Proteomics Reveal the Profiles of Color Change in Brunfelsia acuminata Flowers



Abstract
:1. Introduction
2. Results
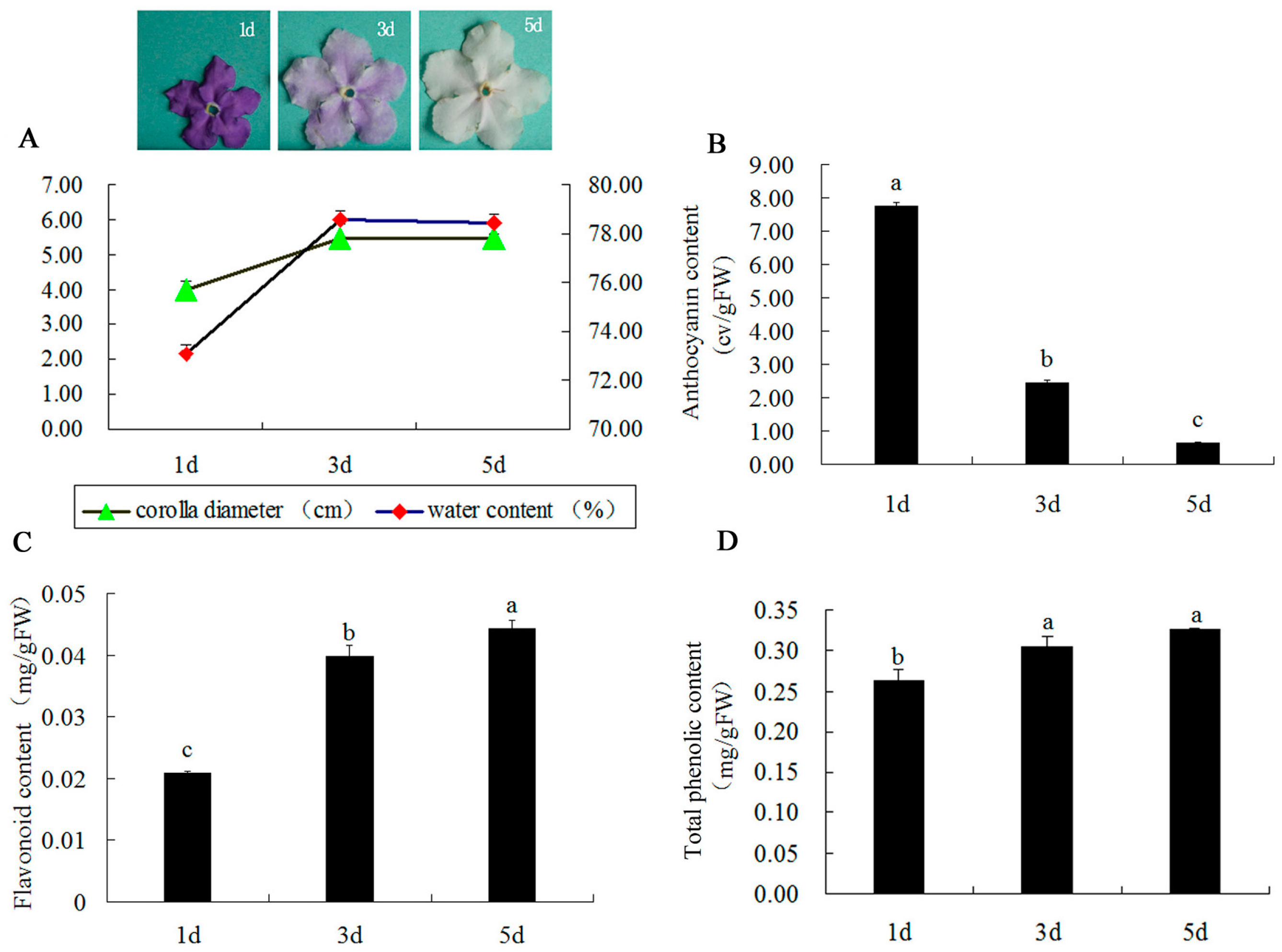
2.1. Changes in Corolla Diameter, Content of Water, Anthocyanin, Flavonoid, Total Phenolic, and Ultra-Structure during Flower Development
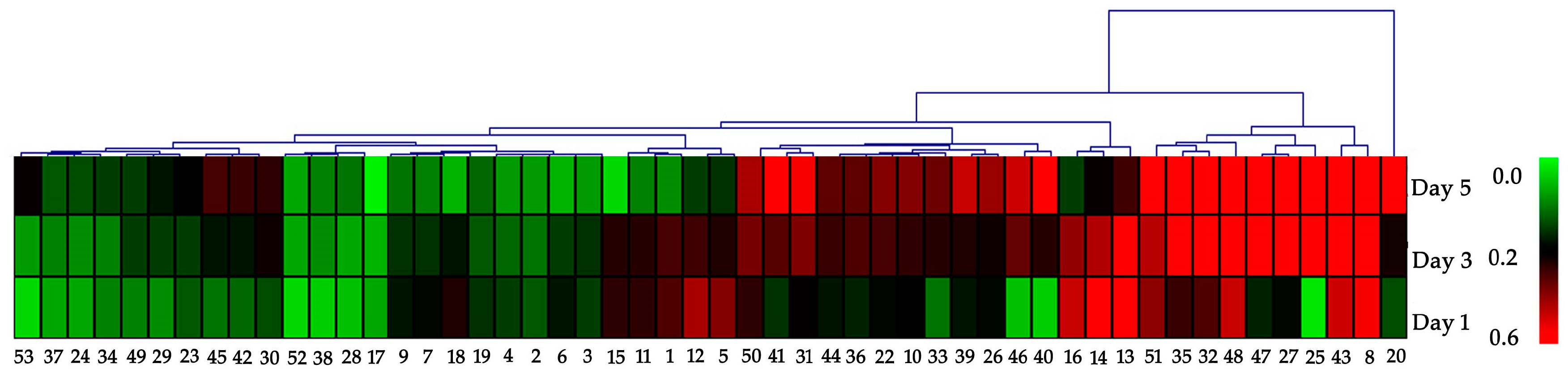
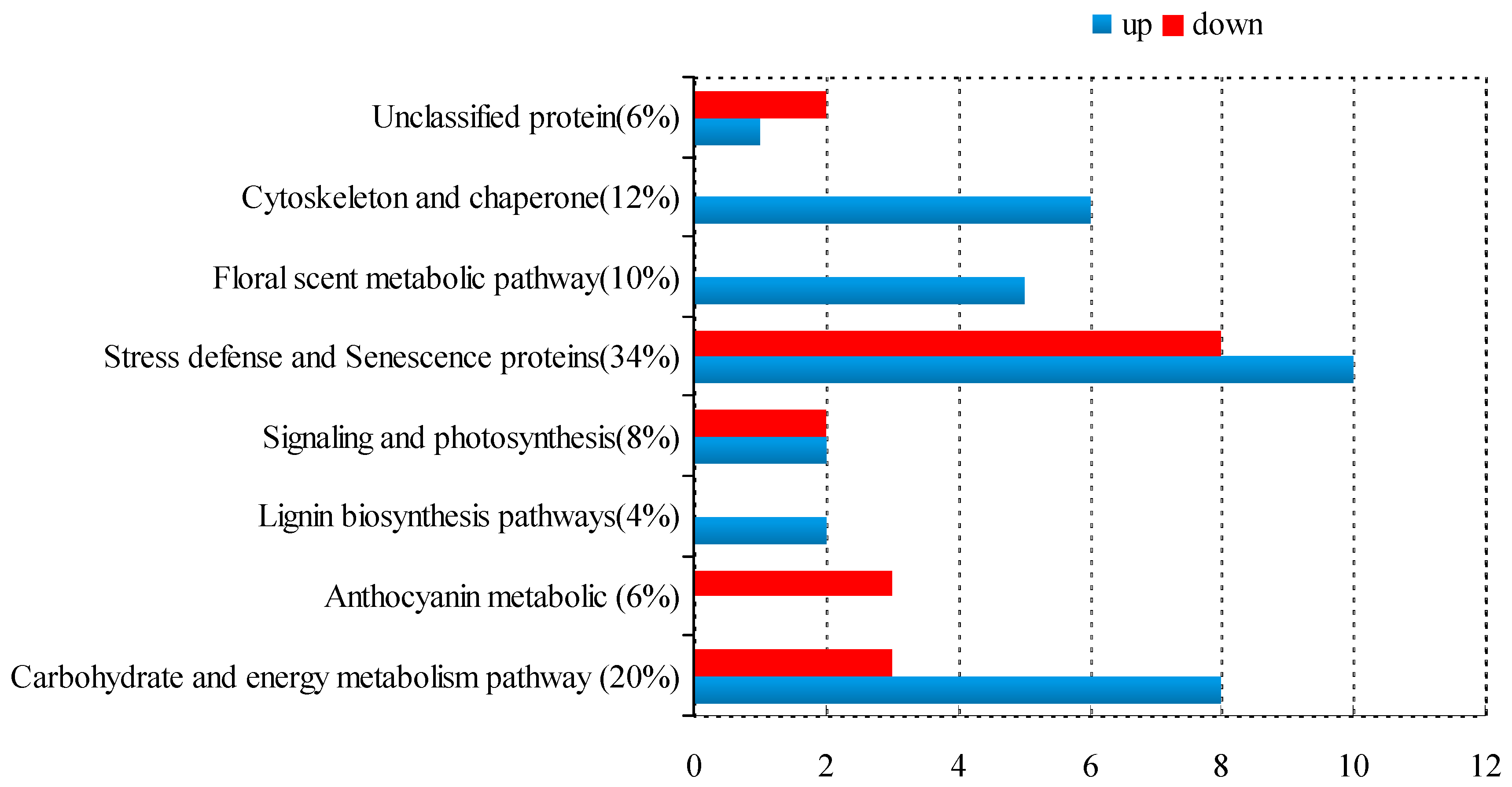
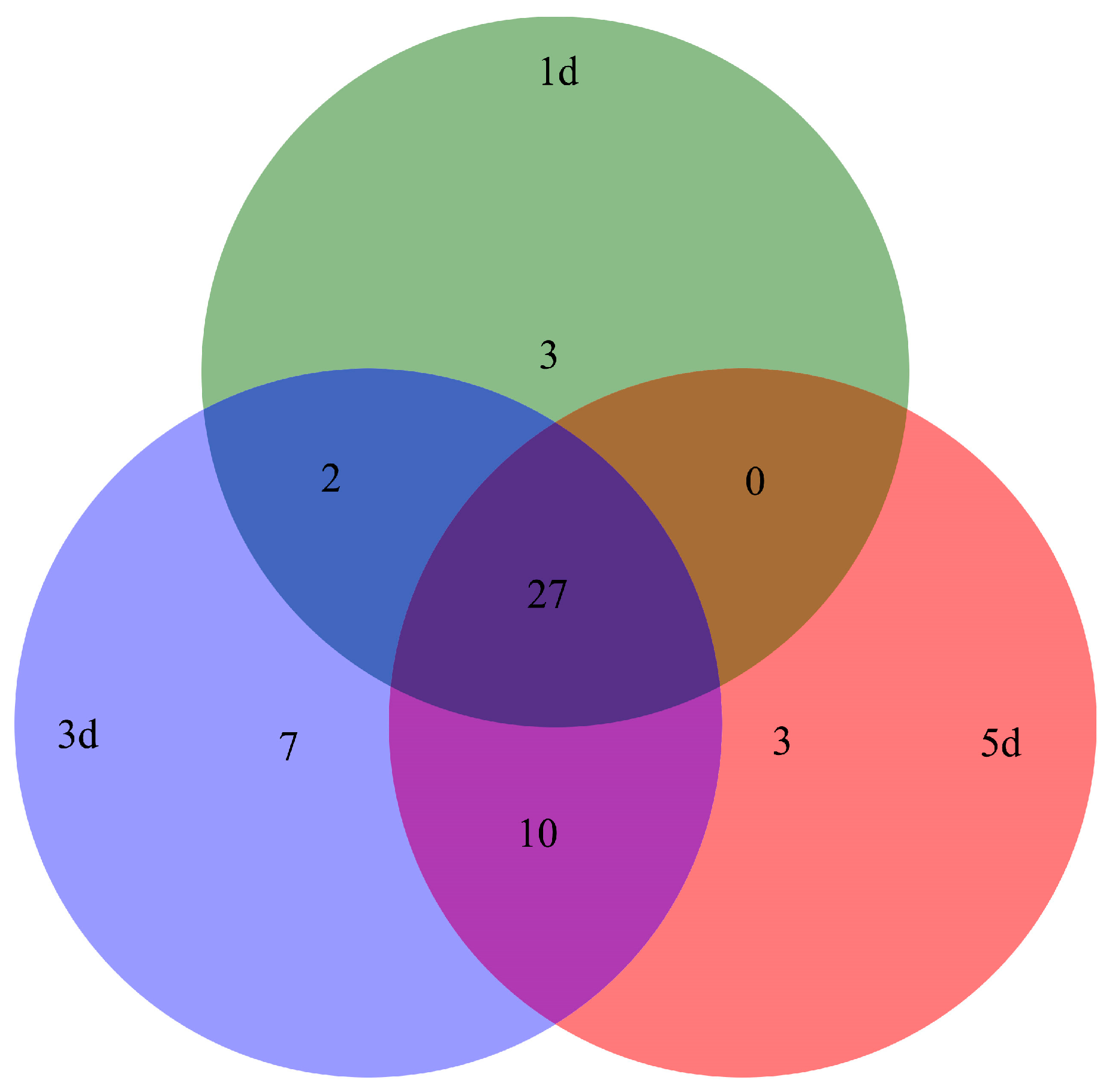
2.2. Protein Identification and Functional Classification
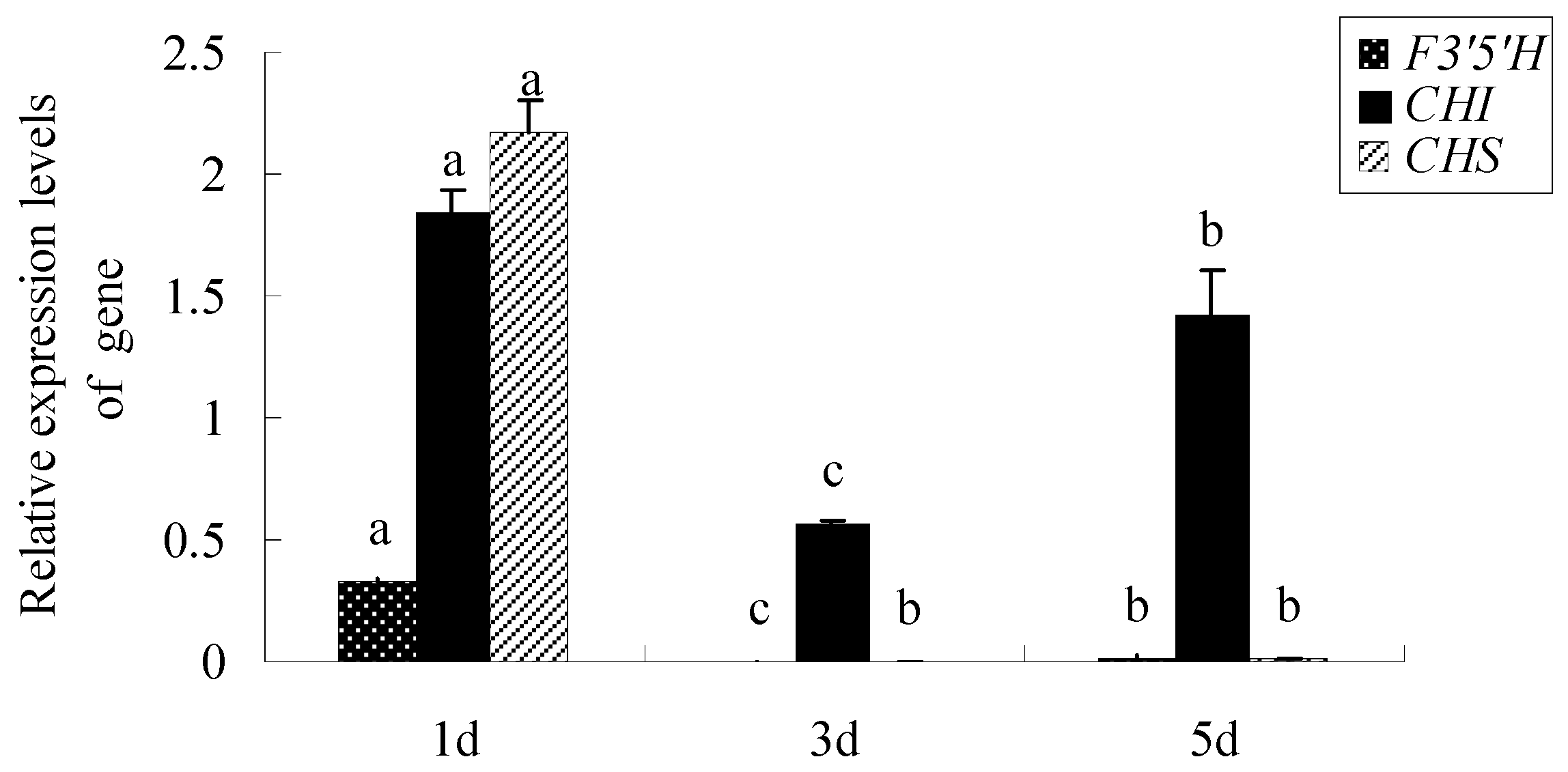
2.3. The Expression Levels of Key Genes Encoding Anthocyanin Synthesis in Different Days
2.4. Analysis of Volatiles
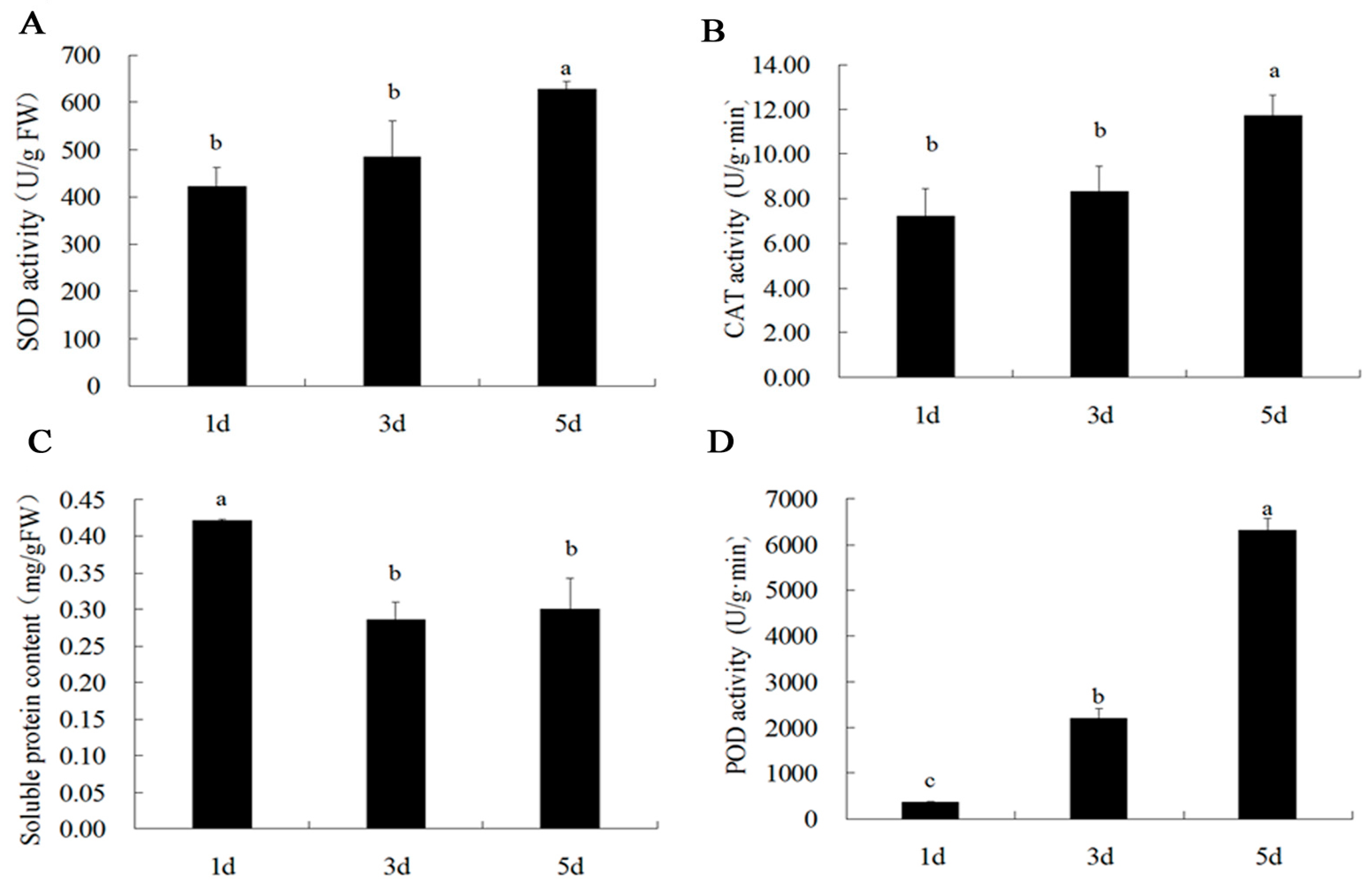
2.5. SOD, CAT, POD Activity and Soluble Protein Content in Petals of B. acuminata
3. Discussion
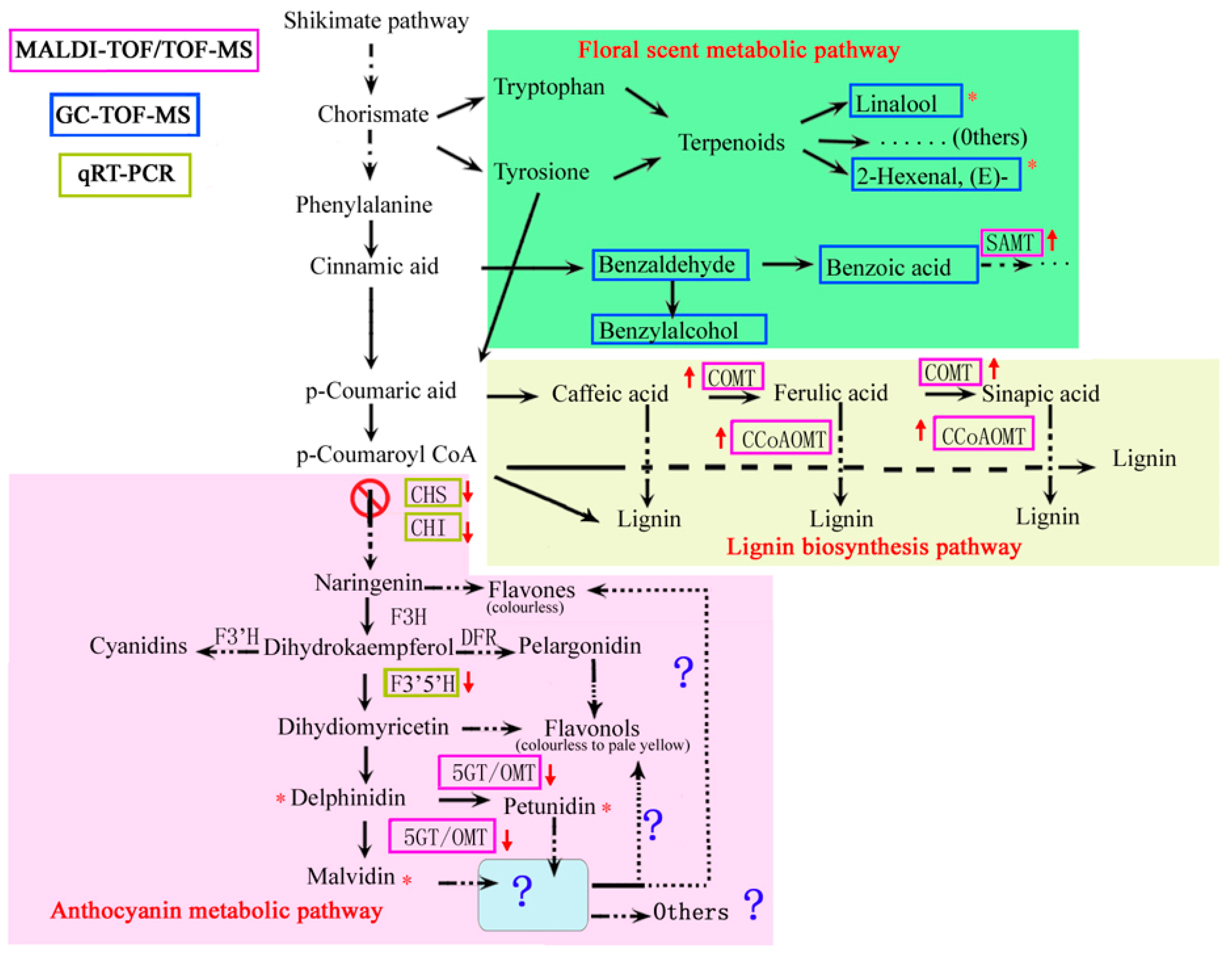
3.1. Main Proteins and Genes Related to Anthocyanin Synthesis
3.2. Other Protein Associated with Anthocyanin Synthesis
3.3. The Proteins Associated with Other Metabolic Pathways
4. Materials and Methods
4.1. Plant Materials and Morphological Indicators
4.2. Observation of Petal Epidermal Cells Structure and Ultra-Structure
4.3. Preparation of Total Protein Extraction
4.4. 2-DE and MALDI-TOF/TOF Analysis
4.5. GC-TOF-MS Analysis
4.6. Quantitative Real-Time (qRT-PCR) Analysis
4.7. Analysis of Physiological Parameters
4.8. Bioinformatic Analysis of Identified Proteins
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vaknin, H.; Barakiva, A.; Ovadia, R.; Nissimlevi, A.; Forer, I.; Weiss, D.; Orenshamir, M. Active anthocyanin degradation in Brunfelsia calycina (yesterday–today–tomorrow) flowers. Planta 2005, 222, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Michal, O.S. Does anthocyanin degradation play a significant role in determining pigment concentration in plants. Plant Sci. 2009, 177, 310–316. [Google Scholar]
- Markham, K.R.; Ryan, K.G.; Gould, K.S.; Rickards, G.K. Cell wall sited flavonoids in lisianthus flower petals. Phytochemistry 2000, 54, 681–687. [Google Scholar] [CrossRef]
- Kay, Q.O.N.; Daoud, H.S.; Stirton, C.H. Pigment distribution, light reflection and cell structure in petals. Bot. J. Linn. Soc. 2008, 83, 57–83. [Google Scholar] [CrossRef]
- Li, M.; Luo, Y.T.; Lu, X.C.; Sun, Y.T.; Qiu, D.L. Changes in Composition of Anthocyanins in Brunfelsia acuminata Flowers. J. Trop. Subtrop. Bot. 2018, 26, 627–632. [Google Scholar]
- Miller, R.; Owens, S.J.; Rørslett, B. Plants and colour: Flowers and pollination. Opt. Laser Technol. 2011, 43, 282–294. [Google Scholar] [CrossRef]
- Akifumi, A.; Hiroshi, Y.; Sato, A. Postharvest light irradiation and appropriate temperature treatment increase anthocyanin accumulation in grape berry skin. Postharvest Biol. Technol. 2019, 147, 89–99. [Google Scholar]
- Welch, C.R.; Wu, Q.; Simon, J.E. Recent Advances in Anthocyanin Analysis and Characterization. Curr. Anal. Chem. 2008, 4, 75. [Google Scholar] [CrossRef]
- Cheynier, V. Phenolic compounds: From plants to foods. Phytochem. Rev. 2012, 11, 153–177. [Google Scholar] [CrossRef]
- Wangia, C.; Orwa, J.; Muregi, F.; Kareru, P.; Cheruiyot, K.; Kibet, J. Comparative Anti-oxidant Activity of Aqueous and Organic Extracts from Kenyan Ruellia linearibracteolata and Ruellia bignoniiflora. Eur. J. Med. Plants 2016, 17, 1–7. [Google Scholar] [CrossRef]
- Liu, F.; Wang, M.; Wang, M. Phenolic compounds and antioxidant activities of flowers, leaves and fruits of five crabapple cultivars (Malus Mill. species). Sci. Hortic. 2018, 235, 460–467. [Google Scholar] [CrossRef]
- Nascimento, L.B.D.S.; Leal-Costa, M.V.; Menezes, E.A.; Lopes, V.R.; Muzitano, M.F.; Costa, S.S.; Tavares, E.S. Ultraviolet-B radiation effects on phenolic profile and flavonoid content of Kalanchoe pinnata. J. Photochem. Photobiol. B 2015, 148, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Vera, H.; Angela, D.; Bernhard, E.; Richard, L.; Stefan, M.; Michael, A. Characterization and structural features of a chalcone synthase mutation in a white-flowering line of Matthiola incana R. Br. (Brassicaceae). Plant Mol. Biol. 2004, 55, 455–465. [Google Scholar]
- Zhang, X.; Liang, Z. Morphology, structure and ultrastructure of staminal nectary in Lamprocapnos (Fumarioideae, Papaveraceae). Flora 2018, 242, 128–136. [Google Scholar] [CrossRef]
- Avalos, A.A.; Lattar, E.C.; Galati, B.G.; Ferrucci, M.S. Nectary structure and ultrastructure in two floral morphs of Koelreuteria elegans subsp. formosana (Sapindaceae). Flora 2017, 226, 29–37. [Google Scholar] [CrossRef]
- Cao, Y.T.; Qiu, D.L. Cloning and Sequence Analysis of Chalcone Isomerase Gene (CHI) from Brunfelsia acuminata Flowers. Master’s Thesis, Fujian Agriculture and Forestry University, Fujian, China, April 2012. [Google Scholar]
- Naing, A.H.; Ai, T.N.; Su, M.J.; Park, K.I.; Lim, K.B.; Chang, K.K. Expression of RsMYB1 in chrysanthemum regulates key anthocyanin biosynthetic genes. Electron. J. Biotechnol. 2015, 18, 359–364. [Google Scholar] [CrossRef]
- Ai, T.N.; Naing, A.H.; Arun, M.; Su, M.J.; Chang, K.K. Expression of RsMYB1 in Petunia enhances anthocyanin production in vegetative and floral tissues. Sci. Hortic. 2017, 214, 58–65. [Google Scholar] [CrossRef]
- Yamagishi, M.; Uchiyama, H.; Handa, T. Floral pigmentation pattern in Oriental hybrid lily (Lilium spp.) cultivar ‘Dizzy’ is caused by transcriptional regulation of anthocyanin biosynthesis genes. J. Plant Physiol. 2018, 228, 85. [Google Scholar] [CrossRef] [PubMed]
- Zipor, G.; Duarte, P.; Carqueijeiro, I.; Shahar, L.; Ovadia, R.; Teper-Bamnolker, P.; Eshel, D.; Levin, Y.; Doron-Faigenboim, A.; Sottomayor, M. In planta anthocyanin degradation by a vacuolar class III peroxidase in Brunfelsia calycina flowers. New Phytol. 2015, 205, 653–665. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Zhou, X.; Dong, L.; Guo, J.; Chen, Y.; Zhang, Y.; Wu, L.; Xu, M. iTRAQ-based analysis of the Arabidopsis proteome reveals insights into the potential mechanisms of anthocyanin accumulation regulation in response to phosphate deficiency. J. Proteomics 2018, 184, 39–53. [Google Scholar] [CrossRef]
- Li, X.; Jackson, A.; Xie, M.; Wu, D.; Tsai, W.C.; Zhang, S. Proteomic insights into floral biology. BBA Proteins Proteomics 2016, 1864, 1050–1060. [Google Scholar] [CrossRef]
- Farzad, M.; Griesbach, R.; Hammond, J.; Weiss, M.R.; Elmendorf, H.G. Differential expression of three key anthocyanin biosynthetic genes in a color-changing flower, Viola cornuta cv. Yesterday, Today and Tomorrow. Plant Sci. 2003, 165, 1333–1342. [Google Scholar] [CrossRef]
- Yamazaki, M.; Gong, Z.; Fukuchimizutani, M.; Fukui, Y.; Tanaka, Y.; Kusumi, T.; Saito, K. Molecular Cloning and Biochemical Characterization of a Novel Anthocyanin 5-O-Glucosyltransferase by mRNA Differential Display for Plant Forms Regarding Anthocyanin. J. Biol. Chem. 1999, 274, 7405. [Google Scholar] [CrossRef] [PubMed]
- Akita, Y.; Hase, Y.; Narumi, I.; Ishizaka, H.; Kondo, E.; Kameari, N.; Nakayama, M.; Tanikawa, N.; Morita, Y.; Tanaka, A. Isolation and characterization of the fragrant cyclamen -methyltransferase involved in flower coloration. Planta 2011, 234, 1127–1136. [Google Scholar] [CrossRef]
- Wei, K.; Wang, L.; Zhang, C.; Wu, L.; Li, H.; Zhang, F.; Cheng, H. Transcriptome Analysis Reveals Key Flavonoid 3′-Hydroxylase and Flavonoid 3′,5′-Hydroxylase Genes in Affecting the Ratio of Dihydroxylated to Trihydroxylated Catechins in Camellia sinensis. PLoS ONE 2015, 10, e0137925. [Google Scholar] [CrossRef]
- Seitz, C.; Ameres, S.; Schlangen, K.; Forkmann, G.; Halbwirth, H. Multiple evolution of flavonoid 3′,5′-hydroxylase. Planta 2015, 242, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Cao, Y.T.; Ye, S.R.; Irshad, M.; Pan, T.F.; Qiu, D.L. Isolation of CHS Gene from Brunfelsia acuminata Flowers and Its Regulation in Anthocyanin Biosysthesis. Molecules 2016, 22, 44. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Q.; Zhu, M.Y.; Jiang, Y.; Zhao, D.Q.; Tao, J. Molecular characterization of chalcone isomerase (CHI) regulating flower color in herbaceouspeony (PaeonialactifloraPall.). J. Integr. Agric. 2018, 17, 122–129. [Google Scholar] [CrossRef]
- Muir, S.R.; Collins, G.J.; Robinson, S.; Hughes, S.; Bovy, A.; de Vos, C.H.R.; van Tunen, A.J.; Martine, E.; Verhoeyen, M.E. Over expression of petunia chalcone isomerase in tomato results in fruit containing increased levels of flavonols. Nat. Biotechnol. 2001, 19, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Barakiva, A.; Ovadia, R.; Rogachev, I.; Baror, C.; Bar, E.; Freiman, Z.; Nissimlevi, A.; Gollop, N.; Lewinsohn, E.; Aharoni, A. Metabolic networking in Brunfelsia calycina petals after flower opening. J. Exp. Bot. 2010, 61, 1393. [Google Scholar] [CrossRef]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin Biosynthesis and Degradation Mechanisms in SolanaceousVegetables: A Review. Front. Chem. 2018, 6, 1–17. [Google Scholar] [CrossRef]
- Zhang, Q.; Tian, Y.Y.; Meng, Y.E.; Li, Y.M.; Wang, H.J.; Wang, L.M.; Zhao, X.S. Research advance in genetic engineering of floral fragrance. J. Henan Agric. Sci. 2014, 43, 11–16. [Google Scholar]
- Gimenez, M.J.; Valverde, J.; Valero, D.; Diazmula, H.M.; Zapata, P.J.; Serrano, M.; Moral, J.M.; Castillo, S. Methyl salicylate treatments of sweet cherry trees improve fruit quality at harvest and during storage. Sci. Hortic. 2015, 197, 665–673. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.X.; Shi, Z.P. Research on the Cell Microstructure of Jasminum sambac’s Petal. J. Hunan Agric. Univ. 1999, 25, 108–111. [Google Scholar]
- Giordano, D.; Provenzano, S.; Ferrandino, A.; Vitali, M.; Pagliarani, C.; Roman, F.; Cardinale, F.; Castellarin, S.D.; Schubert, A. Characterization of a multifunctional caffeoyl-CoA O-methyltransferase activated in grape berries upon drought stress. Plant Physiol. Biochem. 2016, 101, 23. [Google Scholar] [CrossRef]
- Cui, N.; Yu, Z.H.; Han, M.L.; Dong, X.F.; Qu, B.; Li, T.L. Research Advancement of 14-3-3 Proteins in Plants. Acta Bot. Boreali Occident. Sin. 2012, 32, 843–851. [Google Scholar]
- Su, M.; Lv, J.H.; Zhang, Q.X. Research Advance in Floral Fragrance Genetic Engineering. J. Anhui Agric. Sci. 2007, 35, 3169–3171. [Google Scholar]
- Martens, S.; Mithöfer, A. Flavones and flavone synthases. Phytochemistry 2005, 66, 2399–2407. [Google Scholar] [CrossRef]
- Wang, L.; Zheng, Y.; Ding, S.; Zhang, Q.; Chen, Y.; Zhang, J. Molecular cloning, structure, phylogeny and expression analysis of the invertase gene family in sugarcane. BMC Plant Biol. 2017, 17, 109. [Google Scholar] [CrossRef]
- Wang, L.J. Advances on the studies of invertase on sucrose metabolism in higher plant. J. Agric. Sci. 2010, 31, 70–75. [Google Scholar]
- Walker, R.P.; Paoletti, A.; Leegood, R.C.; Famiani, F. Phosphorylation of phosphoenolpyruvate carboxykinase (PEPCK) and phosphoenolpyruvate carboxylase (PEPC) in the flesh of fruits. Plant Physiol. Biochem. 2016, 108, 323–327. [Google Scholar] [CrossRef]
- Chen, Z.; Xia, L.; He, Y.; Zhang, J.; Yan, T.; Liu, X. Physiological investigation of C4 -phosphoenolpyruvate-carboxylase-introduced rice line shows that sucrose metabolism is involved in the improved drought tolerance. Plant Physiol. Biochem. 2017, 115, 328–342. [Google Scholar]
- Silva, L.S.; Seabra, A.R.; Leitão, J.N.; Carvalho, H.G. Possible role of glutamine synthetase of the prokaryotic type (GSI-like) in nitrogen signaling in Medicago truncatula. Plant Sci. 2015, 240, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Hu, W.; Feng, J.; Yang, X.; Huang, Q.; Xiao, J.; Liu, Y.; Yang, G.; He, G. Identification of the ASR gene family from Brachypodium distachyon and functional characterization of BdASR1 in response to drought stress. Plant Cell Rep. 2016, 35, 1221–1234. [Google Scholar] [CrossRef]
- Langenkämper, G.; Manac’H, N.; Broin, M.; Cuiné, S.; Becuwe, N.; Kuntz, M.; Rey, P. Accumulation of plastid lipid-associated proteins (fibrillin/CDSP34) upon oxidative stress, ageing and biotic stress in Solanaceae and in response to drought in other species. J. Exp. Bot. 2001, 52, 1545. [Google Scholar] [CrossRef]
- Pan, Z.Y.; Liu, Q.; Yun, Z.; Rui, G.; Zeng, W.F.; Qiang, X.; Deng, X.X. Comparative proteomics of a lycopene-accumulating mutant reveals the important role of oxidative stress on carotenogenesis in sweet orange (Citrus sinensis [L.] Osbeck). Proteomics 2009, 9, 5455–5470. [Google Scholar] [CrossRef]
- Ihsan, U.; Bassam, O.A.J.; Khalid, M.S.A.G.; Hind, A.A.A.Z.; Yasir, A.; Ahmad, F.; Naser, A.K.; Mohammed, A.A. Endophytic bacteria isolated from Solanum nigrum L., alleviate cadmium (Cd) stress response by their antioxidant potentials, including SOD synthesis by sodA gene. Ecotoxicol. Environ. Saf. 2019, 174, 197–207. [Google Scholar]
- Faten, H.F.; Riadh, K.; Chedly, A. Total phenolic, flavonoid and tannin contents and antioxidant andantimicrobial activities of organic extracts of shoots of the plant Limonium delicatulum. J. Taibah Univ. Sci. 2014, 8, 216–224. [Google Scholar]
- Quintá, H.R.; Galigniana, N.M.; Erlejman, A.G.; Lagadari, M.; Piwien-Pilipuk, G.; Galigniana, M.D. Management of cytoskeleton architecture by molecular chaperones and immunophilins. Cell. Signal. 2011, 23, 1907–1920. [Google Scholar] [CrossRef] [Green Version]
- Orenshamir, M.O.M.; Dela, G.D.G.; Ovadia, R.O.R.; Nissimlevi, A.N.A.; Philosophhadas, S.P.S.; Meir, S.M.S. Differentiation between petal blueing and senescence of cut ‘Mercedes’ rose flowers. J. Hortic. Sci. Biotechnol. 2015, 76, 195–200. [Google Scholar]
- Meng, J.F.; Xu, T.F.; Wang, Z.Z.; Fang, Y.L.; Xi, Z.M.; Zhang, Z.W. The ameliorative effects of exogenous melatonin on grape cuttings under water-deficient stress: Antioxidant metabolites, leaf anatomy, and chloroplast morphology. J. Pineal Res. 2015, 57, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Pan, D.; Lv, X.; Song, X.; Qiu, Z.; Huang, C.; Huang, R.; Wei, C. Proteomic approach reveals that starch degradation contributes to anthocyanin accumulation in tuberous root of purple sweet potato. J. Proteomics 2016, 143, 298–305. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, W.; Wang, Y.; Gao, S.; Du, D.; Fu, J.; Dong, L. Anthocyanin biosynthesis and accumulation in developing flowers of tree peony (Paeonia suffruticosa) ‘Luoyang Hong’. Postharvest Biol. Technol. 2014, 97, 11–22. [Google Scholar] [CrossRef]
- Sangeeta, M.; Muhammad, I.; Biswojit, D.; Lu, X.C.; Li, M.; Chandra, K.D.; Hafiz, M.R.; Qiu, Z.P.; Qiu, D.L. Effect of vineyard soil variability on chlorophyll fluorescence, yield and quality of table grape as influenced by soil moisture, grown under double cropping system in protected condition. Peer J. 2018, 9, 9–25. [Google Scholar]
- Wang, W.X. Principles and Techniques of Plant Physiological Biochemical Experiments; Higher Education Press: Beijing, China, 2006; pp. 282–286. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No a | Protein Name | Species | Accession No. | MW (kDa)/pI b | Score | Cov c | Fold Changes e | |
|---|---|---|---|---|---|---|---|---|
| Carbohydrate and Energy Metabolism Pathway | ||||||||
| D5 d | malate dehydrogenase, cytoplasmic-like | Solanum lycopersicum | gi|460404529 | 35.361/5.91 | 235 | 25% | −2.33 | −1.55 |
| D10 | 2,3-bisphosphoglycerate-independent phosphoglycerate mutase-like | Glycine max | gi|356568270 | 60.799/5.58 | 247 | 11% | −1.92 | −1.32 |
| D19 | 3-isopropylmalate dehydrogenase, chloroplastic-like | Solanum lycopersicum | gi|460386440 | 43.396/6.05 | 232 | 14% | −2.06 | −1.25 |
| U23 d | ATP synthase beta subunit | Eleutherococcus senticosus | gi|343410685 | 39.790/5.77 | 227 | 35% | 1.20 | 1.18 |
| U24 | soluble acid invertase 2 | Orobanche ramosa | gi|294612072 | 61.628/5.24 | 133 | 19% | 2.07 | 1.51 |
| U29 | vacuolar invertase 2 | Gossypium hirsutum | gi|268526570 | 69.303/5.14 | 185 | 8% | 2.16 | 1.98 |
| U32 | ATP synthase subunit beta, mitochondrial-like | Solanum lycopersicum | gi|460382474 | 59.825/5.94 | 869 | 45% | 2.26 | 1.46 |
| U45 | ATP synthase subunit beta, mitochondrial-like | Solanum lycopersicum | gi|460382474 | 59.825/5.94 | 311 | 26% | 3.00 | 2.71 |
| U46 | phosphoenolpyruvate carboxylase kinase 1 | Clusia minor | gi|39842451 | 28.716/6.38 | 88 | 22% | 13.09 | 10.67 |
| U49 | ATP synthase beta subunit | Eleutherococcus senticosus | gi|343410685 | 39.790/5.77 | 227 | 29% | 2.40 | 2.26 |
| U52 | 1,2-beta-fructan 1F-fructosyltransferase | Helianthus tuberosus | gi|3367690 | 69.214/5.02 | 129 | 12% | 2.11 | 2.00 |
| Anthocyanin Metabolic Pathway | ||||||||
| D14 | Anthocyanin-O-methyl transferase | Solanum tuberosum | gi|441433515 | 26.282/5.69 | 90 | 19% | −7.04 | −1.04 |
| D17 | Anthocyanin-5-O-glucosyltransferase | Petunia x hybrida | gi|6683052 | 52.130/5.07 | 98 | 13% | −13.79 | −1.04 |
| D18 | Anthocyanin-5-O-glucosyltransferase | Petunia x hybrida | gi|6683052 | 52.130/5.07 | 100 | 7% | −3.94 | −1.36 |
| Lignin Biosynthesis Pathway | ||||||||
| U41 | caffeate-O-methyltransferase | Liquidambar styraciflua | gi|5732000 | 39.944/5.69 | 80 | 22% | 4.70 | 3.77 |
| U47 | caffeoyl-CoA O-methyltransferase | Broussonetia papyrifera | gi|46394464 | 27.701/5.31 | 412 | 40% | 5.25 | 3.44 |
| Stress Defense and Senescence Proteins | ||||||||
| D1 | polyphenol oxidase E, chloroplastic-like isoform 2 | Solanum lycopersicum | gi|460401035 | 66.181/6.36 | 83 | 18% | −2.66 | −1.02 |
| D2 | polyphenol oxidase | Nicotiana tabacum | gi|92919068 | 57.748/5.92 | 126 | 13% | −2.10 | −1.02 |
| D6 | proteasome subunit alpha type-6-like | Solanum lycopersicum | gi|460412613 | 27.301/6.11 | 102 | 32% | −4.08 | −1.37 |
| U8 | glutathione S-transferase | Solanum commersonii | gi|148616162 | 23.843/5.98 | 82 | 30% | 1.34 | 1.53 |
| D13 | lactoylglutathione lyase-like | Solanum lycopersicum | gi|460373807 | 32.839/5.95 | 283 | 32% | −2.00 | −1.07 |
| U25 | ASR1 protein | Solanum ochranthum | gi|321155417 | 12.547/6.48 | 307 | 34% | 31.33 | 23.55 |
| U28 | CLPC | Theobroma cacao | gi|508775360 | 102.257/6.36 | 373 | 27% | 3.45 | 1.12 |
| U35 | S-adenosyl methionine synthase-like | Solanum tuberosum | gi|78191442 | 43.189/5.52 | 466 | 40% | 2.11 | 2.01 |
| U43 | glutathione S-transferase L3-like | Cicer arietinum | gi|502121795 | 27.092/5.79 | 91 | 15% | 1.59 | 1.51 |
| U44 | putative glutathione S-transferase zeta-class 2 | Brassica napus | gi|330250478 | 25.336/5.53 | 82 | 24% | 2.14 | 1.49 |
| D3 | Adenylosuccinate synthetase, chloroplastic-like | Solanum lycopersicum | gi|460407669 | 55.408/7.55 | 87 | 14% | −2.39 | −1.04 |
| D4 | Adenylosuccinate synthetase1, chloroplastic-like | Solanum lycopersicum | gi|460407669 | 55.408/7.55 | 93 | 18% | −2.11 | −1.38 |
| D15 | Aspartic proteinase | Theobroma cacao | gi|508719874 | 54.428/5.56 | 55 | 3% | −7.85 | −1.08 |
| D16 | endochitinase precursor | Humulus lupulus | gi|4960049 | 33.508/7.42 | 109 | 4% | −3.09 | −1.10 |
| U20 | Small ubiquitin-related modifier 1 | Arabidopsis thaliana | gi|21542462 | 10.969/4.91 | 77 | 48% | 9.19 | 1.68 |
| U26 | annexin p34-like protein-like | Solanum tuberosum | gi|81074127 | 35.909/5.54 | 174 | 24% | 2.01 | 1.59 |
| U27 | Glutamine synthetase 1,4 | Theobroma cacao | gi|508707247 | 39.098/6.02 | 160 | 18% | 5.21 | 2.86 |
| U51 | heat shock 70 protein | Spinacia oleracea | gi|2773050 | 76.094/5.19 | 554 | 25% | 2.02 | 1.07 |
| U53 | Plastid-lipid-associated protein, chloroplast precursor, putative | Ricinus communis | gi|223536371 | 34.979/4.84 | 148 | 24% | 2.06 | 1.78 |
| Floral scent Metabolic Pathway | ||||||||
| U33 | 1-hydroxy-2-methyl-2-(E)-butenyl 4-diphosphate reductase | Ipomoea batatas | gi|325557690 | 51.682/5.90 | 163 | 27% | 2.58 | 2.20 |
| U34 | 1-deoxy-D-xylulose-5-phosphate reductoisomerase | Solanum lycopersicum | gi|350537527 | 51.465/5.94 | 266 | 27% | 2.01 | 1.49 |
| U39 | SAMT | Anthocercis littorea | gi|58201456 | 32.353/4.79 | 151 | 22% | 2.11 | 1.62 |
| U40 | Putative S-adenosyl-L-methionine:Salicylic acid carboxyl methyltransferase | Pisum sativum | gi|37725949 | 40.552/5.17 | 78 | 19% | 15.05 | 7.00 |
| U42 | putative S-adenosyl-L-methionine:Salicylic acid carboxyl methyltransferase | Pisum sativum | gi|37725949 | 40.552/5.17 | 52 | 5% | 2.23 | 1.56 |
| Signaling and Photosynthesis | ||||||||
| D11 | inositol-3-phosphate synthase | Solanum lycopersicum | gi|460388681 | 56.526/5.45 | 189 | 23% | −2.79 | −1.02 |
| D12 | inositol-3-phosphate synthase | Solanum lycopersicum | gi|460388681 | 56.526/5.45 | 318 | 23% | −2.84 | −1.51 |
| U22 | 14-3-3-like protein GF14 Psi | Eutrema salsugineum | gi|309952059 | 28.752/4.78 | 166 | 48% | 2.07 | 1.81 |
| U31 | ruBisCO large subunit-binding protein subunit beta, chloroplastic-like | Cicer arietinum | gi|502125499 | 62.800/5.85 | 348 | 22% | 2.15 | 1.26 |
| Cytoskeleton and Chaperone | ||||||||
| U30 | chaperonin CPN60-2, mitochondrial-like | Solanum lycopersicum | gi|460404682 | 61.521/5.51 | 175 | 18% | 2.37 | 1.99 |
| U36 | beta-actin | Zoysia japonica | gi|284157810 | 41.697/5.23 | 459 | 35% | 2.21 | 2.11 |
| U37 | actin 6 | Populus trichocarpa | gi|222860713 | 40.592/5.05 | 166 | 27% | 1.85 | 1.43 |
| U38 | actin | Gossypium hirsutum | gi|32186904 | 41.878/5.39 | 82 | 27% | 2.5 | 2.25 |
| U50 | 60-kDa chaperonin-60 alpha-polypeptide precursor, partial | Brassica napus | gi|289365 | 57.657/4.84 | 403 | 24% | 2.61 | 1.93 |
| Unclassified Protein | ||||||||
| D7 | predicted protein | Physcomitrella patens subsp. Patens | gi|162667966 | 28.780/5.38 | 79 | 11% | −2.13 | −1.11 |
| D9 | putative transcription factor BTF3-like | Solanum tuberosum | gi|82623431 | 17.472/6.31 | 240 | 34% | −2.06 | −1.12 |
| U48 | cp10-like proteinCP10 | Gossypium hirsutum | gi|21780187 | 26.761/7.77 | 122 | 9% | 1.30 | 1.73 |
| Number | Compounds | Relative Contents (%) | ||
|---|---|---|---|---|
| 1d | 3d | 5d | ||
| 1 | Linalool | 37.59 ± 8.39 | 31.27 ± 2.12 | 19.24 ± 4.12 |
| 2 | 2-Hexenal, (E)- | 44.92 ± 5.11 | 20.98 ± 1.29 | — |
| 3 | trans-Linalool oxide (furanoid) | 5.91 ± 1.47 | 11.61 ± 1.58 | 8.27 ± 1.56 |
| 4 | (E)-4,8-Dimethylnona-1,3,7-triene | 9.62 ± 5.28 | 9.81 ± 1.07 | 10.11 ± 6.31 |
| 5 | 2-Furanmethanol, 5-ethenyltetrahydro-à,à,5-trimethyl-, cis- | 3.47 ± 0.42 | 9.34 ± 1.60 | 6.07 ± 1.37 |
| 6 | 1-Hexanol | 5.22 ± 0.83 | 5.13 ± 1.25 | 12.53 ± 5.98 |
| 7 | Benzeneacetaldehyde | 0.67 ± 0.20 | 4.15 ± 1.70 | 11.40 ± 0.85 |
| 8 | 2-Hexenal | 0.74 ± 0.44 | 0.45 ± 0.13 | 16.78 ± 1.43 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Sun, Y.; Lu, X.; Debnath, B.; Mitra, S.; Qiu, D. Proteomics Reveal the Profiles of Color Change in Brunfelsia acuminata Flowers. Int. J. Mol. Sci. 2019, 20, 2000. https://doi.org/10.3390/ijms20082000
Li M, Sun Y, Lu X, Debnath B, Mitra S, Qiu D. Proteomics Reveal the Profiles of Color Change in Brunfelsia acuminata Flowers. International Journal of Molecular Sciences. 2019; 20(8):2000. https://doi.org/10.3390/ijms20082000
Chicago/Turabian StyleLi, Min, Yueting Sun, Xiaocao Lu, Biswojit Debnath, Sangeeta Mitra, and Dongliang Qiu. 2019. "Proteomics Reveal the Profiles of Color Change in Brunfelsia acuminata Flowers" International Journal of Molecular Sciences 20, no. 8: 2000. https://doi.org/10.3390/ijms20082000
APA StyleLi, M., Sun, Y., Lu, X., Debnath, B., Mitra, S., & Qiu, D. (2019). Proteomics Reveal the Profiles of Color Change in Brunfelsia acuminata Flowers. International Journal of Molecular Sciences, 20(8), 2000. https://doi.org/10.3390/ijms20082000