Therapeutic Effect of Dipsacus asperoides C. Y. Cheng et T. M. Ai in Ovalbumin-Induced Murine Model of Asthma

 ,
, 
 and
and 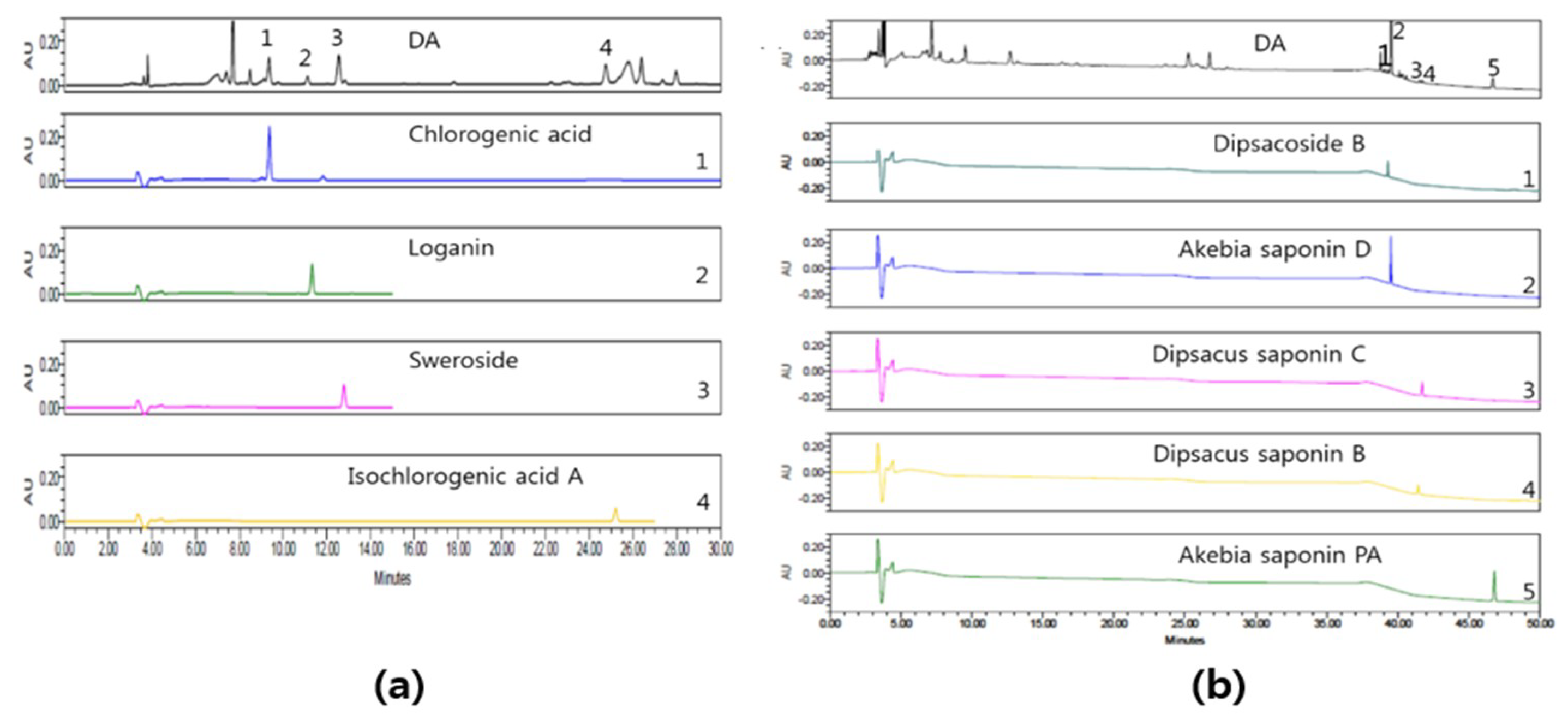{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. HPLC Analysis of DA
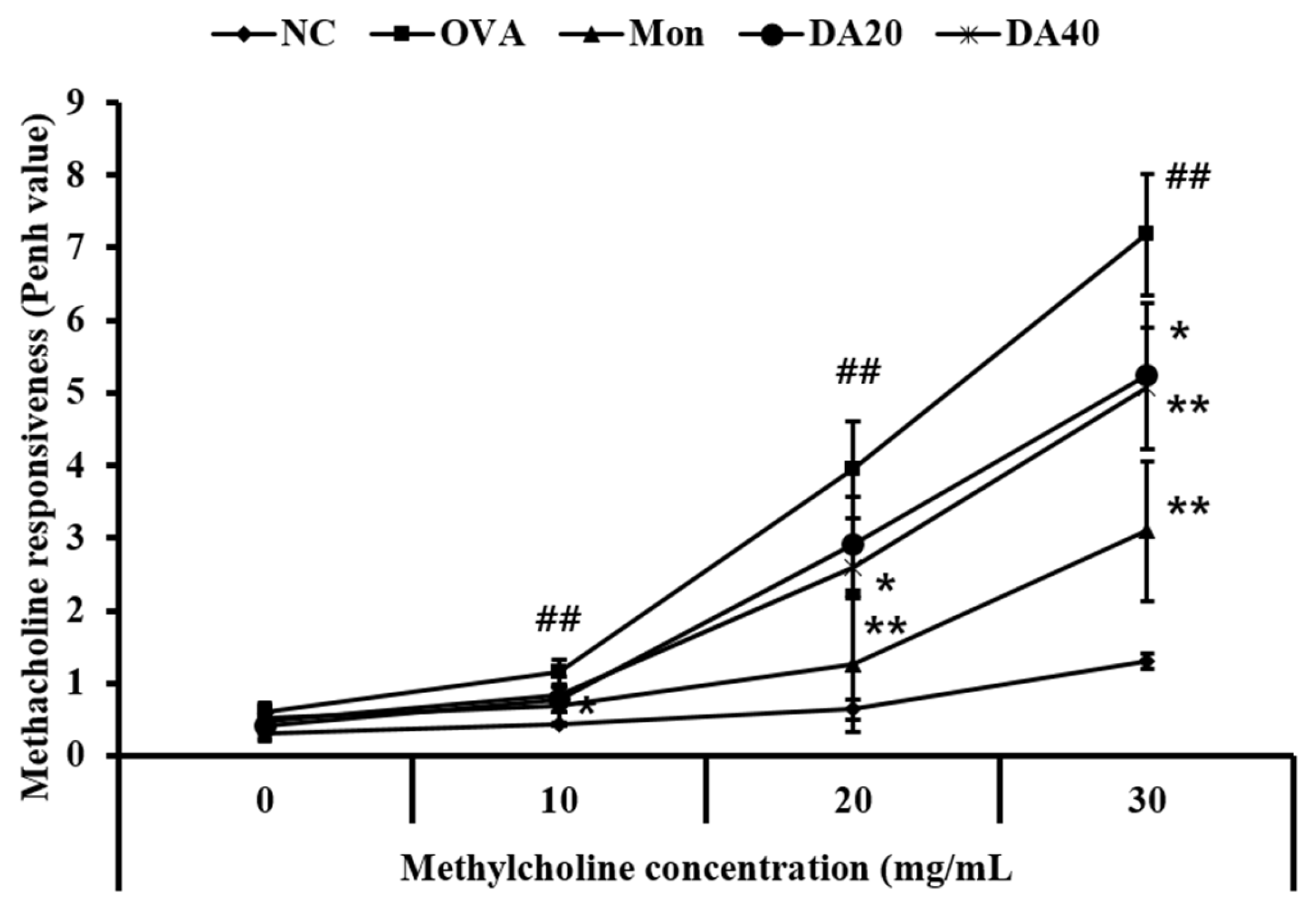
2.2. DA Reduces Methacholine Responsiveness in OVA-Induced Asthma Model
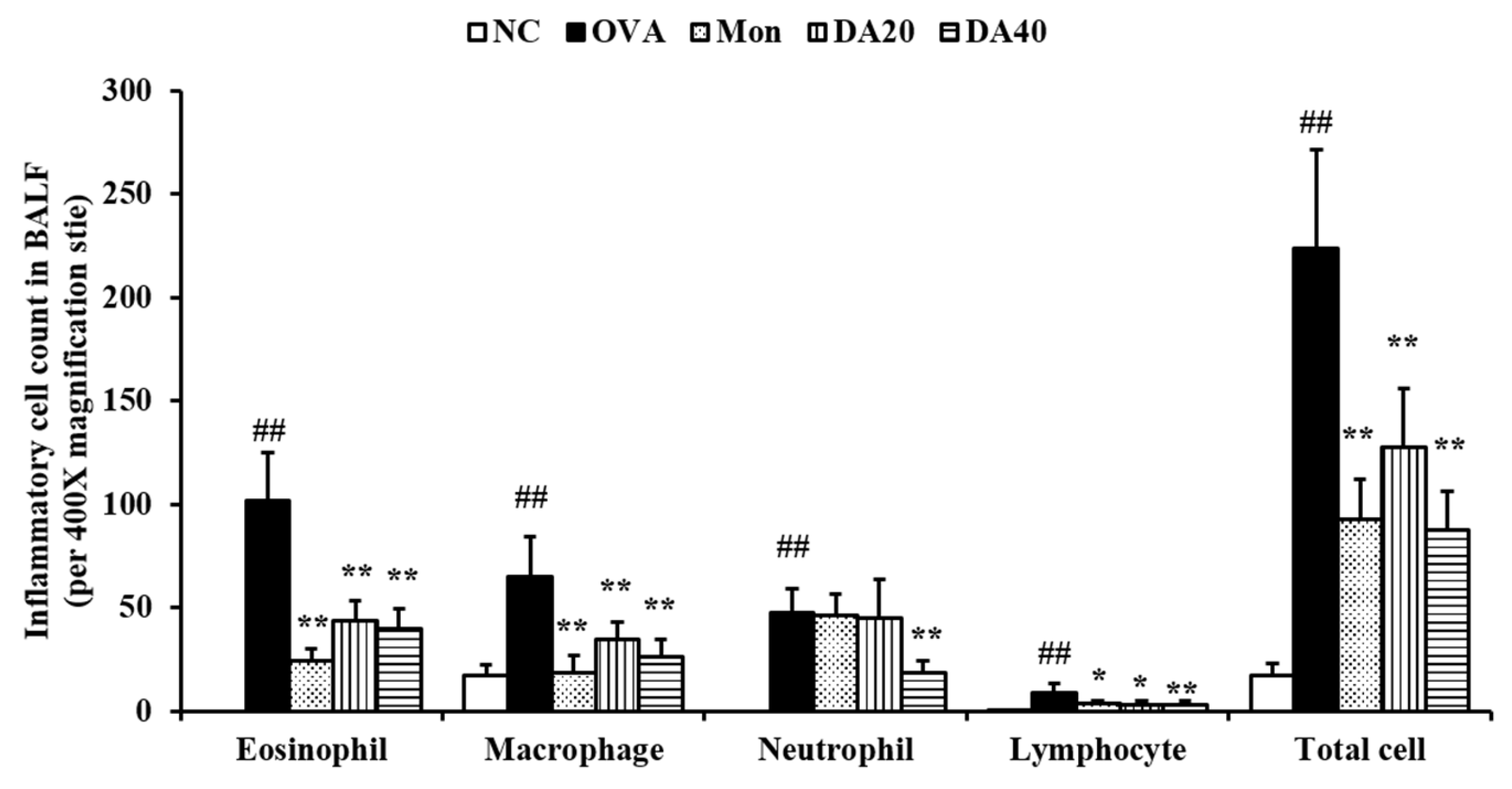
2.3. DA Suppress the Inflammatory Cell Counts in BALF from OVA-Induced Asthma Model
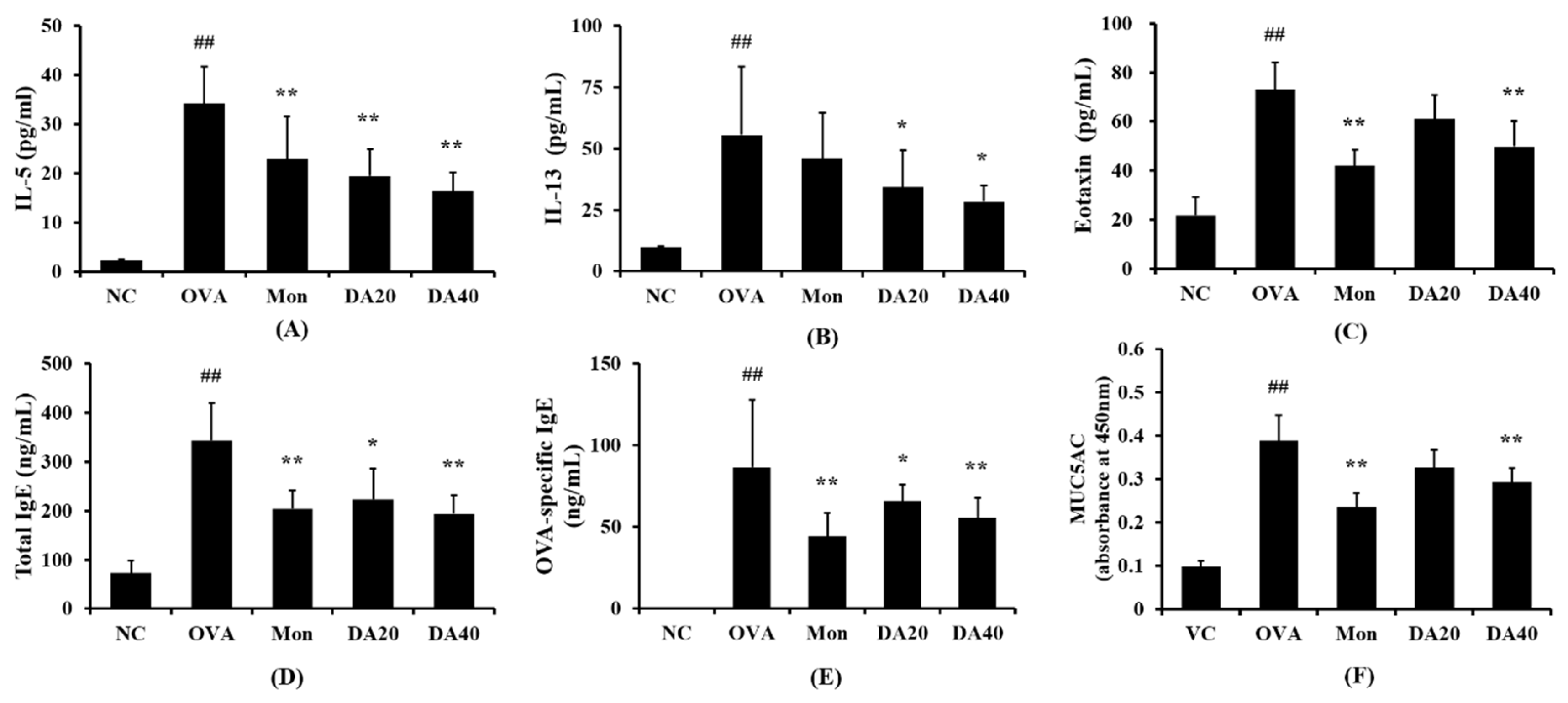
2.4. DA Reduced The Levels of IL-5, IL-13, Eotaxin, and Total Ig E in OVA-Induced Asthma Model
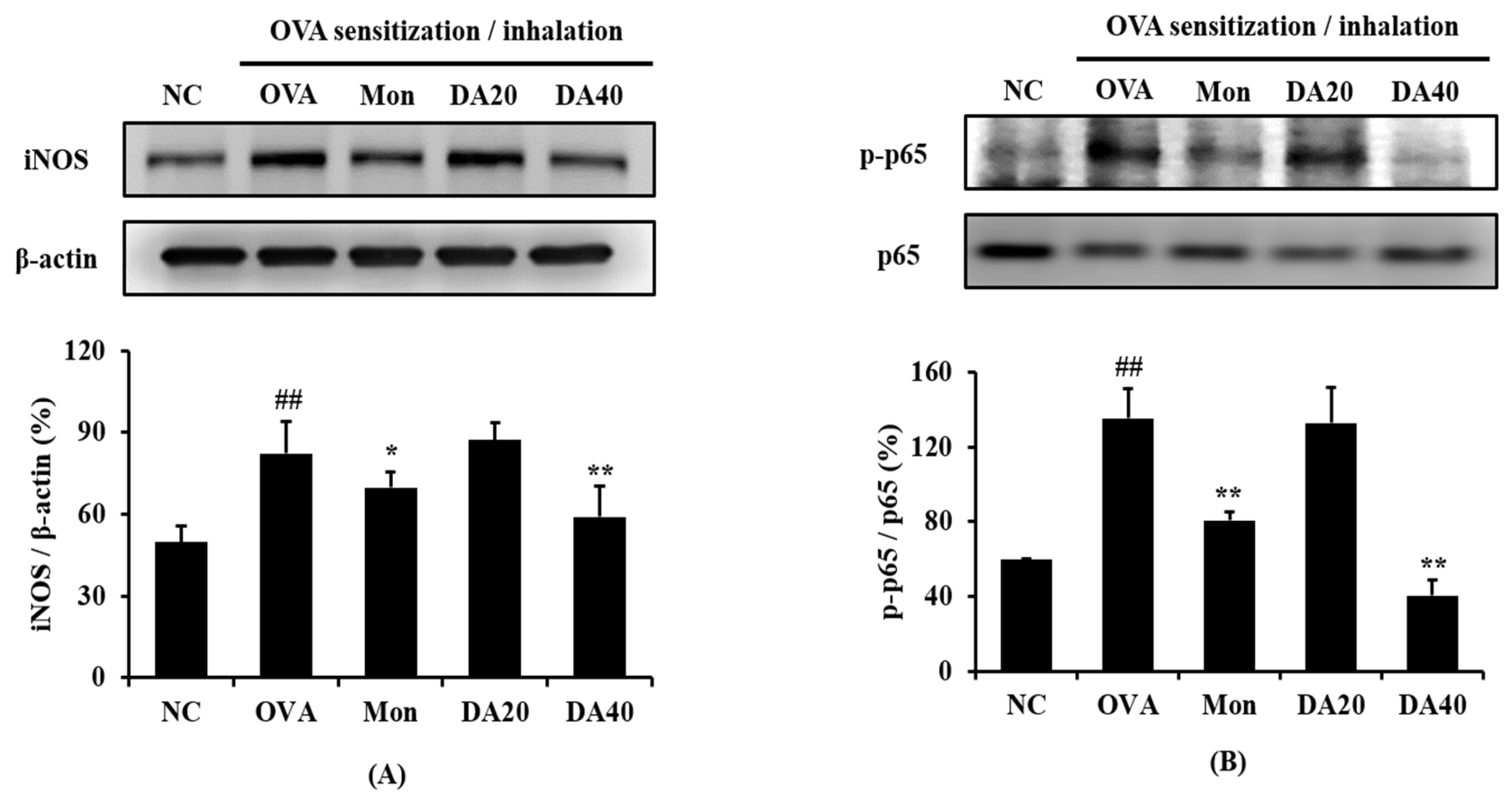
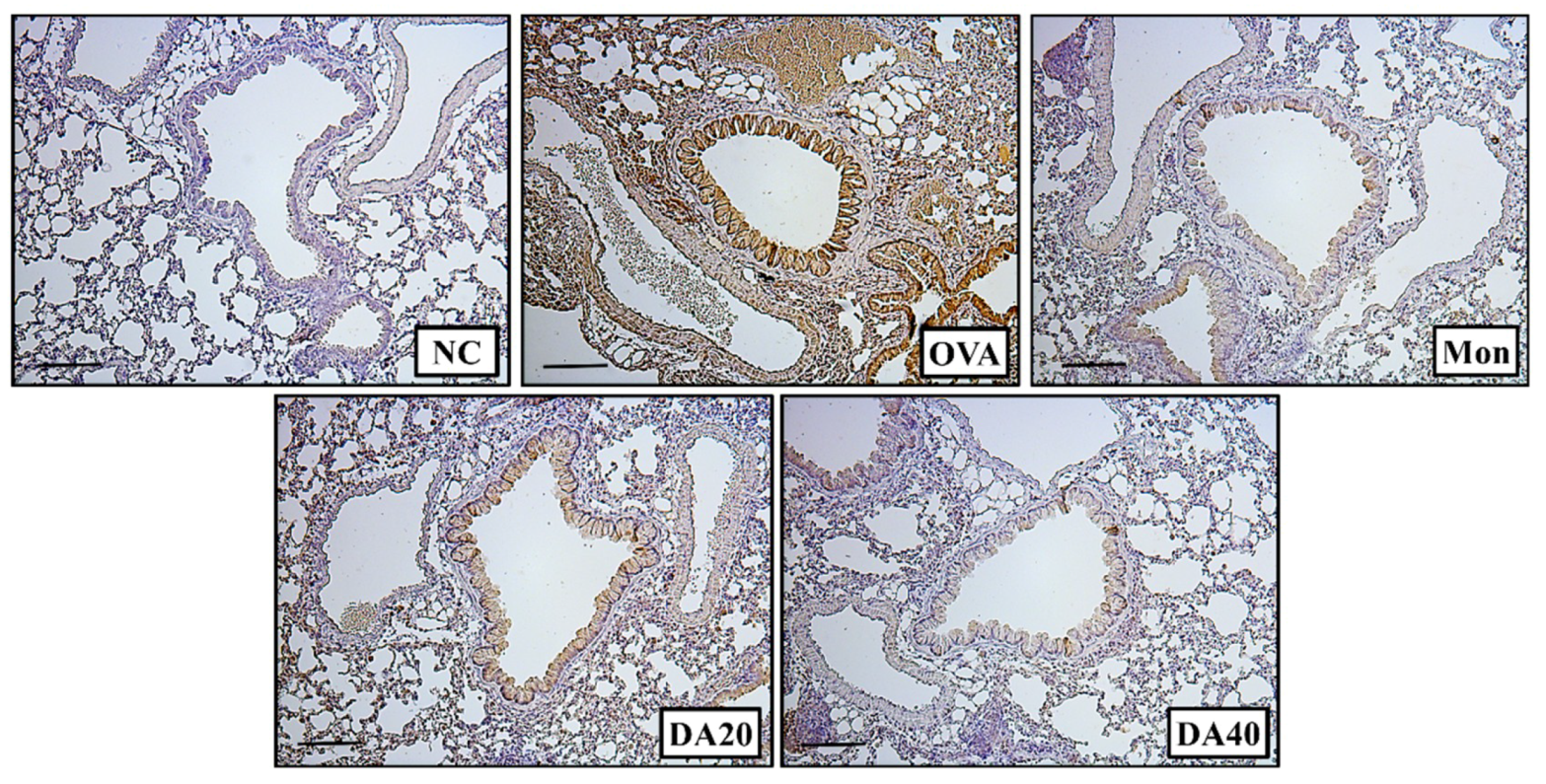
2.5. DA Decreased Expression of iNOS and Phosphorylation of NF-κB in OVA-Induced Asthma Model
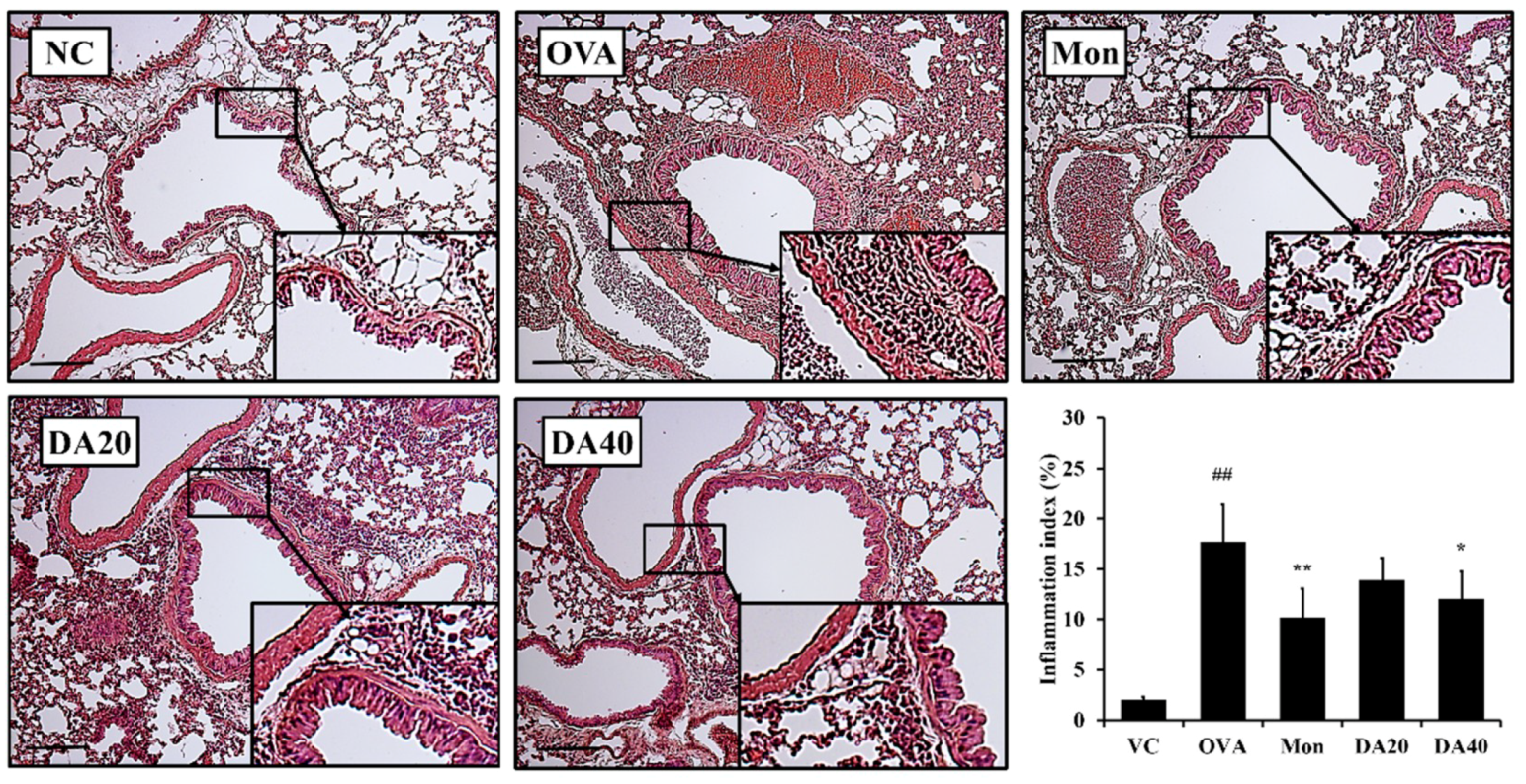
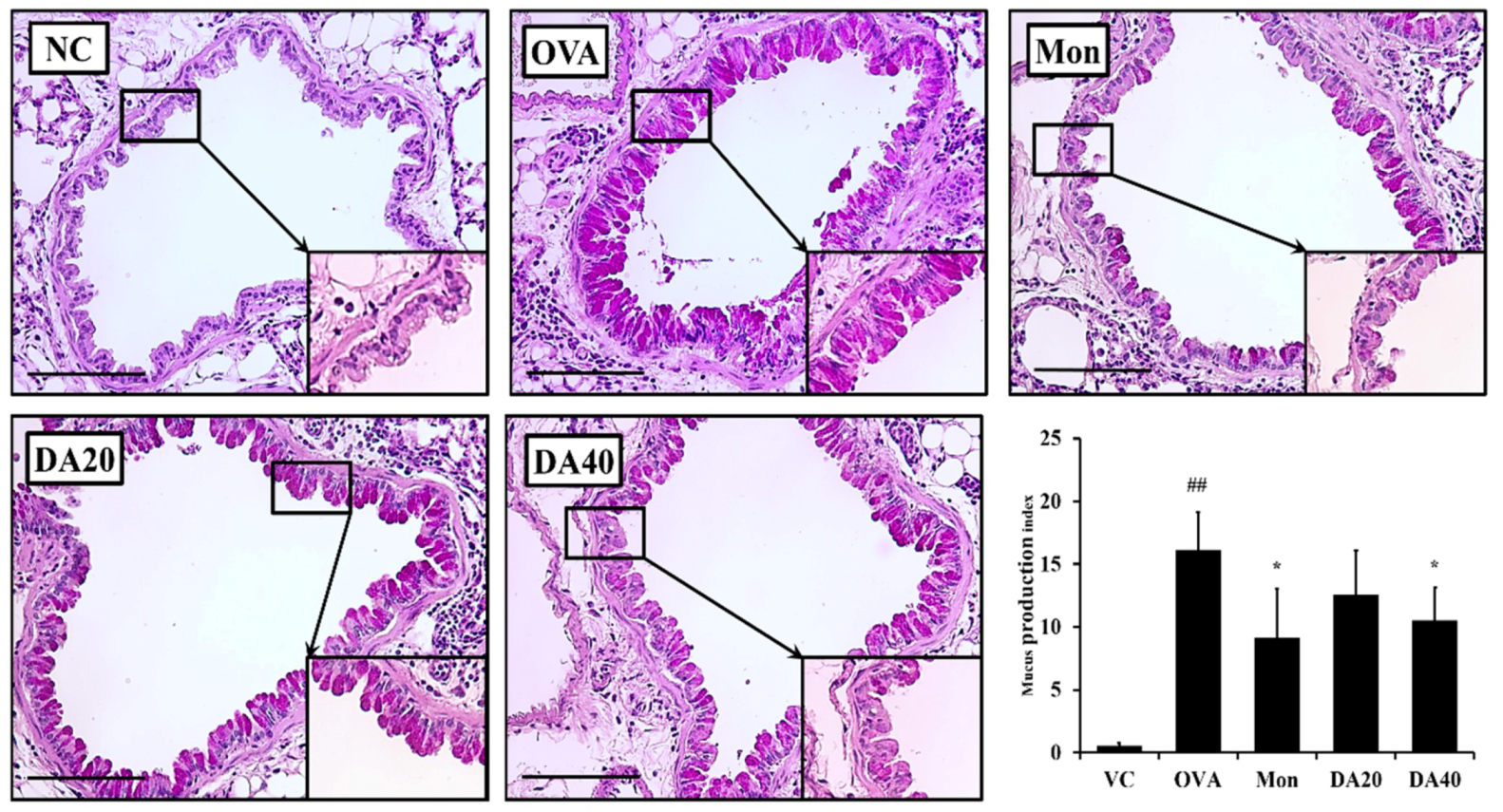
2.6. DA-Reduced Mucus Production in OVA-Induced Asthma Model
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Chemicals
4.2. High-Performance Liquid Chromatography (HPLC) Analysis
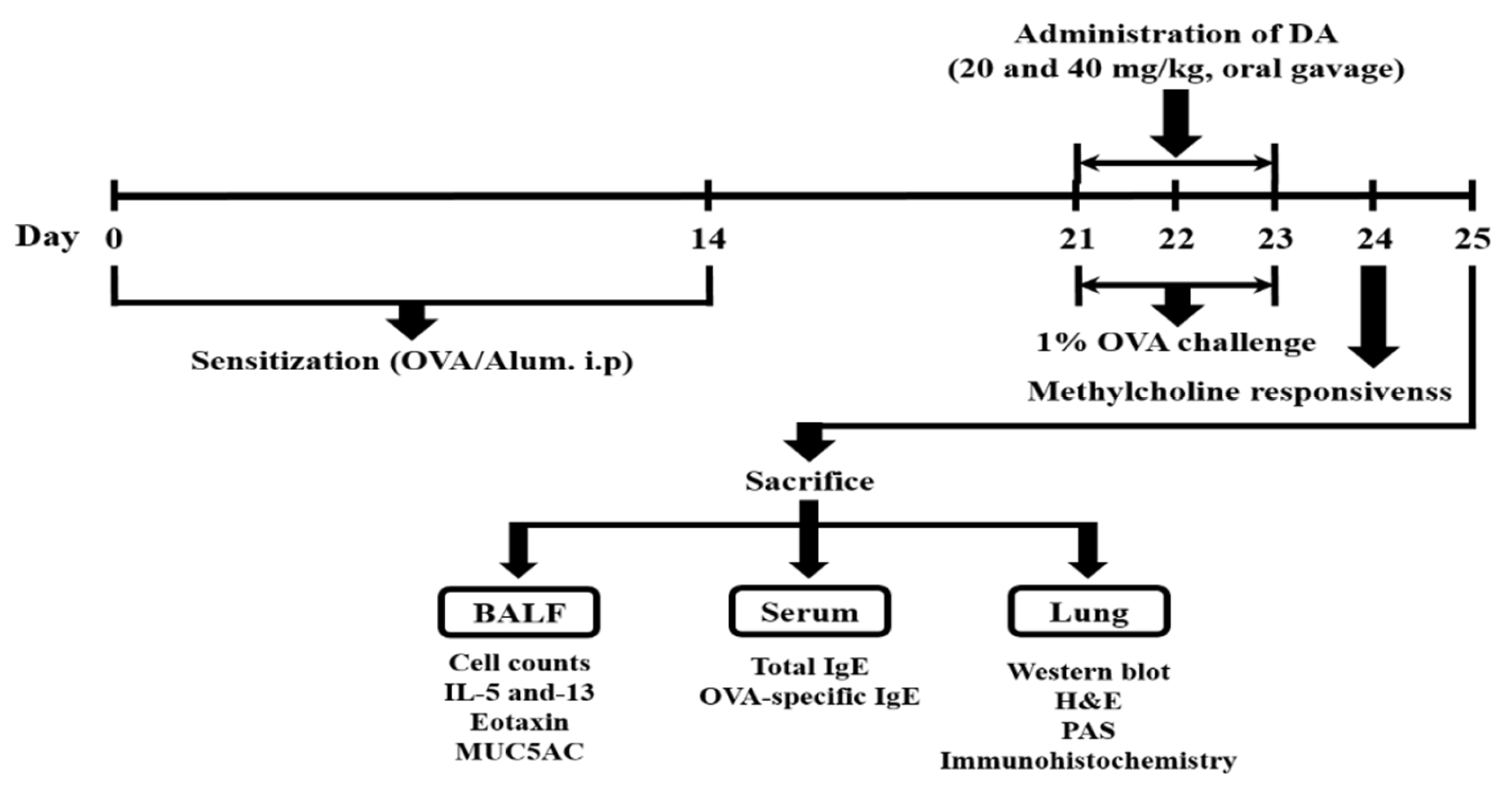
4.3. Experimental Procedure
4.4. BALF and Serum Analysis
4.5. Immunoblotting
4.6. Histopathological Analysis
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Fehrenbach, H.; Wagner, C.; Wegmann, M. Airway remodeling in asthma: What really matters. Cell Tissue Res. 2017, 367, 551–569. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kim, B.K.; Lee, Y.C. Effects of Corni fructus on ovalbumin-induced airway inflammation and airway hyper-responsiveness in a mouse model of allergic asthma. J. Inflamm. 2012, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Barrett, N.A.; Austen, K.F. Innate cells and T helper 2 cell immunity in airway inflammation. Immunity 2009, 31, 425–437. [Google Scholar] [CrossRef]
- Possa, S.S.; Leick, E.A.; Prado, C.M.; Martins, M.A.; Tibério, I.F. Eosinophilic inflammation in allergic asthma. Front. Pharmacol. 2013, 4, 46. [Google Scholar] [CrossRef]
- Coyle, A.J.; Wagner, K.; Bertrand, C.; Tsuyuki, S.; Bews, J.; Heusser, C. Central role of immunoglobulin (Ig) E in the induction of lung eosinophil infiltration and T helper 2 cell cytokine production: Inhibition by a non-anaphylactogenic anti-IgE antibody. J. Exp. Med. 1996, 183, 1303–1310. [Google Scholar] [CrossRef]
- Mahajan, S.; Mehta, A. Role of Cytokines in Pathophysiology of Asthma. Iranian J. Pharmacology Therapeutics 2006, 5, 1–14. [Google Scholar]
- Lawrence, M.G.; Steinke, J.W.; Borish, L. Cytokine-targeting biologics for allergic diseases. Ann. Allergy Asthma. Immunol. 2018, 120, 379–381. [Google Scholar]
- Ghosh, S.; Erzurum, S.C. Nitric oxide metabolism in asthma pathophysiology. Biochim. Biophysica Acta 2011, 1810, 1008–1016. [Google Scholar] [CrossRef]
- Lowry, J.L.; Brovkovych, V.; Zhang, Y.; Skidgel, R.A. Endothelial nitric-oxide synthase activation generates an inducible nitric-oxide synthase-like output of nitric oxide in inflamed endothelium. J. Biol. Chem. 2013, 288, 4174–4193. [Google Scholar] [CrossRef]
- Prado, C.M.; Leick-Maldonado, E.A.; Yano, L.; Leme, A.S.; Capelozzi, V.L.; Martins, M.A.; Tiberio, I.F. Effects of nitric oxide synthases in chronic allergic airway inflammation and remodeling. Am. J. Respir. Cell. Mol. Mol. 2006, 35, 457–465. [Google Scholar] [CrossRef]
- Jeon, W.Y.; Shin, I.S.; Shin, H.K.; Lee, M.Y. Samsoeum water extract attenuates allergic airway inflammation via modulation of Th1/Th2 cytokines and decrease of iNOS expression in asthmatic mice. BMC Complement. Altern. Med. 2015, 15, 47. [Google Scholar] [CrossRef]
- Bao, Z.; Zhang, P.; Yao, Y.; Lu, G.; Tong, Z.; Yan, B.; Tu, L.; Yang, G.; Zhou, J. Deguelin Attenuates Allergic Airway Inflammation via Inhibition of NF-κb Pathway in Mice. Int. J. Biol. Sci. 2017, 13, 492–504. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Zhang, Q.; Du, Q.; Shen, H.; Zhu, Z. Pinocembrin attenuates allergic airway inflammation via inhibition of NF-κB pathway in mice. Int. Immunopharmacol. 2017, 53, 90–95. [Google Scholar] [CrossRef]
- Bui, T.T.; Piao, C.H.; Song, C.H.; Shin, H.S.; Chai, O.H. Bupleurum chinense extract ameliorates an OVA-induced murine allergic asthma through the reduction of the Th2 and Th17 cytokines production by inactivation of NFκB pathway. Biomed. Pharmacother. 2017, 91, 1085–1095. [Google Scholar] [CrossRef]
- Inam, A.; Shahzad, M.; Shabbir, A.; Shahid, H.; Shahid, K.; Javeed, A. Carica papaya ameliorates allergic asthma via down regulation of IL-4, IL-5, eotaxin, TNF-α, NF-ĸB, and iNOS levels. Phytomedicine 2017, 32, 1–7. [Google Scholar] [CrossRef]
- Hung, T.M.; Na, M.K.; Thuong, P.T.; Su, N.D.; Sok, D.E.; Song, K.S.; Seong, Y.H.; Bae, K.H. Antioxidant activity of caffeoyl quinic acid derivatives from the roots of Dipsacus asper Wall. J. Ethnopharmacol. 2006, 108, 188–192. [Google Scholar] [CrossRef]
- Liu, Z.G.; Zhang, R.; Li, C.; Ma, X.; Liu, L.; Wang, J.P.; Mei, Q.B. The osteroprotective effect of Radix Dipsaci extract in ovariectomized rate. J. Ethnopharmacol. 2009, 123, 74–81. [Google Scholar] [CrossRef]
- Chen, J.; Yao, D.; Yuan, H.; Zhang, S.; Tian, J.; Guo, W.; Liang, W.; Li, H.; Zhang, Y. Dipsacus asperoides polysaccharide induces apoptosis in osteosarcoma cells by modulating the PI3K/Akt pathway. Carbohydr. Polym. 2013, 95, 780–784. [Google Scholar] [CrossRef]
- Cong, G.; Cui, L.; Zang, M.; Hao, L. Attenuation of renal ischemia/reperfusion injury by a polysaccharide from the roots of Dipsacus asperoides. Int. J. Biol. Macromol. 2013, 56, 14–19. [Google Scholar] [CrossRef]
- Huang, Y.X.; Liu, E.W.; Wang, L.; Huo, Y.; Wang, Q.; Olaleye, O.; Wang, T.; Gao, X.M. LC/MS/MS determination and pharmacokinetic studies of six compounds in rat plasma following oral administration of the single and combined extracts of Eucommia ulmoides and Dipsacus asperoides. Chin. J. Nat. Med. 2014, 12, 469–476. [Google Scholar] [CrossRef]
- Niu, Y.B.; Kong, X.H.; Li, Y.H.; Fan, L.; Pan, Y.L.; Li, C.R.; Wu, X.L.; Mei, Q.B. Radix Dipsaci total saponins stimulate MC3T3-E1 cell differentiation via the bone morphogenetic protein-2/MAPK/Smad-dependent Runx2 pathway. Mol. Med. Rep. 2015, 11, 4468–4472. [Google Scholar] [CrossRef]
- Niu, Y.; Li, C.; Pan, Y.; Li, Y.; Kong, X.; Wang, S.; Zhai, Y.; Wu, X.; Fan, W.; Mei, Q. Treatment of Radix Dipsaci extract prevents long bone loss induced by modeled microgravity in hindlimb unloading rats. Pharm. Biol. 2015, 53, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.W.; Jung, J.K.; Son, K.H.; Lee, D.H.; Kang, T.M.; Kim, Y.S.; Park, Y.K. Inhibitory effects of the root extract of Dipsacus asperoides C.Y. Cheng et al. T.M.Ai on collagen-induced arthritis in mice. J. Ethnopharmacol. 2012, 139, 98–130. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Dong, X.; Ma, H.; Cui, Y.; Tong, J. Purification, characterisation and protective effects of polysaccharidesfrom alfalfa on hepatocytes. Carbohydr. Polym. 2014, 112, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.H.; Zhang, D.; Yu, B.; Zhao, S.P.; Wang, J.W.; Yao, L.; Cao, W.G. Antioxidant activity and optimization of extraction of polysaccharide from the roots of Dipsacus asperoides. Int. J. Biol. Macromol. 2015, 81, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Park, S.D.; Koh, Y.J.; Kim, D.I.; Lee, J.H. Aqueous extract of Dipsacus asperoides suppresses lipopolysaccharide-stimulated inflammatory responses by inhibiting the ERK1/2 signaling pathway in RAW 264.7 macrophages. J. Ethnopharmacol. 2019, 231, 253–261. [Google Scholar] [CrossRef]
- Ji, D.; Zhang, C.; Li, J.; Yang, H.; Shen, J.; Yang, Z. A new iridoid glycoside from the roots of Dipsacus asper. Molecules 2012, 17, 1419–1424. [Google Scholar] [CrossRef]
- Yu, X.; Wang, L.N.; Du, Q.M.; Ma, L.; Chen, L.; You, R.; Liu, L.; Ling, J.J.; Yang, Z.L.; Ji, H. Akebia Saponin D attenuates amyloid β-induced cognitive deficits and inflammatory response in rats: Involvement of Akt/NF-κB pathway. Behav. Brain. Res. 2012, 235, 200–209. [Google Scholar] [CrossRef]
- Li, C.; Gao, Y.; Tian, J.; Xing, Y.; Zhu, H.; Shen, J. Long-term oral Asperosaponin VI attenuates cardiac dysfunction, myocardial fibrosis in a rat model of chronic myocardial infarction. Food Chem. Toxicol. 2012, 50, 1432–1438. [Google Scholar] [CrossRef]
- Gong, L.L.; Yang, S.; Liu, H.; Zhang, W.; Ren, L.L.; Han, F.F.; Lv, Y.L.; Wan, Z.R.; Liu, L.H. Anti-nociceptive and anti-inflammatory potentials of Akebia saponin D. Eur. J. Pharmacol. 2018, 845, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Pawankar, R. Allergic diseases and asthma: A global public health concern and a call to action. World Allergy Organ. J. 2014, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, D.K.; Shao, Z. Pathogenesis of allergic airway inflammation. Curr. Allergy Asthma Rep. 2010, 10, 39–48. [Google Scholar] [CrossRef]
- Moldoveanu, B.; Otmishi, P.; Jani, P.; Walker, J.; Sarmiento, X.; Guardiola, J.; Saad, M.; Yu, J. Inflammatory mechanisms in the lung. J. Inflamm. Res. 2009, 2, 1–11. [Google Scholar] [PubMed]
- Batra, J.; Chatterjee, R.; Ghosh, B. Inducible nitric oxide synthase (iNOS): Role in asthma pathogenesis. Indian J. Biochem. Biophysics 2007, 44, 303–309. [Google Scholar]
- Shin, I.S.; Shin, N.R.; Jeon, C.M.; Kwon, O.K.; Hong, J.M.; Kim, H.S.; Oh, S.R.; Ahn, K.S. Thuja orientalis reduces airway inflammation in ovalbumin-induced allergic asthma. Mol. Med. Rep. 2015, 12, 4640–4646. [Google Scholar] [CrossRef]
- Yadav, U.C.; Naura, A.S.; Aguilera-Aguirre, L.; Ramana, K.V.; Boldogh, I.; Sur, S.; Boulares, H.A.; Srivastava, S.K. Aldose reductase inhibition suppresses the expression of Th2 cytokines and airway inflammation in ovalbumin-induced asthma in mice. J. Immunol. 2009, 183, 4723–4732. [Google Scholar] [CrossRef]
- Shin, N.R.; Ko, J.W.; Park, S.H.; Cho, Y.K.; Oh, S.R.; Ahn, K.S.; Ryu, J.M.; Kim, J.C.; Seo, C.S.; Shin, I.S. Protective effect of HwangRyunHaeDok-Tang water extract against chronic obstructive pulmonary disease induced by cigarette smoke and lipopolysaccharide in a mouse model. J. Ethnopharmacol. 2017, 200, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.S.; Lee, B.S.; Han, S.R.; Han, H.Y.; Chung, M.K.; Min, B.S.; Seok, J.H.; Kim, Y.B. A subchronic toxicity study of Radix Dipsaci water extract by oral administration in F344 rats. Regul. Toxicol. Pharmacol. 2016, 81, 136–145. [Google Scholar]
- Melgert, B.N.; Postma, D.S.; Kuipers, I.; Geerlings, M.; Luinge, M.A.; van der Strate, B.W.; Kerstjens, H.A.; Timens, W.; Hylkema, M.N. Female mice are more susceptible to the development of allergic airway inflammation than male mice. Clin. Exp. Allergy 2005, 35, 1496–1503. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Tanabe, M.; Ito, W.; Ueki, S.; Konnno, Y.; Chihara, M.; Itoga, M.; Kobayashi, Y.; Moritoki, Y.; Kayaba, H.; et al. Gender difference in allergic airway remodeling and immunoglobulin production in mouse model of asthma. Respirology 2013, 18, 797–806. [Google Scholar] [CrossRef]
- Shin, I.S.; Shin, N.R.; Jeon, C.M.; Kwon, O.K.; Sohn, K.Y.; Lee, T.S.; Kim, J.W.; Ahn, K.S.; Oh, S.R. EC-18, a synthetic monoacetyldiglyceride (1-palmitoyl-2-linoleoyl-3-acetylglycerol), attenuates the asthmatic response in an aluminum hydroxide/ovalbumin-induced model of asthma. Int. Immunopharmacol. 2014, 18, 116–123. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, N.-R.; Lee, A.Y.; Park, G.; Ko, J.-W.; Kim, J.-C.; Shin, I.-S.; Kim, J.-S. Therapeutic Effect of Dipsacus asperoides C. Y. Cheng et T. M. Ai in Ovalbumin-Induced Murine Model of Asthma. Int. J. Mol. Sci. 2019, 20, 1855. https://doi.org/10.3390/ijms20081855
Shin N-R, Lee AY, Park G, Ko J-W, Kim J-C, Shin I-S, Kim J-S. Therapeutic Effect of Dipsacus asperoides C. Y. Cheng et T. M. Ai in Ovalbumin-Induced Murine Model of Asthma. International Journal of Molecular Sciences. 2019; 20(8):1855. https://doi.org/10.3390/ijms20081855
Chicago/Turabian StyleShin, Na-Rae, A Yeong Lee, Gunhyuk Park, Je-Won Ko, Jong-Choon Kim, In-Sik Shin, and Joong-Sun Kim. 2019. "Therapeutic Effect of Dipsacus asperoides C. Y. Cheng et T. M. Ai in Ovalbumin-Induced Murine Model of Asthma" International Journal of Molecular Sciences 20, no. 8: 1855. https://doi.org/10.3390/ijms20081855
APA StyleShin, N. -R., Lee, A. Y., Park, G., Ko, J. -W., Kim, J. -C., Shin, I. -S., & Kim, J. -S. (2019). Therapeutic Effect of Dipsacus asperoides C. Y. Cheng et T. M. Ai in Ovalbumin-Induced Murine Model of Asthma. International Journal of Molecular Sciences, 20(8), 1855. https://doi.org/10.3390/ijms20081855