Two Component Regulatory Systems and Antibiotic Resistance in Gram-Negative Pathogens

 , and
, and
Abstract
:1. Introduction
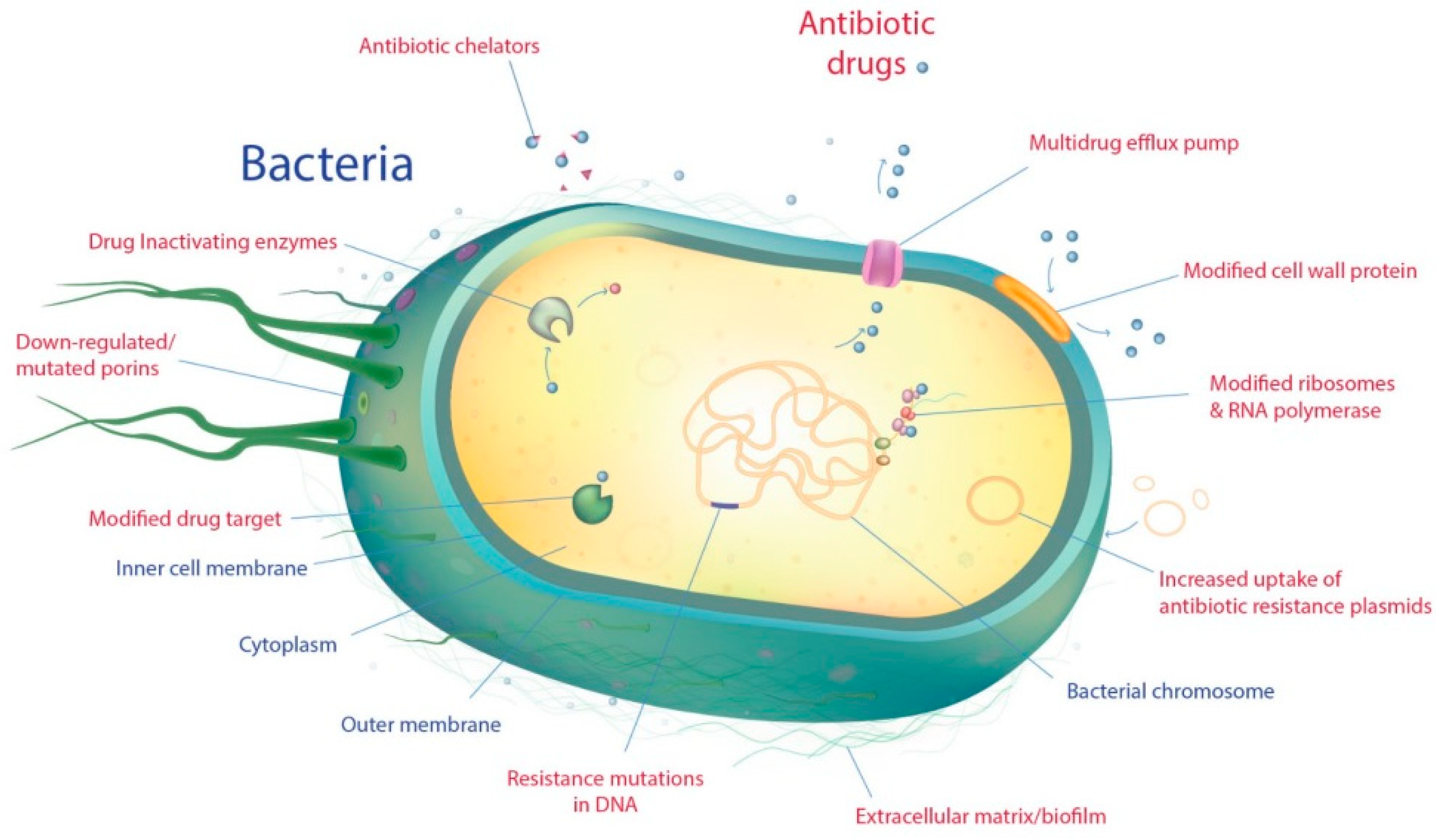
2. Antimicrobial Resistance in Gram-Negative Pathogenic Bacteria
2.1. Intrinsic Resistance
2.2. Acquired Resistance
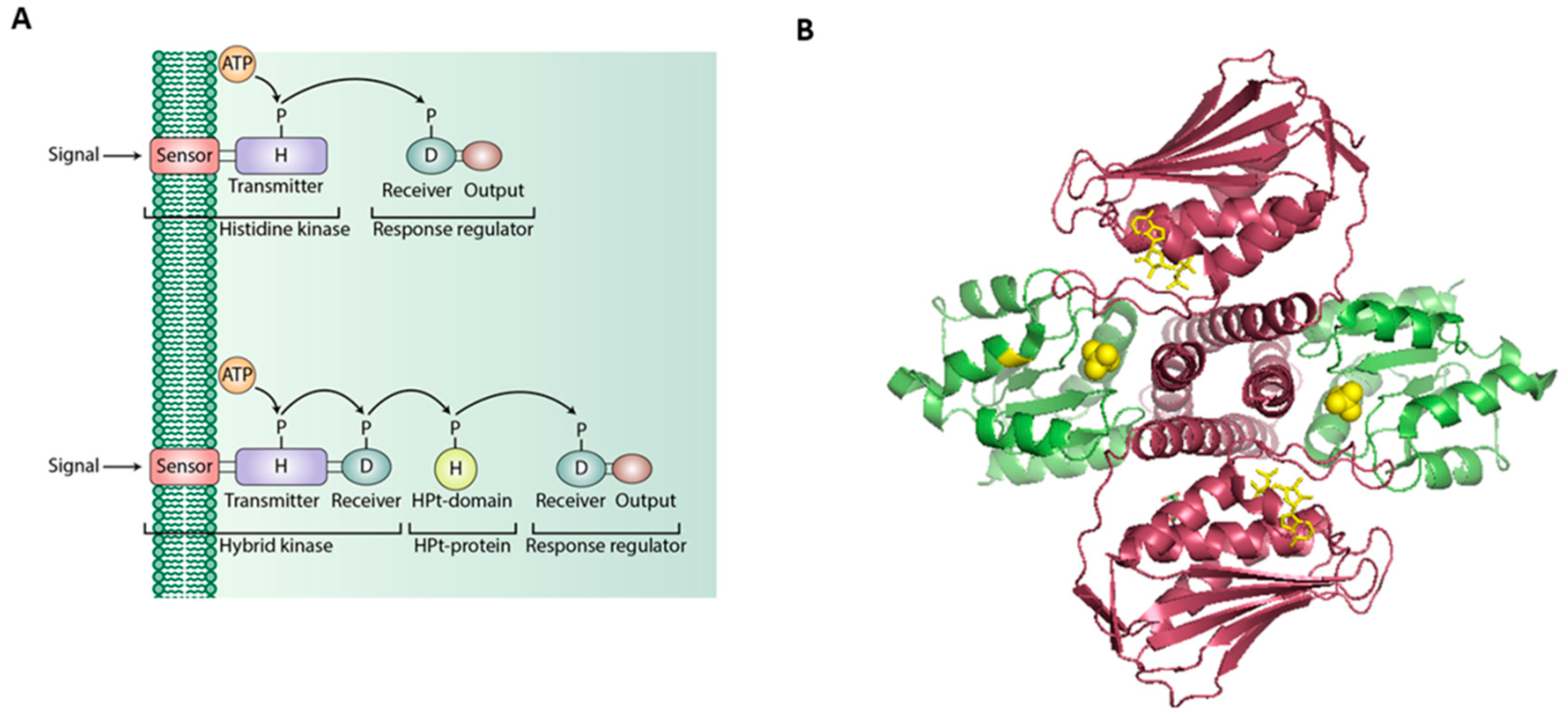
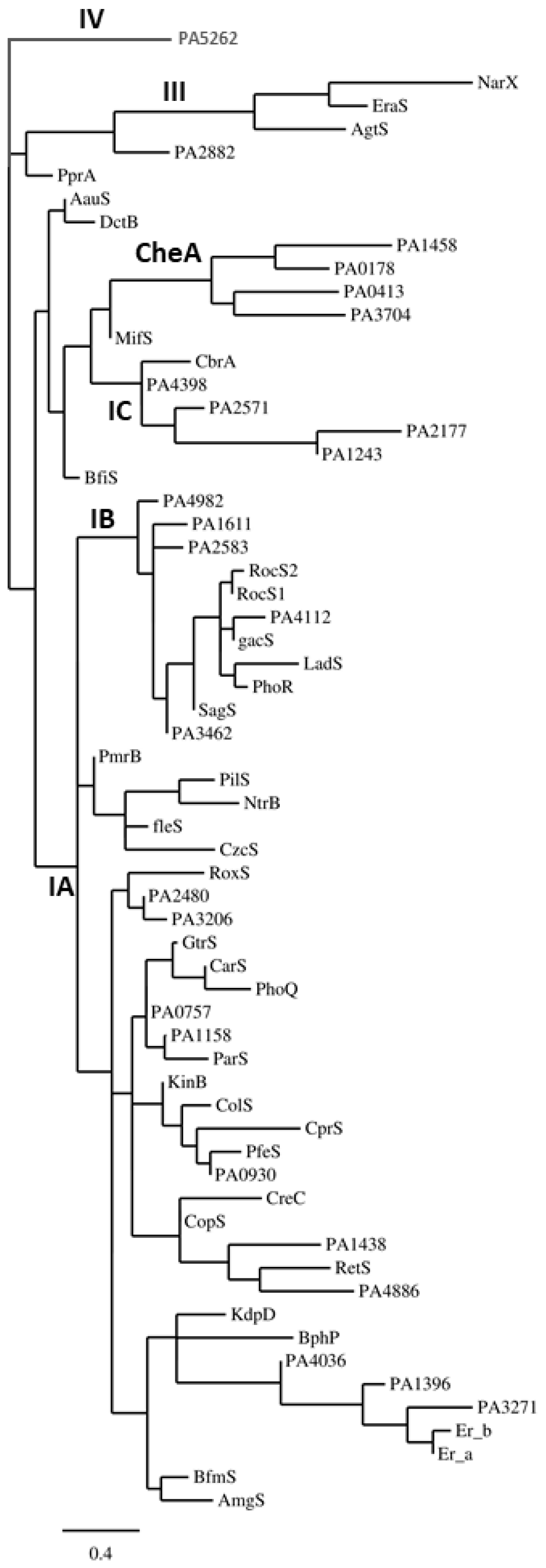
3. Two-Component Regulatory Systems in Gram-Negative Pathogenic Bacteria
3.1. One-Component Signaling Systems
3.2. Hybrid Histidine Kinase [HHK] and Direct-Interaction-Mediated Signaling
4. Role of Two-Component Regulatory Systems in Antimicrobial Resistance in Gram- Negative Pathogenic Bacteria
4.1. The PmrAB System
4.2. The GacSA System
4.3. The AdeRS and the BaeSR Systems
5. Two-Component Regulatory Systems as Potential Drug Targets
6. Conclusions and Perspective
Funding
Conflicts of Interest
Abbreviations
| AAC | Aminoglycoside Acetyltransferase |
| ABC | ATP Binding Cassette |
| AMR | Antimicrobial Resistance |
| CAPs | Cationic Antimicrobial Peptides |
| CDC | Centers for Disease Control and Prevention |
| CF | Cystic Fibrosis |
| CHDL | Carbapenem-Hydrolysing Class D β-Lactamase |
| Csr | Carbon Storage Regulation |
| DSF | Diffusible Signal Factor |
| ESBL | Extended-Spectrum β-lactamase |
| ESCAPE | Enterococcus faecium, Staphylococcus aureus, Clostridium difficile, A. baumannii, P. aeruginosa, and Enterobacteriaceae |
| ESKAPE | Enterococcus faecium, Staphylococcus aureus, K. pneumoniae, A. baumannii, P. aeruginosa, and Enterobacteriaceae |
| HAP | Histidyl-Aspartyl Phosphorelay |
| HGT | Horizontal Gene Transfer |
| HHK | Hybrid Histidine Kinase |
| HK | Histidine Kinase |
| Hpt | Histidine Phosphotransfer Protein |
| HTH | Helix-turn-helix |
| ICU | Intensive Care Unit |
| L-Ara4N | 4-Amino-4-Deoxy-L-Arabinose |
| LPSs | Lipopolysaccharides |
| MATE | Multidrug and Toxin Extrusion |
| MBL | Metallo β-lactamases |
| MDR | Multidrug-Resistant |
| MFS | Major Facilitator Superfamily |
| NDM-1 | New Delhi Metallo- β-lactamase 1 |
| NNIS | National Nosocomial Infections Surveillance |
| OCSs | One-Component Systems |
| OM | Outer Membrane |
| PAA | Phenylacetic Acid |
| PAS | Period clock protein, Aryl hydrocarbon receptor, and Single-minded protein |
| PDR | Pan-Drug Resistant |
| pEtN | Phosphoethanolamine |
| RND | Resistance-Nodulation-Cell Division |
| RR | Response Regulator |
| SCV | Small Colony Variants |
| SMR | Small Multidrug Resistance |
| TCA | Tricarboxylic Acid |
| TCSs | Two-Component Systems |
| WHO | World Health Organization |
References
- World Health Organization. Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug-Resistant Bacterial Infections, Including Tuberculosis; World Health Organization: Geneva, Switzerland, 2017; p. 12. [Google Scholar]
- Chairat, S.; Ben Yahia, H.; Rojo-Bezares, B.; Saenz, Y.; Torres, C.; Ben Slama, K. High prevalence of imipenem-resistant and metallo-beta-lactamase-producing Pseudomonas aeruginosa in the Burns Hospital in Tunisia: Detection of a novel class 1 integron. J. Chemother. 2019, 1–7. [Google Scholar] [CrossRef]
- Zhe, S.; Qianru, Z.; Liying, Z.; Zhidong, Z.; Ling, J.; He, H. Draft genome sequence of a multidrug-resistant beta-lactamase-harboring Bacillus cereusS66, isolated from China. J. Glob. Antimicrob. Resist. 2019. [Google Scholar] [CrossRef]
- Goic-Barisic, I.; Seruga Music, M.; Kovacic, A.; Tonkic, M.; Hrenovic, J. Pan drug-resistant environmental Isolate of Acinetobacter baumannii from Croatia. Microb. Drug Resist. 2017, 23, 494–496. [Google Scholar] [CrossRef]
- Li, L.; Yu, T.; Ma, Y.; Yang, Z.; Wang, W.; Song, X.; Shen, Y.; Guo, T.; Kong, J.; Wang, M.; Xu, H. The genetic structures of an Extensively Drug Resistant (XDR) Klebsiella pneumoniae and Its plasmids. Front. Cell Infect. Microbiol. 2018, 8, 446. [Google Scholar] [CrossRef]
- Wang, C.Y.; Jerng, J.S.; Chen, K.Y.; Lee, L.N.; Yu, C.J.; Hsueh, P.R.; Yang, P.C. Pandrug-resistant Pseudomonas aeruginosa among hospitalised patients: Clinical features, risk-factors and outcomes. Clin. Microbiol. Infect. 2006, 12, 63–68. [Google Scholar] [CrossRef]
- Sonnevend, Á.; Ghazawi, A.; Hashmey, R.; Haidermota, A.; Girgis, S.; Alfaresi, M.; Omar, M.; Paterson, D.L.; Zowawi, H.M.; Pál, T. Multihospital occurrence of pan-resistant Klebsiella pneumoniae sequence type 147 with an ISEcp1-directed blaOXA-181 Insertion in the mgrB gene in the United Arab Emirates. Antimicrob. Agents Chemother. 2017, 61, e00418-17. [Google Scholar] [CrossRef]
- Hsueh, P.-R.; Teng, L.-J.; Chen, C.-Y.; Chen, W.-H.; Yu, C.-J.; Ho, S.-W.; Luh, K.-T. Pandrug-resistant Acinetobacter baumannii causing nosocomial infections in a university hospital, Taiwan. Emerg. Infect. Dis. 2002, 8, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.; Vira, D.; Medikonda, R.; Kumar, N. Extensively and pan-drug resistant Pseudomonas aeruginosa keratitis: Clinical features, risk factors, and outcome. Graefes Arch. Clin. Exp. Ophthalmol. 2016, 254, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Ozer, E.A.; Krapp, F.; Hauser, A.R.; Qi, C. Case report of an extensively drug-resistant Klebsiella pneumoniae Infection with genomic characterization of the strain and review of similar cases in the United States. Open. Forum Infect. Dis. 2018, 5. [Google Scholar] [CrossRef]
- Peterson, L.R. Bad bugs, no drugs: No ESCAPE revisited. Clin. Infect. Dis. 2009, 49, 992–993. [Google Scholar] [CrossRef]
- Cabot, G.; Zamorano, L.; Moya, B.; Juan, C.; Navas, A.; Blazquez, J.; Oliver, A. Evolution of Pseudomonas aeruginosa antimicrobial resistance and fitness under low and high mutation rates. Antimicrob. Agents Chemother. 2016, 60, 1767–1778. [Google Scholar] [CrossRef]
- Tsakiridou, E.; Makris, D.; Daniil, Z.; Manoulakas, E.; Chatzipantazi, V.; Vlachos, O.; Xidopoulos, G.; Charalampidou, O.; Zakynthinos, E. Acinetobacter baumannii infection in prior ICU bed occupants is an independent risk factor for subsequent cases of ventilator-associated pneumonia. BioMed. Res. Int. 2014, 2014, 193516. [Google Scholar] [CrossRef]
- Martin, R.M.; Bachman, M.A. Colonization, infection, and the accessory genome of Klebsiella pneumoniae. Front. Cell Infect. Microbiol. 2018, 8, 4. [Google Scholar] [CrossRef]
- Gaynes, R.; Edwards, J.R. Overview of nosocomial infections caused by gram-negative bacilli. Clin. Infect. Dis. 2005, 41, 848–854. [Google Scholar]
- Landman, D.; Trehan, M.; Panwar, M.; Kochar, S.; Bratu, S.; Quale, J.; Doymaz, M. Evolution of antimicrobial resistance among Pseudomonas aeruginosa, Acinetobacter baumannii and Klebsiella pneumoniae in Brooklyn, NY. J. Antimicrob. Chemother. 2007, 60, 78–82. [Google Scholar] [CrossRef]
- Streit, J.M.; Jones, R.N.; Sader, H.S.; Fritsche, T.R. Assessment of pathogen occurrences and resistance profiles among infected patients in the intensive care unit: Report from the SENTRY Antimicrobial Surveillance Program (North America, 2001). Int. J. Antimicrob. Agents 2004, 24, 111–118. [Google Scholar] [CrossRef]
- Lingzhi, L.; Haojie, G.; Dan, G.; Hongmei, M.; Yang, L.; Mengdie, J.; Chengkun, Z.; Xiaohui, Z. The role of two-component regulatory system in beta-lactam antibiotics resistance. Microbiol. Res. 2018, 215, 126–129. [Google Scholar] [CrossRef]
- Gutu, A.D.; Sgambati, N.; Strasbourger, P.; Brannon, M.K.; Jacobs, M.A.; Haugen, E.; Kaul, R.K.; Johansen, H.K.; Hoiby, N.; Moskowitz, S.M. Polymyxin resistance of Pseudomonas aeruginosa phoQ mutants is dependent on additional two-component regulatory systems. Antimicrob. Agents Chemother. 2013, 57, 2204–2215. [Google Scholar] [CrossRef]
- Hirsch, E.B.; Tam, V.H. Impact of multidrug-resistant Pseudomonas aeruginosa infection on patient outcomes. Expert Rev. Pharm. Outcomes Res. 2010, 10, 441–451. [Google Scholar]
- Cella, E.; Ciccozzi, M.; Lo Presti, A.; Fogolari, M.; Azarian, T.; Prosperi, M.; Salemi, M.; Equestre, M.; Antonelli, F.; Conti, A.; et al. Multi-drug resistant Klebsiella pneumoniae strains circulating in hospital setting: Whole-genome sequencing and Bayesian phylogenetic analysis for outbreak investigations. Sci. Rep. 2017, 7, 3534. [Google Scholar] [CrossRef]
- Mira, A.; Apalara, J.; Thyagarajan, R.; Sengstock, D.M.; Kaye, K.S.; Chopra, T. Multidrug-resistant Acinetobacter baumannii: An emerging pathogen among older adults in community hospitals and nursing homes. Clin. Infect. Dis. 2010, 50, 1611–1616. [Google Scholar]
- Martinez, J.L.; Baquero, F. Mutation frequencies and antibiotic resistance. Antimicrob. Agents Chemother. 2000, 44, 1771–1777. [Google Scholar] [CrossRef]
- Andam, C.P.; Fournier, G.P.; Gogarten, J.P. Multilevel populations and the evolution of antibiotic resistance through horizontal gene transfer. FEMS Microbiol. Rev. 2011, 35, 756–767. [Google Scholar] [CrossRef] [Green Version]
- Sommer, M.O.A.; Dantas, G.; Church, G.M. Functional characterization of the antibiotic resistance reservoir in the human microflora. Science 2009, 325, 1128–1131. [Google Scholar] [CrossRef]
- Davies, J.E. Origins, acquisition and dissemination of antibiotic resistance determinants. Ciba Found Symp. 1997, 207, 15–27. [Google Scholar]
- Blanquart, F.; Lehtinen, S.; Lipsitch, M.; Fraser, C. The evolution of antibiotic resistance in a structured host population. J. R. Soc. Interface 2018, 15, 20180040. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Lee, S.; Zhao, X.; Dong, Y.; Drlica, K.; Amin, A.; Musser, J.M.; Ramaswamy, S.; Domagala, J. Selection of antibiotic-resistant bacterial mutants: Allelic diversity among fluoroquinolone-resistant mutations. J. Infect. Dis. 2000, 182, 517–525. [Google Scholar] [CrossRef]
- Gniadkowski, M. Evolution of extended-spectrum beta-lactamases by mutation. Clin. Microbiol. Infect. 2008, 14, 11–32. [Google Scholar] [CrossRef]
- Bush, K. Past and present perspectives on beta-Lactamases. Antimicrob. Agents Chemother. 2018, 62, e01076-18. [Google Scholar] [CrossRef]
- Gutkind, G.O.; Di Conza, J.; Power, P.; Radice, M. Beta-lactamase-mediated resistance: A biochemical, epidemiological and genetic overview. Curr. Pharm. Des. 2013, 19, 164–208. [Google Scholar] [CrossRef]
- Bajaj, P.; Singh, N.S.; Virdi, J.S. Escherichia coli β-Lactamases: What Really Matters. Front. Microbiol. 2016, 7, 417. [Google Scholar] [CrossRef]
- Martinez, J.L. The antibiotic resistome: Challenge and opportunity for therapeutic intervention. Future Med. Chem. 2012, 4, 347–359. [Google Scholar] [CrossRef]
- Perry, J.A.; Wright, G.D. The antibiotic resistance “mobilome”: Searching for the link between environment and clinic. Front. Microbiol. 2013, 4, 138. [Google Scholar] [CrossRef]
- Corona, F.; Blanco, P.; Alcalde-Rico, M.; Hernando-Amado, S.; Lira, F.; Bernardini, A.; Sanchez, M.B.; Martinez, J.L. The analysis of the antibiotic resistome offers new opportunities for therapeutic intervention. Future Med. Chem. 2016, 8, 1133–1151. [Google Scholar] [CrossRef]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef]
- Veleba, M.; Higgins, P.G.; Gonzalez, G.; Seifert, H.; Schneiders, T. Characterization of RarA, a novel AraC family multidrug resistance regulator in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2012, 56, 4450–4458. [Google Scholar] [CrossRef]
- Jana, B.; Cain, A.K.; Doerrler, W.T.; Boinett, C.J.; Fookes, M.C.; Parkhill, J.; Guardabassi, L. The secondary resistome of multidrug-resistant Klebsiella pneumoniae. Sci. Rep. 2017, 7, 42483. [Google Scholar] [CrossRef]
- Magnet, S.; Courvalin, P.; Lambert, T. Resistance-nodulation-cell division-type efflux pump involved in aminoglycoside resistance in Acinetobacter baumannii strain BM4454. Antimicrob. Agents Chemother. 2001, 45, 3375–3380. [Google Scholar] [CrossRef]
- Damier-Piolle, L.; Magnet, S.; Bremont, S.; Lambert, T.; Courvalin, P. AdeIJK, a resistance-nodulation-cell division pump effluxing multiple antibiotics in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2008, 52, 557–562. [Google Scholar] [CrossRef]
- Yoon, E.J.; Chabane, Y.N.; Goussard, S.; Snesrud, E.; Courvalin, P.; De, E.; Grillot-Courvalin, C. Contribution of resistance-nodulation-cell division efflux systems to antibiotic resistance and biofilm formation in Acinetobacter baumannii. mBio 2015, 6, e00309-15. [Google Scholar] [CrossRef]
- Coyne, S.; Courvalin, P.; Perichon, B. Efflux-mediated antibiotic resistance in Acinetobacter spp. Antimicrob. Agents Chemother. 2011, 55, 947–953. [Google Scholar] [CrossRef]
- Zgurskaya, H.I.; Löpez, C.A.; Gnanakaran, S. Permeability barrier of Gram-negative cell envelopes and approaches to bypass It. ACS Infect. Dis. 2015, 1, 512–522. [Google Scholar] [CrossRef]
- Nakae, T. Outer membrane of Salmonella. Isolation of protein complex that produces transmembrane channels. J. Biol. Chem. 1976, 251, 2176–2178. [Google Scholar]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. MMBR 2003, 67, 593–656. [Google Scholar] [CrossRef]
- Wise, M.G.; Horvath, E.; Young, K.; Sahm, D.F.; Kazmierczak, K.M. Global survey of Klebsiella pneumoniae major porins from ertapenem non-susceptible isolates lacking carbapenemases. J. Med. Microbiol. 2018, 67, 289–295. [Google Scholar] [CrossRef]
- Hong, J.H.; Clancy, C.J.; Cheng, S.; Shields, R.K.; Chen, L.; Doi, Y.; Zhao, Y.; Perlin, D.S.; Kreiswirth, B.N.; Nguyen, M.H. Characterization of porin expression in Klebsiella pneumoniae Carbapenemase (KPC)-producing K. pneumoniae identifies isolates most susceptible to the combination of colistin and carbapenems. Antimicrob. Agents Chemother. 2013, 57, 2147–2153. [Google Scholar] [CrossRef]
- Pulzova, L.; Navratilova, L.; Comor, L. Alterations in outer membrane permeability favor drug-resistant phenotype of Klebsiella pneumoniae. Microb. Drug Resist. 2017, 23, 413–420. [Google Scholar] [CrossRef]
- Domenech-Sanchez, A.; Hernandez-Alles, S.; Martinez-Martinez, L.; Benedi, V.J.; Alberti, S. Identification and characterization of a new porin gene of Klebsiella pneumoniae: Its role in beta-lactam antibiotic resistance. J. Bacteriol. 1999, 181, 2726–2732. [Google Scholar]
- Martinez-Martinez, L.; Pascual, A.; Conejo Mdel, C.; Garcia, I.; Joyanes, P.; Domenech-Sanchez, A.; Benedi, V.J. Energy-dependent accumulation of norfloxacin and porin expression in clinical isolates of Klebsiella pneumoniae and relationship to extended-spectrum beta-lactamase production. Antimicrob. Agents Chemother. 2002, 46, 3926–3932. [Google Scholar] [CrossRef]
- Smani, Y.; Fabrega, A.; Roca, I.; Sanchez-Encinales, V.; Vila, J.; Pachon, J. Role of OmpA in the multidrug resistance phenotype of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2014, 58, 1806–1808. [Google Scholar] [CrossRef]
- Clemmer, K.M.; Bonomo, R.A.; Rather, P.N. Genetic analysis of surface motility in Acinetobacter baumannii. Microbiology 2011, 157, 2534–2544. [Google Scholar] [CrossRef]
- Gaddy, J.A.; Tomaras, A.P.; Actis, L.A. The Acinetobacter baumannii 19606 OmpA protein plays a role in biofilm formation on abiotic surfaces and in the interaction of this pathogen with eukaryotic cells. Infect. Immun. 2009, 77, 3150–3160. [Google Scholar] [CrossRef]
- Aeschlimann, J.R. The role of multidrug efflux pumps in the antibiotic resistance of Pseudomonas aeruginosa and other gram-negative bacteria. Insights from the Society of Infectious Diseases Pharmacists. Pharmacotherapy 2003, 23, 916–924. [Google Scholar] [CrossRef]
- Evans, K.; Adewoye, L.; Poole, K. MexR repressor of the mexAB-oprM multidrug efflux operon of Pseudomonas aeruginosa: Identification of MexR binding sites in the mexA-mexR Intergenic Region. J. Bacteriol. 2001, 183, 807–812. [Google Scholar] [CrossRef]
- Sun, J.; Deng, Z.; Yan, A. Bacterial multidrug efflux pumps: Mechanisms, physiology and pharmacological exploitations. Biochem. Biophys. Res. Commun. 2014, 453, 254–267. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Chen, L.; Shen, L.; Surette, M.; Duan, K. Inactivation of MuxABC-OpmB transporter system in Pseudomonas aeruginosa leads to increased ampicillin and carbenicillin resistance and decreased virulence. J. Microbiol. 2011, 49, 107–114. [Google Scholar] [CrossRef]
- Marchand, I.; Damier-Piolle, L.; Courvalin, P.; Lambert, T. Expression of the RND-type efflux pump AdeABC in Acinetobacter baumannii is regulated by the AdeRS two-component system. Antimicrob. Agents Chemother. 2004, 48, 3298–3304. [Google Scholar] [CrossRef]
- Wieczorek, P.; Sacha, P.; Hauschild, T.; Zorawski, M.; Krawczyk, M.; Tryniszewska, E. Multidrug resistant Acinetobacter baumannii—The role of AdeABC (RND family) efflux pump in resistance to antibiotics. Folia Histochem. Cytobiol. 2008, 46, 257–267. [Google Scholar] [CrossRef]
- Yoon, E.J.; Balloy, V.; Fiette, L.; Chignard, M.; Courvalin, P.; Grillot-Courvalin, C. Contribution of the Ade resistance-nodulation-cell division-type efflux pumps to fitness and pathogenesis of Acinetobacter baumannii. mBio 2016, 7, e00697-16. [Google Scholar] [CrossRef]
- Ruzin, A.; Visalli, M.A.; Keeney, D.; Bradford, P.A. Influence of transcriptional activator RamA on expression of multidrug efflux pump AcrAB and tigecycline susceptibility in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2005, 49, 1017–1022. [Google Scholar] [CrossRef]
- Li, D.W.; Onishi, M.; Kishino, T.; Matsuo, T.; Ogawa, W.; Kuroda, T.; Tsuchiya, T. Properties and expression of a multidrug efflux pump AcrAB-KocC from Klebsiella pneumoniae. Biol. Pharm. Bull. 2008, 31, 577–582. [Google Scholar] [CrossRef]
- Ogawa, W.; Onishi, M.; Ni, R.; Tsuchiya, T.; Kuroda, T. Functional study of the novel multidrug efflux pump KexD from Klebsiella pneumoniae. Gene 2012, 498, 177–182. [Google Scholar] [CrossRef]
- El Zowalaty, M.E.; Al Thani, A.A.; Webster, T.J.; El Zowalaty, A.E.; Schweizer, H.P.; Nasrallah, G.K.; Marei, H.E.; Ashour, H.M. Pseudomonas aeruginosa: Arsenal of resistance mechanisms, decades of changing resistance profiles, and future antimicrobial therapies. Future Microbiol. 2015, 10, 1683–1706. [Google Scholar] [CrossRef]
- Van Hoek, A.H.A.M.; Mevius, D.; Guerra, B.; Mullany, P.; Roberts, A.P.; Aarts, H.J.M. Acquired antibiotic resistance genes: An overview. Front. Microbiol. 2011, 2, 203. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Updat 2010, 13, 151–171. [Google Scholar] [CrossRef] [Green Version]
- Wright, G.D. Bacterial resistance to antibiotics: Enzymatic degradation and modification. Adv. Drug Deliv. Rev. 2005, 57, 1451–1470. [Google Scholar] [CrossRef]
- Hoiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, R.F.; Sá-Correia, I.; Valvano, M.A. Lipopolysaccharide modification in Gram-negative bacteria during chronic infection. FEMS Microbiol. Rev. 2016, 40, 480–493. [Google Scholar] [CrossRef] [Green Version]
- Lerminiaux, N.A.; Cameron, A.D.S. Horizontal transfer of antibiotic resistance genes in clinical environments. Can. J. Microbiol. 2019, 65, 34–44. [Google Scholar] [CrossRef]
- O’Toole, G.; Kaplan, H.B.; Kolter, R. Biofilm formation as microbial development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef]
- Stewart, P.S. Mechanisms of antibiotic resistance in bacterial biofilms. Zentralbl. Bakteriol. 2002, 292, 107–113. [Google Scholar] [CrossRef]
- Mah, T.F.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Curr. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Hoffman, L.R.; D’Argenio, D.A.; MacCoss, M.J.; Zhang, Z.; Jones, R.A.; Miller, S.I. Aminoglycoside antibiotics induce bacterial biofilm formation. Nature 2005, 436, 1171–1175. [Google Scholar] [CrossRef]
- Perez-Martinez, I.; Haas, D. Azithromycin inhibits expression of the GacA-dependent small RNAs RsmY and RsmZ in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2011, 55, 3399–3405. [Google Scholar] [CrossRef]
- Ahmed, M.N.; Porse, A.; Sommer, M.O.A.; Høiby, N.; Ciofu, O. Evolution of antibiotic resistance in biofilm and planktonic Pseudomonas aeruginosa populations exposed to subinhibitory levels of Ciprofloxacin. Antimicrob. Agents Chemother. 2018, 62, e00320-18. [Google Scholar] [CrossRef]
- Cordero, O.X.; Hogeweg, P. The impact of long-distance horizontal gene transfer on prokaryotic genome size. Proc. Natl. Acad. Sci. USA 2009, 106, 21748–21753. [Google Scholar] [CrossRef] [Green Version]
- Von Wintersdorff, C.J.H.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef]
- Otto, M. Bacterial sensing of antimicrobial peptides. Contrib. Microbiol. Immunol. 2009, 16, 136–149. [Google Scholar] [Green Version]
- Rodrigue, A.; Quentin, Y.; Lazdunski, A.; Méjean, V.; Foglino, M. Cell signalling by oligosaccharides. Two-component systems in Pseudomonas aeruginosa: Why so many? Curr. Trends Microbiol. 2000, 8, 498–504. [Google Scholar] [CrossRef]
- Ma, S.; Wozniak, D.; Ohman, D. Identification of the histidine protein kinase KinB in Pseudomonas aeruginosa and its phosphorylation of the alginate regulator algB. J. Biol. Chem. 1997, 272, 17952–17960. [Google Scholar] [CrossRef]
- Benkert, B.; Quack, N.; Schreiber, K.; Jaensch, L.; Jahn, D.; Schobert, M. Nitrate-responsive NarX-NarL represses arginine-mediated induction of the Pseudomonas aeruginosa arginine fermentation arcDABC operon. Microbiolgy (Reading, England) 2008, 154, 3053–3060. [Google Scholar] [CrossRef]
- Kimbara, K.; Chakrabarty, A.M. Control of alginate synthesis in Pseudomonas aeruginosa: Regulation of the algR1 gene. Biochem. Biophys. Res. Commun. 1989, 164, 601–608. [Google Scholar] [CrossRef]
- Bielecki, P.; Jensen, V.; Schulze, W.; Godeke, J.; Strehmel, J.; Eckweiler, D.; Nicolai, T.; Bielecka, A.; Wille, T.; Gerlach, R.G.; et al. Cross talk between the response regulators PhoB and TctD allows for the integration of diverse environmental signals in Pseudomonas aeruginosa. Nucleic Acids Res. 2015, 43, 6413–6425. [Google Scholar] [CrossRef]
- Dean, C.R.; Neshat, S.; Poole, K. PfeR, an enterobactin-responsive activator of ferric enterobactin receptor gene expression in Pseudomonas aeruginosa. J. Bacteriol. 1996, 178, 5361–5369. [Google Scholar] [CrossRef]
- Ritchings, B.W.; Almira, E.C.; Lory, S.; Ramphal, R. Cloning and phenotypic characterization of fleS and fleR, new response regulators of Pseudomonas aeruginosa which regulate motility and adhesion to mucin. Infect. Immun. 1995, 63, 4868–4876. [Google Scholar]
- Yeung, A.T.; Bains, M.; Hancock, R.E. The sensor kinase CbrA is a global regulator that modulates metabolism, virulence, and antibiotic resistance in Pseudomonas aeruginosa. J. Bacteriol. 2011, 193, 918–931. [Google Scholar] [CrossRef]
- Lukat, G.S.; Stock, J.B. Response regulation in bacterial chemotaxis. J. Cell Biochem. 1993, 51, 41–46. [Google Scholar] [CrossRef]
- Casino, P.; Rubio, V.; Marina, A. Structural insight into partner specificity and phosphoryl transfer in two-component signal transduction. Cell 2009, 139, 325–336. [Google Scholar] [CrossRef]
- Kim, D.; Forst, S. Genomic analysis of the histidine kinase family in bacteria and archaea. Microbiology 2001, 147, 1197–1212. [Google Scholar] [CrossRef] [Green Version]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; Claverie, J.M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Dereeper, A.; Audic, S.; Claverie, J.M.; Blanc, G. BLAST-EXPLORER helps you building datasets for phylogenetic analysis. BMC Evol. Biol. 2010, 10, 8. [Google Scholar] [CrossRef]
- Dago, A.E.; Schug, A.; Procaccini, A.; Hoch, J.A.; Weigt, M.; Szurmant, H. Structural basis of histidine kinase autophosphorylation deduced by integrating genomics, molecular dynamics, and mutagenesis. Proc. Natl. Acad. Sci. USA 2012, 109, E1733–E1742. [Google Scholar] [CrossRef] [Green Version]
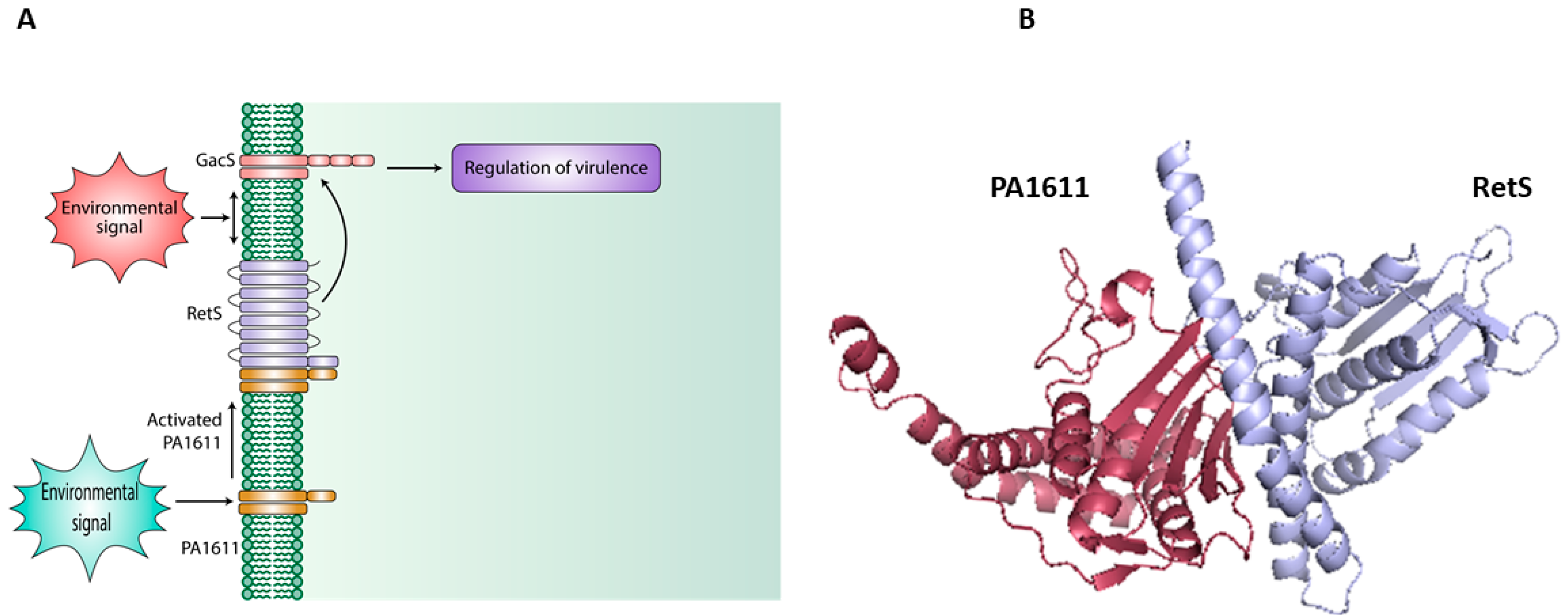
- Bhagirath, A.Y.; Pydi, S.P.; Li, Y.; Lin, C.; Kong, W.; Chelikani, P.; Duan, K. Characterization of the direct Interaction between hybrid sensor kinases PA1611 and RetS that controls biofilm formation and the type III secretion system in Pseudomonas aeruginosa. ACS Infect. Dis. 2017, 3, 162–175. [Google Scholar] [CrossRef]
- Ulrich, L.; Koonin, E.; Zhulin, I. One-component systems dominate signal transduction in prokaryotes. Curr. Trends Microbiol. 2005, 13, 52–56. [Google Scholar] [CrossRef] [Green Version]
- Koretke, K.; Lupas, A.; Warren, P.; Rosenberg, M.; Brown, J. Evolution of two-component signal transduction. Mol. Biol. Evol. 2000, 17, 1956–1970. [Google Scholar] [CrossRef]
- Goodman, A.L.; Kulasekara, B.; Rietsch, A.; Boyd, D.; Smith, R.S.; Lory, S. A signaling network reciprocally regulates genes associated with acute infection and chronic persistence in Pseudomonas aeruginosa. Dev. Cell 2004, 7, 745–754. [Google Scholar] [CrossRef]
- Goodman, A.L.; Merighi, M.; Hyodo, M.; Ventre, I.; Filloux, A.; Lory, S. Direct interaction between sensor kinase proteins mediates acute and chronic disease phenotypes in a bacterial pathogen. Genes Dev. 2009, 23, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Chambonnier, G.; Roux, L.; Redelberger, D.; Fadel, F.; Filloux, A.; Sivaneson, M.; de Bentzmann, S.; Bordi, C. The hybrid histidine kinase LadS forms a multicomponent signal transduction system with the GacS/GacA two-component system in Pseudomonas aeruginosa. PLoS Genet. 2016, 12, e1006032. [Google Scholar] [CrossRef]
- Mitrophanov, A.; Groisman, E. Signal integration in bacterial two-component regulatory systems. Genes Dev. 2008, 22, 2601–2611. [Google Scholar] [CrossRef] [Green Version]
- Ramos, P.I.; Custodio, M.G.; Quispe Saji, G.D.; Cardoso, T.; da Silva, G.L.; Braun, G.; Martins, W.M.; Girardello, R.; de Vasconcelos, A.T.; Fernandez, E.; et al. The polymyxin B-induced transcriptomic response of a clinical, multidrug-resistant Klebsiella pneumoniae involves multiple regulatory elements and intracellular targets. BMC Genom. 2016, 17, 737. [Google Scholar] [CrossRef]
- Babouee Flury, B.; Dona, V.; Buetti, N.; Furrer, H.; Endimiani, A. First two cases of severe multifocal infections caused by Klebsiella pneumoniae in Switzerland: Characterization of an atypical non-K1/K2-serotype strain causing liver abscess and endocarditis. J. Glob. Antimicrob. Resist. 2017, 10, 165–170. [Google Scholar] [CrossRef]
- Dorman, M.J.; Feltwell, T.; Goulding, D.A.; Parkhill, J.; Short, F.L. The capsule regulatory network of Klebsiella pneumoniae defined by density-TraDISort. mBio 2018, 9, e01863-18. [Google Scholar] [CrossRef]
- Merighi, M.; Carroll-Portillo, A.; Septer, A.N.; Bhatiya, A.; Gunn, J.S. Role of Salmonella enterica serovar typhimurium two-component system PreA/PreB in modulating PmrA-regulated gene transcription. J. Bacteriol. 2006, 188, 141–149. [Google Scholar] [CrossRef]
- Mourey, L.; Da Re, S.; Pedelacq, J.; Tolstykh, T.; Faurie, C.; Guillet, V.; Stock, J.; Samama, J. Crystal structure of the CheA histidine phosphotransfer domain that mediates response regulator phosphorylation in bacterial chemotaxis. J. Biol. Chem. 2001, 276, 31074–31082. [Google Scholar] [CrossRef]
- Schaller, G.E.; Shiu, S.H.; Armitage, J.P. Two-component systems and their co-option for eukaryotic signal transduction. Curr. Biol. 2011, 21, R320–R330. [Google Scholar] [CrossRef]
- Pena-Sandoval, G.R.; Kwon, O.; Georgellis, D. Requirement of the receiver and phosphotransfer domains of ArcB for efficient dephosphorylation of phosphorylated ArcA in vivo. J. Bacteriol. 2005, 187, 3267–3272. [Google Scholar] [CrossRef]
- Dingemans, J.; Al-Feghali, R.E.; Lau, G.W.; Sauer, K. Controlling chronic Pseudomonas aeruginosa infections by strategically interfering with the sensory function of SagS. Mol. Microbiol. 2019. [Google Scholar] [CrossRef]
- Dingemans, J.; Poudyal, B.; Sondermann, H.; Sauer, K. The yin and yang of SagS: Distinct residues in the HmsP domain of SagS Independently regulate biofilm formation and biofilm drug tolerance. mSphere 2018, 3, e00192-18. [Google Scholar] [CrossRef]
- Pamp, S.J.; Gjermansen, M.; Johansen, H.K.; Tolker-Nielsen, T. Tolerance to the antimicrobial peptide colistin in Pseudomonas aeruginosa biofilms is linked to metabolically active cells, and depends on the pmr and mexAB-oprM genes. Mol. Microbiol. 2008, 68, 223–240. [Google Scholar] [CrossRef]
- Gupta, K.; Liao, J.; Petrova, O.E.; Cherny, K.E.; Sauer, K. Elevated levels of the second messenger c-di-GMP contribute to antimicrobial resistance of Pseudomonas aeruginosa. Mol. Microbiol. 2014, 92, 488–506. [Google Scholar] [CrossRef]
- Gardner, S.G.; Miller, J.B.; Dean, T.; Robinson, T.; Erickson, M.; Ridge, P.G.; McCleary, W.R. Genetic analysis, structural modeling, and direct coupling analysis suggest a mechanism for phosphate signaling in Escherichia coli. BMC Genet. 2015, 16, S2. [Google Scholar] [CrossRef]
- Mitchell, S.L.; Ismail, A.M.; Kenrick, S.A.; Camilli, A. The VieB auxiliary protein negatively regulates the VieSA signal transduction system in Vibrio cholerae. BMC Microbiol. 2015, 15, 59. [Google Scholar] [CrossRef]
- Airola, M.V.; Watts, K.J.; Bilwes, A.M.; Crane, B.R. Structure of concatenated HAMP domains provides a mechanism for signal transduction. Structure 2010, 18, 436–448. [Google Scholar] [CrossRef]
- Hsing, W.; Silhavy, T.J. Function of conserved histidine-243 in phosphatase activity of EnvZ, the sensor for porin osmoregulation in Escherichia coli. J. Bacteriol. 1997, 179, 3729–3735. [Google Scholar] [CrossRef]
- Matamouros, S.; Hager, K.R.; Miller, S.I. HAMP domain rotation and tilting movements associated with signal transduction in the PhoQ sensor kinase. mBio 2015, 6, e00616-15. [Google Scholar] [CrossRef]
- Meena, N.; Kaur, H.; Mondal, A.K. Interactions among HAMP domain repeats act as an osmosensing molecular switch in group III hybrid histidine kinases from fungi. J. Biol. Chem. 2010, 285, 12121–12132. [Google Scholar] [CrossRef]
- Muzamal, U.; Gomez, D.; Kapadia, F.; Golemi-Kotra, D. Diversity of two-component systems: Insights into the signal transduction mechanism by the Staphylococcus aureus two-component system GraSR. F1000Res 2014, 3, 252. [Google Scholar] [CrossRef]
- Hellingwerf, K.J.; Postma, P.W.; Tommassen, J.; Westerhoff, H.V. Signal transduction in bacteria: Phospho-neural network(s) in Escherichia coli? FEMS Microbiol. Rev. 1995, 16, 309–321. [Google Scholar] [CrossRef]
- Gronlund, A. Networking genetic regulation and neural computation: Directed network topology and its effect on the dynamics. Phys. Rev. E Stat. Nonlin Soft Matter Phys. 2004, 70, 061908. [Google Scholar] [CrossRef]
- Huynh, T.N.; Chen, L.L.; Stewart, V. Sensor-response regulator interactions in a cross-regulated signal transduction network. Microbiology 2015, 161, 1504–1515. [Google Scholar] [CrossRef]
- Firon, A.; Tazi, A.; Da Cunha, V.; Brinster, S.; Sauvage, E.; Dramsi, S.; Golenbock, D.T.; Glaser, P.; Poyart, C.; Trieu-Cuot, P. The Abi-domain protein Abx1 interacts with the CovS histidine kinase to control virulence gene expression in group B Streptococcus. PLoS Pathog. 2013, 9, e1003179. [Google Scholar] [CrossRef]
- Gendrin, C.; Lembo, A.; Whidbey, C.; Burnside, K.; Berry, J.; Ngo, L.; Banerjee, A.; Xue, L.; Arrington, J.; Doran, K.S.; Tao, W.A.; et al. The sensor histidine kinase RgfC affects group B streptococcal virulence factor expression independent of its response regulator RgfA. Infect. Immun. 2015, 83, 1078–1088. [Google Scholar] [CrossRef]
- He, Y.W.; Wang, C.; Zhou, L.; Song, H.; Dow, J.M.; Zhang, L.H. Dual signaling functions of the hybrid sensor kinase RpfC of Xanthomonas campestris involve either phosphorelay or receiver domain-protein interaction. J. Biol. Chem. 2006, 281, 33414–33421. [Google Scholar] [CrossRef]
- Reisinger, S.J.; Huntwork, S.; Viollier, P.H.; Ryan, K.R. DivL performs critical cell cycle functions in Caulobacter crescentus independent of kinase activity. J. Bacteriol. 2007, 189, 8308–8320. [Google Scholar] [CrossRef]
- Gao, R.; Stock, A.M. Biological insights from structures of two-component proteins. Annu. Rev. Microbiol. 2009, 63, 133–154. [Google Scholar] [CrossRef]
- Kong, W.; Chen, L.; Zhao, J.; Shen, T.; Surette, M.; Shen, L.; Duan, K. Hybrid sensor kinase PA1611 in Pseudomonas aeruginosa regulates transitions between acute and chronic infection through direct interaction with RetS. Mol. Microbiol. 2013, 88, 784–797. [Google Scholar] [CrossRef]
- Chen, L.; Duan, K. A PhoPQ-regulated ABC transporter system exports Tetracycline in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2016, 60, 3016–3024. [Google Scholar] [CrossRef]
- Wilton, M.; Charron-Mazenod, L.; Moore, R.; Lewenza, S. Extracellular DNA acidifies biofilms and Induces aminoglycoside resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2016, 60, 544–553. [Google Scholar] [CrossRef]
- McPhee, J.B.; Bains, M.; Winsor, G.; Lewenza, S.; Kwasnicka, A.; Brazas, M.D.; Brinkman, F.S.; Hancock, R.E. Contribution of the PhoP-PhoQ and PmrA-PmrB two-component regulatory systems to Mg2+-induced gene regulation in Pseudomonas aeruginosa. J. Bacteriol. 2006, 188, 3995–4006. [Google Scholar] [CrossRef]
- Jochumsen, N.; Marvig, R.L.; Damkiaer, S.; Jensen, R.L.; Paulander, W.; Molin, S.; Jelsbak, L.; Folkesson, A. The evolution of antimicrobial peptide resistance in Pseudomonas aeruginosa is shaped by strong epistatic interactions. Nat. Commun. 2016, 7, 13002. [Google Scholar] [CrossRef]
- Moskowitz, S.M.; Ernst, R.K.; Miller, S.I. PmrAB, a two-component regulatory system of Pseudomonas aeruginosa that modulates resistance to cationic antimicrobial peptides and addition of aminoarabinose to lipid A. J. Bacteriol. 2004, 186, 575–579. [Google Scholar] [CrossRef]
- Barbosa, C.; Trebosc, V.; Kemmer, C.; Rosenstiel, P.; Beardmore, R.; Schulenburg, H.; Jansen, G. Alternative evolutionary paths to bacterial antibiotic resistance cause distinct collateral effects. Mol. Biol. Evol. 2017, 34, 2229–2244. [Google Scholar] [CrossRef]
- Tian, Z.X.; Yi, X.X.; Cho, A.; O’Gara, F.; Wang, Y.P. CpxR Activates MexAB-OprM Efflux Pump Expression and Enhances Antibiotic Resistance in Both Laboratory and Clinical nalB-Type Isolates of Pseudomonas aeruginosa. PLoS Pathog. 2016, 12, e1005932. [Google Scholar] [CrossRef]
- Fernandez, L.; Gooderham, W.J.; Bains, M.; McPhee, J.B.; Wiegand, I.; Hancock, R.E. Adaptive resistance to the “last hope” antibiotics polymyxin B and colistin in Pseudomonas aeruginosa is mediated by the novel two-component regulatory system ParR-ParS. Antimicrob. Agents Chemother. 2010, 54, 3372–3382. [Google Scholar] [CrossRef]
- Muller, C.; Plesiat, P.; Jeannot, K. A two-component regulatory system interconnects resistance to polymyxins, aminoglycosides, fluoroquinolones, and beta-lactams in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2011, 55, 1211–1221. [Google Scholar] [CrossRef]
- Rasamiravaka, T.; Labtani, Q.; Duez, P.; El Jaziri, M. The formation of biofilms by Pseudomonas aeruginosa: A review of the natural and synthetic compounds interfering with control mechanisms. BioMed. Res. Int. 2015, 2015, 759348. [Google Scholar] [CrossRef]
- Mikkelsen, H.; Ball, G.; Giraud, C.; Filloux, A. Expression of Pseudomonas aeruginosa cupD fimbrial genes is antagonistically controlled by RcsB and the EAL-containing PvrR response regulators. PLoS ONE 2009, 4, e6018. [Google Scholar] [CrossRef]
- Mikkelsen, H.; Hui, K.; Barraud, N.; Filloux, A. The pathogenicity island encoded PvrSR/RcsCB regulatory network controls biofilm formation and dispersal in Pseudomonas aeruginosa PA14. Mol. Microbiol. 2013, 89, 450–463. [Google Scholar] [CrossRef]
- Drenkard, E.; Ausubel, F.M. Pseudomonas biofilm formation and antibiotic resistance are linked to phenotypic variation. Nature 2002, 416, 740–743. [Google Scholar] [CrossRef]
- Sivaneson, M.; Mikkelsen, H.; Ventre, I.; Bordi, C.; Filloux, A. Two-component regulatory systems in Pseudomonas aeruginosa: An intricate network mediating fimbrial and efflux pump gene expression. Mol. Microbiol. 2011, 79, 1353–1366. [Google Scholar] [CrossRef]
- Lau, C.H.; Fraud, S.; Jones, M.; Peterson, S.N.; Poole, K. Mutational activation of the AmgRS two-component system in aminoglycoside-resistant Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2013, 57, 2243–2251. [Google Scholar] [CrossRef]
- Lau, C.H.; Krahn, T.; Gilmour, C.; Mullen, E.; Poole, K. AmgRS-mediated envelope stress-inducible expression of the mexXY multidrug efflux operon of Pseudomonas aeruginosa. Microbiologyopen 2015, 4, 121–135. [Google Scholar] [CrossRef]
- Fruci, M.; Poole, K. Aminoglycoside-inducible expression of the mexAB-oprM multidrug efflux operon in Pseudomonas aeruginosa: Involvement of the envelope stress-responsive AmgRS two-component system. PLoS ONE 2018, 13, e0205036. [Google Scholar] [CrossRef]
- Petrova, O.E.; Sauer, K. A novel signaling network essential for regulating Pseudomonas aeruginosa biofilm development. PLoS Pathog. 2009, 5, e1000668. [Google Scholar] [CrossRef]
- Hsu, J.L.; Chen, H.C.; Peng, H.L.; Chang, H.Y. Characterization of the histidine-containing phosphotransfer protein B-mediated multistep phosphorelay system in Pseudomonas aeruginosa PAO1. J. Biol. Chem. 2008, 283, 9933–9944. [Google Scholar] [CrossRef]
- Bhuwan, M.; Lee, H.J.; Peng, H.L.; Chang, H.Y. Histidine-containing phosphotransfer protein-B (HptB) regulates swarming motility through partner-switching system in Pseudomonas aeruginosa PAO1 strain. J. Biol. Chem. 2012, 287, 1903–1914. [Google Scholar] [CrossRef]
- Kulasekara, H.D.; Ventre, I.; Kulasekara, B.R.; Lazdunski, A.; Filloux, A.; Lory, S. A novel two-component system controls the expression of Pseudomonas aeruginosa fimbrial cup genes. Mol. Microbiol. 2005, 55, 368–380. [Google Scholar] [CrossRef]
- Beaudoin, T.; Zhang, L.; Hinz, A.J.; Parr, C.J.; Mah, T.F. The biofilm-specific antibiotic resistance gene ndvB is important for expression of ethanol oxidation genes in Pseudomonas aeruginosa biofilms. J. Bacteriol. 2012, 194, 3128–3136. [Google Scholar] [CrossRef]
- Blus-Kadosh, I.; Zilka, A.; Yerushalmi, G.; Banin, E. The effect of pstS and phoB on quorum sensing and swarming motility in Pseudomonas aeruginosa. PLoS ONE 2013, 8, e74444. [Google Scholar] [CrossRef]
- Luo, Y.; Zhao, K.; Baker, A.E.; Kuchma, S.L.; Coggan, K.A.; Wolfgang, M.C.; Wong, G.C.; O’Toole, G.A. A Hierarchical Cascade of Second Messengers Regulates Pseudomonas aeruginosa Surface Behaviors. mBio 2015, 6, e02456-14. [Google Scholar] [CrossRef]
- Fulcher, N.B.; Holliday, P.M.; Klem, E.; Cann, M.J.; Wolfgang, M.C. The Pseudomonas aeruginosa Chp chemosensory system regulates intracellular cAMP levels by modulating adenylate cyclase activity. Mol. Microbiol. 2010, 76, 889–904. [Google Scholar] [CrossRef]
- Intile, P.J.; Diaz, M.R.; Urbanowski, M.L.; Wolfgang, M.C.; Yahr, T.L. The AlgZR two-component system recalibrates the RsmAYZ posttranscriptional regulatory system to inhibit expression of the Pseudomonas aeruginosa type III secretion system. J. Bacteriol. 2014, 196, 357–366. [Google Scholar] [CrossRef]
- Zamorano, L.; Moya, B.; Juan, C.; Mulet, X.; Blazquez, J.; Oliver, A. The Pseudomonas aeruginosa CreBC two-component system plays a major role in the response to beta-lactams, fitness, biofilm growth, and global regulation. Antimicrob. Agents Chemother. 2014, 58, 5084–5095. [Google Scholar] [CrossRef]
- Vasil, M.L.; Ochsner, U.A. The response of Pseudomonas aeruginosa to iron: Genetics, biochemistry and virulence. Mol. Microbiol. 1999, 34, 399–413. [Google Scholar] [CrossRef]
- Dasgupta, N.; Wolfgang, M.C.; Goodman, A.L.; Arora, S.K.; Jyot, J.; Lory, S.; Ramphal, R. A four-tiered transcriptional regulatory circuit controls flagellar biogenesis in Pseudomonas aeruginosa. Mol. Microbiol. 2003, 50, 809–824. [Google Scholar] [CrossRef]
- Ryan, R.P.; Fouhy, Y.; Garcia, B.F.; Watt, S.A.; Niehaus, K.; Yang, L.; Tolker-Nielsen, T.; Dow, J.M. Interspecies signalling via the Stenotrophomonas maltophilia diffusible signal factor influences biofilm formation and polymyxin tolerance in Pseudomonas aeruginosa. Mol. Microbiol. 2008, 68, 75–86. [Google Scholar] [CrossRef]
- Perron, K.; Caille, O.; Rossier, C.; Van Delden, C.; Dumas, J.L.; Kohler, T. CzcR-CzcS, a two-component system involved in heavy metal and carbapenem resistance in Pseudomonas aeruginosa. J. Biol. Chem. 2004, 279, 8761–8768. [Google Scholar] [CrossRef]
- Caille, O.; Rossier, C.; Perron, K. A copper-activated two-component system interacts with zinc and imipenem resistance in Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 4561–4568. [Google Scholar] [CrossRef]
- Laskowski, M.A.; Osborn, E.; Kazmierczak, B.I. A novel sensor kinase-response regulator hybrid regulates type III secretion and is required for virulence in Pseudomonas aeruginosa. Mol. Microbiol. 2004, 54, 1090–1103. [Google Scholar] [CrossRef]
- Kreamer, N.N.; Costa, F.; Newman, D.K. The ferrous iron-responsive BqsRS two-component system activates genes that promote cationic stress tolerance. mBio 2015, 6, e02549. [Google Scholar] [CrossRef]
- Sage, A.E.; Proctor, W.D.; Phibbs, P.V., Jr. A two-component response regulator, gltR, is required for glucose transport activity in Pseudomonas aeruginosa PAO1. J. Bacteriol. 1996, 178, 6064–6066. [Google Scholar] [CrossRef]
- Wolfgang, M.C.; Lee, V.T.; Gilmore, M.E.; Lory, S. Coordinate regulation of bacterial virulence genes by a novel adenylate cyclase-dependent signaling pathway. Dev. Cell 2003, 4, 253–263. [Google Scholar] [CrossRef]
- Huangyutitham, V.; Guvener, Z.T.; Harwood, C.S. Subcellular clustering of the phosphorylated WspR response regulator protein stimulates its diguanylate cyclase activity. mBio 2013, 4, e00242-13. [Google Scholar] [CrossRef]
- Borlee, B.R.; Goldman, A.D.; Murakami, K.; Samudrala, R.; Wozniak, D.J.; Parsek, M.R. Pseudomonas aeruginosa uses a cyclic-di-GMP-regulated adhesin to reinforce the biofilm extracellular matrix. Mol. Microbiol. 2010, 75, 827–842. [Google Scholar] [CrossRef]
- Hickman, J.W.; Tifrea, D.F.; Harwood, C.S. A chemosensory system that regulates biofilm formation through modulation of cyclic diguanylate levels. Proc. Natl. Acad. Sci. USA 2005, 102, 14422–14427. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ha, U.; Zeng, L.; Jin, S. Regulation of membrane permeability by a two-component regulatory system in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2003, 47, 95–101. [Google Scholar] [CrossRef]
- Giraud, C.; Bernard, C.S.; Calderon, V.; Yang, L.; Filloux, A.; Molin, S.; Fichant, G.; Bordi, C.; de Bentzmann, S. The PprA-PprB two-component system activates CupE, the first non-archetypal Pseudomonas aeruginosa chaperone-usher pathway system assembling fimbriae. Environ. Microbiol. 2011, 13, 666–683. [Google Scholar] [CrossRef]
- De Bentzmann, S.; Giraud, C.; Bernard, C.S.; Calderon, V.; Ewald, F.; Plesiat, P.; Nguyen, C.; Grunwald, D.; Attree, I.; Jeannot, K.; et al. Unique biofilm signature, drug susceptibility and decreased virulence in Drosophila through the Pseudomonas aeruginosa two-component system PprAB. PLoS Pathog. 2012, 8, e1003052. [Google Scholar] [CrossRef]
- Hurley, B.P.; Goodman, A.L.; Mumy, K.L.; Murphy, P.; Lory, S.; McCormick, B.A. The two-component sensor response regulator RoxS/RoxR plays a role in Pseudomonas aeruginosa interactions with airway epithelial cells. Microbes Infect. 2010, 12, 190–198. [Google Scholar] [CrossRef]
- Kilmury, S.L.N.; Burrows, L.L. The Pseudomonas aeruginosa PilSR two-component system regulates both twitching and swimming motilities. mBio 2018, 9, e01310-18. [Google Scholar] [CrossRef]
- Beckmann, C.; Brittnacher, M.; Ernst, R.; Mayer-Hamblett, N.; Miller, S.I.; Burns, J.L. Use of phage display to identify potential Pseudomonas aeruginosa gene products relevant to early cystic fibrosis airway infections. Infect. Immun. 2005, 73, 444–452. [Google Scholar] [CrossRef]
- Yang, Z.; Lu, C.D. Functional genomics enables identification of genes of the arginine transaminase pathway in Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 3945–3953. [Google Scholar] [CrossRef]
- Li, W.; Lu, C.D. Regulation of carbon and nitrogen utilization by CbrAB and NtrBC two-component systems in Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 5413–5420. [Google Scholar] [CrossRef]
- Valentini, M.; Storelli, N.; Lapouge, K. Identification of C(4)-dicarboxylate transport systems in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2011, 193, 4307–4316. [Google Scholar] [CrossRef]
- Chand, N.S.; Clatworthy, A.E.; Hung, D.T. The two-component sensor KinB acts as a phosphatase to regulate Pseudomonas aeruginosa Virulence. J. Bacteriol. 2012, 194, 6537–6547. [Google Scholar] [CrossRef]
- Tatke, G.; Kumari, H.; Silva-Herzog, E.; Ramirez, L.; Mathee, K. Pseudomonas aeruginosa MifS-MifR two-component system is specific for alpha-ketoglutarate utilization. PLoS ONE 2015, 10, e0129629. [Google Scholar] [CrossRef]
- Srinivasan, V.B.; Vaidyanathan, V.; Mondal, A.; Rajamohan, G. Role of the two component signal transduction system CpxAR in conferring cefepime and chloramphenicol resistance in Klebsiella pneumoniae NTUH-K2044. PLoS ONE 2012, 7, e33777. [Google Scholar] [CrossRef]
- Raivio, T.L. Envelope stress responses and Gram-negative bacterial pathogenesis. Mol. Microbiol. 2005, 56, 1119–1128. [Google Scholar] [CrossRef]
- Cannatelli, A.; D’Andrea, M.M.; Giani, T.; Di Pilato, V.; Arena, F.; Ambretti, S.; Gaibani, P.; Rossolini, G.M. In vivo emergence of colistin resistance in Klebsiella pneumoniae producing KPC-type carbapenemases mediated by insertional inactivation of the PhoQ/PhoP mgrB regulator. Antimicrob. Agents Chemother. 2013, 57, 5521–5526. [Google Scholar] [CrossRef]
- Jayol, A.; Nordmann, P.; Brink, A.; Poirel, L. Heteroresistance to colistin in Klebsiella pneumoniae associated with alterations in the PhoPQ regulatory system. Antimicrob. Agents Chemother. 2015, 59, 2780–2784. [Google Scholar] [CrossRef]
- Lippa, A.M.; Goulian, M. Feedback inhibition in the PhoQ/PhoP signaling system by a membrane peptide. PLoS Genet. 2009, 5, e1000788. [Google Scholar] [CrossRef]
- Sperandio, V.; Torres, A.G.; Kaper, J.B. Quorum sensing Escherichia coli regulators B and C (QseBC): A novel two-component regulatory system involved in the regulation of flagella and motility by quorum sensing in E. coli. Mol. Microbiol. 2002, 43, 809–821. [Google Scholar] [CrossRef]
- Lai, Y.C.; Lin, G.T.; Yang, S.L.; Chang, H.Y.; Peng, H.L. Identification and characterization of KvgAS, a two-component system in Klebsiella pneumoniae CG43. Fems Microbiol. Lett. 2003, 218, 121–126. [Google Scholar] [CrossRef]
- Lin, C.T.; Huang, T.Y.; Liang, W.C.; Peng, H.L. Homologous response regulators KvgA, KvhA and KvhR regulate the synthesis of capsular polysaccharide in Klebsiella pneumoniae CG43 in a coordinated manner. J. Biochem. 2006, 140, 429–438. [Google Scholar] [CrossRef]
- Lin, C.T.; Peng, H.L. Regulation of the homologous two-component systems KvgAS and KvhAS in Klebsiella pneumoniae CG43. J. Biochem. 2006, 140, 639–648. [Google Scholar] [CrossRef]
- Cannatelli, A.; Di Pilato, V.; Giani, T.; Arena, F.; Ambretti, S.; Gaibani, P.; D’Andrea, M.M.; Rossolini, G.M. In vivo evolution to colistin resistance by PmrB sensor kinase mutation in KPC-producing Klebsiella pneumoniae is associated with low-dosage colistin treatment. Antimicrob. Agents Chemother. 2014, 58, 4399–4403. [Google Scholar] [CrossRef]
- Su, K.; Zhou, X.; Luo, M.; Xu, X.; Liu, P.; Li, X.; Xue, J.; Chen, S.; Xu, W.; Li, Y.; Qiu, J. Genome-wide identification of genes regulated by RcsA, RcsB, and RcsAB phosphorelay regulators in Klebsiella pneumoniae NTUH-K2044. Microb. Pathog. 2018, 123, 36–41. [Google Scholar] [CrossRef]
- Nixon, B.T.; Ronson, C.W.; Ausubel, F.M. Two-component regulatory systems responsive to environmental stimuli share strongly conserved domains with the nitrogen assimilation regulatory genes ntrB and ntrC. Proc. Natl. Acad. Sci. USA 1986, 83, 7850–7854. [Google Scholar] [CrossRef]
- Hsing, W.H.; Russo, F.D.; Bernd, K.K.; Silhavy, T.J. Mutations that alter the kinase and phosphatase activities of the two-component sensor EnvZ. J. Bacteriol. 1998, 180, 4538–4546. [Google Scholar]
- Lin, T.H.; Chen, Y.; Kuo, J.T.; Lai, Y.C.; Wu, C.C.; Huang, C.F.; Lin, C.T. Phosphorylated OmpR Is required for type 3 fimbriae expression in Klebsiella pneumoniae under hypertonic conditions. Front. Microbiol. 2018, 9, 2405. [Google Scholar] [CrossRef]
- Zahid, N.; Zulfiqiar, S.; Shakoori, A.R. Functional analysis of cus operon promoter of Klebsiella pneumoniae using E. coli lacZ assay. Gene 2012, 495, 81–88. [Google Scholar] [CrossRef]
- Chaturvedi, K.S.; Henderson, J.P. Pathogenic adaptations to host-derived antibacterial copper. Front. Cell. Infect. Microbiol. 2014, 4, 3. [Google Scholar] [CrossRef]
- Hanczvikkel, A.; Fuzi, M.; Ungvari, E.; Toth, A. Transmissible silver resistance readily evolves in high-risk clone isolates of Klebsiella pneumoniae. Acta Microbiol. Immunol. Hung 2018, 65, 387–403. [Google Scholar] [CrossRef]
- Goulian, M. Two-component signaling circuit structure and properties. Curr. Opin. Microbiol. 2010, 13, 184–189. [Google Scholar] [CrossRef]
- Pfluger-Grau, K.; Gorke, B. Regulatory roles of the bacterial nitrogen-related phosphotransferase system. Curr. Trends Microbiol. 2010, 18, 205–214. [Google Scholar] [CrossRef]
- Zhou, L.; Lei, X.H.; Bochner, B.R.; Wanner, B.L. Phenotype microarray analysis of Escherichia coli K-12 mutants with deletions of all two-component systems. J. Bacteriol. 2003, 185, 4956–4972. [Google Scholar] [CrossRef]
- Nagakubo, S.; Nishino, K.; Hirata, T.; Yamaguchi, A. The putative response regulator BaeR stimulates multidrug resistance of Escherichia coli via a novel multidrug exporter system, MdtABC. J. Bacteriol. 2002, 184, 4161–4167. [Google Scholar] [CrossRef]
- Perrenoud, A.; Sauer, U. Impact of global transcriptional regulation by ArcA, ArcB, Cra, Crp, Cya, Fnr, and Mlc on glucose catabolism in Escherichia coli. J. Bacteriol. 2005, 187, 3171–3179. [Google Scholar] [CrossRef]
- Lin, J.T.; Goldman, B.S.; Stewart, V. Structures of genes nasA and nasB, encoding assimilatory nitrate and nitrite reductases in Klebsiella Pneumoniae M5al. J. Bacteriol. 1993, 175, 2370–2378. [Google Scholar] [CrossRef]
- Moreno-Vivian, C.; Cabello, P.; Martinez-Luque, M.; Blasco, R.; Castillo, F. Prokaryotic nitrate reduction: Molecular properties and functional distinction among bacterial nitrate reductases. J. Bacteriol. 1999, 181, 6573–6584. [Google Scholar]
- Verhamme, D.T.; Postma, P.W.; Crielaard, W.; Hellingwerf, K.J. Cooperativity in signal transfer through the Uhp system of Escherichia coli. J. Bacteriol. 2002, 184, 4205–4210. [Google Scholar] [CrossRef]
- Castaneda-Garcia, A.; Blazquez, J.; Rodriguez-Rojas, A. Molecular mechanisms and clinical impact of acquired and intrinsic fosfomycin resistance. Antibiotics (Basel) 2013, 2, 217–236. [Google Scholar] [CrossRef]
- Luo, S.C.; Lou, Y.C.; Rajasekaran, M.; Chang, Y.W.; Hsiao, C.D.; Chen, C. Structural basis of a physical blockage mechanism for the interaction of response regulator PmrA with connector protein PmrD from Klebsiella pneumoniae. J. Biol. Chem. 2013, 288, 25551–25561. [Google Scholar] [CrossRef]
- Nishino, K.; Yamaguchi, A. EvgA of the two-component signal transduction system modulates production of the YhiUV multidrug transporter in Escherichia coli. J. Bacteriol. 2002, 184, 2319–2323. [Google Scholar] [CrossRef]
- Chen, Y.M.; Backman, K.; Magasanik, B. Characterization of a Gene, glnl, the Product of Which Is Involved in the Regulation of Nitrogen-Utilization in Escherichia coli. J. Bacteriol. 1982, 150, 214–220. [Google Scholar]
- Claveriemartin, F.; Magasanik, B. Positive and negative effects of DNA bending on activation of transcription from a distant site. J. Mol. Biol. 1992, 227, 996–1008. [Google Scholar] [CrossRef]
- Sallai, L.; Tucker, P.A. Crystal structure of the central and C-terminal domain of the sigma(54)-activator ZraR. J. Struct. Biol. 2005, 151, 160–170. [Google Scholar] [CrossRef]
- Scheu, P.D.; Witan, J.; Rauschmeier, M.; Graf, S.; Liao, Y.F.; Ebert-Jung, A.; Basche, T.; Erker, W.; Unden, G. CitA/CitB two-component system regulating citrate fermentation in Escherichia coli and its relation to the DcuS/DcuR system in vivo. J. Bacteriol. 2012, 194, 636–645. [Google Scholar] [CrossRef]
- Meyer, M.; Dimroth, P.; Bott, M. Catabolite repression of the citrate fermentation genes in Klebsiella pneumoniae: Evidence for involvement of the cyclic AMP receptor protein. J. Bacteriol. 2001, 183, 5248–5256. [Google Scholar] [CrossRef]
- Cheng, Y.H.; Lin, T.L.; Lin, Y.T.; Wang, J.T. Amino acid substitutions of CrrB responsible for resistance to colistin through CrrC in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2016, 60, 3709–3716. [Google Scholar] [CrossRef]
- Lean, S.S.; Yeo, C.C.; Suhaili, Z.; Thong, K.L. Comparative genomics of two ST 195 carbapenem-resistant Acinetobacter baumannii with different susceptibility to polymyxin revealed underlying resistance mechanism. Front. Microbiol. 2015, 6, 1445. [Google Scholar] [CrossRef]
- Jaidane, N.; Naas, T.; Mansour, W.; Radhia, B.B.; Jerbi, S.; Boujaafar, N.; Bouallegue, O.; Bonnin, R.A. Genomic analysis of in vivo acquired resistance to colistin and rifampicin in Acinetobacter baumannii. Int. J. Antimicrob. Agents 2018, 51, 266–269. [Google Scholar] [CrossRef]
- Richmond, G.E.; Evans, L.P.; Anderson, M.J.; Wand, M.E.; Bonney, L.C.; Ivens, A.; Chua, K.L.; Webber, M.A.; Sutton, J.M.; Peterson, M.L.; et al. The Acinetobacter baumannii two-component system AdeRS regulates genes required for multidrug efflux, biofilm formation, and virulence in a strain-specific manner. mBio 2016, 7, e00430-16. [Google Scholar] [CrossRef]
- Lin, M.F.; Lin, Y.Y.; Yeh, H.W.; Lan, C.Y. Role of the BaeSR two-component system in the regulation of Acinetobacter baumannii adeAB genes and its correlation with tigecycline susceptibility. BMC Microbiol. 2014, 14, 119. [Google Scholar] [CrossRef]
- Lin, M.F.; Lin, Y.Y.; Lan, C.Y. The role of the two-component system BaeSR in disposing chemicals through regulating transporter systems in Acinetobacter baumannii. PLoS ONE 2015, 10, e0132843. [Google Scholar] [CrossRef]
- Tomaras, A.P.; Flagler, M.J.; Dorsey, C.W.; Gaddy, J.A.; Actis, L.A. Characterization of a two-component regulatory system from Acinetobacter baumannii that controls biofilm formation and cellular morphology. Microbiology 2008, 154, 3398–3409. [Google Scholar] [CrossRef]
- Liou, M.L.; Soo, P.C.; Ling, S.R.; Kuo, H.Y.; Tang, C.Y.; Chang, K.C. The sensor kinase BfmS mediates virulence in Acinetobacter baumannii. J. Microbiol. Immunol. Infect. 2014, 47, 275–281. [Google Scholar] [CrossRef]
- Russo, T.A.; Manohar, A.; Beanan, J.M.; Olson, R.; MacDonald, U.; Graham, J.; Umland, T.C. The response regulator BfmR is a potential drug target for Acinetobacter baumannii. mSphere 2016, 1, e00082-16. [Google Scholar] [CrossRef]
- Cerqueira, G.M.; Kostoulias, X.; Khoo, C.; Aibinu, I.; Qu, Y.; Traven, A.; Peleg, A.Y. A global virulence regulator in Acinetobacter baumannii and its control of the phenylacetic acid catabolic pathway. J. Infect. Dis. 2014, 210, 46–55. [Google Scholar] [CrossRef]
- Bhuiyan, M.S.; Ellett, F.; Murray, G.L.; Kostoulias, X.; Cerqueira, G.M.; Schulze, K.E.; Mahamad Maifiah, M.H.; Li, J.; Creek, D.J.; Lieschke, G.J.; et al. Acinetobacter baumannii phenylacetic acid metabolism influences infection outcome through a direct effect on neutrophil chemotaxis. Proc. Natl. Acad. Sci. USA 2016, 113, 9599–9604. [Google Scholar] [CrossRef]
- Chen, R.; Lv, R.; Xiao, L.; Wang, M.; Du, Z.; Tan, Y.; Cui, Y.; Yan, Y.; Luo, Y.; Yang, R.; et al. A1S_2811, a CheA/Y-like hybrid two-component regulator from Acinetobacter baumannii ATCC17978, is involved in surface motility and biofilm formation in this bacterium. Microbiologyopen 2017, 6, e00510. [Google Scholar] [CrossRef]
- Samir, R.; Hussein, S.H.; Elhosseiny, N.M.; Khattab, M.S.; Shawky, A.E.; Attia, A.S. Adaptation to potassium-limitation is essential for Acinetobacter baumannii pneumonia pathogenesis. J. Infect. Dis. 2016, 214, 2006–2013. [Google Scholar] [CrossRef]
- Ninfa, A.J.; Magasanik, B. Covalent modification of the glnG product, NRI, by the glnL product, NRII, regulates the transcription of the glnALG operon in Escherichia coli. Proc. Natl. Acad. Sci. USA 1986, 83, 5909–5913. [Google Scholar] [CrossRef]
- Gavigan, J.A.; Marshall, L.M.; Dobson, A.D. Regulation of polyphosphate kinase gene expression in Acinetobacter baumannii 252. Microbiology 1999, 145, 2931–2937. [Google Scholar] [CrossRef]
- Williams, C.L.; Neu, H.M.; Gilbreath, J.J.; Michel, S.L.; Zurawski, D.V.; Merrell, D.S. Copper resistance of the emerging pathogen Acinetobacter baumannii. Appl. Environ. Microbiol. 2016, 82, 6174–6188. [Google Scholar] [CrossRef]
- Tipton, K.A.; Rather, P.N. An OmpR-EnvZ two-component system ortholog regulates phase variation, osmotic tolerance, motility, and virulence in Acinetobacter baumannii Strain AB5075. J. Bacteriol. 2017, 199, e00705-16. [Google Scholar] [CrossRef]
- Mulcahy, H.; Charron-Mazenod, L.; Lewenza, S. Extracellular DNA chelates cations and induces antibiotic resistance in Pseudomonas aeruginosa biofilms. PLoS Pathog. 2008, 4, e1000213. [Google Scholar] [CrossRef]
- McPhee, J.B.; Lewenza, S.; Hancock, R.E. Cationic antimicrobial peptides activate a two-component regulatory system, PmrA-PmrB, that regulates resistance to polymyxin B and cationic antimicrobial peptides in Pseudomonas aeruginosa. Mol. Microbiol. 2003, 50, 205–217. [Google Scholar] [CrossRef]
- Schniederjans, M.; Koska, M.; Häussler, S. Transcriptional and mutational profiling of an aminoglycoside-resistant Pseudomonas aeruginosa small-colony variant. Antimicrob. Agents Chemother. 2017, 61, e01178-17. [Google Scholar] [CrossRef]
- Fothergill, J.L.; Neill, D.R.; Loman, N.; Winstanley, C.; Kadioglu, A. Pseudomonas aeruginosa adaptation in the nasopharyngeal reservoir leads to migration and persistence in the lungs. Nat. Commun. 2014, 5, 4780. [Google Scholar] [CrossRef]
- Bricio-Moreno, L.; Sheridan, V.H.; Goodhead, I.; Armstrong, S.; Wong, J.K.L.; Waters, E.M.; Sarsby, J.; Panagiotou, S.; Dunn, J.; Chakraborty, A.; et al. Evolutionary trade-offs associated with loss of PmrB function in host-adapted Pseudomonas aeruginosa. Nat. Commun. 2018, 9, 2635. [Google Scholar] [CrossRef]
- Nowicki, E.M.; O’Brien, J.P.; Brodbelt, J.S.; Trent, M.S. Extracellular zinc induces phosphoethanolamine addition to Pseudomonas aeruginosa lipid A via the ColRS two-component system. Mol. Microbiol. 2015, 97, 166–178. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Chen, Y.F.; Peng, H.L. Molecular characterization of the PhoPQ-PmrD-PmrAB mediated pathway regulating polymyxin B resistance in Klebsiella pneumoniae CG43. J. Biomed. Sci. 2010, 17, 60. [Google Scholar] [CrossRef]
- Carretero-Ledesma, M.; García-Quintanilla, M.; Martín-Peña, R.; Pulido, M.R.; Pachón, J.; McConnell, M.J. Phenotypic changes associated with Colistin resistance due to Lipopolysaccharide loss in Acinetobacter baumannii. Virulence 2018, 9, 930–942. [Google Scholar] [CrossRef]
- Kim, Y.; Bae, I.K.; Jeong, S.H.; Yong, D.; Lee, K. In vivo selection of pan-drug resistant Acinetobacter baumannii during antibiotic treatment. Yonsei Med. J. 2015, 56, 928–934. [Google Scholar] [CrossRef]
- Lopez-Rojas, R.; Dominguez-Herrera, J.; McConnell, M.J.; Docobo-Perez, F.; Smani, Y.; Fernandez-Reyes, M.; Rivas, L.; Pachon, J. Impaired virulence and in vivo fitness of colistin-resistant Acinetobacter baumannii. J. Infect. Dis. 2011, 203, 545–548. [Google Scholar] [CrossRef]
- Durante-Mangoni, E.; Del Franco, M.; Andini, R.; Bernardo, M.; Giannouli, M.; Zarrilli, R. Emergence of colistin resistance without loss of fitness and virulence after prolonged colistin administration in a patient with extensively drug-resistant Acinetobacter baumannii. Diagn Microbiol. Infect. Dis. 2015, 82, 222–226. [Google Scholar] [CrossRef]
- Lopez-Rojas, R.; Garcia-Quintanilla, M.; Labrador-Herrera, G.; Pachon, J.; McConnell, M.J. Impaired growth under iron-limiting conditions associated with the acquisition of colistin resistance in Acinetobacter baumannii. Int. J. Antimicrob. Agents 2016, 47, 473–477. [Google Scholar] [CrossRef]
- Wand, M.E.; Bock, L.J.; Bonney, L.C.; Sutton, J.M. Retention of virulence following adaptation to colistin in Acinetobacter baumannii reflects the mechanism of resistance. J. Antimicrob. Chemother. 2015, 70, 2209–2216. [Google Scholar] [CrossRef]
- Da Silva, G.J.; Domingues, S. Interplay between colistin resistance, virulence and fitness in Acinetobacter baumannii. Antibiotics 2017, 6, 28. [Google Scholar] [CrossRef]
- Janssen, K.H.; Diaz, M.R.; Golden, M.; Graham, J.W.; Sanders, W.; Wolfgang, M.C.; Yahr, T.L. Functional analyses of the RsmY and RsmZ small noncoding regulatory RNAs in Pseudomonas aeruginosa. J. Bacteriol. 2018, 200, e00736-17. [Google Scholar] [CrossRef]
- Marden, J.N.; Diaz, M.R.; Walton, W.G.; Gode, C.J.; Betts, L.; Urbanowski, M.L.; Redinbo, M.R.; Yahr, T.L.; Wolfgang, M.C. An unusual CsrA family member operates in series with RsmA to amplify post-transcriptional responses in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2013, 110, 15055–15060. [Google Scholar] [CrossRef]
- Llanes, C.; Köhler, T.; Patry, I.; Dehecq, B.; van Delden, C.; Plésiat, P. Role of the MexEF-OprN efflux system in low-level resistance of Pseudomonas aeruginosa to Ciprofloxacin. Antimicrob. Agents Chemother. 2011, 55, 5676–5684. [Google Scholar] [CrossRef]
- Lamarche, M.G.; Déziel, E. MexEF-OprN efflux pump exports the Pseudomonas Quinolone Signal (PQS) precursor HHQ (4-hydroxy-2-heptylquinoline). PLoS ONE 2011, 6, e24310. [Google Scholar] [CrossRef]
- Linares, J.F.; Gustafsson, I.; Baquero, F.; Martinez, J.L. Antibiotics as intermicrobial signaling agents instead of weapons. Proc. Natl. Acad. Sci. USA 2006, 103, 19484–19489. [Google Scholar] [CrossRef] [Green Version]
- Valentini, M.; Filloux, A. Biofilms and Cyclic di-GMP (c-di-GMP) signaling: Lessons from Pseudomonas aeruginosa and other bacteria. J. Biol. Chem. 2016, 291, 12547–12555. [Google Scholar] [CrossRef]
- Ventre, I.; Goodman, A.L.; Vallet-Gely, I.; Vasseur, P.; Soscia, C.; Molin, S.; Bleves, S.; Lazdunski, A.; Lory, S.; Filloux, A. Multiple sensors control reciprocal expression of Pseudomonas aeruginosa regulatory RNA and virulence genes. Proc. Natl. Acad. Sci. USA 2006, 103, 171–176. [Google Scholar] [CrossRef]
- Moscoso, J.A.; Mikkelsen, H.; Heeb, S.; Williams, P.; Filloux, A. The Pseudomonas aeruginosa sensor RetS switches type III and type VI secretion via c-di-GMP signalling. Environ. Microbiol. 2011, 13, 3128–3138. [Google Scholar] [CrossRef]
- Evans, T.J. Small colony variants of Pseudomonas aeruginosa in chronic bacterial infection of the lung in cystic fibrosis. Future Microbiol. 2015, 10, 231–239. [Google Scholar] [CrossRef]
- Kentache, T.; Ben Abdelkrim, A.; Jouenne, T.; De, E.; Hardouin, J. Global dynamic proteome study of a pellicle-forming Acinetobacter baumannii strain. Mol. Cell. Proteomics MCP 2017, 16, 100–112. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Tampakakis, E.; Fuchs, B.B.; Eliopoulos, G.M.; Moellering, R.C., Jr.; Mylonakis, E. Prokaryote-eukaryote interactions identified by using Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2008, 105, 14585–14590. [Google Scholar] [CrossRef]
- Dorsey, C.W.; Tomaras, A.P.; Actis, L.A. Genetic and phenotypic analysis of Acinetobacter baumannii insertion derivatives generated with a transposome system. Appl. Environ. Microbiol. 2002, 68, 6353–6360. [Google Scholar] [CrossRef]
- Suzuki, K.; Wang, X.; Weilbacher, T.; Pernestig, A.-K.; Melefors, O.; Georgellis, D.; Babitzke, P.; Romeo, T. Regulatory circuitry of the CsrA/CsrB and BarA/UvrY systems of Escherichia coli. J. Bacteriol. 2002, 184, 5130–5140. [Google Scholar] [CrossRef]
- Tomenius, H.; Pernestig, A.K.; Mendez-Catala, C.F.; Georgellis, D.; Normark, S.; Melefors, O. Genetic and functional characterization of the Escherichia coli BarA-UvrY two-component system: Point mutations in the HAMP linker of the BarA sensor give a dominant-negative phenotype. J. Bacteriol. 2005, 187, 7317–7324. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Audia, J.P.; Roy, R.N.; Schellhorn, H.E. Transcriptional induction of the conserved alternative sigma factor RpoS in Escherichia coli is dependent on BarA, a probable two-component regulator. Mol. Microbiol. 2000, 37, 371–381. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Schellhorn, H.E. Identification and characterization of hydrogen peroxide-sensitive mutants of Escherichia coli: Genes that require OxyR for expression. J. Bacteriol. 1997, 179, 330–338. [Google Scholar] [CrossRef]
- Michael, B.; Smith, J.N.; Swift, S.; Heffron, F.; Ahmer, B.M. SdiA of Salmonella enterica is a LuxR homolog that detects mixed microbial communities. J. Bacteriol. 2001, 183, 5733–5742. [Google Scholar] [CrossRef]
- Fernando, D.; Kumar, A. Growth phase-dependent expression of RND efflux pump- and outer membrane porin-encoding genes in Acinetobacter baumannii ATCC 19606. J. Antimicrob. Chemother. 2012, 67, 569–572. [Google Scholar] [CrossRef]
- Costa, S.S.; Viveiros, M.; Amaral, L.; Couto, I. Multidrug efflux pumps in Staphylococcus aureus: An update. Open Microbiol. J. 2013, 7, 59–71. [Google Scholar] [CrossRef]
- Ruzin, A.; Keeney, D.; Bradford, P.A. AdeABC multidrug efflux pump is associated with decreased susceptibility to tigecycline in Acinetobacter calcoaceticus-Acinetobacter baumannii complex. J. Antimicrob. Chemother. 2007, 59, 1001–1004. [Google Scholar] [CrossRef]
- Vila, J.; Marti, S.; Sanchez-Cespedes, J. Porins, efflux pumps and multidrug resistance in Acinetobacter baumannii. J. Antimicrob. Chemother. 2007, 59, 1210–1215. [Google Scholar] [CrossRef]
- Sun, J.R.; Chan, M.C.; Chang, T.Y.; Wang, W.Y.; Chiueh, T.S. Overexpression of the adeB gene in clinical isolates of tigecycline-nonsusceptible Acinetobacter baumannii without insertion mutations in adeRS. Antimicrob. Agents Chemother. 2010, 54, 4934–4938. [Google Scholar] [CrossRef]
- Higgins, P.G.; Schneiders, T.; Hamprecht, A.; Seifert, H. In vivo selection of a missense mutation in adeR and conversion of the novel blaOXA-164 gene into blaOXA-58 in carbapenem-resistant Acinetobacter baumannii isolates from a hospitalized patient. Antimicrob. Agents Chemother. 2010, 54, 5021–5027. [Google Scholar] [CrossRef]
- Kröger, C.; Kary, S.C.; Schauer, K.; Cameron, A.D.S. Genetic regulation of virulence and antibiotic resistance in Acinetobacter baumannii. Genes 2016, 8, 12. [Google Scholar] [CrossRef]
- Sun, J.R.; Perng, C.L.; Chan, M.C.; Morita, Y.; Lin, J.C.; Su, C.M.; Wang, W.Y.; Chang, T.Y.; Chiueh, T.S. A truncated AdeS kinase protein generated by ISAba1 insertion correlates with tigecycline resistance in Acinetobacter baumannii. PLoS ONE 2012, 7, e49534. [Google Scholar] [CrossRef]
- Sun, J.R.; Jeng, W.Y.; Perng, C.L.; Yang, Y.S.; Soo, P.C.; Chiang, Y.S.; Chiueh, T.S. Single amino acid substitution Gly186Val in AdeS restores tigecycline susceptibility of Acinetobacter baumannii. J. Antimicrob. Chemother. 2016, 71, 1488–1492. [Google Scholar] [CrossRef]
- Raffa, R.G.; Raivio, T.L. A third envelope stress signal transduction pathway in Escherichia coli. Mol. Microbiol. 2002, 45, 1599–1611. [Google Scholar] [CrossRef]
- Anand, P.; Chandra, N. Characterizing the pocketome of Mycobacterium tuberculosis and application in rationalizing polypharmacological target selection. Sci. Rep. 2014, 4, 6356. [Google Scholar] [CrossRef]
- Ramakrishnan, G.; Chandra, N.R.; Srinivasan, N. Recognizing drug targets using evolutionary information: Implications for repurposing FDA-approved drugs against Mycobacterium tuberculosis H37Rv. Mol. Biosyst. 2015, 11, 3316–3331. [Google Scholar] [CrossRef]
- Zheng, H.; Aleiwi, B.; Ellsworth, E.; Abramovitch, R.B. Inhibition of Mycobacterium tuberculosis DosRST two-component regulatory system signaling by targeting response regulator DNA binding and sensor kinase heme. bioRxivorg 2018. [Google Scholar] [CrossRef]
- Stephenson, K.; Yamaguchi, Y.; Hoch, J.A. The mechanism of action of inhibitors of bacterial two-component signal transduction systems. J. Biol. Chem. 2000, 275, 38900–38904. [Google Scholar] [CrossRef]
- Wilke, K.E.; Francis, S.; Carlson, E.E. Inactivation of multiple bacterial histidine kinases by targeting the ATP-binding domain. ACS Chem. Biol. 2015, 10, 328–335. [Google Scholar] [CrossRef]
- Williams, C.; Cotter, P. Autoregulation is essential for precise temporal and steady-state regulation by the Bordetella BvgAS phosphorelay. J. Bacteriol. 2007, 189, 1974–1982. [Google Scholar] [CrossRef]
- Miyashiro, T.; Goulian, M. High stimulus unmasks positive feedback in an autoregulated bacterial signaling circuit. Proc. Natl. Acad. Sci. USA 2008, 105, 17457–17462. [Google Scholar] [CrossRef] [Green Version]
- Ansaldi, M.; Simon, G.; Lepelletier, M.; Mejean, V. The TorR high-affinity binding site plays a key role in both torR autoregulation and torCAD operon expression in Escherichia coli. J. Bacteriol. 2000, 182, 961–966. [Google Scholar] [CrossRef]
- Svenningsen, S.; Tu, K.; Bassler, B. Gene dosage compensation calibrates four regulatory RNAs to control Vibrio cholerae quorum sensing. EMBO J. 2009, 28, 429–439. [Google Scholar] [CrossRef] [Green Version]
- Hoffer, S.; Westerhoff, H.; Hellingwerf, K.; Postma, P.; Tommassen, J. Autoamplification of a two-component regulatory system results in “learning” behavior. J. Bacteriol. 2001, 183, 4914–4917. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Two-Component System | Confirmed or Predicted Function | Reference(s) |
|---|---|---|
| P. aeruginosa | ||
| PhoQ/PhoP | Regulating ABC transporter system; Resistance to antimicrobial peptides, polymyxins, and aminoglycosides; Regulating virulence, swarming motility and biofilm formation; Mg2+ sensing. | [130,131,132,133] |
| PmrA/PmrB | Activated by low Mg2+ and cationic antimicrobial peptides; Resistance against polymyxin B, colistin and other antimicrobial peptides | [131,134,135] |
| CpxA/CpxR | Role in cell envelope stress response; Activates MexAB-OprM efflux pump expression | [136] |
| CprS/CprR | Role in LPS modification and antimicrobial peptide resistance | [137] |
| ParR/ParS | Role in resistance to colistin and polymyxins; Role in quorum sensing, phenazine production, and motility | [137,138] |
| GacS/GacA | Regulating virulence factors; Biofilm formation; Antibiotic resistance; Motility; Iron metabolism; Type III and type VI secretion | [139] |
| PvrS/PvrR | Regulation of the MexAB-OprM efflux pump; Biofilm formation. Controls of fimbrial genes | [140,141,142,143] |
| RcsC-RcsB | Role in biofilm formation and control of fimbrial genes | [140,141] |
| AmgS-AmgR | Involved in aminoglycoside resistance and cell envelope stress response | [144,145,146] |
| PA1611 | Biofilm formation and virulence regulation | [96,129] |
| BfiS/BfiR | Biofilm maturation | [147] |
| HptB/HsbR | Involved in swarming motility and biofilm formation | [148,149] |
| RocS2/RocA2 | Regulation of fimbrial adhesins and antimicrobial resistance | [143,150] |
| ErbR/EraR | Control of biofilm specific antibiotic resistance | [151] |
| TctE/TctD | Controls expression of tricarboxylic acid (TCA) uptake system | [86] |
| PhoR–PhoB | Plays a role in quorum sensing and swarming motility | [86,152] |
| ChpA/PilG/PilH/ChpB | Regulation of the chemosensory pili (Pil–Chp) system, twitching motility and cAMP levels; Regulates virulence genes | [153,154] |
| FimS (AlgZ)/AlgR | Regulation of virulence; Alginate biosynthesis; Motility; Biofilm formation; Cytotoxicity and type III secretion system expression | [155] |
| ColS/ColR | Polymyxin resistance; Virulence and cell adherence | [20] |
| CreC–CreB | Role in catabolism; Swarming and swimming motility; Antibiotic resistance; Biofilm and global gene regulation | [156] |
| PirR–PirS | Iron acquisition | [157] |
| FleS–FleR | Flagellar motility; Adhesion to mucins | [158] |
| PA1396/PA1397 | Plays a role in interspecies signaling; Responds to diffusible signal factor (DSF); Regulates biofilm formation and antibiotic resistance | [159] |
| CzcS–CzcR | Regulates heavy metal resistance; Controls antibiotic resistance and pathogenicity | [160,161] |
| RetS | Regulates virulence; Biofilm formation; Regulates Type III and VI secretion/cytotoxicity | [99,162] |
| LadS | Regulates virulence; Biofilm formation; Type III secretion/cytotoxicity | [99,101] |
| BqsS/BqrR/CarS/CarR | Biofilm formation; Iron sensing; Antibiotic resistance and cationic stress tolerance. Maintains Ca2+ homeostasis; Regulates pyocyanin secretion; Motility. | [163] |
| PfeS–PfeR | Iron acquisition | [87] |
| CopS–CopR | Tolerance to Cu2+, Zn2+; Imipenem resistance | [161] |
| GtrS/GltR | Regulates glucose transport and Type III secretion system | [164,165] |
| WspE–WspR | Regulates biofilm formation, autoaggregation, and cyclic-di-GMP synthesis | [166,167,168] |
| NarX–NarL | Nitrate sensing and respiration; Biofilm formation; Motility | [84] |
| BfmS/BfmR | Biofilm formation/maintenance | [147] |
| PprA–PprB | Regulates outer membrane permeability; Aminoglycoside resistance; Controls virulence including type III secretion system and biofilm formation | [169,170,171] |
| RoxS/RoxR | Confers cyanide tolerance | [172] |
| PilS–PilR | Involved in regulating the expression of the T4P major subunit PilA; Biofilm formation; Motility; Positively regulates the transcription of flagellar regulatory genes | [173] |
| CbrA–CbrB | Metabolic regulation of carbon and nitrogen utilization. Modulates biofilm formation; Cytotoxicity; Motility; Antibiotic resistance | [89] |
| AruS/AruR | Controls the expression of the arginine transaminase pathway | [174,175] |
| NtrB/NtrC | Responds to cellular nitrogen levels and activates nitrogen scavenging genes | [176] |
| DctB/DctD | Controls the expression of C4-dicarboxylate transporters | [177] |
| KinA/AlgB | Regulates alginate biosynthesis; Regulates virulence | [178] |
| MifS/MifR | Role in biofilm formation and metabolism | [179] |
| K. pneumoniae | ||
| CpxA/CpxR | Sensing extracellular pH and membrane composition; Regulating cell envelope protein folding and protein degradation | [180,181] |
| PhoP/PhoQ | Activates pmrHFIJKLM; Responsible for L-amino arabinose synthesis and polymyxin resistance | [182,183,184] |
| PhoR/PhoB | Phosphate assimilation | [180] |
| QseC/QseB | Involved in regulation of the flagella and motility genes | [185] |
| KvgA/KvgS | Involved in tolerating free radical stresses and sensing iron-limiting conditions | [186] |
| KvhA/KvhS | Regulates capsular polysaccharide synthesis | [187,188] |
| PmrA/PmrB | Regulator of genes for lipopolysaccharide modification | [189] |
| RcsC/RcsB | Involved in the capsular polysaccharide biosynthesis; Type III system; Regulates the production of major pilin protein MrkA; Confers resistance to low pH | [190] |
| EnvZ/OmpR | Senses osmotic signals; Regulates the c-di-GMP signaling pathway; Regulates type III fimbriae and biofilm formation | [191,192,193] |
| CusS/CusR | Induced by Copper and regulates the CusCFBARS efflux system; Tolerance to silver | [194,195,196] |
| KdpD/KdpE | Potassium transporter system | [197,198,199] |
| BaeS/BaeR | Regulates Multidrug efflux pump AdeABC; Regulates Modification of lipopolysaccharides | [199,200] |
| ArcB/ArcA | Involved in modulating the expression of genes encoding for proteins with membrane modification functions and TCA cycle enzymes depending upon oxygen levels. | [199,201] |
| NarX/NarL | Role in nitrate and nitrite reductase synthesis | [202,203] |
| UhpB/UhpA | Role in uptake of hexose phosphates | [199,204,205] |
| EvgS/EvgA | Regulates capsular polysaccharide biosynthesis | [206,207] |
| GlnL/GlnG | Role in glutamate metabolism | [208,209] |
| ZraR/ZraS | Zinc-responsive TCS; Activated under high calcium and iron conditions | [210] |
| CitA/CitB | Regulates citrate metabolism under anaerobic conditions | [211,212] |
| CrrA/CrrB | Involved in polymyxin resistance | [213] |
| A. baumannii | ||
| PmrA/PmrB | Regulates genes involved in lipopolysaccharide modification | [214,215] |
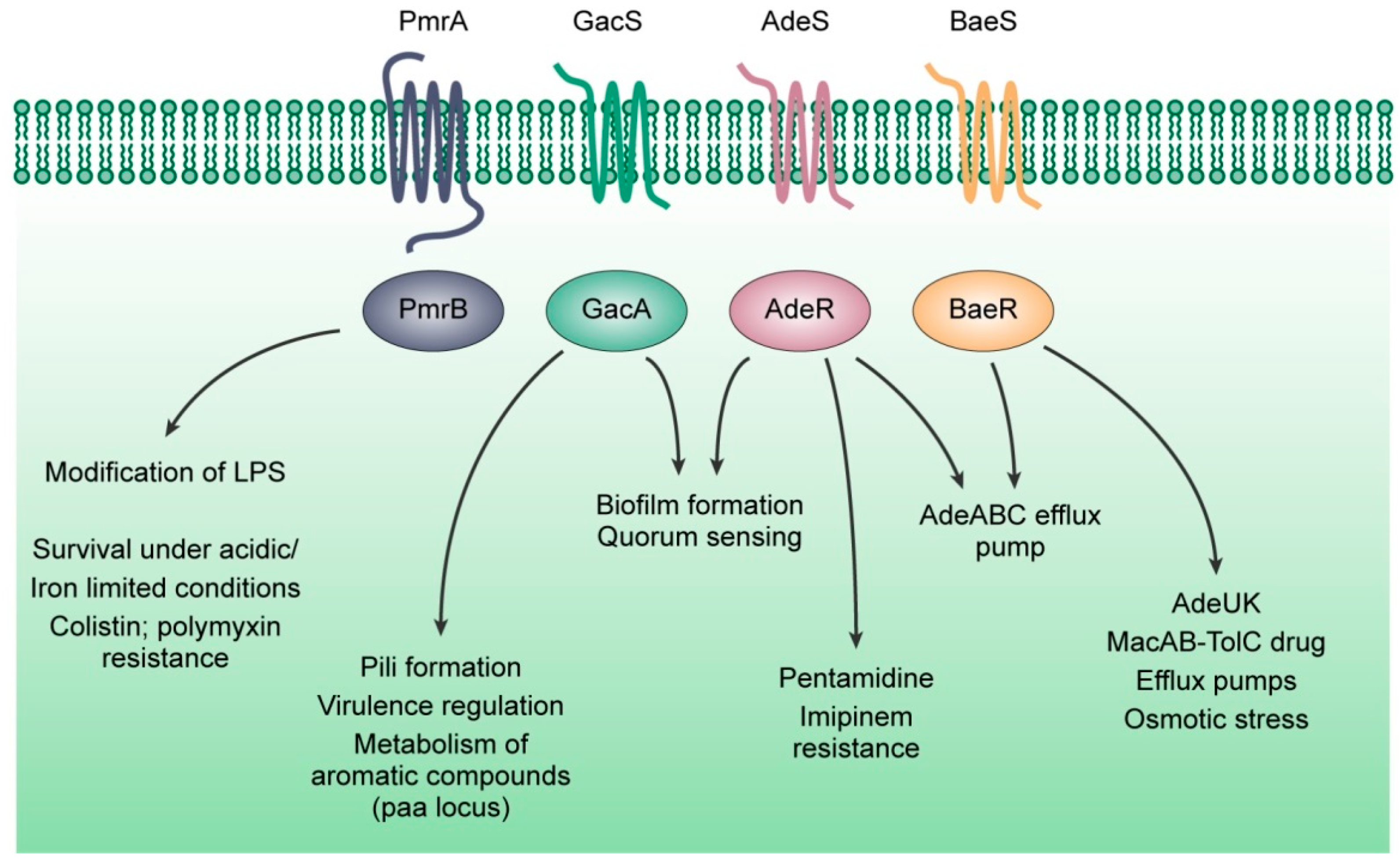
| AdeS/AdeR | Regulates genes encoding the AdeABC pump | [59,216] |
| BaeS/BaeR | Stress response under high osmotic conditions | [60,217,218] |
| BfmS/BfmR | Regulates biofilm formation and antibiotic resistance | [219,220,221] |
| GacS/GacA | Regulates genes associated with pili and biofilm development, motility and resistance against host antimicrobial peptides | [222,223] |
| A1S_2811 | Involved in surface motility and biofilm formation | [224] |
| KdpD/KpdE | Regulates potassium transport | [225] |
| GlnL/GlnG | Involved in nitrogen assimilation | [226] |
| PhoR/PhoB | Regulates phosphate assimilation | [227] |
| CusS/CusR | Senses copper ions and upregulates the expression of an RND family efflux pump that removes copper ions from the cell | [228] |
| OmpR/EnvZ | Regulates virulence; Phase variation; Osmotic tolerance | [229] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhagirath, A.Y.; Li, Y.; Patidar, R.; Yerex, K.; Ma, X.; Kumar, A.; Duan, K. Two Component Regulatory Systems and Antibiotic Resistance in Gram-Negative Pathogens. Int. J. Mol. Sci. 2019, 20, 1781. https://doi.org/10.3390/ijms20071781
Bhagirath AY, Li Y, Patidar R, Yerex K, Ma X, Kumar A, Duan K. Two Component Regulatory Systems and Antibiotic Resistance in Gram-Negative Pathogens. International Journal of Molecular Sciences. 2019; 20(7):1781. https://doi.org/10.3390/ijms20071781
Chicago/Turabian StyleBhagirath, Anjali Y., Yanqi Li, Rakesh Patidar, Katherine Yerex, Xiaoxue Ma, Ayush Kumar, and Kangmin Duan. 2019. "Two Component Regulatory Systems and Antibiotic Resistance in Gram-Negative Pathogens" International Journal of Molecular Sciences 20, no. 7: 1781. https://doi.org/10.3390/ijms20071781
APA StyleBhagirath, A. Y., Li, Y., Patidar, R., Yerex, K., Ma, X., Kumar, A., & Duan, K. (2019). Two Component Regulatory Systems and Antibiotic Resistance in Gram-Negative Pathogens. International Journal of Molecular Sciences, 20(7), 1781. https://doi.org/10.3390/ijms20071781