MicroRNA-214-3p Targeting Ctnnb1 Promotes 3T3-L1 Preadipocyte Differentiation by Interfering with the Wnt/β-Catenin Signaling Pathway

 and
and
Abstract
:1. Introduction
2. Results
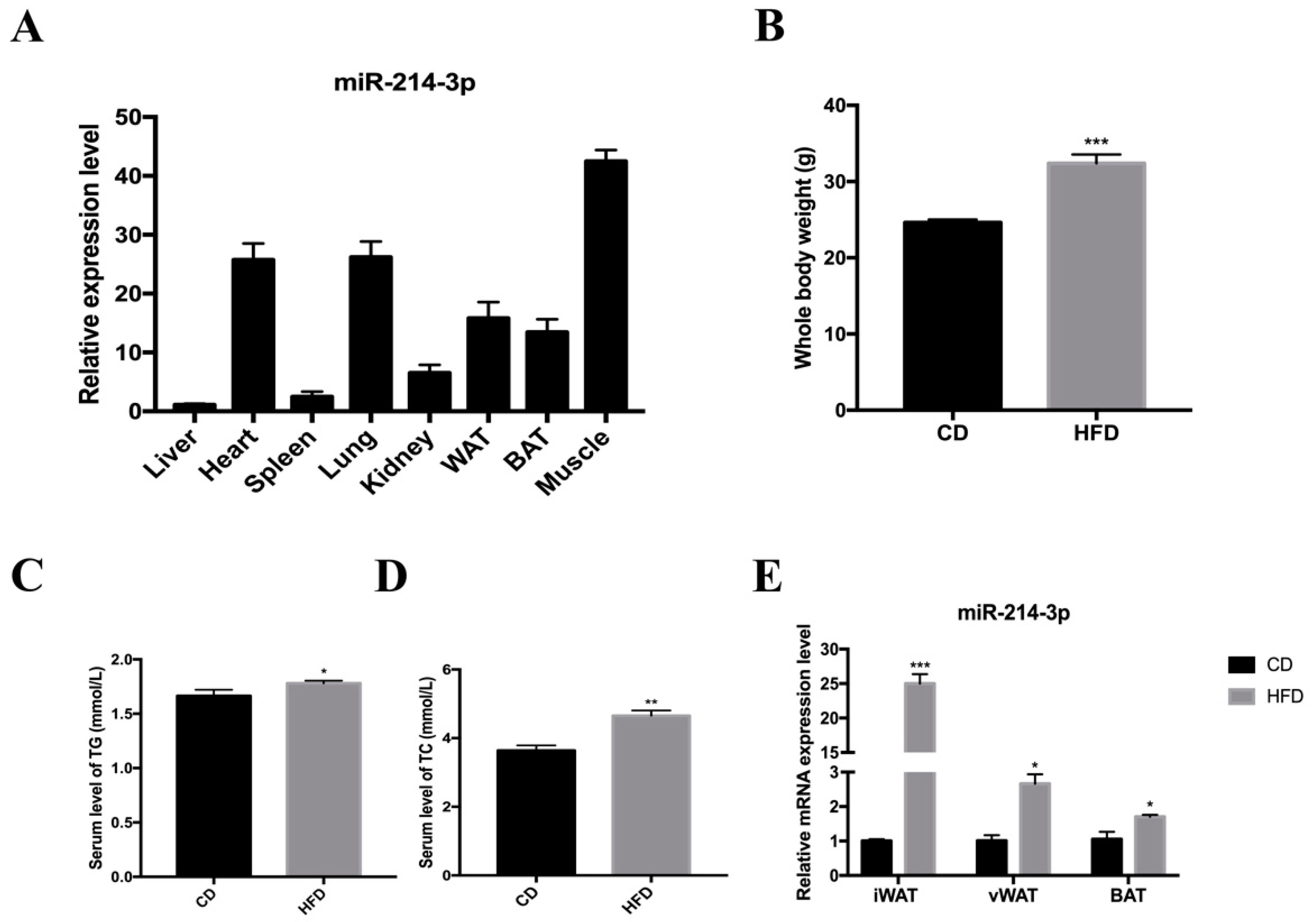
2.1. The Expression of miR-214-3p Is Strongly Associated with Lipogenesis
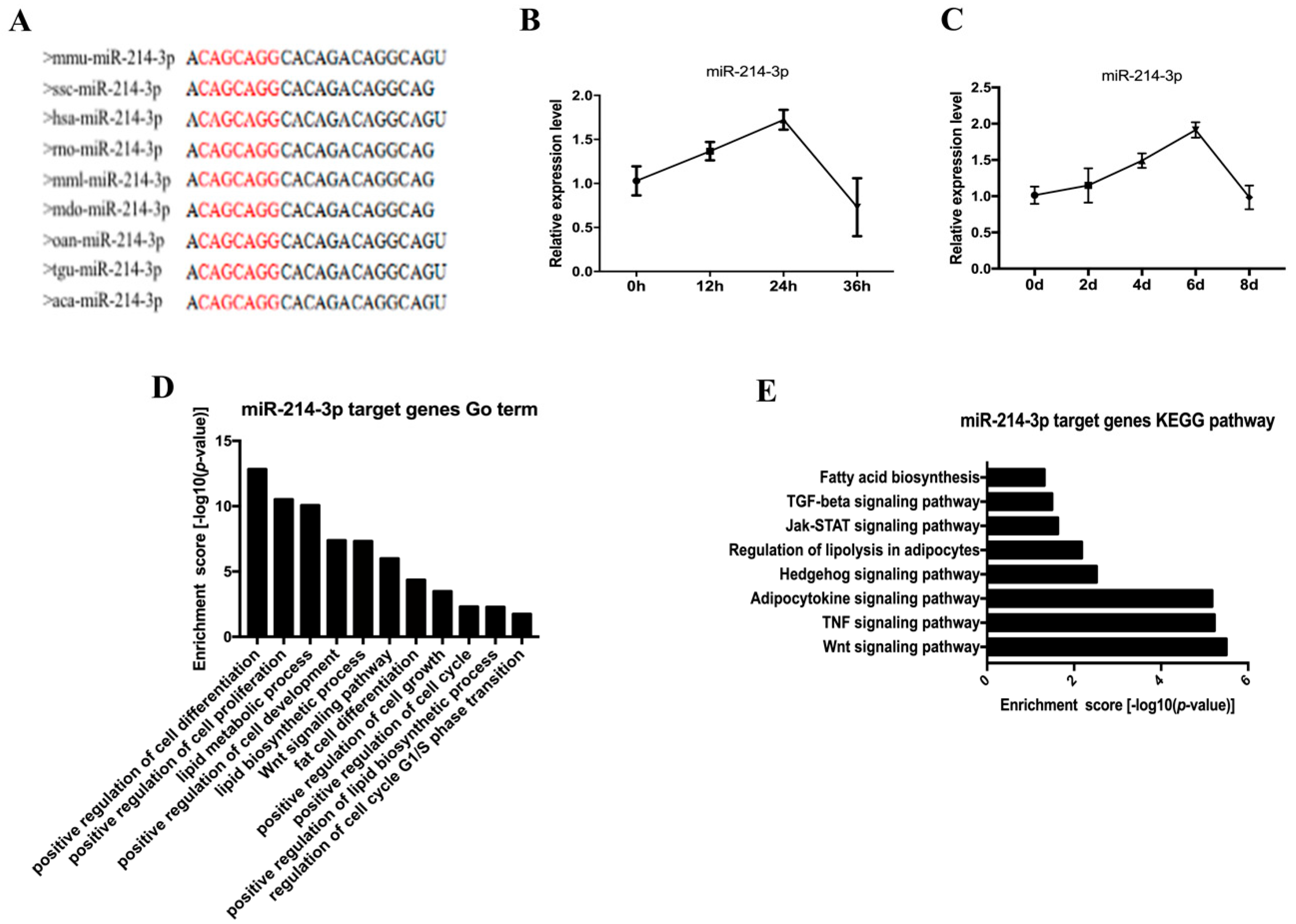
2.2. The Profile of miR-214-3p in Adipogenesis
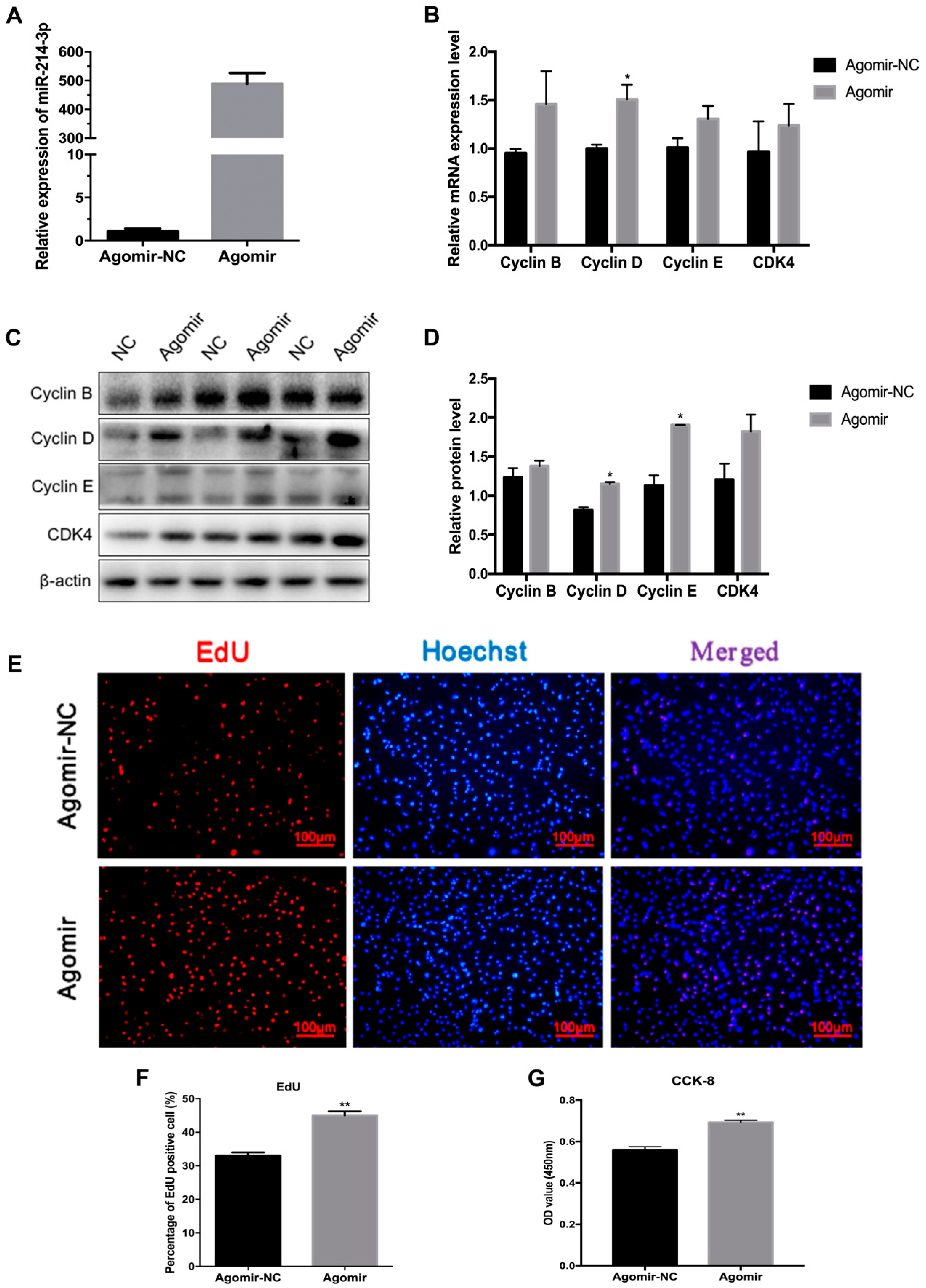
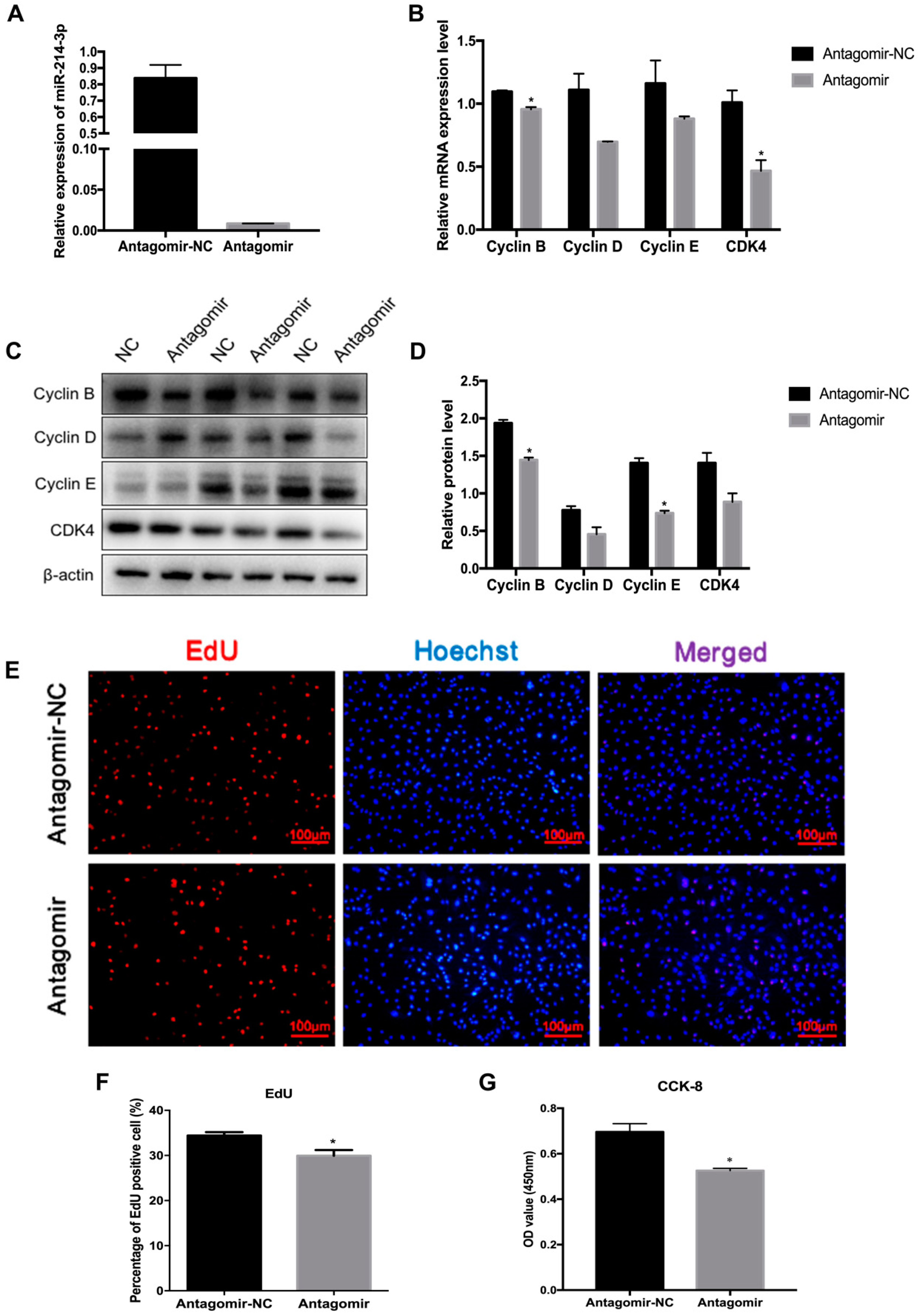
2.3. MicroRNA-214-3p Promotes 3T3-L1 Preadipocyte Proliferation
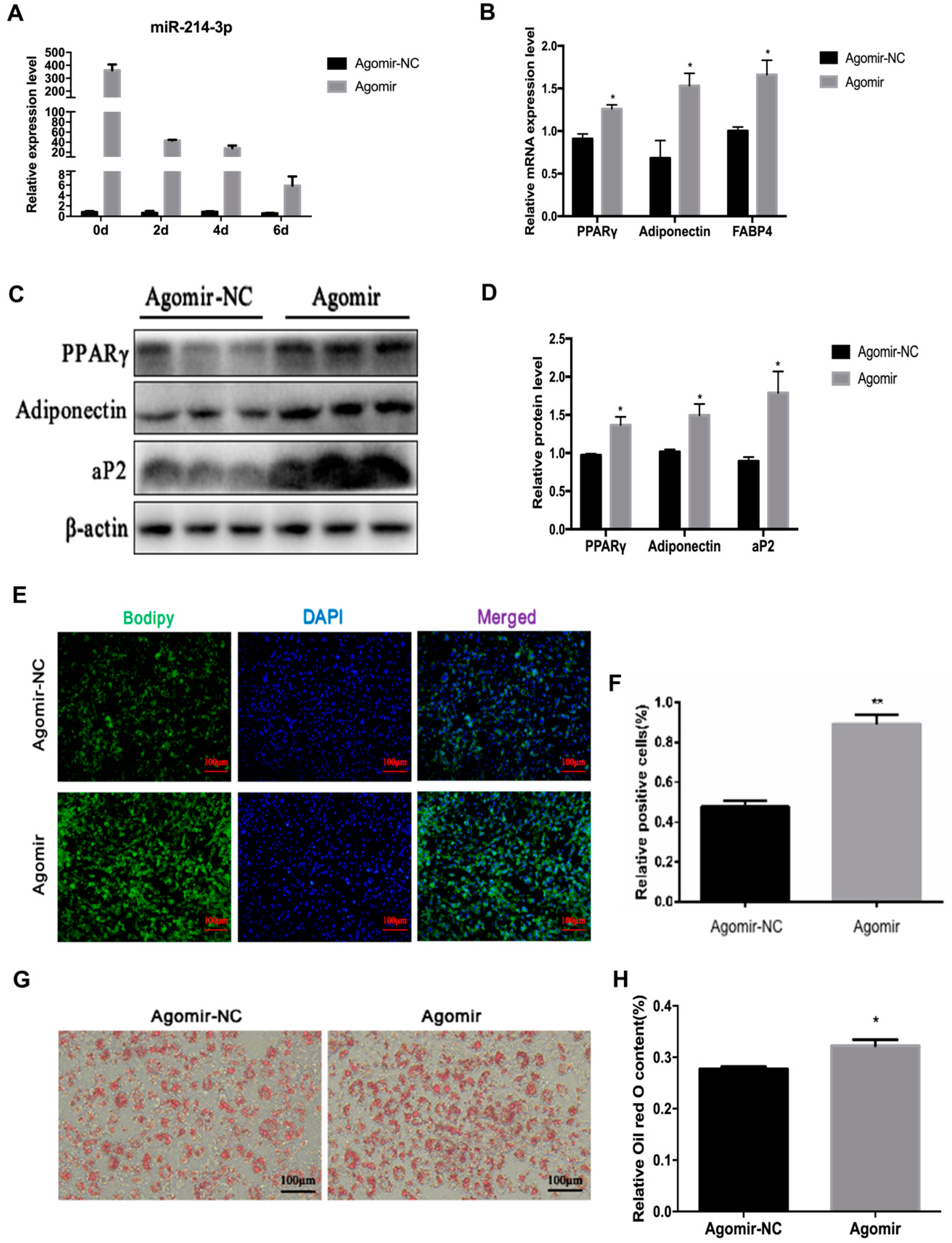
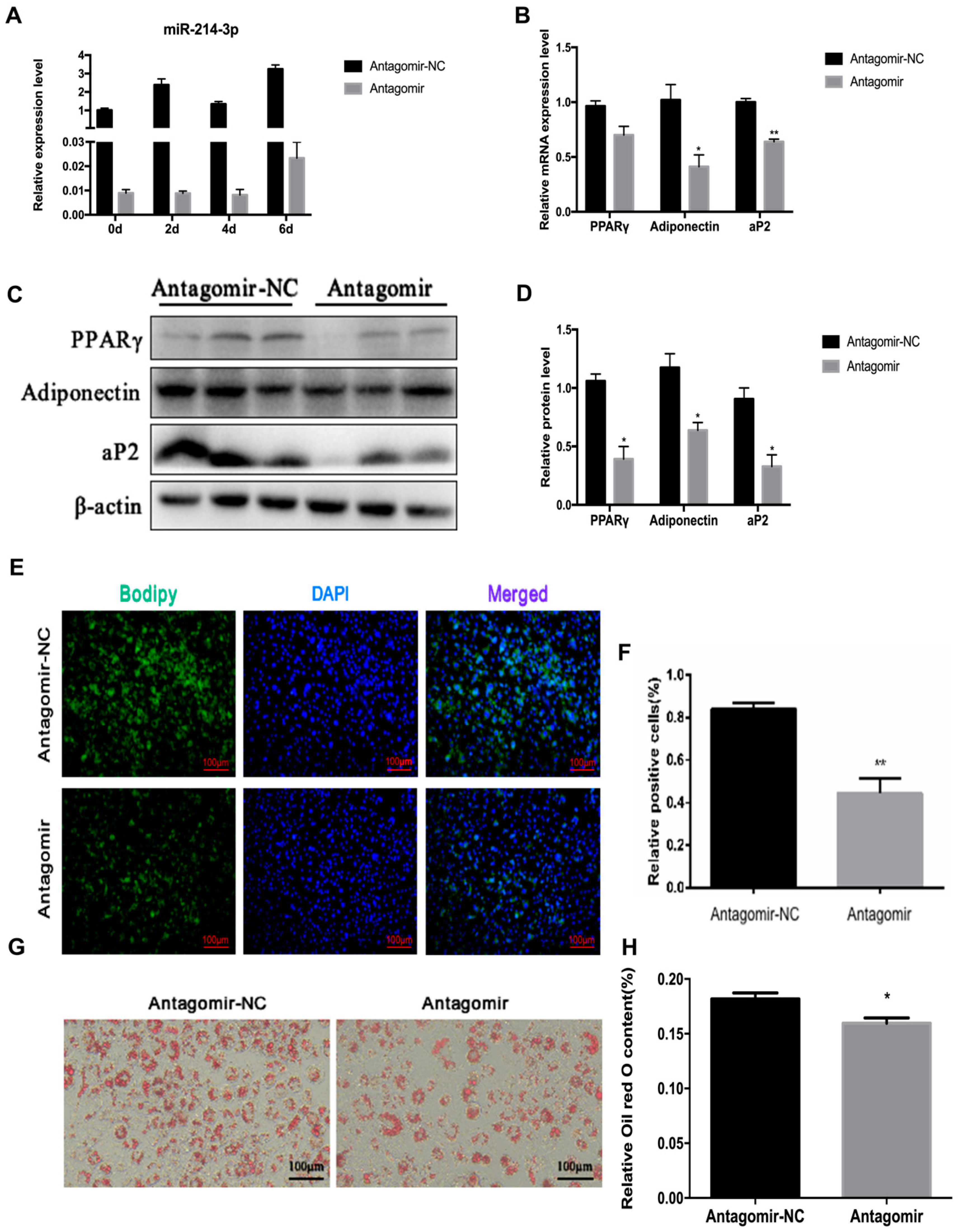
2.4. MicroRNA-214-3p Enhances 3T3-L1 Preadipocyte Differentiation
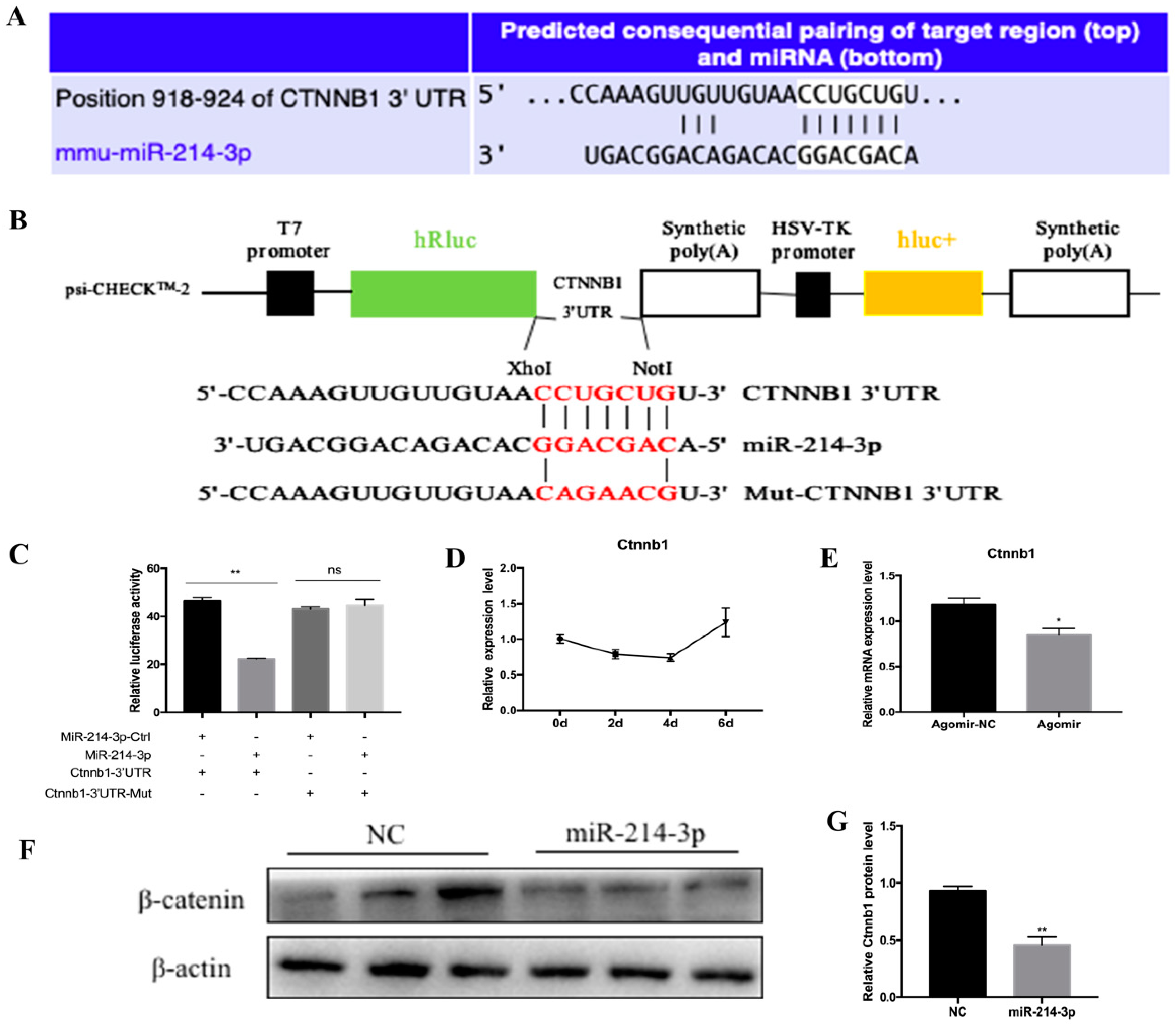
2.5. MicroRNA-214-3p Promotes 3T3-L1 Preadipocyte Differentiation by Targeting Ctnnb1 and Regulating the Canonical Wnt/β-Catenin Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Serum Analysis
4.3. Cell Culture and Differentiation
4.4. Transfection of miRNA Agomir and Antagomir
4.5. RNA Isolation and RT-qPCR
4.6. Western Blot Analysis
4.7. EdU Staining
4.8. Cell Counting Kit
4.9. Oil Red O staining
4.10. BODIPY Staining
4.11. Luciferase Reporter Assay
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| aP2 | Adipocyte fatty acid binding protein 4 (FABP4) |
| Cyclin B | Cell cycle protein B |
| Cyclin D | Cell cycle protein D |
| Cyclin E | Cell cycle protein E |
| CDK4 | Cyclin-dependent kinase 4 |
| mmu | Mus musculus |
| ssc | Sus scrofa |
| hsa | Homo sapiens |
| rno | Rattus norvegicus |
| mml | Macaca mulatta |
| mdo | Monodelphis domestica |
| oan | Ornithorhynchus anatinus |
| tgu | Taeniopygia guttata |
| aca | Anolis carolinensis |
References
- Bou, M.; Todorcevic, M.; Rodriguez, J.; Capilla, E.; Gutierrez, J.; Navarro, I. Interplay of adiponectin, TNFalpha and insulin on gene expression, glucose uptake and PPARgamma, AKT and TOR pathways in rainbow trout cultured adipocytes. Gen. Comp. Endocrinol. 2014, 205, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Vienberg, S.; Geiger, J.; Madsen, S.; Dalgaard, L.T. MicroRNAs in metabolism. Acta Physiol. 2017, 219, 346–361. [Google Scholar] [CrossRef]
- Rosen, E.D.; Spiegelman, B.M. Adipocytes as regulators of energy balance and glucose homeostasis. Nature 2006, 444, 847–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singla, P.; Bardoloi, A.; Parkash, A.A. Metabolic effects of obesity: A review. World J. Diabetes 2010, 1, 76–88. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/ (accessed on 12 December 2018).
- Bhupathiraju, S.N.; Hu, F.B. Epidemiology of Obesity and Diabetes and Their Cardiovascular Complications. Circ. Res. 2016, 118, 1723–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Despres, J.P. Is visceral obesity the cause of the metabolic syndrome? Ann. Med. 2006, 38, 52–63. [Google Scholar] [CrossRef]
- Gallagher, E.J.; LeRoith, D. Obesity and Diabetes: The Increased Risk of Cancer and Cancer-Related Mortality. Physiol. Rev. 2015, 95, 727–748. [Google Scholar] [CrossRef] [Green Version]
- Kahn, B.B.; Flier, J.S. Obesity and insulin resistance. J. Clin. Investig. 2000, 106, 473–481. [Google Scholar] [CrossRef] [Green Version]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell. Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Graves, P.; Zeng, Y. Biogenesis of mammalian microRNAs: A global view. Genom. Proteom. Bioinform. 2012, 10, 239–245. [Google Scholar] [CrossRef]
- Li, M.; Liu, Z.; Zhang, Z.; Liu, G.; Sun, S.; Sun, C. miR-103 promotes 3T3-L1 cell adipogenesis through AKT/mTOR signal pathway with its target being MEF2D. Biol. Chem. 2015, 396, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Chen, F.F.; Ge, J.; Zhu, J.Y.; Shi, X.E.; Li, X.; Yu, T.Y.; Chu, G.Y.; Yang, G.S. miR-429 Inhibits Differentiation and Promotes Proliferation in Porcine Preadipocytes. Int. J. Mol. Sci. 2016, 17, 2047. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Du, J.; Wang, L.; Niu, L.; Zhao, Y.; Tang, G.; Jiang, Y.; Shuai, S.; Bai, L.; Li, X.; et al. MicroRNA-143a-3p modulates preadipocyte proliferation and differentiation by targeting MAPK7. Biomed. Pharmacother. 2018, 108, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Sato, T.; Amano, T.; Kawamura, Y.; Kawamura, N.; Kawaguchi, H.; Yamashita, N.; Kurihara, H.; Nakaoka, T. Dnm3os, a non-coding RNA, is required for normal growth and skeletal development in mice. Dev. Dyn. 2008, 237, 3738–3748. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.B.; Bantounas, I.; Lee, D.Y.; Phylactou, L.; Caldwell, M.A.; Uney, J.B. Twist-1 regulates the miR-199a/214 cluster during development. Nucleic Acids Res. 2009, 37, 123–128. [Google Scholar] [CrossRef]
- Loebel, D.A.; Tsoi, B.; Wong, N.; Tam, P.P. A conserved noncoding intronic transcript at the mouse Dnm3 locus. Genomics 2005, 85, 782–789. [Google Scholar] [CrossRef]
- Li, D.; Liu, J.; Guo, B.; Liang, C.; Dang, L.; Lu, C.; He, X.; Cheung, H.Y.; Xu, L.; Lu, C.; et al. Osteoclast-derived exosomal miR-214-3p inhibits osteoblastic bone formation. Nat. Commun. 2016, 7, 10872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Guo, B.; Li, Q.; Peng, J.; Yang, Z.; Wang, A.; Li, D.; Hou, Z.; Lv, K.; Kan, G.; et al. miR-214 targets ATF4 to inhibit bone formation. Nat. Med. 2013, 19, 93–100. [Google Scholar] [CrossRef]
- Flynt, A.S.; Li, N.; Thatcher, E.J.; Solnica-Krezel, L.; Patton, J.G. Zebrafish miR-214 modulates Hedgehog signaling to specify muscle cell fate. Nat. Genet. 2007, 39, 259–263. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Kuek, V.; Liu, Y.; Tickner, J.; Yuan, Y.; Chen, L.; Zeng, Z.; Shao, M.; He, W.; Xu, J. MiR-214 is an important regulator of the musculoskeletal metabolism and disease. J. Cell Physiol. 2018, 234, 231–245. [Google Scholar] [CrossRef]
- Wang, R.; Sun, Y.; Yu, W.; Yan, Y.; Qiao, M.; Jiang, R.; Guan, W.; Wang, L. Downregulation of miRNA-214 in cancer-associated fibroblasts contributes to migration and invasion of gastric cancer cells through targeting FGF9 and inducing EMT. J. Exp. Clin. Cancer Res. 2019, 38, 20. [Google Scholar] [CrossRef]
- Qin, L.; Chen, Y.; Niu, Y.; Chen, W.; Wang, Q.; Xiao, S.; Li, A.; Xie, Y.; Li, J.; Zhao, X.; et al. A deep investigation into the adipogenesis mechanism: Profile of microRNAs regulating adipogenesis by modulating the canonical Wnt/beta-catenin signaling pathway. BMC Genom. 2010, 11, 320. [Google Scholar] [CrossRef]
- Knelangen, J.M.; van der Hoek, M.B.; Kong, W.-C.; Owens, J.A.; Fischer, B.; Santos, A.N. MicroRNA expression profile during adipogenic differentiation in mouse embryonic stem cells. Physiol. Genom. 2011, 43, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell. Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef]
- Guo, Y.; Li, L.; Gao, J.; Chen, X.; Sang, Q. miR-214 suppresses the osteogenic differentiation of bone marrow-derived mesenchymal stem cells and these effects are mediated through the inhibition of the JNK and p38 pathways. Int. J. Mol. Med. 2017, 39, 71–80. [Google Scholar] [CrossRef]
- Kaur, Y.; de Souza, R.J.; Gibson, W.T.; Meyre, D. A systematic review of genetic syndromes with obesity. Obes. Rev. 2017, 18, 603–634. [Google Scholar] [CrossRef]
- Mota de Sa, P.; Richard, A.J.; Hang, H.; Stephens, J.M. Transcriptional Regulation of Adipogenesis. Compr. Physiol. 2017, 7, 635–674. [Google Scholar]
- Maurizi, G.; Babini, L.; Della Guardia, L. Potential role of microRNAs in the regulation of adipocytes liposecretion and adipose tissue physiology. J. Cell Physiol. 2018, 233, 9077–9086. [Google Scholar] [CrossRef]
- He, L.; Tang, M.; Xiao, T.; Liu, H.; Liu, W.; Li, G.; Zhang, F.; Xiao, Y.; Zhou, Z.; Liu, F.; et al. Obesity-Associated miR-199a/214 Cluster Inhibits Adipose Browning via PRDM16-PGC-1alpha Transcriptional Network. Diabetes 2018, 67, 2585–2600. [Google Scholar] [CrossRef]
- Yi, S.J.; Li, L.L.; Tu, W.B. MiR-214 negatively regulates proliferation and WNT/beta-catenin signaling in breast cancer. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 5148–5154. [Google Scholar]
- Cai, H.; Miao, M.; Wang, Z. miR-214-3p promotes the proliferation, migration and invasion of osteosarcoma cells by targeting CADM1. Oncol. Lett. 2018, 16, 2620–2628. [Google Scholar] [CrossRef]
- Feng, Y.; Cao, J.H.; Li, X.Y.; Zhao, S.H. Inhibition of miR-214 expression represses proliferation and differentiation of C2C12 myoblasts. Cell Biochem. Funct. 2011, 29, 378–383. [Google Scholar] [CrossRef]
- Mazumder, S.; DuPre, E.L.; Almasan, A. A dual role of Cyclin E in cell proliferation and apotosis may provide a target for cancer therapy. Curr. Cancer Drug Targets 2004, 4, 65–75. [Google Scholar] [CrossRef]
- Tan, Z.; Du, J.; Shen, L.; Liu, C.; Ma, J.; Bai, L.; Jiang, Y.; Tang, G.; Li, M.; Li, X.; et al. miR-199a-3p affects adipocytes differentiation and fatty acid composition through targeting SCD. Biochem. Biophys Res. Commun. 2017, 492, 82–88. [Google Scholar] [CrossRef]
- Shi, X.E.; Li, Y.F.; Jia, L.; Ji, H.L.; Song, Z.Y.; Cheng, J.; Wu, G.F.; Song, C.C.; Zhang, Q.L.; Zhu, J.Y.; et al. MicroRNA-199a-5p affects porcine preadipocyte proliferation and differentiation. Int. J. Mol. Sci. 2014, 15, 8526–8538. [Google Scholar] [CrossRef]
- Giralt, M.; Villarroya, F. White, brown, beige/brite: Different adipose cells for different functions? Endocrinology 2013, 154, 2992–3000. [Google Scholar] [CrossRef]
- Miller, R.K.; Hong, J.Y.; Munoz, W.A.; McCrea, P.D. Beta-catenin versus the other armadillo catenins: Assessing our current view of canonical Wnt signaling. Prog. Mol. Biol. Transl. Sci. 2013, 116, 387–407. [Google Scholar]
- Fagotto, F. Looking beyond the Wnt pathway for the deep nature of beta-catenin. EMBO Rep. 2013, 14, 422–433. [Google Scholar] [CrossRef]
- Chen, N.; Wang, J. Wnt/beta-Catenin Signaling and Obesity. Front. Physiol. 2018, 9, 792. [Google Scholar] [CrossRef]
- Ross, S.E. Inhibition of Adipogenesis by Wnt Signaling. Science 2000, 289, 950–953. [Google Scholar] [CrossRef]
- Chen, C.; Peng, Y.; Peng, Y.; Peng, J.; Jiang, S. miR-135a-5p inhibits 3T3-L1 adipogenesis through activation of canonical Wnt/beta-catenin signaling. J. Mol. Endocrinol. 2014, 52, 311–320. [Google Scholar] [CrossRef]
- Chen, H.; Wang, S.; Chen, L.; Chen, Y.; Wu, M.; Zhang, Y.; Yu, K.; Huang, Z.; Qin, L.; Mo, D. MicroRNA-344 inhibits 3T3-L1 cell differentiation via targeting GSK3beta of Wnt/beta-catenin signaling pathway. FEBS Lett. 2014, 588, 429–435. [Google Scholar] [CrossRef]
- Xu, Y.; Lu, S. Regulation of beta-catenin-mediated esophageal cancer growth and invasion by miR-214. Am J. Transl. Res. 2015, 7, 2316–2325. [Google Scholar]
- Chandrasekaran, K.S.; Sathyanarayanan, A.; Karunagaran, D. miR-214 activates TP53 but suppresses the expression of RELA, CTNNB1, and STAT3 in human cervical and colorectal cancer cells. Cell Biochem. Funct. 2017, 35, 464–471. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| PPARγ | CGCTGATGCACTGCCTATGA | AGAGGTCCACAGAGCTGATTCC |
| aP2 | CGATCCCAATGAGCAAGTGG | TGGGTCAAGCAACTCTGGAT |
| Adiponectin | GGCAGGAAAGGAGAACCTGG | AGCCTTGTCCTTCTTGAAGAG |
| CDK4 | AGTTTCTAAGCGGCCTGGAT | AACTTCAGGAGCTCGGTACC |
| Cyclin B | AACTTCAGCCTGGGTCG | CAGGGAGTCTTCACTGTAGGA |
| Cyclin E | GCTTGCTCCGGGGATGAAAT | GCGAGGACACCATAAGGAAATCTG |
| Cyclin D | TAGGCCCTCAGCCTCACTC | CCACCCCTGGGATAAAGCAC |
| Ctnnb1 | TCCCATCCACGCAGTTTGAC | TCCTCATCGTTTAGCAGTTTTGT |
| β-actin | GTCCCTGACCCTCCCAAAAG | GCTGCCTCAACACCTCAACCC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xi, F.-x.; Wei, C.-s.; Xu, Y.-t.; Ma, L.; He, Y.-l.; Shi, X.-e.; Yang, G.-s.; Yu, T.-y. MicroRNA-214-3p Targeting Ctnnb1 Promotes 3T3-L1 Preadipocyte Differentiation by Interfering with the Wnt/β-Catenin Signaling Pathway. Int. J. Mol. Sci. 2019, 20, 1816. https://doi.org/10.3390/ijms20081816
Xi F-x, Wei C-s, Xu Y-t, Ma L, He Y-l, Shi X-e, Yang G-s, Yu T-y. MicroRNA-214-3p Targeting Ctnnb1 Promotes 3T3-L1 Preadipocyte Differentiation by Interfering with the Wnt/β-Catenin Signaling Pathway. International Journal of Molecular Sciences. 2019; 20(8):1816. https://doi.org/10.3390/ijms20081816
Chicago/Turabian StyleXi, Feng-xue, Chang-sheng Wei, Yan-ting Xu, Lu Ma, Yu-lin He, Xin-e Shi, Gong-she Yang, and Tai-yong Yu. 2019. "MicroRNA-214-3p Targeting Ctnnb1 Promotes 3T3-L1 Preadipocyte Differentiation by Interfering with the Wnt/β-Catenin Signaling Pathway" International Journal of Molecular Sciences 20, no. 8: 1816. https://doi.org/10.3390/ijms20081816
APA StyleXi, F. -x., Wei, C. -s., Xu, Y. -t., Ma, L., He, Y. -l., Shi, X. -e., Yang, G. -s., & Yu, T. -y. (2019). MicroRNA-214-3p Targeting Ctnnb1 Promotes 3T3-L1 Preadipocyte Differentiation by Interfering with the Wnt/β-Catenin Signaling Pathway. International Journal of Molecular Sciences, 20(8), 1816. https://doi.org/10.3390/ijms20081816