Inhibitory Effect of Berberine on Broiler P-glycoprotein Expression and Function: In Situ and In Vitro Studies

 ,
,  and
and
Abstract
:
1. Introduction
2. Results
2.1. Effect of Berberine on Abcb1 and CXR mRNA Expression in Broilers
2.2. Berberine Affected Rho123 Uptake in the Jejunum of Broilers by In Situ Perfusion
2.3. Effect of Berberine on P-gp Medicated Rho123 Efflux in MDCK-chAbcb1 Cells
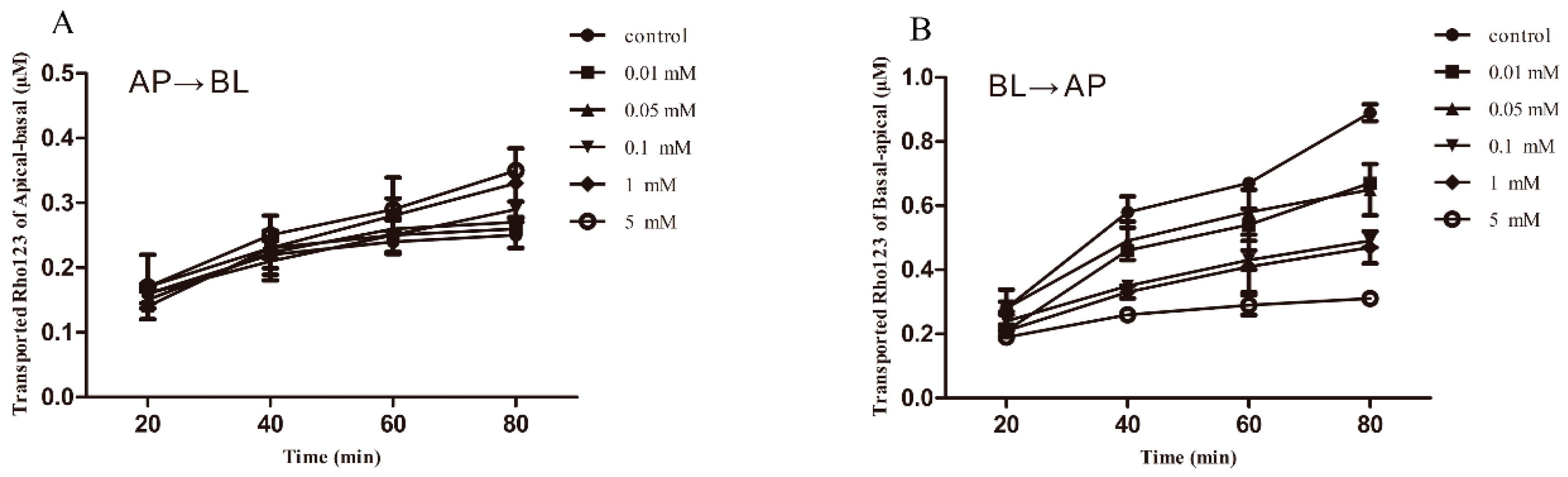
2.4. Transporting Interactions of Berberine with Veterinary Drugs in MDCK-chAbcb1 Cells
2.5. Berberine Is a Substrate for Chicken P-gp Proved Based on a Bidirectional Transport Assay in MDCK chAbcb1 Cells
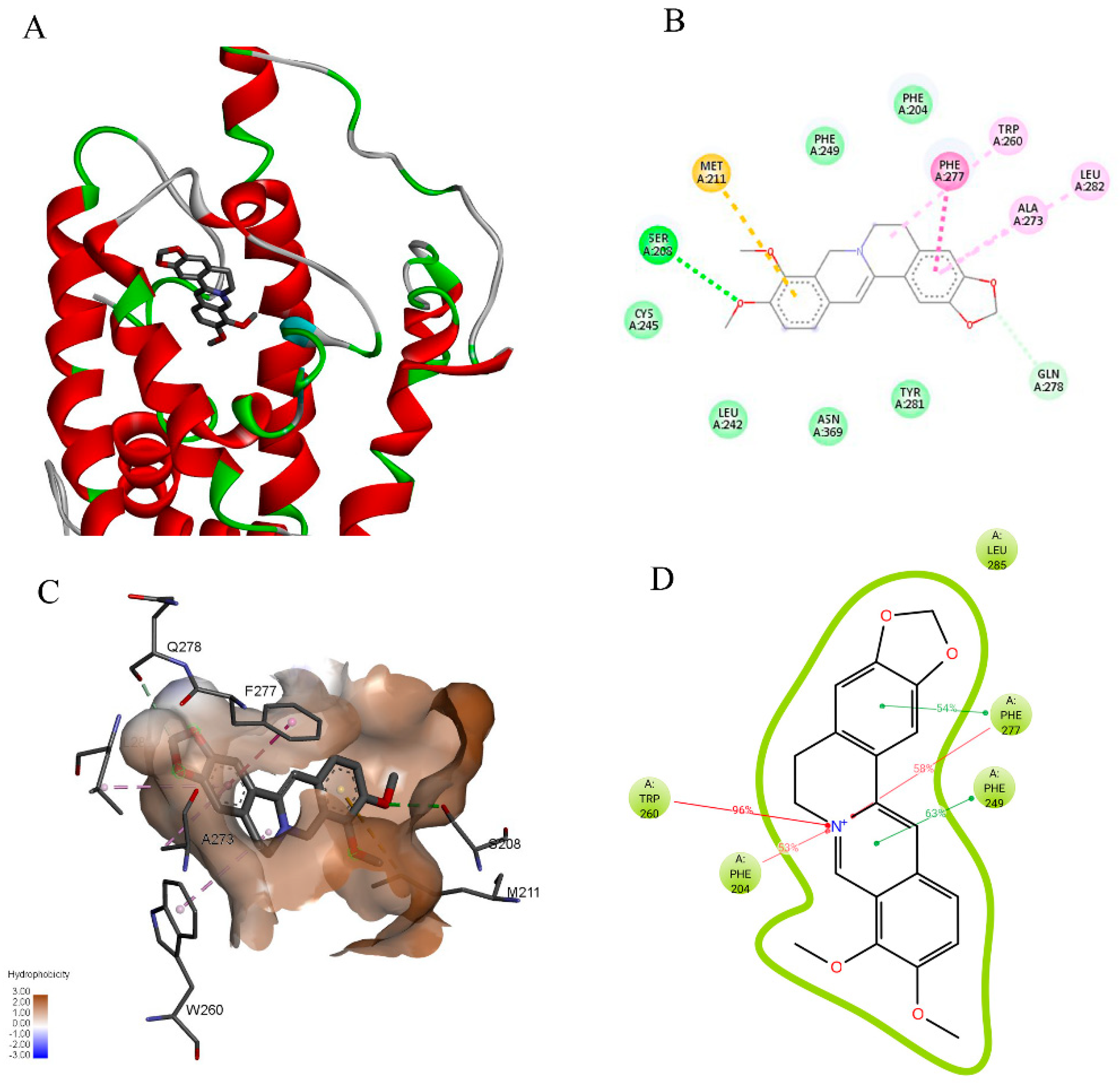
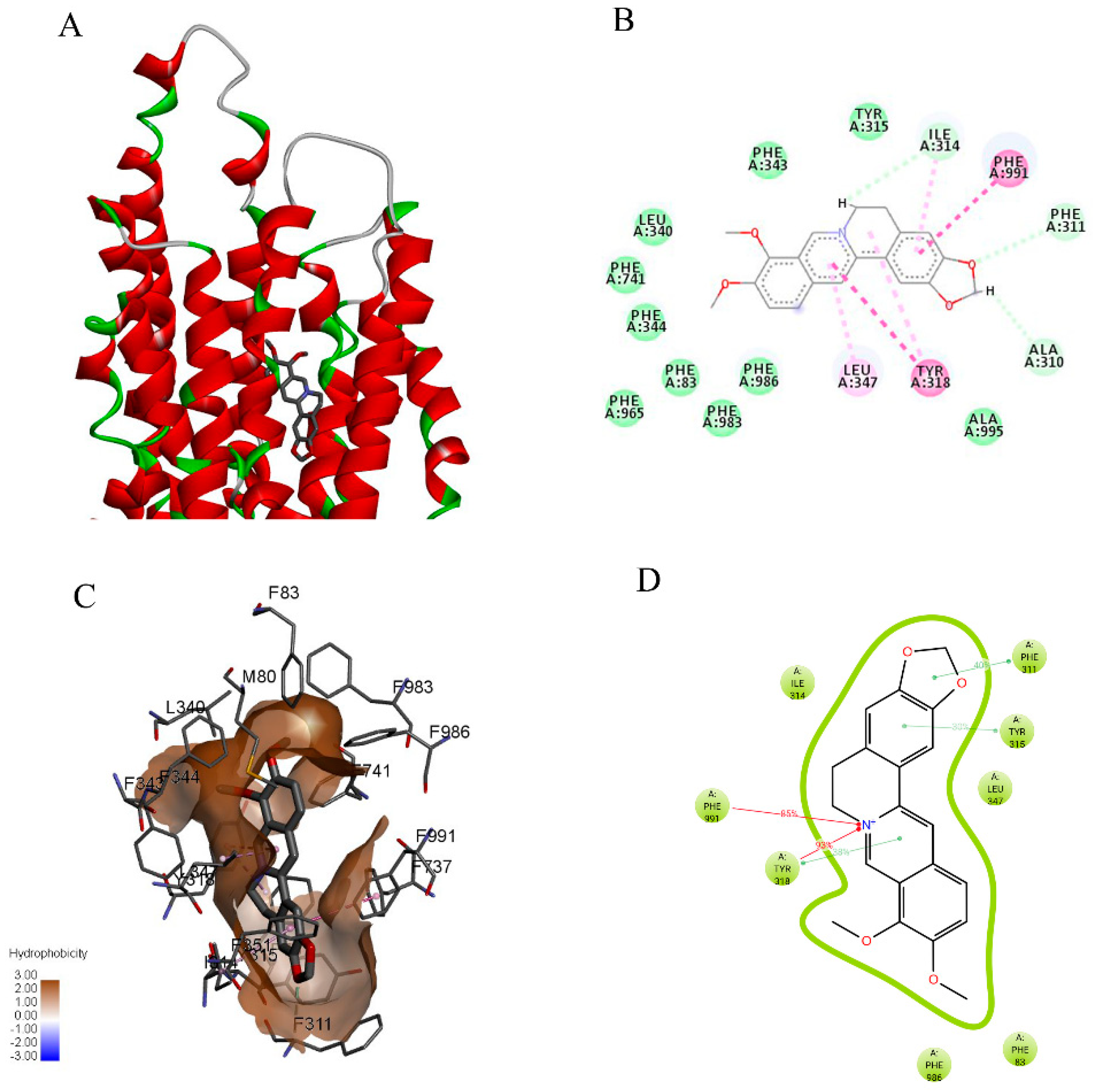
2.6. Berberine might Favorably Interact with P-gp and CXR, as Analyzed by Molecular Docking Modeling
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Line and Animals
4.3. Effect of Berberine on Abcb1 and Chicken Xenobiotic Receptor mRNA Expression in the Ileum of Broilers Determined by Real-Time RT-PCR
4.4. In Situ Single-Pass Intestinal Perfusion in Berberine-Treated Broilers
4.5. Bidirectional Transport Experiments Using MDCK-Abcb1 Cells
4.6. Molecular Modelling
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Otani, M.; Shitan, N.; Sakai, K.; Martinoia, E.; Sato, F.; Yazaki, K. Characterization of vacuolar transport of the endogenous alkaloid berberine in Coptis japonica. Plant Physiol. 2005, 138, 1939–1946. [Google Scholar] [CrossRef]
- Luo, J.; Yan, D.; Yang, M.; Dong, X.; Xiao, X. Multicomponent Therapeutics of Berberine Alkaloids. Evid. Based Complement. Altern. Med. 2013, 2013, 545898. [Google Scholar] [CrossRef]
- Shen, Y.B.; Piao, X.S.; Kim, S.W.; Wang, L.; Liu, P. The effects of berberine on the magnitude of the acute inflammatory response induced by Escherichia coli lipopolysaccharide in broiler chickens. Poult Sci. 2010, 89, 13. [Google Scholar] [CrossRef]
- Qiu, W.; Jiang, X.H.; Liu, C.X.; Ju, Y.; Jin, J.X. Effect of berberine on the pharmacokinetics of substrates of CYP3A and P-gp. Phytother. Res. 2009, 23, 1553–1558. [Google Scholar] [CrossRef]
- Shan, Y.Q.; Zhu, Y.P.; Pang, J.; Wang, Y.X.; Song, D.Q.; Kong, W.J.; Jiang, J.D. Tetrandrine potentiates the hypoglycemic efficacy of berberine by inhibiting P-glycoprotein function. Biol. Pharm. Bull. 2013, 36, 1562. [Google Scholar] [CrossRef]
- Montanari, F.; Ecker, G.F. Prediction of drug–ABC-transporter interaction—Recent advances and future challenges. Adv. Drug Deliv. Rev. 2015, 86, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Revalde, J.; Paxton, J.W. The effects of dietary and herbal phytochemicals on drug transporters. Adv. Drug Deliv. Rev. 2017, 116, 45–62. [Google Scholar] [CrossRef] [PubMed]
- Peachey, L.E.; Pinchbeck, G.L.; Matthews, J.B.; Burden, F.A.; Lespine, A.; von Samson-Himmelstjerna, G.; Krücken, J.; Hodgkinson, J.E. P-glycoproteins play a role in ivermectin resistance in cyathostomins. Int. J. Parasitol. 2017, 7, 388–398. [Google Scholar] [CrossRef]
- Guo, T.; Huang, J.; Zhang, H.; Dong, L.; Guo, D.; Guo, L.; He, F.; Bhutto, Z.A.; Wang, L. Abcb1 in Pigs: Molecular cloning, tissues distribution, functional analysis, and its effect on pharmacokinetics of enrofloxacin. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Ballent, M.; Maté, L.; Virkel, G.; Sallovitz, J.; Viviani, P.; Lanusse, C.; Lifschitz, A. Intestinal drug transport: Ex vivo evaluation of the interactions between ABC transporters and anthelmintic molecules. J. Vet. Pharmacol. Ther. 2014, 37, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Kathawala, R.J.; Gupta, P.; Ashby, C.R.; Chen, Z.-S. The modulation of ABC transporter-mediated multidrug resistance in cancer: A review of the past decade. Drug Resist. Updates 2015, 18, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.Q.; Ren, G.; Wang, Y.X.; Pang, J.; Zhao, Z.Y.; Yao, J.; You, X.F.; Si, S.Y.; Song, D.Q.; Kong, W.J.; et al. Berberine analogue IMB-Y53 improves glucose-lowering efficacy by averting cellular efflux especially P-glycoprotein efflux. Metabolism 2013, 62, 446–456. [Google Scholar] [CrossRef]
- Pang, Y.N.; Liang, Y.W.; Feng, T.S.; Zhao, S.; Wu, H.; Chai, Y.S.; Lei, F.; Ding, Y.; Xing, D.M.; Du, L.J. Transportation of berberine into HepG2, HeLa and SY5Y cells: A correlation to its anti-cancer effect. PLoS ONE 2014, 9, e112937. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, J.; Liu, Y.; Guo, T.; Wang, L. Using the lentiviral vector system to stably express chicken P-gp and BCRP in MDCK cells for screening the substrates and studying the interplay of both transporters. Arch. Toxicol. 2018. [Google Scholar] [CrossRef]
- Bhutto, Z.A.; He, F.; Zloh, M.; Yang, J.; Huang, J.; Guo, T.; Wang, L. Use of quercetin in animal feed: Effects on the P-gp expression and pharmacokinetics of orally administrated enrofloxacin in chicken. Sci. Rep. 2018, 8, 4400. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Jin, J.; Zeng, J.; Shi, R.; Dai, Y.; Wu, J.; Li, Y.; Wang, T.; Ma, Y. Involvement of P-glycoprotein and multidrug and toxin extrusion protein 1 in hepatic and renal berberine efflux in mice. RSC Adv. 2017, 7, 34801–34809. [Google Scholar] [CrossRef]
- Zha, W.; Wang, G.; Xu, W.; Liu, X.; Wang, Y.; Zha, B.S.; Shi, J.; Zhao, Q.; Gerk, P.M.; Studer, E.; et al. Inhibition of P-Glycoprotein by HIV Protease Inhibitors Increases Intracellular Accumulation of Berberine in Murine and Human Macrophages. PLOS ONE 2013, 8, e54349. [Google Scholar] [CrossRef] [PubMed]
- David, X.Y.; Zhiyong, H.; Wenyue, W.; Colin, P.; Zhi, C.X. The Effects of Berberine on Clostridium Perfringens Induced Necrotic Enteritis in Broiler Chickens. Arch. Clin. Microbiol. 2017, 8, 3. [Google Scholar]
- Malik, T.A.; Kamili, A.N.; Chishti, M.Z.; Tanveer, S.; Ahad, S.; Johri, R.K. Synergistic approach for treatment of chicken coccidiosis using berberine--A plant natural product. Microb. Pathog. 2016, 93, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.X.; He, Z.; Pouton, C.; Hoerr, F.J.; Xiao, Z.C. Target Animal Safety and Residual Study for Berberine and other Phytogenic Compounds in Broiler Chickens. Arch. Clin. Microbiol. 2017, 8, 69. [Google Scholar]
- Slosky, L.M.; Thompson, B.J.; Sanchez-Covarrubias, L.; Zhang, Y.; Laracuente, M.L.; Vanderah, T.W.; Ronaldson, P.T.; Davis, T.P. Acetaminophen modulates P-glycoprotein functional expression at the blood-brain barrier by a constitutive androstane receptor-dependent mechanism. Mol. Pharmacol. 2013, 84, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Handschin, C.; Podvinec, M.; Meyer, U.A. CXR, a chicken xenobiotic-sensing orphan nuclear receptor, is related to both mammalian pregnane X receptor (PXR) and constitutive androstane receptor (CAR). Proc. Natl. Acad. Sci. USA 2000, 97, 10769–10774. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Tang, Y.; Robbins, G.T.; Nie, D. Camptothecin attenuates cytochrome P450 3A4 induction by blocking the activation of human pregnane X receptor. J. Pharmacol. Exp. Ther. 2010, 334, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.; Safarpour, Y.; Zhang, T.; Guo, P.; Chen, G.; Wu, X.; Fu, Q.; Wang, Y. Berberine Upregulates P-Glycoprotein in Human Caco-2 Cells and in an Experimental Model of Colitis in the Rat via Activation of Nrf2-Dependent Mechanisms. J. Pharmacol. Exp. Ther. 2018, 366, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.M. Expression of P-glycoprotein in the chicken. Comp. Biochem. Physiol. Part A 2001, 130, 301–310. [Google Scholar] [CrossRef]
- Tomita, M.; Kanbayashi, A.; Murata, H.; Tanaka, A.; Nakaike, M.; Hatanaka, M.; Hayashi, M. Effect of lipopolysaccharide on P-glycoprotein-mediated intestinal and biliary excretion of rhodamine123 in rats. Int. J. Pharm. 2010, 392, 35–41. [Google Scholar] [CrossRef]
- Piquette-Miller, M.; Pak, A.; Kim, H.; Anari, R.; Shahzamani, A. Decreased Expression and Activity of P-GIycoprotein in Rat Liver During Acute Inflammation. Pharm. Res. 1998, 15, 706–711. [Google Scholar] [CrossRef]
- Handschin, C.; Meyer, U.A. A Conserved Nuclear Receptor Consensus Sequence (DR-4) Mediates Transcriptional Activation of the Chicken CYP2H1 Gene by Phenobarbital in a Hepatoma Cell Line. J. Biol. Chem. 2000, 275, 13362–13369. [Google Scholar] [CrossRef] [Green Version]
- Prüfer, K.; Barsony, J. Retinoid X receptor dominates the nuclear import and export of the unliganded vitamin D receptor. Mol. Endocrinol. 2002, 16, 1738–1751. [Google Scholar] [CrossRef]
- Walther, R.F.; Lamprecht, C.; Ridsdale, A.; Groulx, I.; Lee, S.; Lefebvre, Y.A.; Haché, R.J. Nuclear export of the glucocorticoid receptor is accelerated by cell fusion-dependent release of calreticulin. J. Biol. Chem. 2003, 278, 37858–37864. [Google Scholar] [CrossRef] [PubMed]
- Shitan, N.; Tanaka, M.; Terai, K.; Ueda, K.; Yazaki, K. Human MDR1 and MRP1 recognize berberine as their transport substrate. Biosci. Biotechnol. Biochem. 2007, 71, 242–245. [Google Scholar] [CrossRef]
- Najar, I.A.; Sachin, B.S.; Sharma, S.C.; Satti, N.K.; Suri, K.A.; Johri, R.K. Modulation of P-glycoprotein ATPase activity by some phytoconstituents. Phytother. Res. 2010, 24, 454–458. [Google Scholar] [CrossRef]
- Mitsunaga, Y.; Takanaga, H.; Matsuo, H.; Naito, M.; Tsuruo, T.; Ohtani, H.; Sawadaa, Y. Effect of bioflavonoids on vincristine transport across blood–brain barrier. Eur. J. Pharmacol. 2000, 395, 193. [Google Scholar] [CrossRef]
- Hamabe, W.; Maeda, T.; Fukazawa, Y.; Kumamoto, K.; Shang, L.Q.; Yamamoto, A.; Yamamoto, C.; Tokuyama, S.; Kishioka, S. P-glycoprotein ATPase activating effect of opioid analgesics and their P-glycoprotein-dependent antinociception in mice. Pharmacol. Biochem. Behav. 2006, 85, 629–636. [Google Scholar] [CrossRef]
- Sun, Y.; Guo, T.; Guo, D.; Guo, L.; Chen, L.; Zhang, Y.; Wang, L. Establishment and characterization of an MDCK cell line stably-transfected with chicken Abcb1 encoding P-glycoprotein. Res. Vet. Sci. 2016, 106, 37–44. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound databases. Nucleic. Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef] [PubMed]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminformatics 2012, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Osguthorpe, D.J.; Sherman, W.; Hagler, A.T. Exploring protein flexibility: Incorporating structural ensembles from crystal structures and simulation into virtual screening protocols. J. Phys. Chem. B 2012, 116, 6952–6959. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Pedretti, A.; Villa, L.; Vistoli, G. VEGA – An open platform to develop chemo-bio-informatics applications, using plug-in architecture and script programming. J.Comput. Aided Mol. Des. 2004, 18, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K. Improvements to the APBS biomolecular solvation software suite. Protein Sci. 2018, 27, 112–128. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | Berberine (24 h) | |
|---|---|---|---|
| 40 mg/kg | 80 mg/kg | ||
| Ka (1/min) | 1.12 ± 0.28 | 1.87 ± 0.32 * | 2.25 ± 0.29 * |
| Papp (×10−4 cm/s) | 1.23 ± 0.42 | 2.2 ± 1.41 | 1.26 ± 0.42 |
| Parameter | Control | Berberine (2 h) | Control | Berberine (8 h) | ||||
|---|---|---|---|---|---|---|---|---|
| 5 μM | 20 μM | 40 μM | 5 μM | 20 μM | 40 μM | |||
| PappAP→BL(×10−6 cm/s) | 8.86 ± 1.38 | 5.43 ± 0.41 | 6.90 ± 0.76 | 9.94 ± 1.47 | 8.62 ± 1.24 | 6.43 ± 1.01 | 8.31 ± 0.8 | 7.35 ± 0.36 |
| PappBL→AP(×10−6 cm/s) | 62.9 ± 7.69 | 23.26 ± 1.77 | 23.26 ± 1.77 * | 18.49 ± 5.88 ** | 60.1 ± 5.13 | 27.11 ± 2.27 | 22.18 ± 2.43 * | 17.35 ± 0.82 ** |
| Efflux ratio | 7.28 ± 0.98 | 4.26 ± 0.85 | 3.4 ± 0.66 * | 1.92 ± 0.79 ** | 7.24 ± 0.49 | 4.37 ± 0.93 | 2.71 ± 0.51 * | 2.3 ± 0.16 ** |
| Inhibition rate | 0 | 41.48 | 53.30 | 73.63 | 0 | 39.64 | 62.57 | 68.23 |
| Verapamil (µM) | Papp AP→BL (×10−6 cm/s) | Papp BL→AP (×10−6 cm/s) | Efflux Ratio | Inhibition Rate |
|---|---|---|---|---|
| 0 | 8.04 ± 0.76 | 54.4 ± 4.34 | 6.24 ± 0.7 | 0 |
| 10 | 11.7 ± 0.53 | 41.1 ± 0.159 | 4.52 ± 0.11 | 27.56 |
| 50 | 7.4 ± 0.25 | 33.4 ± 4.02 | 4.16 ± 0.866 | 33.33 |
| 100 | 8.6 ± 1.5 | 27.7 ± 0.68 * | 3.51 ± 0.23 * | 43.75 |
| 1 000 | 9.1 ± 0.25 | 22.8 ± 1.56 * | 1.94 ± 0.098 * | 68.91 |
| 5 000 | 8.04 ± 0.76 | 10.7 ± 0.65 ** | 1.37 ± 0.21 ** | 78.04 |
| Name | CXR | CP-gp | |||||
|---|---|---|---|---|---|---|---|
| Docking Score (kcal/mol) & | Rescore * | Docking Score (kcal/mol) & | Rescore * | ||||
| Xscore | DSX | Xscore | DSX | ||||
| berberine | −7.8 | −9.2 | −160.8 | −9.5 | −9.41 | −196.9 | |
| morphine | −8.1 | −9.5 | |||||
| rhodamine123 | −9.8 | −9.4 | |||||
| indinavir | −9.1 | −10.3 | |||||
| ritonavir | −8.8 | −10.1 | |||||
| saquinavir | −10.8 | −10.8 | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Guo, L.; Huang, J.; Sun, Y.; He, F.; Zloh, M.; Wang, L. Inhibitory Effect of Berberine on Broiler P-glycoprotein Expression and Function: In Situ and In Vitro Studies. Int. J. Mol. Sci. 2019, 20, 1966. https://doi.org/10.3390/ijms20081966
Zhang Y, Guo L, Huang J, Sun Y, He F, Zloh M, Wang L. Inhibitory Effect of Berberine on Broiler P-glycoprotein Expression and Function: In Situ and In Vitro Studies. International Journal of Molecular Sciences. 2019; 20(8):1966. https://doi.org/10.3390/ijms20081966
Chicago/Turabian StyleZhang, Yujuan, Li Guo, Jinhu Huang, Yong Sun, Fang He, Mire Zloh, and Liping Wang. 2019. "Inhibitory Effect of Berberine on Broiler P-glycoprotein Expression and Function: In Situ and In Vitro Studies" International Journal of Molecular Sciences 20, no. 8: 1966. https://doi.org/10.3390/ijms20081966
APA StyleZhang, Y., Guo, L., Huang, J., Sun, Y., He, F., Zloh, M., & Wang, L. (2019). Inhibitory Effect of Berberine on Broiler P-glycoprotein Expression and Function: In Situ and In Vitro Studies. International Journal of Molecular Sciences, 20(8), 1966. https://doi.org/10.3390/ijms20081966