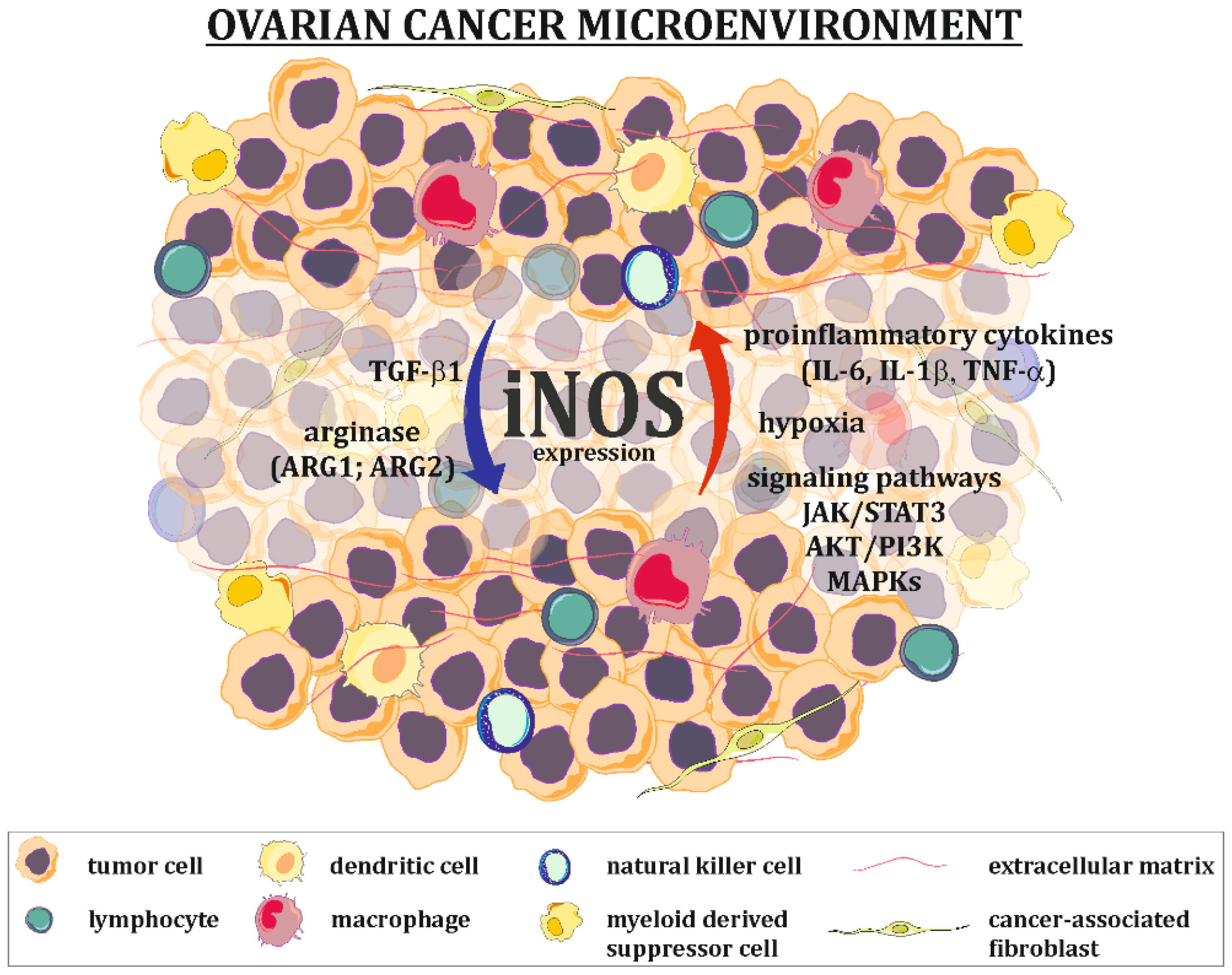
3. iNOS Expression and Regulation in Ovarian Tumors
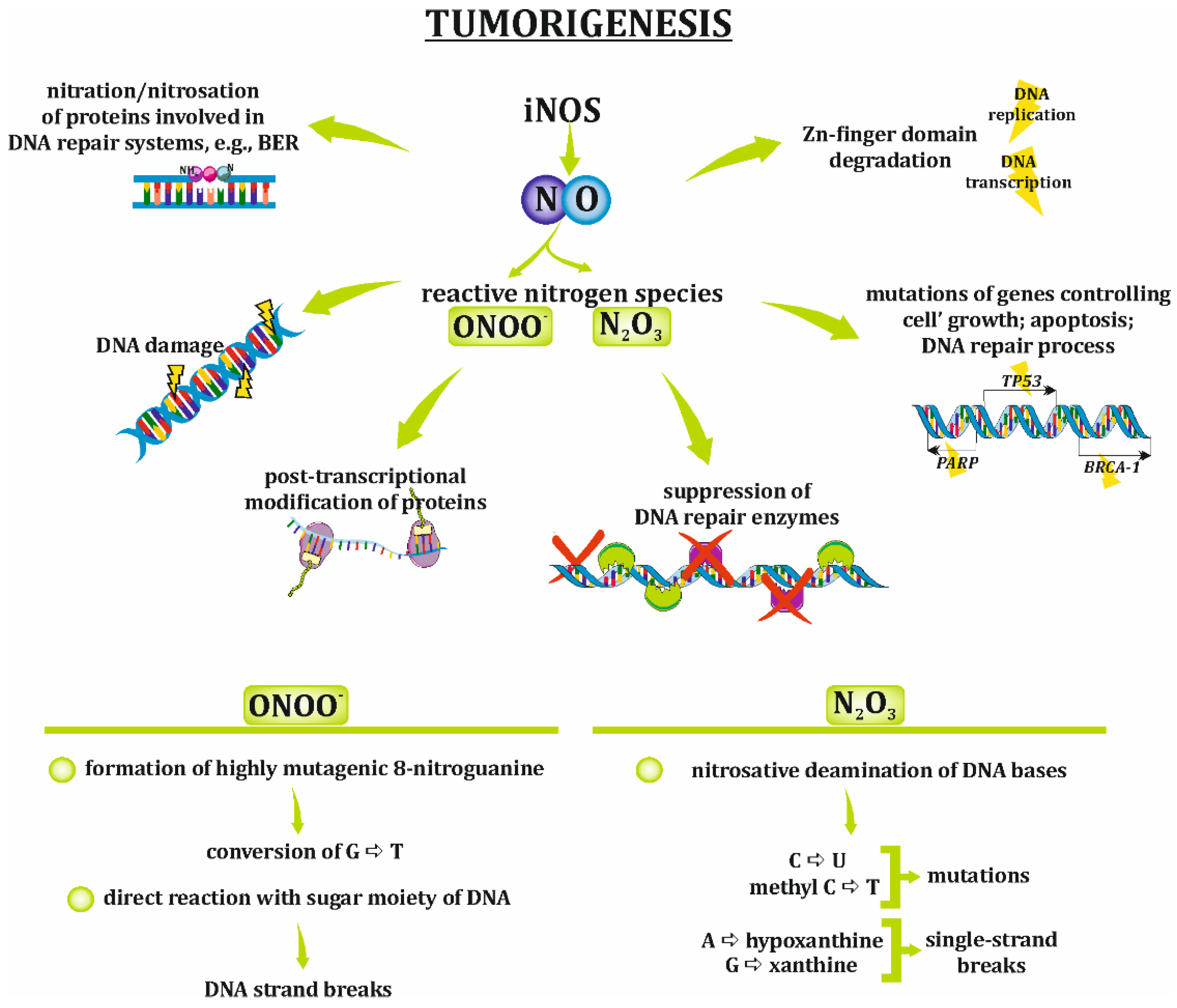
4. Implication of NO/RNS in the Development of Ovarian Cancer
5. Can iNOS Expression be a Prognostic Factor in Ovarian Cancer?
5.1. Votes For
5.2. Votes Against
5.3. NO Correct Answer
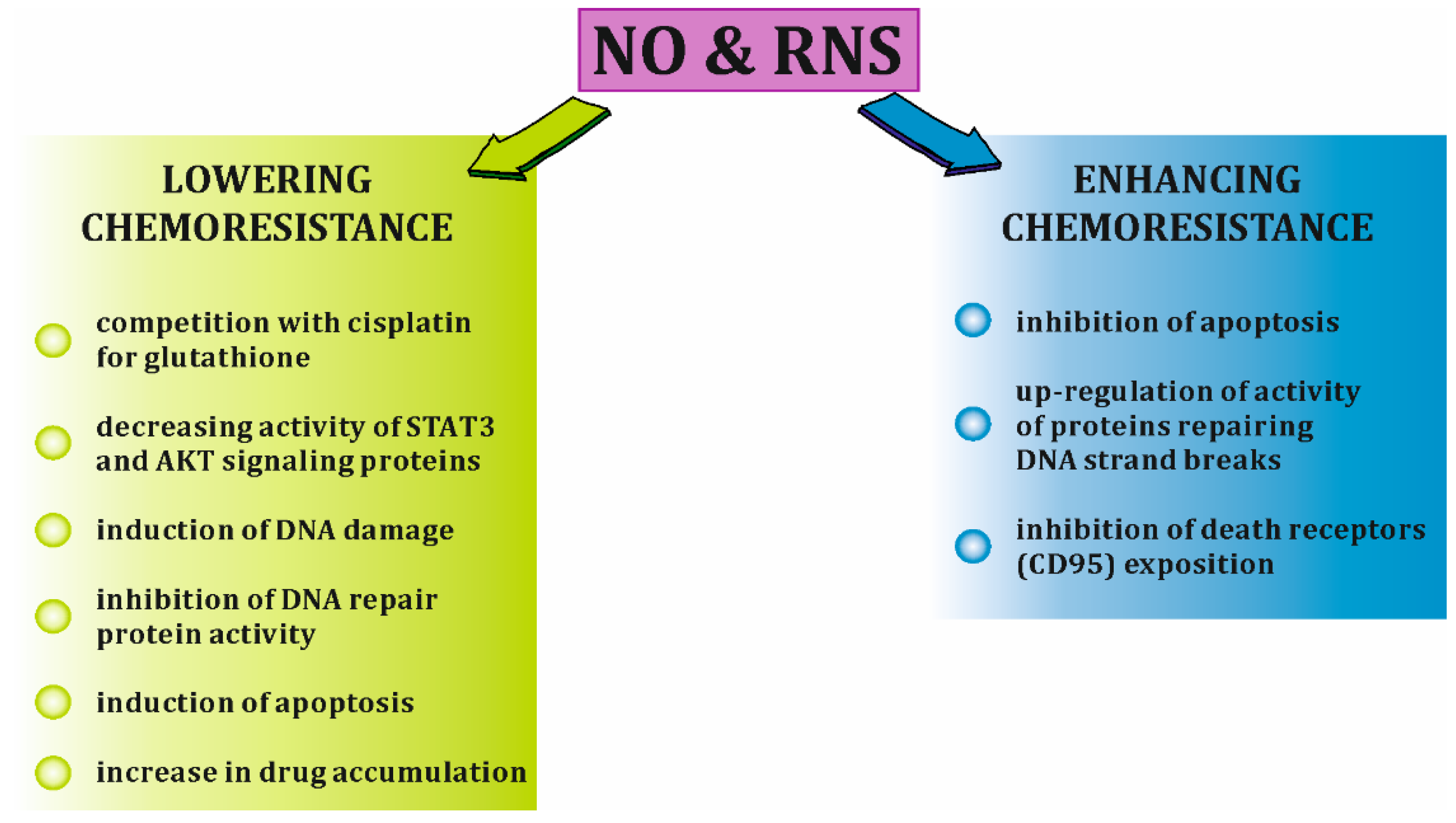
6. iNOS Expression Versus Chemoresistance in Ovarian Cancer Cells
7. Targeting of iNOS in Ovarian Cancer
8. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ·O2- | superoxide anion |
| 3′-UTR | 3′-untranslated region |
| AKT | serine-threonine protein kinase |
| ARE | AU-rich element |
| BH4 | tetrahydropterin |
| FAD | flavin adenine dinucleotide |
| FIGO | International Federation of Gynecology and Obstetrics |
| FMN | flavin mononucleotide |
| HIF | hypoxia-inducible factor |
| IFN-γ | interferon-γ |
| IL | interleukin |
| iNOS | inducible nitric oxide synthase |
| JAK | Janus tyrosine kinase |
| MAPK | mitogen-activated protein kinase |
| miRNA | microRNA |
| NADPH | reduced form of nicotinamide adenine dinucleotide phosphate |
| NO | nitric oxide |
| ONOO- | peroxynitrite anion |
| RNS | reactive nitrogen species |
| STAT | signal transducer and activator of transcription |
| TGF-β1 | transforming growth factor-β1 |
| TNF-α | tumor necrosis factor-α |
References
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric oxide synthases: Structure, function and inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef] [PubMed]
- Groves, J.T.; Wang, C.C. Nitric oxide synthase: Models and mechanisms. Curr. Opin. Chem. Biol. 2000, 4, 687–695. [Google Scholar] [CrossRef]
- Kröncke, K.D.; Fehsel, K.; Kolb-Bachofen, V. Inducible nitric oxide synthase and its product nitric oxide, a small molecule with complex biological activities. Biol. Chem. Hoppe Seyler 1995, 376, 327–343. [Google Scholar] [CrossRef]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Pautz, A.; Art, J.; Hahn, S.; Nowag, S.; Voss, C.; Kleinert, H. Regulation of the expression of inducible nitric oxide synthase. In Nitric Oxide; Academic Press: Cambridge, MA, USA, 2010; Volume 23, pp. 75–93. [Google Scholar]
- Keshet, R.; Erez, A. Arginine and the metabolic regulation of nitric oxide synthesis in cancer. Dis. Model. Mech. 2018, 11, dmm033332. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y. Superoxide generation from nitric oxide synthases. Antioxid. Redox Signal. 2007, 9, 1773–1778. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide synthase in innate and adaptive immunity: An update. Trends Immunol. 2015, 36, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Geller, D.A. microRNA and human inducible nitric oxide synthase. In Vitamins & Hormones; Academic Press: Cambridge, MA, USA, 2014; Volume 96, pp. 19–27. [Google Scholar]
- Lirk, P.; Hoffmann, G.; Rieder, J. Inducible nitric oxide synthase--time for reappraisal. Curr. Drug Targets Inflamm. Allergy 2002, 1, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Kleinert, H.; Pautz, A.; Linker, K.; Schwarz, P.M. Regulation of the expression of inducible nitric oxide synthase. Eur. J. Pharmacol. 2004, 500, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Kleinert, H.; Schwarz, P.M.; Förstermann, U. Regulation of the expression of inducible nitric oxide synthase. Biol. Chem. 2003, 384, 1343–1364. [Google Scholar] [CrossRef] [PubMed]
- Lowenstein, C.J.; Padalko, E. iNOS (NOS2) at a glance. J. Cell Sci. 2004, 117, 2865–2867. [Google Scholar] [CrossRef] [PubMed]
- Pautz, A.; Linker, K.; Altenhöfer, S.; Heil, S.; Schmidt, N.; Art, J.; Knauer, S.; Stauber, R.; Sadri, N.; Pont, A.; et al. Similar regulation of human inducible nitric-oxide synthase expression by different isoforms of the RNA-binding protein AUF1. J. Biol. Chem. 2009, 284, 2755–2766. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Pascual, F.; Hausding, M.; Ihrig-Biedert, I.; Furneaux, H.; Levy, A.P.; Förstermann, U.; Kleinert, H. Complex contribution of the 3′-untranslated region to the expressional regulation of the human inducible nitric-oxide synthase gene. Involvement of the RNA-binding protein HuR. J. Biol. Chem. 2000, 275, 26040–26049. [Google Scholar] [CrossRef]
- Linker, K.; Pautz, A.; Fechir, M.; Hubrich, T.; Greeve, J.; Kleinert, H. Involvement of KSRP in the post-transcriptional regulation of human iNOS expression-complex interplay of KSRP with TTP and HuR. Nucleic Acids Res. 2005, 33, 4813–4827. [Google Scholar] [CrossRef]
- Guo, Z.; Shao, L.; Zheng, L.; Du, Q.; Li, P.; John, B.; Geller, D.A. miRNA-939 regulates human inducible nitric oxide synthase posttranscriptional gene expression in human hepatocytes. Proc. Natl. Acad. Sci. USA 2012, 109, 5826–5831. [Google Scholar] [CrossRef]
- Li, X.; Gibson, G.; Kim, J.-S.; Kroin, J.; Xu, S.; van Wijnen, A.J.; Im, H.-J. MicroRNA-146a is linked to pain-related pathophysiology of osteoarthritis. Gene 2011, 480, 34–41. [Google Scholar] [CrossRef]
- Zhu, H.; Vishwamitra, D.; Curry, C.V.; Manshouri, R.; Diao, L.; Khan, A.; Amin, H.M. NPM-ALK up-regulates iNOS expression through a STAT3/microRNA-26a-dependent mechanism. J. Pathol. 2013, 230, 82–94. [Google Scholar] [CrossRef]
- Luss, H.; Li, R.K.; Shapiro, R.A.; Tzeng, E.; McGowan, F.X.; Yoneyama, T.; Hatakeyama, K.; Geller, D.A.; Mickle, D.A.; Simmons, R.L.; et al. Dedifferentiated human ventricular cardiac myocytes express inducible nitric oxide synthase mRNA but not protein in response to IL-1, TNF, IFNgamma, and LPS. J. Mol. Cell. Cardiol. 1997, 29, 1153–1165. [Google Scholar] [CrossRef] [PubMed]
- Felley-Bosco, E.; Bender, F.; Quest, A.F.G. Caveolin-1-mediated post-transcriptional regulation of inducible nitric oxide synthase in human colon carcinoma cells. Biol. Res. 2002, 35, 169–176. [Google Scholar] [CrossRef]
- Choudhari, S.K.; Chaudhary, M.; Bagde, S.; Gadbail, A.R.; Joshi, V. Nitric oxide and cancer: A review. World J. Surg. Oncol. 2013, 11, 118. [Google Scholar] [CrossRef]
- Bian, K.; Ghassemi, F.; Sotolongo, A.; Siu, A.; Shauger, L.; Kots, A.; Murad, F. NOS-2 signaling and cancer therapy. IUBMB Life 2012, 64, 676–683. [Google Scholar] [CrossRef]
- Lechner, M.; Lirk, P.; Rieder, J. Inducible nitric oxide synthase (iNOS) in tumor biology: The two sides of the same coin. Semin. Cancer Biol. 2005, 15, 277–289. [Google Scholar] [CrossRef]
- Burke, A.J.; Sullivan, F.J.; Giles, F.J.; Glynn, S.A. The yin and yang of nitric oxide in cancer progression. Carcinogenesis 2013, 34, 503–512. [Google Scholar] [CrossRef]
- Burke, A.J.; Garrido, P.; Johnson, C.; Sullivan, F.J.; Glynn, S.A. Inflammation and Nitrosative Stress Effects in Ovarian and Prostate Pathology and Carcinogenesis. Antioxid. Redox Signal. 2017, 26, 1078–1090. [Google Scholar] [CrossRef]
- Nomelini, R.S.; de Abreu Ribeiro, L.C.; Tavares-Murta, B.M.; Adad, S.J.; Murta, E.F.C. Production of nitric oxide and expression of inducible nitric oxide synthase in ovarian cystic tumors. Mediat. Inflamm. 2008, 2008, 186584. [Google Scholar] [CrossRef]
- Klimp, A.H.; Hollema, H.; Kempinga, C.; van der Zee, A.G.; de Vries, E.G.; Daemen, T. Expression of cyclooxygenase-2 and inducible nitric oxide synthase in human ovarian tumors and tumor-associated macrophages. Cancer Res. 2001, 61, 7305–7309. [Google Scholar]
- Salimian Rizi, B.; Caneba, C.; Nowicka, A.; Nabiyar, A.W.; Liu, X.; Chen, K.; Klopp, A.; Nagrath, D. Nitric oxide mediates metabolic coupling of omentum-derived adipose stroma to ovarian and endometrial cancer cells. Cancer Res. 2015, 75, 456–471. [Google Scholar] [CrossRef]
- El-Sehemy, A.; Postovit, L.-M.; Fu, Y. Nitric oxide signaling in human ovarian cancer: A potential therapeutic target. Nitric Oxide Biol. Chem. 2016, 54, 30–37. [Google Scholar] [CrossRef]
- Macciò, A.; Madeddu, C. Inflammation and ovarian cancer. Cytokine 2012, 58, 133–147. [Google Scholar] [CrossRef]
- Kulbe, H.; Chakravarty, P.; Leinster, D.A.; Charles, K.A.; Kwong, J.; Thompson, R.G.; Coward, J.I.; Schioppa, T.; Robinson, S.C.; Gallagher, W.M.; et al. A dynamic inflammatory cytokine network in the human ovarian cancer microenvironment. Cancer Res. 2012, 72, 66–75. [Google Scholar] [CrossRef]
- Yu, H.; Kortylewski, M.; Pardoll, D. Crosstalk between cancer and immune cells: Role of STAT3 in the tumour microenvironment. Nat. Rev. Immunol. 2007, 7, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Paez, J.; Sellers, W.R. PI3K/PTEN/AKT pathway. A critical mediator of oncogenic signaling. Cancer Treat Res. 2003, 115, 145–167. [Google Scholar] [PubMed]
- Burotto, M.; Chiou, V.L.; Lee, J.-M.; Kohn, E.C. The MAPK pathway across different malignancies: A new perspective. Cancer 2014, 120, 3446–3456. [Google Scholar] [CrossRef]
- McCubrey, J.A.; Steelman, L.S.; Chappell, W.H.; Abrams, S.L.; Wong, E.W.T.; Chang, F.; Lehmann, B.; Terrian, D.M.; Milella, M.; Tafuri, A.; et al. Roles of the Raf/MEK/ERK pathway in cell growth, malignant transformation and drug resistance. Biochim. Biophys. Acta 2007, 1773, 1263–1284. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.A.; Baumgardner, J.E.; Otto, C.M. Oxygen-dependent regulation of nitric oxide production by inducible nitric oxide synthase. Free Radic. Biol. Med. 2011, 51, 1952–1965. [Google Scholar] [CrossRef]
- Timosenko, E.; Hadjinicolaou, A.V.; Cerundolo, V. Modulation of cancer-specific immune responses by amino acid degrading enzymes. Immunotherapy 2017, 9, 83–97. [Google Scholar] [CrossRef]
- Molinier-Frenkel, V.; Castellano, F. Immunosuppressive enzymes in the tumor microenvironment. FEBS Lett. 2017, 591, 3135–3157. [Google Scholar] [CrossRef]
- Abendstein, B.; Stadlmann, S.; Knabbe, C.; Buck, M.; Müller-Holzner, E.; Zeimet, A.G.; Marth, C.; Obrist, P.; Krugmann, J.; Offner, F.A. Regulation of transforming growth factor-beta secretion by human peritoneal mesothelial and ovarian carcinoma cells. Cytokine 2000, 12, 1115–1119. [Google Scholar] [CrossRef]
- Toutirais, O.; Chartier, P.; Dubois, D.; Bouet, F.; Lévêque, J.; Catros-Quemener, V.; Genetet, N. Constitutive expression of TGF-bêta1, interleukin-6 and interleukin-8 by tumor cells as a major component of immune escape in human ovarian carcinoma. Eur. Cytokine Netw. 2003, 14, 246–255. [Google Scholar]
- Feng, X.; Wang, C.-X.; Ou, Z.-Y. TGF-β1 and IL-10 expression in epithelial ovarian cancer cell line A2780. Trop. J. Pharm. Res. 2015, 14, 2179–2185. [Google Scholar] [CrossRef]
- Berg, D.T.; Gupta, A.; Richardson, M.A.; O’Brien, L.A.; Calnek, D.; Grinnell, B.W. Negative regulation of inducible nitric-oxide synthase expression mediated through transforming growth factor-beta-dependent modulation of transcription factor TCF11. J. Biol. Chem. 2007, 282, 36837–36844. [Google Scholar] [CrossRef]
- Saad, A.F.; Hu, W.; Sood, A.K. Microenvironment and pathogenesis of epithelial ovarian cancer. Horm. Cancer 2010, 1, 277–290. [Google Scholar] [CrossRef] [PubMed]
- King, M.-C.; Marks, J.H.; Mandell, J.B. New York Breast Cancer Study Group Breast and ovarian cancer risks due to inherited mutations in BRCA1 and BRCA2. Science 2003, 302, 643–646. [Google Scholar] [CrossRef]
- Crijnen, T.E.M.; Janssen-Heijnen, M.L.G.; Gelderblom, H.; Morreau, J.; Nooij, M.A.; Kenter, G.G.; Vasen, H.F.A. Survival of patients with ovarian cancer due to a mismatch repair defect. Fam. Cancer 2005, 4, 301–305. [Google Scholar] [CrossRef]
- Singer, G.; Oldt, R.; Cohen, Y.; Wang, B.G.; Sidransky, D.; Kurman, R.J.; Shih, I.-M. Mutations in BRAF and KRAS characterize the development of low-grade ovarian serous carcinoma. J. Natl. Cancer Inst. 2003, 95, 484–486. [Google Scholar] [CrossRef]
- Zhang, D.; Piao, H.-L.; Li, Y.-H.; Qiu, Q.; Li, D.-J.; Du, M.-R.; Tsang, B.K. Inhibition of AKT sensitizes chemoresistant ovarian cancer cells to cisplatin by abrogating S and G2/M arrest. Exp. Mol. Pathol. 2016, 100, 506–513. [Google Scholar] [CrossRef]
- Suzuki, N.; Yasui, M.; Geacintov, N.E.; Shafirovich, V.; Shibutani, S. Miscoding events during DNA synthesis past the nitration-damaged base 8-nitroguanine. Biochemistry 2005, 44, 9238–9245. [Google Scholar] [CrossRef]
- Burney, S.; Caulfield, J.L.; Niles, J.C.; Wishnok, J.S.; Tannenbaum, S.R. The chemistry of DNA damage from nitric oxide and peroxynitrite. Mutat. Res. 1999, 424, 37–49. [Google Scholar] [CrossRef]
- Grisham, M.B.; Jourd’heuil, D.; Wink, D.A. Review article: Chronic inflammation and reactive oxygen and nitrogen metabolism--implications in DNA damage and mutagenesis. Aliment. Pharmacol. Ther. 2000, 14 (Suppl. 1), 3–9. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.E.; Ying, L.; Hofseth, A.B.; Jelezcova, E.; Sobol, R.W.; Ambs, S.; Harris, C.C.; Espey, M.G.; Hofseth, L.J.; Wyatt, M.D. Differential effects of reactive nitrogen species on DNA base excision repair initiated by the alkyladenine DNA glycosylase. Carcinogenesis 2009, 30, 2123–2129. [Google Scholar] [CrossRef] [PubMed]
- Parrish, M.C.; Chaim, I.A.; Nagel, Z.D.; Tannenbaum, S.R.; Samson, L.D.; Engelward, B.P. Nitric oxide induced S-nitrosation causes base excision repair imbalance. DNA Repair 2018, 68, 25–33. [Google Scholar] [CrossRef]
- Chang, L.-C.; Huang, C.-F.; Lai, M.-S.; Shen, L.-J.; Wu, F.-L.L.; Cheng, W.-F. Prognostic factors in epithelial ovarian cancer: A population-based study. PLoS ONE 2018, 13, e0194993. [Google Scholar] [CrossRef]
- Davidson, B.; Tropé, C.G. Ovarian cancer: Diagnostic, biological and prognostic aspects. Womens Health 2014, 10, 519–533. [Google Scholar] [CrossRef]
- Ezzati, M.; Abdullah, A.; Shariftabrizi, A.; Hou, J.; Kopf, M.; Stedman, J.K.; Samuelson, R.; Shahabi, S. Recent Advancements in Prognostic Factors of Epithelial Ovarian Carcinoma. Int. Sch. Res. Not. 2014, 2014, 953509. [Google Scholar] [CrossRef]
- Xu, K.; Yang, S.; Zhao, Y. Prognostic significance of BRCA mutations in ovarian cancer: An updated systematic review with meta-analysis. Oncotarget 2017, 8, 285–302. [Google Scholar] [CrossRef]
- Woopen, H.; Pietzner, K.; Richter, R.; Fotopoulou, C.; Joens, T.; Braicu, E.I.; Mellstedt, H.; Mahner, S.; Lindhofer, H.; Darb-Esfahani, S.; et al. Overexpression of the epithelial cell adhesion molecule is associated with a more favorable prognosis and response to platinum-based chemotherapy in ovarian cancer. J. Gynecol. Oncol. 2014, 25, 221–228. [Google Scholar] [CrossRef]
- Shen, W.; Li, H.-L.; Liu, L.; Cheng, J.-X. Expression levels of PTEN, HIF-1α, and VEGF as prognostic factors in ovarian cancer. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2596–2603. [Google Scholar]
- Raspollini, M.R.; Amunni, G.; Villanucci, A.; Boddi, V.; Baroni, G.; Taddei, A.; Taddei, G.L. Expression of inducible nitric oxide synthase and cyclooxygenase-2 in ovarian cancer: Correlation with clinical outcome. Gynecol. Oncol. 2004, 92, 806–812. [Google Scholar] [CrossRef]
- Ali-Fehmi, R.; Semaan, A.; Sethi, S.; Arabi, H.; Bandyopadhyay, S.; Hussein, Y.R.; Diamond, M.P.; Saed, G.; Morris, R.T.; Munkarah, A.R. Molecular typing of epithelial ovarian carcinomas using inflammatory markers. Cancer 2011, 117, 301–309. [Google Scholar] [CrossRef]
- Engels, K.; du Bois, A.; Harter, P.; Fisseler-Eckhoff, A.; Kommoss, F.; Stauber, R.; Kaufmann, M.; Nekljudova, V.; Loibl, S. VEGF-A and i-NOS expression are prognostic factors in serous epithelial ovarian carcinomas after complete surgical resection. J. Clin. Pathol. 2009, 62, 448–454. [Google Scholar] [CrossRef]
- Saied, E.M.; El-Etreby, N.M. The role and prognostic value of inducible nitric oxide synthase (iNOS) and interleukin-33 (IL-33) in serous and mucinous epithelial ovarian tumours. Ann. Diagn. Pathol. 2017, 27, 62–68. [Google Scholar] [CrossRef]
- Li, L.; Zhu, L.; Hao, B.; Gao, W.; Wang, Q.; Li, K.; Wang, M.; Huang, M.; Liu, Z.; Yang, Q.; et al. iNOS-derived nitric oxide promotes glycolysis by inducing pyruvate kinase M2 nuclear translocation in ovarian cancer. Oncotarget 2017, 8, 33047–33063. [Google Scholar] [CrossRef]
- Anttila, M.A.; Voutilainen, K.; Merivalo, S.; Saarikoski, S.; Kosma, V.-M. Prognostic significance of iNOS in epithelial ovarian cancer. Gynecol. Oncol. 2007, 105, 97–103. [Google Scholar] [CrossRef]
- Ozel, E.; Peştereli, H.E.; Simşek, T.; Erdoğan, G.; Karaveli, F.S. Expression of cyclooxygenase-2 and inducible nitric oxide synthase in ovarian surface epithelial carcinomas: Is there any correlation with angiogenesis or clinicopathologic parameters? Int. J. Gynecol. Cancer 2006, 16, 549–555. [Google Scholar] [CrossRef]
- Martins Filho, A.; Jammal, M.P.; de Castro Côbo, E.; Silveira, T.P.; Adad, S.J.; Murta, E.F.C.; Nomelini, R.S. Correlation of cytokines and inducible nitric oxide synthase expression with prognostic factors in ovarian cancer. Immunol. Lett. 2014, 158, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Ushijima, K. Treatment for recurrent ovarian cancer-at first relapse. J. Oncol. 2010, 2010, 497429. [Google Scholar] [CrossRef] [PubMed]
- Tapia, G.; Diaz-Padilla, I. Molecular Mechanisms of Platinum Resistance in Ovarian Cancer. In Ovarian Cancer-A Clinical and Translational Update; Intech Open: London, UK, 2013. [Google Scholar]
- Brasseur, K.; Gévry, N.; Asselin, E. Chemoresistance and targeted therapies in ovarian and endometrial cancers. Oncotarget 2017, 8, 4008–4042. [Google Scholar] [CrossRef]
- Turchi, J.J. Nitric oxide and cisplatin resistance: NO easy answers. Proc. Natl. Acad. Sci. USA 2006, 103, 4337–4338. [Google Scholar] [CrossRef]
- Kielbik, M.; Klink, M.; Brzezinska, M.; Szulc, I.; Sulowska, Z. Nitric oxide donors: Spermine/NO and diethylenetriamine/NO induce ovarian cancer cell death and affect STAT3 and AKT signaling proteins. Nitric Oxide 2013, 35, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Gupta, A.K. Nitric oxide: Role in tumour biology and iNOS/NO-based anticancer therapies. Cancer Chemother. Pharmacol. 2011, 67, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Sinha, B.K. Nitric Oxide: Friend or Foe in Cancer Chemotherapy and Drug Resistance: A Perspective. J. Cancer Sci. Ther. 2016, 8, 244–251. [Google Scholar] [CrossRef]
- Perrotta, C.; Cervia, D.; Di Renzo, I.; Moscheni, C.; Bassi, M.T.; Campana, L.; Martelli, C.; Catalani, E.; Giovarelli, M.; Zecchini, S.; et al. Nitric Oxide Generated by Tumor-Associated Macrophages Is Responsible for Cancer Resistance to Cisplatin and Correlated With Syntaxin 4 and Acid Sphingomyelinase Inhibition. Front. Immunol. 2018, 9, 1186. [Google Scholar] [CrossRef]
- Leung, E.L.; Fraser, M.; Fiscus, R.R.; Tsang, B.K. Cisplatin alters nitric oxide synthase levels in human ovarian cancer cells: Involvement in p53 regulation and cisplatin resistance. Br. J. Cancer 2008, 98, 1803–1809. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Xie, Q.; Liu, W.; Guo, Y.; Xu, N.; Xu, L.; Liu, S.; Li, S.; Xu, Y.; Sun, L. Increased intracellular Ca2+ decreases cisplatin resistance by regulating iNOS expression in human ovarian cancer cells. Biomed. Pharmacother. 2017, 86, 8–15. [Google Scholar] [CrossRef]
- Belotte, J.; Fletcher, N.M.; Awonuga, A.O.; Alexis, M.; Abu-Soud, H.M.; Saed, M.G.; Diamond, M.P.; Saed, G.M. The role of oxidative stress in the development of cisplatin resistance in epithelial ovarian cancer. Reprod. Sci. 2014, 21, 503–508. [Google Scholar] [CrossRef]
- Malone, J.M.; Saed, G.M.; Diamond, M.P.; Sokol, R.J.; Munkarah, A.R. The effects of the inhibition of inducible nitric oxide synthase on angiogenesis of epithelial ovarian cancer. Am. J. Obstet. Gynecol. 2006, 194, 1110–1116, discussion 1116–1118. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Yu, C.; Zhou, S.; Lau, W.B.; Lau, B.; Luo, Z.; Lin, Q.; Yang, H.; Xuan, Y.; Yi, T.; et al. Epigenetic repression of PDZ-LIM domain-containing protein 2 promotes ovarian cancer via NOS2-derived nitric oxide signaling. Oncotarget 2016, 7, 1408–1420. [Google Scholar] [CrossRef]
- Saed, G.M.; Ali-Fehmi, R.; Jiang, Z.L.; Fletcher, N.M.; Diamond, M.P.; Abu-Soud, H.M.; Munkarah, A.R. Myeloperoxidase serves as a redox switch that regulates apoptosis in epithelial ovarian cancer. Gynecol. Oncol. 2010, 116, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Son, K.K.; Hall, K.J. Nitric oxide-mediated tumor cell killing of cisplatin-based interferon-gamma gene therapy in murine ovarian carcinoma. Cancer Gene Ther. 2000, 7, 1324–1328. [Google Scholar] [CrossRef]
- Xu, W.; Liu, L.; Charles, I.G. Microencapsulated iNOS-expressing cells cause tumor suppression in mice. FASEB J. 2002, 16, 213–215. [Google Scholar] [CrossRef]
- Hendren, S.K.; Prabakaran, I.; Buerk, D.G.; Karakousis, G.; Feldman, M.; Spitz, F.; Menon, C.; Fraker, D.L. Interferon-beta gene therapy improves survival in an immunocompetent mouse model of carcinomatosis. Surgery 2004, 135, 427–436. [Google Scholar] [CrossRef]
- Trinh, B.; Ko, S.Y.; Haria, D.; Barengo, N.; Naora, H. The homeoprotein DLX4 controls inducible nitric oxide synthase-mediated angiogenesis in ovarian cancer. Mol. Cancer 2015, 14, 97. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Votes For | Ref. |
| iNOS expression in ovarian serous carcinoma (FIGO III) is an independent predictor of disease relapse and patients death | [60] |
| High iNOS expression in type II ovarian cancer is a poor prognostic factor | [61] |
| iNOS-negative ovarian serous carcinoma indicate longer progression-free survival | [62] |
| High iNOS expression in ovarian serous and mucinous carcinoma is a poor prognostic factor | [63] |
| High iNOS expression in ovarian serous and mucinous carcinoma is associated with more aggressive phenotype of tumor and poor survival outcome | [64] |
| Votes Against | |
| iNOS expression does not correlate with FIGO and grade and has no prognostic value | [65] |
| iNOS expression has no impact on patients survival | [66] |
| iNOS expression has no impact on FIGO, grade, response to chemotherapy or patients survival | [67] |
| Basal iNOS Expression, NO Production | Cisplatin-Induced iNOS Expression, NO Production | Ref. | ||
|---|---|---|---|---|
| Cisplatin Resistant Ovarian Cancer Cell Line | Cisplatin Sensitive Ovarian Cancer Cell Lines | Cisplatin Resistant Ovarian Cancer Cell Line | Cisplatin Sensitive Ovarian Cancer Cell Lines | |
| Low protein level | High protein level | No induction of iNOS | High induction of iNOS | [76] |
| Low protein level | High protein level | Low induction of iNOS | High induction of iNOS | [77] |
| High mRNA level | Low mRNA level | Nd | nd | [78] |
| Low mRNA level | Low mRNA level | Nd | nd | [78] |
| High NO production | Low NO production | Nd | nd | [71] |
| Ovarian Cancer Model | Treatment | Effect | Ref. |
|---|---|---|---|
| iNOS Inhibition | |||
| Ovarian cancer cell lines | L-NAME -NOS inhibitor | Reduction of VEGF production | [79] |
| Ovarian cancer cell lines | 1400W- iNOS inhibitor iNOS siRNA | Inhibition of cell growth | [80] |
| Ovarian cancer cell lines | iNOS siRNA | Increased activity of caspase 3 Induction of cell apoptosis | [81] |
| Xenograft model | iNOS siRNA | Decrease in number of M2-type of TAM in tumor | [80] |
| iNOS Induction | |||
| Murine ovarian carcinoma | IFN-γ + cisplatin | High level of NO in ascites Enhanced survival of mice | [82] |
| Murine ovarian carcinoma | iNOS-expressed micro-encapsulated cells | Inhibition of tumor growth | [83] |
| Mouse ovarian teratoma | IFN-β | Inhibition of tumor growth Enhanced survival of mice | [84] |
| Ovarian cancer cell lines | DLX4 | Stimulation of STAT1 activity Stimulation of VEGF-A production | [85] |
| Xenograft model | DLX4 | Stimulation of tumor angiogenesis | [85] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kielbik, M.; Szulc-Kielbik, I.; Klink, M. The Potential Role of iNOS in Ovarian Cancer Progression and Chemoresistance. Int. J. Mol. Sci. 2019, 20, 1751. https://doi.org/10.3390/ijms20071751
Kielbik M, Szulc-Kielbik I, Klink M. The Potential Role of iNOS in Ovarian Cancer Progression and Chemoresistance. International Journal of Molecular Sciences. 2019; 20(7):1751. https://doi.org/10.3390/ijms20071751
Chicago/Turabian StyleKielbik, Michal, Izabela Szulc-Kielbik, and Magdalena Klink. 2019. "The Potential Role of iNOS in Ovarian Cancer Progression and Chemoresistance" International Journal of Molecular Sciences 20, no. 7: 1751. https://doi.org/10.3390/ijms20071751
APA StyleKielbik, M., Szulc-Kielbik, I., & Klink, M. (2019). The Potential Role of iNOS in Ovarian Cancer Progression and Chemoresistance. International Journal of Molecular Sciences, 20(7), 1751. https://doi.org/10.3390/ijms20071751