Genetic Mapping and Molecular Characterization of a Broad-spectrum Phytophthora sojae Resistance Gene in Chinese Soybean


Abstract
:1. Introduction
2. Results
2.1. Xiu94-11 Has Broad-spectrum Resistance to Phytophthora sojae Which is Controlled by a Dominant Single Gene
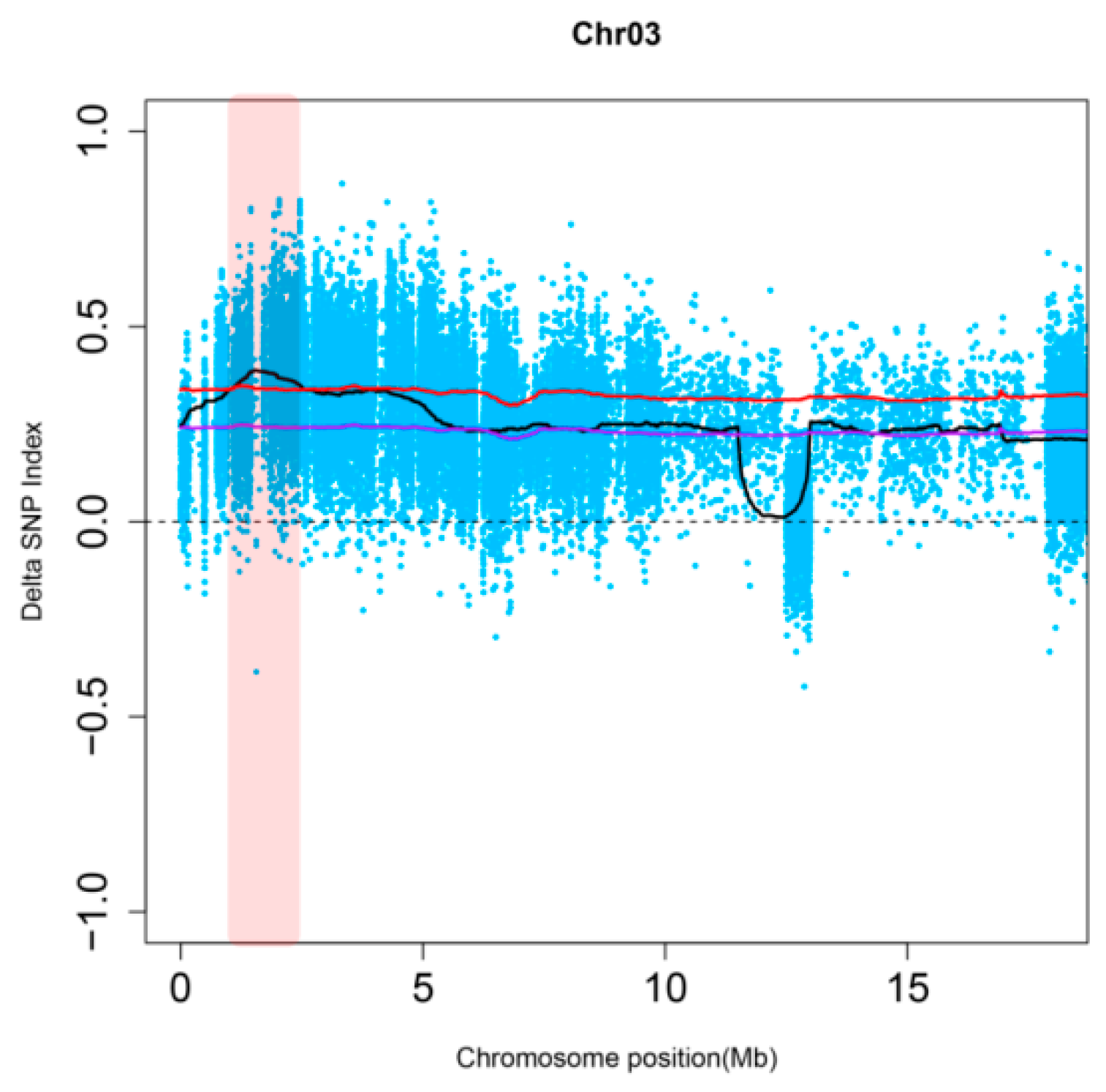
2.2. The Resistance Gene RpsX in Xiu94-11 is Located on Soybean Chromosome 3
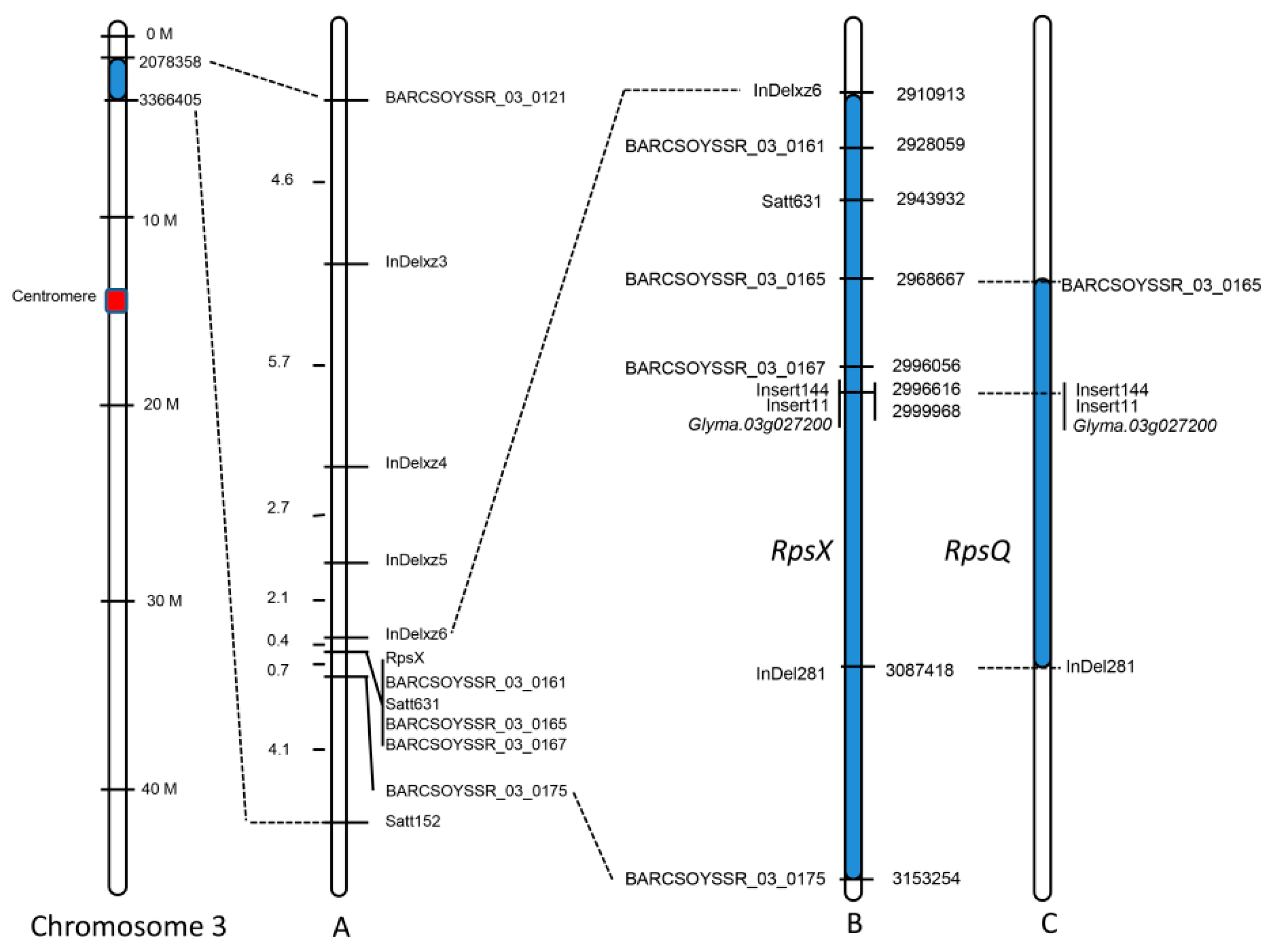
2.3. RpsX Was Finely Mapped to the 242 kb Region on Chromosome 3
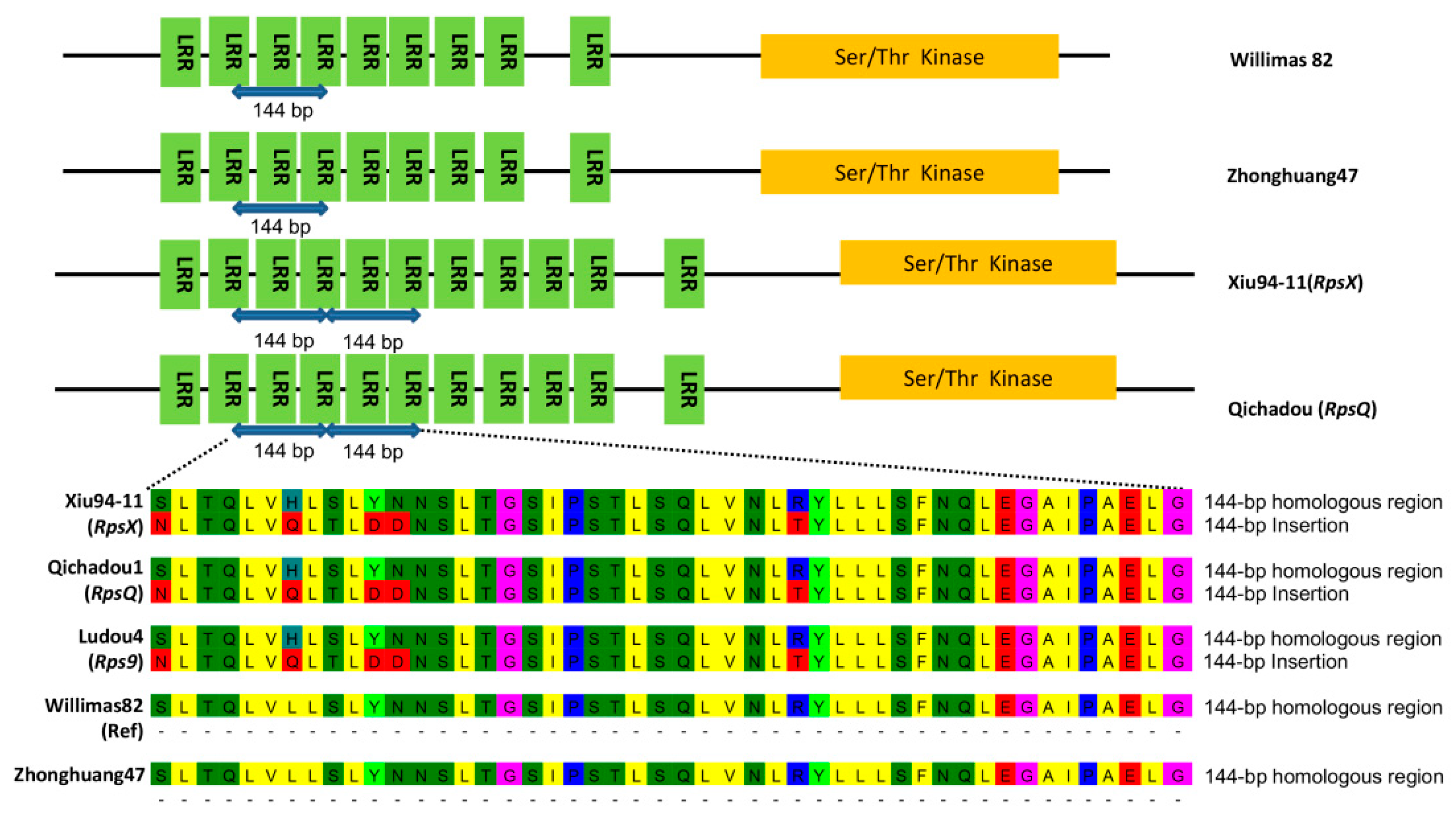
2.4. RpsX and RpsQShared the Same Candidate Gene Model
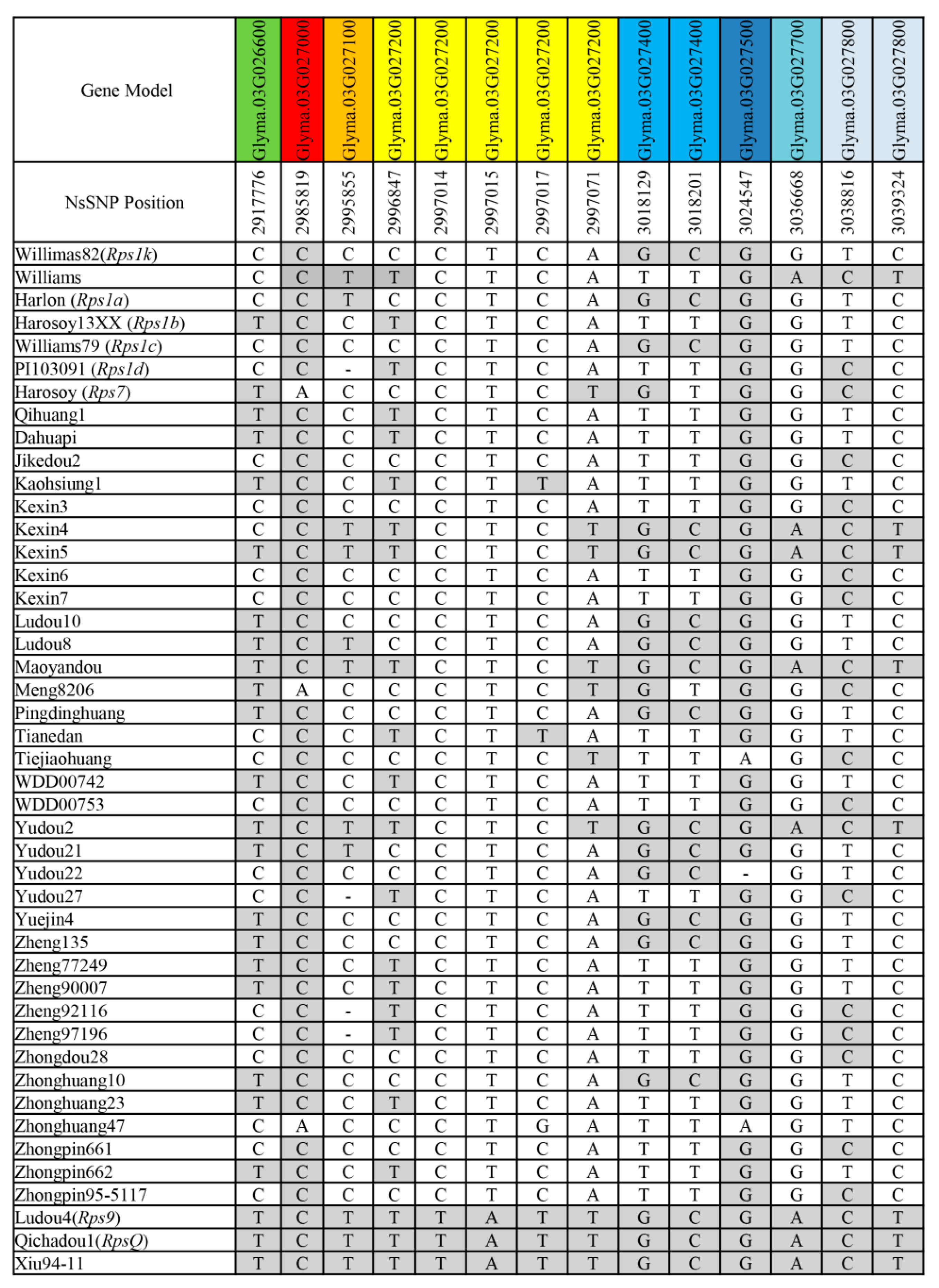
2.5. 144-bp Insertion in LRR Domain is Present in Resistant Haplotype of RpsX Locus
2.6. Developed Marker Insert144 Is Able to Efficiently Detect Resistant Haplotypes at RpsX Locus
3. Discussion
4. Materials and Methods
4.1. Phenotyping for PRR Resistance
4.2. Next-generation Sequencing and QTL-seq Analysis of Resistant and Susceptible Bulks
4.3. Linkage Analysis and Genetic Mapping of the Candidate Region
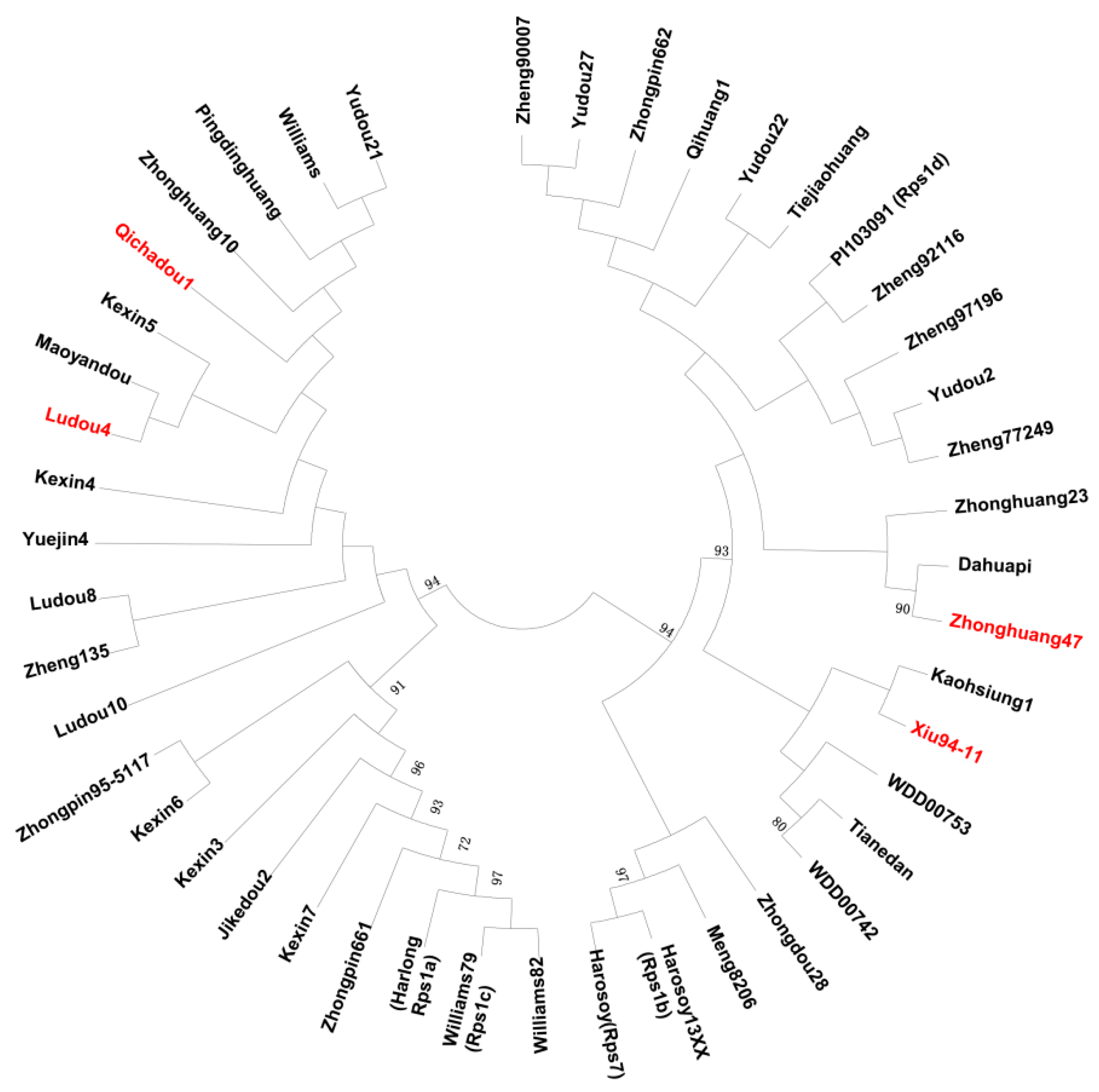
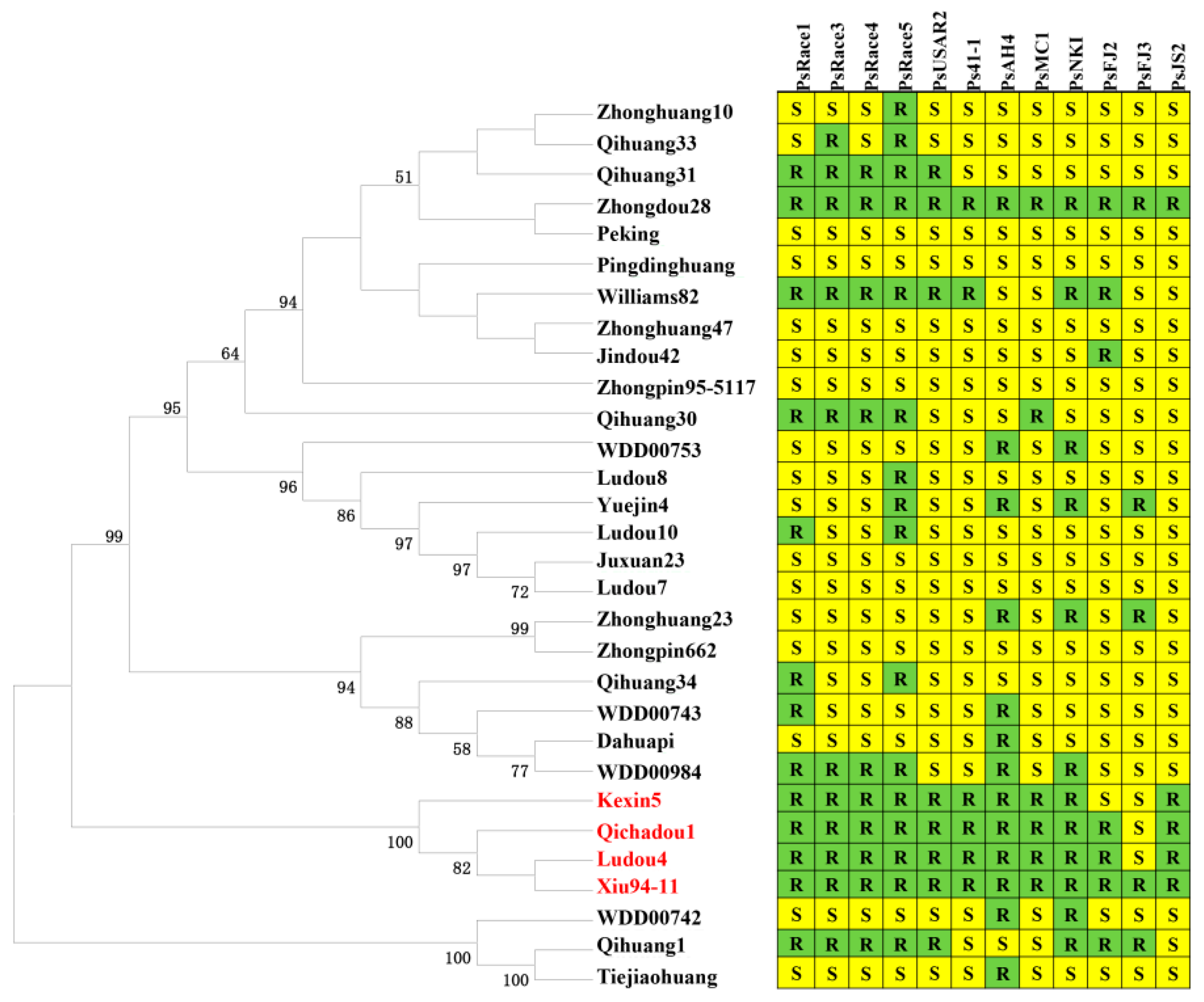
4.4. Phylogenetic Analysis of Soybean Genotypes
4.5. Analysis of the Allelic Sequences of the Candidate Gene Locus
4.6. Validation and Screening of RpsX and Its Alleles in Soybean Genotypes Using the Functional Marker Insert144
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| LRR | Leucine-rich repeat |
| PRR | Phytophthora root rot |
| NGS | Next-generation sequencing |
| SNP | Single nucleotide polymorphism |
| InDel | Insertion/deletion |
| STK | Serine/threonine protein kinase |
| NBS | Nucleotide-binding site |
References
- Zhou, Z.; Jiang, Y.; Wang, Z.; Gou, Z.; Lyu, J.; Li, W.; Yu, Y.; Shu, L.; Zhao, Y.; Ma, Y.; et al. Resequencing 302 wild and cultivated accessions identifies genes related to domestication and improvement in soybean. Nat. Biotechnol. 2015, 33, 408–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufmann, M.; Gerdemann, J. Root and stem rot of soybean caused by Phytophthora sojae n. sp. Phytopathology 1958, 48, 201–208. [Google Scholar]
- Schmitthenner, A.F. Problems and progress in control of Phytophthora root rot of soybean. Plant Dis. 1985, 69, 362–368. [Google Scholar] [CrossRef]
- Kamoun, S.; Furzer, O.; Jones, J.D.; Judelson, H.S.; Ali, G.S.; Dalio, R.J.; Roy, S.G.; Schena, L.; Zambounis, A.; Panabières, F.; et al. The Top 10 oomycete pathogens in molecular plant pathology. Mol. Plant Pathol. 2015, 16, 413–434. [Google Scholar] [CrossRef] [PubMed]
- Koenning, S.R.; Wrather, J.A. Suppression of soybean yield potential in the continental United States by plant diseases from 2006 to 2009. Plant Health Prog. 2010. Available online: https://www.plantmanagementnetwork.org/pub/php/research/2010/yield/ (accessed on 9 April 2019). [CrossRef]
- Dorrance, A.E. Management of Phytophthora sojae of soybean: A review and future perspectives. Can. J. Plant Pathol. 2018, 40, 210–219. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y. Phytophthora sojae. In Biological Invasions and Its Management in China; Wan, F., Jiang, M., Zhan, A., Eds.; Springer: Singapore, 2017; pp. 199–223. [Google Scholar]
- Tyler, B.M. Phytophthora sojae: Root rot pathogen of soybean and model oomycete. Mol. Plant Pathol. 2007, 8, 1–8. [Google Scholar] [CrossRef]
- Sugimoto, T.; Kato, M.; Yoshida, S.; Matsumoto, I.; Kobayashi, T.; Kaga, A.; Hajika, M.; Yamamoto, R.; Watanabe, K.; Aino, M.; et al. Pathogenic diversity of Phytophthora sojae and breeding strategies to develop Phytophthora-resistant soybeans. Breed. Sci. 2012, 61, 511–522. [Google Scholar] [CrossRef]
- Tooley, P.W.; Grau, C.R. The relationship between rate-reducing resistance to Phytophthora megasperma f. sp. glycinea and yield of soybean. Phytopathology 1984, 74, 1209–1216. [Google Scholar] [CrossRef]
- Dorrance, A.E.; St. McClure, S.A.; Martin, S.K. Effect of partial resistance on Phytophthora stem rot incidence and yield of soybean in Ohio. Plant Dis. 2003, 87, 308–312. [Google Scholar] [CrossRef]
- Dorrance, A.E.; Jia, H.; Abney, T.S. Evaluation of soybean differentials for their interaction with Phytophthora sojae. Plant Health Prog. 2004. [Google Scholar] [CrossRef]
- Zhang, J.; Xia, C.; Duan, C.; Sun, S.; Wang, X.; Wu, X.; Zhu, Z. Identification and candidate gene analysis of a novel Phytophthora resistance gene Rps10 in a Chinese soybean cultivar. PLoS ONE 2013, 8, e69799. [Google Scholar] [CrossRef]
- Zhang, J.; Xia, C.; Wang, X.; Duan, C.; Sun, S.; Wu, X.; Zhu, Z. Genetic characterization and fine mapping of the novel Phytophthora resistance gene in a Chinese soybean cultivar. Theor. Appl. Genet. 2013, 126, 1555–1561. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Zhao, M.; Ping, J.; Johnson, A.; Zhang, B.; Abney, T.S.; Hughes, T.; Ma, J. Molecular mapping of two genes conferring resistance to Phytophthora sojae in a soybean landrace PI 567139B. Theor. Appl. Genet. 2013, 126, 2177–2185. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, L.; Zhao, J.; Huang, J.; Yan, Q.; Xing, H.; Guo, N. Genetic analysis and fine mapping of RpsJS, a novel resistance gene to Phytophthora sojae in soybean [Glycine max (L.) Merr.]. Theor. Appl. Genet. 2014, 127, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Ping, J.; Fitzgerald, C.; Zhang, C.; Lin, F.; Bai, Y.; Wang, D.; Aggarwal, R.; Rehman, M.; Crasta, O.; Ma, J. Identification and molecular mapping of Rps11, a novel gene conferring resistance to Phytophthora sojae in soybean. Theor. Appl. Genet. 2016, 129, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lin, F.; Wang, W.; Ping, J.; Fitzgerald, J.; Zhao, M.; Li, S.; Sun, L.; Cai, C.; Ma, J. Fine mapping and candidate gene analysis of two loci conferring resistance to Phytophthora sojae in soybean. Theor. Appl. Genet. 2016, 129, 2379–2386. [Google Scholar] [CrossRef]
- Cheng, Y.; Ma, Q.; Ren, H.; Xia, Q.; Song, E.; Tan, Z.; Li, S.; Zhang, G.; Nian, H. Fine mapping of a Phytophthora-resistance gene RpsWY in soybean (Glycine max L.) by high-throughput genome-wide sequencing. Theor. Appl. Genet. 2017, 130, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, S.; Zhong, C.; Wang, X.; Wu, X.; Zhu, Z. Genetic mapping and development of co-segregating markers of RpsQ, which provides resistance to in soybean. Theor. Appl. Genet. 2017, 130, 1223–1233. [Google Scholar] [CrossRef]
- Niu, J.; Guo, N.; Sun, J.; Li, L.; Cao, Y.; Li, S.; Huang, J.; Zhao, J.; Zhao, T.; Xing, H. Fine mapping of a resistance gene RpsHN that controls Phytophthora sojae using recombinant inbred lines and secondary populations. Front. Plant Sci. 2017, 8, 538. [Google Scholar] [CrossRef]
- Sahoo, D.K.; Abeysekara, N.S.; Cianzio, S.R.; Robertson, A.E.; Bhattacharyya, M.K. A Novel Phytophthora sojae Resistance Rps12 Gene Mapped to a Genomic Region That Contains Several Rps Genes. PLoS ONE 2017, 12, e0169950. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Song, Q.; Shi, A.; Li, S.; Zhang, M.; Zhang, B. Genome-wide association mapping of resistance to Phytophthora sojae in a soybean [Glycine max (L.) Merr.] germplasm panel from maturity groups IV and V. PLoS ONE 2017, 12, e0184613. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Sun, S.; Li, Y.; Duan, C.; Zhu, Z. Next-generation sequencing to identify candidate genes and develop diagnostic markers for a novel Phytophthora resistance gene, RpsHC18, in soybean. Theor. Appl. Genet. 2017, 131, 525–538. [Google Scholar] [CrossRef]
- Zhong, C.; Sun, S.; Yao, L.; Ding, J.; Duan, C.; Zhu, Z. Fine mapping and identification of a novel phytophthora root rot resistance locus RpsZS18 on Chromosome 2 in Soybean. Front. Plant Sci. 2018, 9, 44. [Google Scholar] [CrossRef]
- Tian, M.; Zhao, L.; Li, S.; Huang, J.; Sui, Z.; Wen, J.; Li, Y. Pathotypes and metalaxyl sensitivity of Phytophthora sojae and their distribution in Heilongjiang, China 2011–2015. J. Gen. Plant Pathol. 2016, 82, 132–141. [Google Scholar] [CrossRef]
- Wu, M.; Li, B.; Liu, P.; Weng, Q.; Zhan, J.; Chen, Q. Genetic analysis of Phytophthora sojae populations in Fujian, China. Plant Pathol. 2017, 66, 1182–1190. [Google Scholar] [CrossRef]
- Dorrance, A.E.; Kurle, J.; Robertson, A.E.; Bradley, C.A.; Giesler, L.; Wise, K.; Concibido, V.C. Pathotype diversity of Phytophthora sojae in eleven states in the United States. Plant Dis. 2016, 100, 1429–1437. [Google Scholar] [CrossRef]
- Schmitthenner, A.F. Phytophthora rot of soybean. In Compendium of Soybean Diseases, 4th ed.; The American Phytopathological Society Press: St. Paul, MN, USA, 1999; pp. 39–42. [Google Scholar]
- Costamilan, L.M.; Clebsch, C.C.; Soares, R.M.; Seixas, C.D.S.; Godoy, C.V.; Dorrance, A.E. Pathogenic diversity of Phytophthora sojae pathotypes from Brazil. Eur. J. Plant Pathol. 2013, 135, 845–853. [Google Scholar] [CrossRef]
- Dorrance, A.E.; Schmitthenner, A.F. New sources of resistance to Phytophthora sojae in the soybean plant introductions. Plant Dis. 2000, 84, 1303–1308. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, S.; Wang, G.; Duan, C.; Wang, X.; Wu, X.; Zhu, Z. Characterization of Phytophthora resistance in soybean cultivars/lines bred in Henan province. Euphytica 2014, 196, 375–384. [Google Scholar] [CrossRef]
- Scheben, A.; Batley, J.; Edwards, D. Genotyping by sequencing approaches to characterise crop genomes: Choosing the right tool for the right application. Plant Biotechnol. J. 2016, 15, 149–161. [Google Scholar] [CrossRef]
- Wallace, J.G.; Mitchell, S.E. Genotyping-by-sequencing. Curr. Protoc. Plant Biol. 2017, 2, 64–77. [Google Scholar]
- Grant, D.; Nelson, R.T.; Cannon, S.B.; Shoemaker, R.C. SoyBase, the USDA-ARS soybean genetics and genomics database. Nucleic Acids Res. 2009, 38, 843–846. [Google Scholar] [CrossRef]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef]
- Song, Q.; Jenkins, J.; Jia, G.; Hyten, D.L.; Pantalone, V.; Jackson, S.A.; Schmutz, J.; Cregan, P.B. Construction of high resolution genetic linkage maps to improve the soybean genome sequence assembly Glyma1. 01. BMC Genom. 2016, 17, 33. [Google Scholar] [CrossRef]
- Takagi, H.; Abe, A.; Yoshida, K.; Kosugi, S.; Natsume, S.; Mitsuoka, C.; Uemura, A.; Utsushi, H.; Tamiru, M.; Takuno, S.; et al. QTL-seq: Rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J. 2013, 74, 174–183. [Google Scholar] [CrossRef]
- Lu, H.; Lin, T.; Klein, J.; Wang, S.; Qi, J.; Zhou, Q.; Sun, J.; Zhang, Z.; Weng, Y.; Huang, S. QTL-seq identifies an early flowering QTL located near flowering locus T in cucumber. Theor. Appl. Genet. 2014, 127, 1491–1499. [Google Scholar] [CrossRef]
- Illa-Berenguer, E.; Van Houten, J.; Huang, Z.; van der Knaap, E. Rapid and reliable identification of tomato fruit weight and locule number loci byQTL-seq. Theor. Appl. Genet. 2015, 128, 1329–1342. [Google Scholar] [CrossRef]
- Das, S.; Upadhyaya, H.D.; Bajaj, D.; Kujur, A.; Badoni, S.; Laxmi; Kumar, V.; Tripathi, S.; Gowda, C.L.; Sharma, S.; et al. Deploying QTL-seq for rapid delineation of a potential candidate gene underlying major trait-associated QTL in chickpea. DNA Res. 2015, 22, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.K.; Khan, A.W.; Jaganathan, D.; Thudi, M.; Roorkiwal, M.; Takagi, H.; Garg, V.; Kumar, V.; Chitikineni, A.; Gaur, P.M.; et al. QTL-seq for rapid identification of candidate genesfor 100-seed weight and root/total plant dry weight ratio under rained conditions in chickpea. Plant Biotechnol. J. 2016, 14, 2110–2119. [Google Scholar] [CrossRef]
- Singh, V.K.; Khan, A.W.; Saxena, R.K.; Kumar, V.; Kale, S.M.; Sinha, P.; Chitikineni, A.; Pazhamala, L.T.; Garg, V.; Sharma, M.; et al. Next-generation sequencing for identification of candidate genes for Fusarium wilt and sterility mosaic disease in pigeonpea (Cajanus cajan). Plant Biotechnol. J. 2016, 14, 1183–1194. [Google Scholar] [CrossRef]
- Singh, V.K.; Khan, A.W.; Saxena, R.K.; Sinha, P.; Kale, S.M.; Parupalli, S.; Kumar, V.; Chitikineni, A.; Vechalapu, S.; Sameer Kumar, C.V.; et al. Indel-seq: A fast-forward genetics approach for identification of trait-associated putative candidate genomic regions and its application in pigeonpea (Cajanus cajan). Plant Biotechnol. J. 2017, 15, 906–914. [Google Scholar] [CrossRef]
- Pandey, M.K.; Khan, A.W.; Singh, V.K.; Vishwakarma, M.K.; Shasidhar, Y.; Kumar, V.; Garg, V.; Bhat, R.S.; Chitikineni, A.; Janila, P.; et al. QTL-seq approach identified genomic regions and diagnostic markers for rust and late leaf spot resistance in groundnut (Arachis hypogaea L.). Plant Biotechnol. J. 2017, 15, 927–941. [Google Scholar] [CrossRef]
- Lin, M.S.; Lin, S.F. Pedigree analysis of soybean varieties. Bot. Bull. Acad. Sin. 1994, 35, 87–93. [Google Scholar]
- Gao, H.; Bhattacharyya, M.K. The soybean-Phytophthora resistance locus Rps1-k encompasses coiled coil-nucleotide binding leucine rich repeat-like genes and repetitive sequences. BMC Plant Biol. 2008, 8, 29. [Google Scholar] [CrossRef]
- Gao, H.; Narayanan, N.N.; Ellison, L.; Bhattacharyya, M.K. Two classes of highly similar coiled coil-nucleotide binding-leucine rich repeat genes isolated from the Rps1-k locus encode Phytophthora resistance in soybean. Mol. Plant Microb. Interact. 2005, 18, 1035–1045. [Google Scholar] [CrossRef]
- Feuillet, C.; Schachermayr, G.; Keller, B. Molecular cloning of a new receptor-like kinase gene encoded at the Lrl0 disease resistance locus of wheat. Plant J. 1997, 11, 45–52. [Google Scholar] [CrossRef]
- Song, W.Y.; Wang, G.L.; Chen, L.L.; Kim, H.S.; Pi, L.Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.X.; Zhu, L.H.; et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 27, 1804–1806. [Google Scholar] [CrossRef]
- Wang, G.L.; Ruan, D.L.; Song, W.Y.; Sideris, S.; Chen, L.; Pi, L.Y.; Zhang, S.; Zhang, Z.; Fauquet, C.; Gautd, B.S.; et al. Xa21D encodes a receptor-like molecule with a leucine-rich repeat domain that determines race-specific recognition and is subject to adaptive evolution. Plant Cell 1998, 10, 765–779. [Google Scholar] [CrossRef]
- Sun, X.; Cao, Y.; Yang, Z.; Xu, C.; Li, X.; Wang, S.; Zhang, Q. Xa26, a gene conferring resistance to Xanthomonas oryzae pv. oryzae in rice, encodes an LRR receptor kinase-like protein. Plant J. 2004, 37, 517–527. [Google Scholar] [CrossRef]
- Padmarasu, S.; Sargent, D.J.; Patocchi, A.; Troggio, M.; Baldi, P.; Linsmith, G.; Poles, L.; Jänsch, M.; Kellerhals, M.; Tartarini, S.; et al. Identification of a leucine-rich repeat receptor-like serine/threonine-protein kinase as a candidate gene for Rvi12 (Vb)-based apple scab resistance. Mol. Breed. 2018, 38, 73. [Google Scholar] [CrossRef]
- Dodds, P.N.; Lawrence, G.J.; Ellis, J.G. Six amino acidchanges confined to the leucine-rich repeat beta-strand/beta-turnmotif determine the difference between the P and P2 rust resistance specificities in flax. Plant Cell 2001, 13, 163–178. [Google Scholar]
- Jones, D.; Jones, J.D. The role of leucine-rich repeat proteins in plant defenses. Adv. Bot. Res. 1997, 24, 89–167. [Google Scholar]
- Ellis, J.; Dodds, P.; Pryor, T. The generation of plant disease resistance gene specificities. Trends Plant Sci. 2000, 5, 373–379. [Google Scholar] [CrossRef]
- Ramkumar, G.; Madhav, M.S.; Devi, S.R.; Umakanth, B.; Pandey, M.K.; Prasad, M.S.; Sundaram, R.M.; Viraktamath, B.C.; Babu, V.R. Identification and validation of novel alleles of rice blast resistant gene Pi54, and analysis of their nucleotide diversity in landraces and wild Oryza species. Euphytica 2016, 209, 725–737. [Google Scholar] [CrossRef]
- Thakur, S.; Singh, P.K.; Das, A.; Rathour, R.; Variar, M.; Prashanthi, S.K.; Singh, A.K.; Singh, U.D.; Chand, D.; Singh, N.K.; et al. Extensive sequence variation in rice blast resistance gene Pi54 makes it broad spectrum in nature. Front. Plant Sci. 2015, 6, 345. [Google Scholar] [CrossRef]
- Wicker, T.; Yahiaoui, N.; Keller, B. Illegitimate recombination is a major evolutionary mechanism for initiating size variation in plant resistance genes. Plant J. 2007, 51, 631–641. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, G.J.; Anderson, P.A.; Dodds, P.N.; Ellis, J.G. Relationships between rust resistance genes at the M locus in flax. Mol. Plant Pathol. 2010, 11, 19–32. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; DePristo, M.A. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Lincoln, S.E.; Daly, M.J.; Lander, E.S. Constructing Genetic Linkage Maps with MAPMAKER/EXP Version 3.0: A Tutorial and Reference Manual. A Whitehead Institute for Biomedical Research Technical Report. 1993, Volume 3. Available online: http://home.cc.umanitoba.ca/~psgendb/birchhomedir/doc/mapmaker/mapmaker.tutorial.pdf (accessed on 9 April 2019).
- Liu, R.H.; Meng, J.L. MapDraw: A microsoft excel macro for drawing genetic linkage maps based on given genetic linkage data. Hereditas 2003, 25, 317–321. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, 222–226. [Google Scholar] [CrossRef]
- Letunic, I.; Doerks, T.; Bork, P. SMART: Recent updates, new developments and status in 2015. Nucleic Acids Res. 2015, 43, 257–260. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar (Rps gene) | PsRace1 | PsRace3 | PsRace4 | PsRace5 | PsUSAR2 | Ps41-1 | PsAH4 | PsMC1 | PsNKI | PsFJ2 | PsFJ3 | PsJS2 | Ps6497 | Ps7063 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Harlon (Rps1a) | S 1 | S | S | S | R | S | R | S | R | S | S | S | R | S |
| Harosoy13XX (Rps1b) | R | R | R | R | S | S | S | S | S | S | S | S | S | R |
| Williams79 (Rps1c) | R | R | R | R | R | R | S | R | R | R | R | S | R | R |
| PI103091 (Rps1d) | R | S | S | R | R | S | S | S | S | S | S | S | S | S |
| Williams82 (Rps1k) | R | R | R | R | R | R | S | S | R | R | S | S | R | R |
| L76-988 (Rps2) | R | R | R | R | S | S | S | S | S | S | S | S | S | S |
| L83-570 (Rps3a) | R | R | R | R | R | S | S | S | S | S | S | S | R | S |
| PRX146-36 (Rps3b) | R | R | S | R | R | S | S | S | S | S | S | S | R | R |
| PRX145-48 (Rps3c) | R | R | R | R | S | S | S | S | S | S | S | S | S | R |
| L85-2352 (Rps4) | R | R | R | R | R | S | S | S | S | S | S | S | R | S |
| L85-3059 (Rps5) | R | R | R | R | S | S | S | S | S | S | S | S | R | S |
| Harosoy62XX (Rps6) | R | R | R | R | R | S | R | S | S | S | S | S | R | S |
| Harosoy (Rps7) | R | R | R | S | R | S | S | S | S | S | S | S | S | S |
| PI399073 (Rps8) | R | R | R | R | R | S | R | S | S | S | S | S | R | S |
| Youbian30 (RpsYB30) | R | R | S | R | R | R | S | S | R | R | S | S | S | S |
| Yudou25 (RpsYD25) | R | R | R | R | R | R | S | R | R | S | R | S | R | R |
| Yudou29 (RpsYD29) | R | R | R | R | R | R | S | R | R | R | R | S | R | R |
| Ludou4 (Rps9) | R | R | R | R | R | R | R | R | R | R | S | R | R | R |
| Qichadou 1 (RpsQ) | R | R | R | R | R | R | R | R | R | R | S | R | R | R |
| Wandou15 (Rps10) | R | R | R | R | R | R | R | R | R | S | R | S | R | S |
| Zaoshu18 (RpsZS18) | R | R | R | R | R | R | S | S | R | R | R | S | R | S |
| Huachun18 (RpsHC18) | R | R | R | R | R | R | R | R | R | R | R | R | R | S |
| Xiu94-11 (RpsX) | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| Zhonghuang13 | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Williams (rps) | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Jikedou 2 | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Zhonghuang 47 | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Parent and the Cross | Generation | Total Plants | Observed Number | Except Ratio and Goodness of Fit | ||||
|---|---|---|---|---|---|---|---|---|
| R 1 | Rs | S | R:Rs:S | χ2 | P | |||
| Xiu94-11 | P1 | 15 | 15 | - | - | |||
| Zhonghuang47 | P2 | 15 | - | - | 15 | |||
| Zhonghuang47 × Xiu94-11 | F2:3 | 137 | 38 | 63 | 36 | 1:2:1 | 0.93 | 0.62 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, C.; Li, Y.; Sun, S.; Duan, C.; Zhu, Z. Genetic Mapping and Molecular Characterization of a Broad-spectrum Phytophthora sojae Resistance Gene in Chinese Soybean. Int. J. Mol. Sci. 2019, 20, 1809. https://doi.org/10.3390/ijms20081809
Zhong C, Li Y, Sun S, Duan C, Zhu Z. Genetic Mapping and Molecular Characterization of a Broad-spectrum Phytophthora sojae Resistance Gene in Chinese Soybean. International Journal of Molecular Sciences. 2019; 20(8):1809. https://doi.org/10.3390/ijms20081809
Chicago/Turabian StyleZhong, Chao, Yinping Li, Suli Sun, Canxing Duan, and Zhendong Zhu. 2019. "Genetic Mapping and Molecular Characterization of a Broad-spectrum Phytophthora sojae Resistance Gene in Chinese Soybean" International Journal of Molecular Sciences 20, no. 8: 1809. https://doi.org/10.3390/ijms20081809
APA StyleZhong, C., Li, Y., Sun, S., Duan, C., & Zhu, Z. (2019). Genetic Mapping and Molecular Characterization of a Broad-spectrum Phytophthora sojae Resistance Gene in Chinese Soybean. International Journal of Molecular Sciences, 20(8), 1809. https://doi.org/10.3390/ijms20081809