Theaflavin-Enriched Fraction Stimulates Adipogenesis in Human Subcutaneous Fat Cells




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
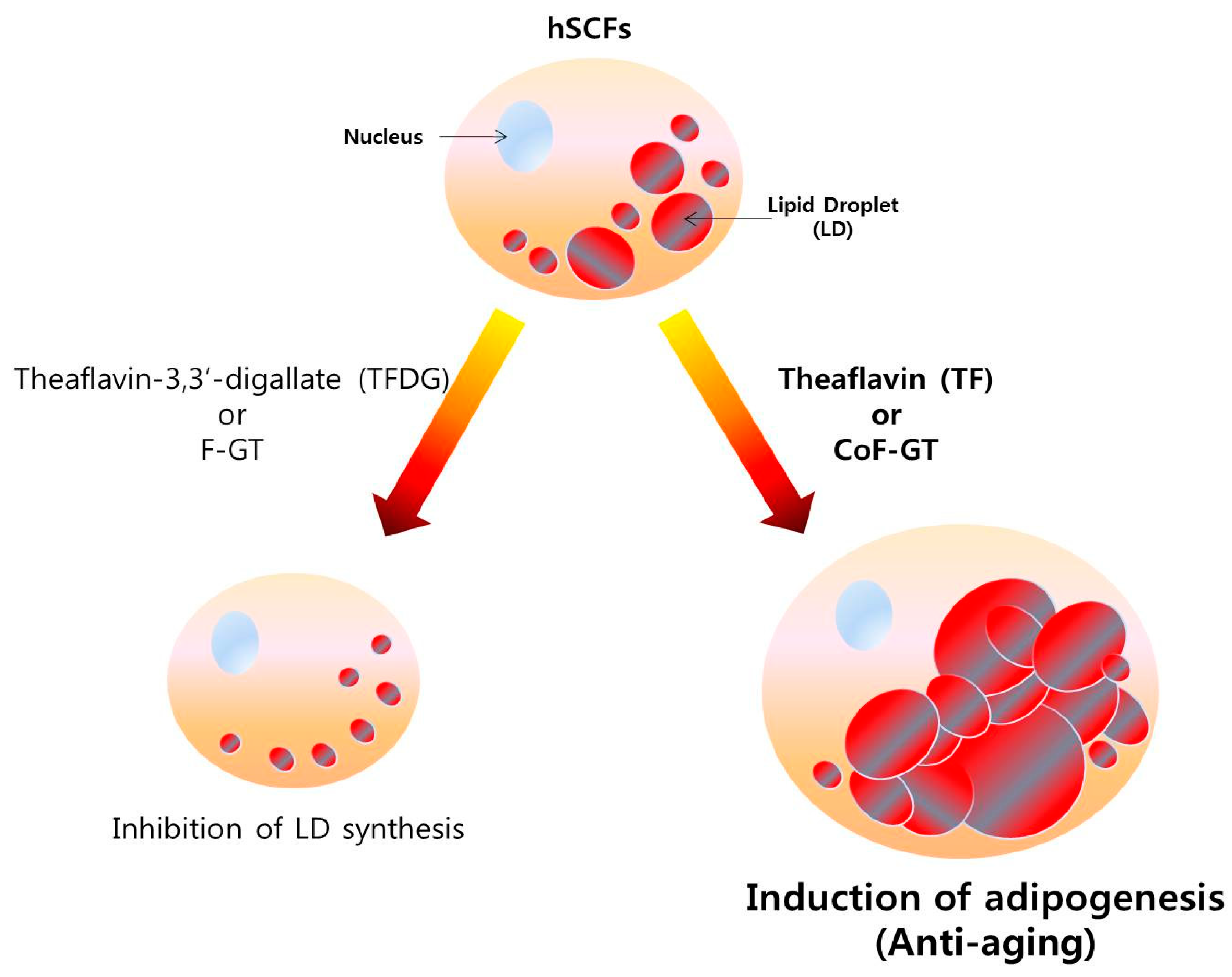
:1. Introduction
2. Results
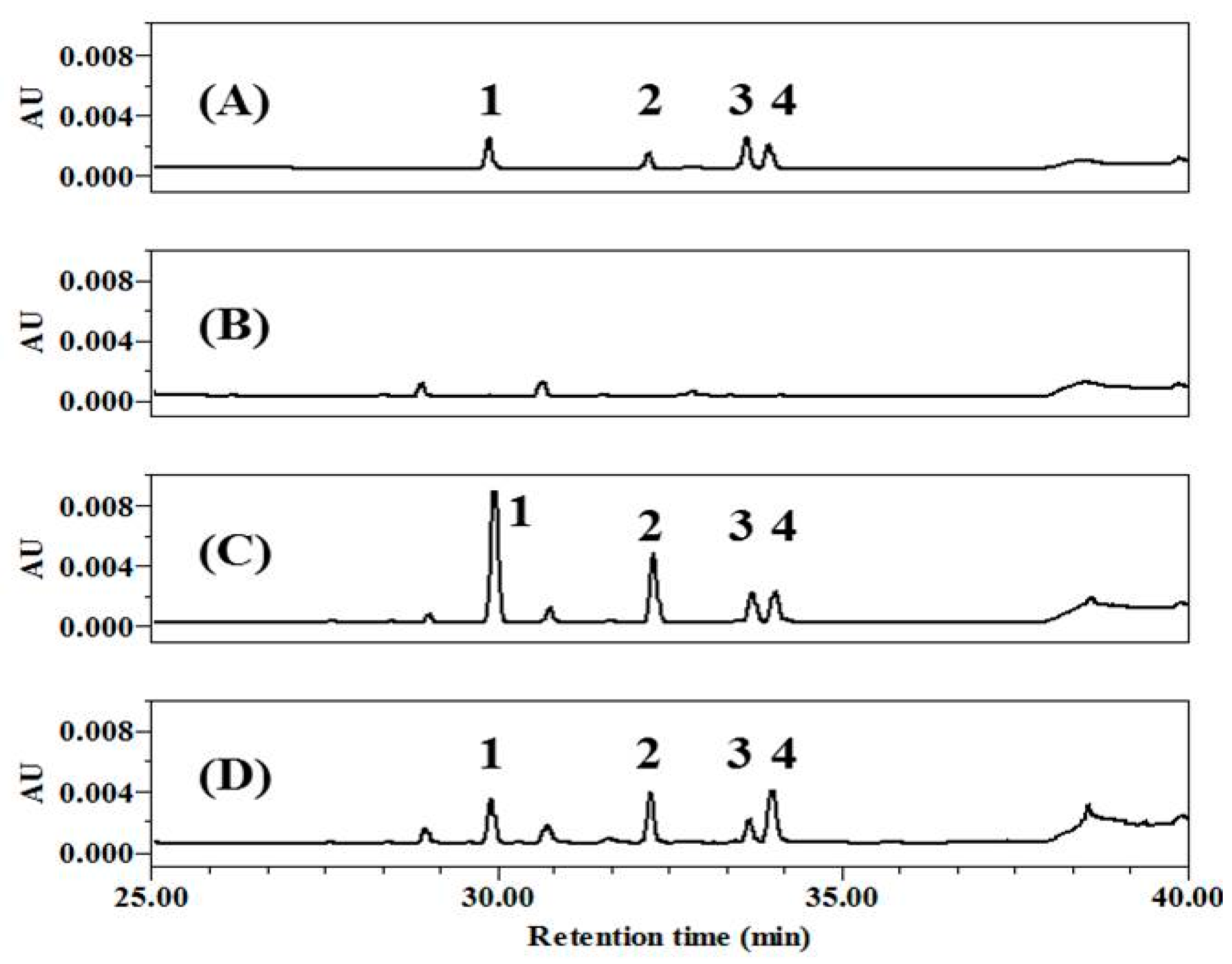
2.1. Preparation of Theaflavin (TF)-Enriched Fraction by Co-Fermentation with Chrysanthemum Coronarium (GC)
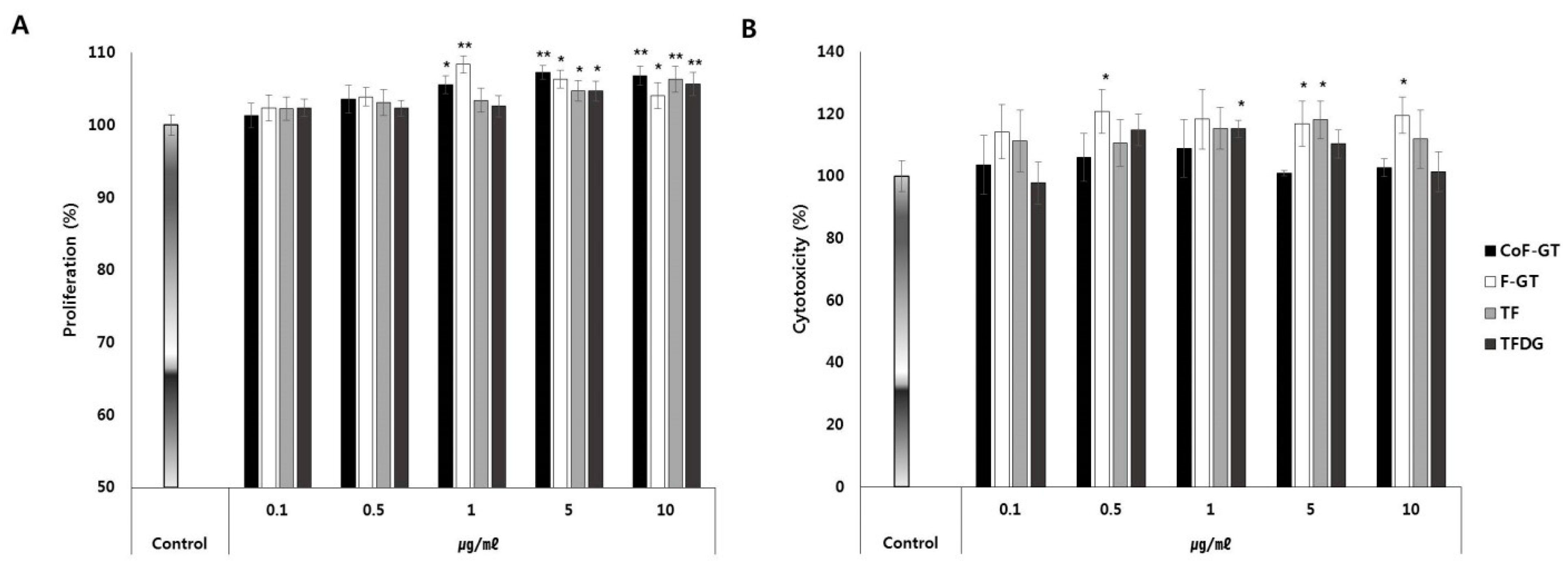
2.2. TF-Enriched Fraction Does Not Adversely Affect Human Subcutaneous Fat Cells (hSCF) Cell Viability
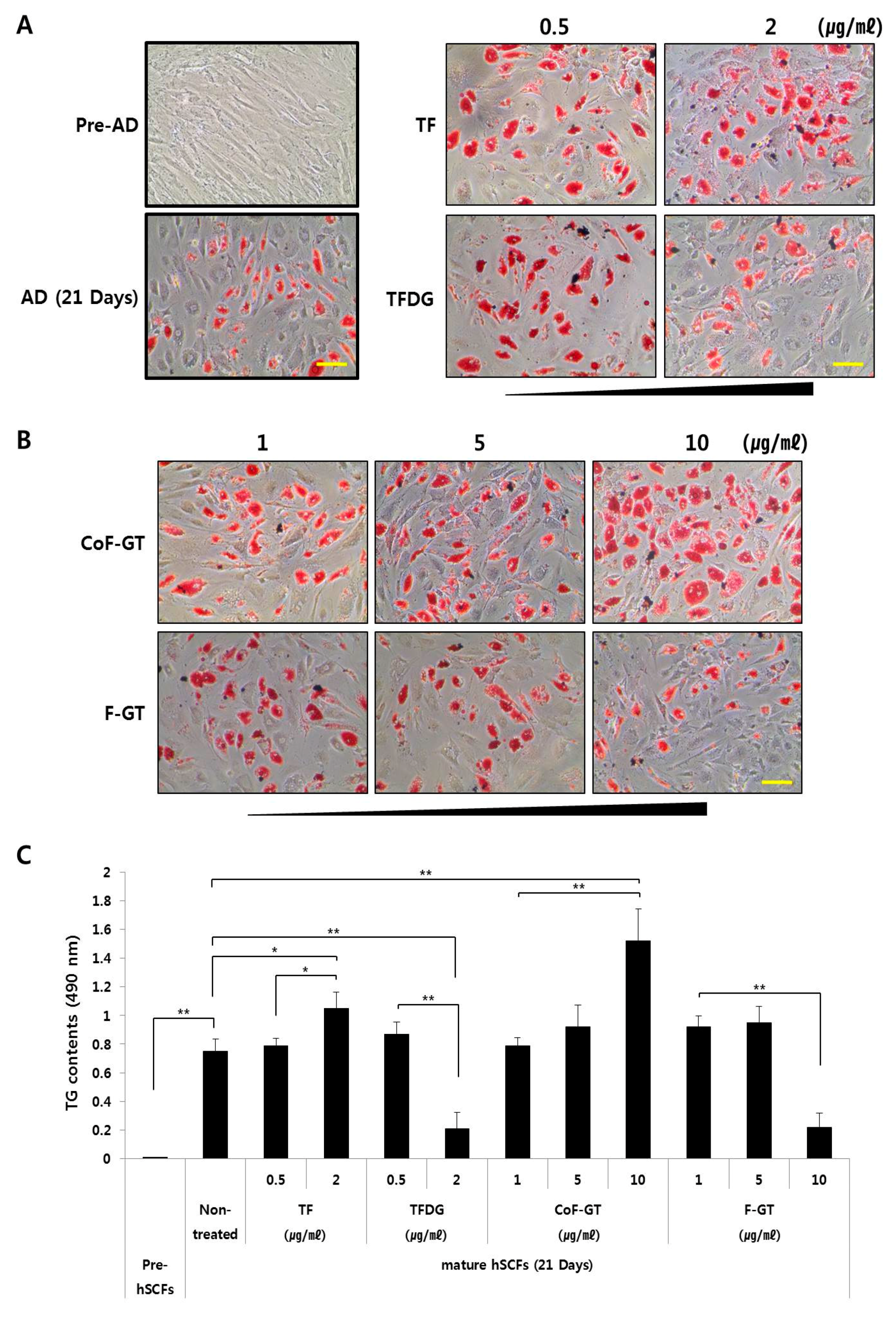
2.3. TF-Enriched Fraction Induces Lipogenesis in hSCF Cells
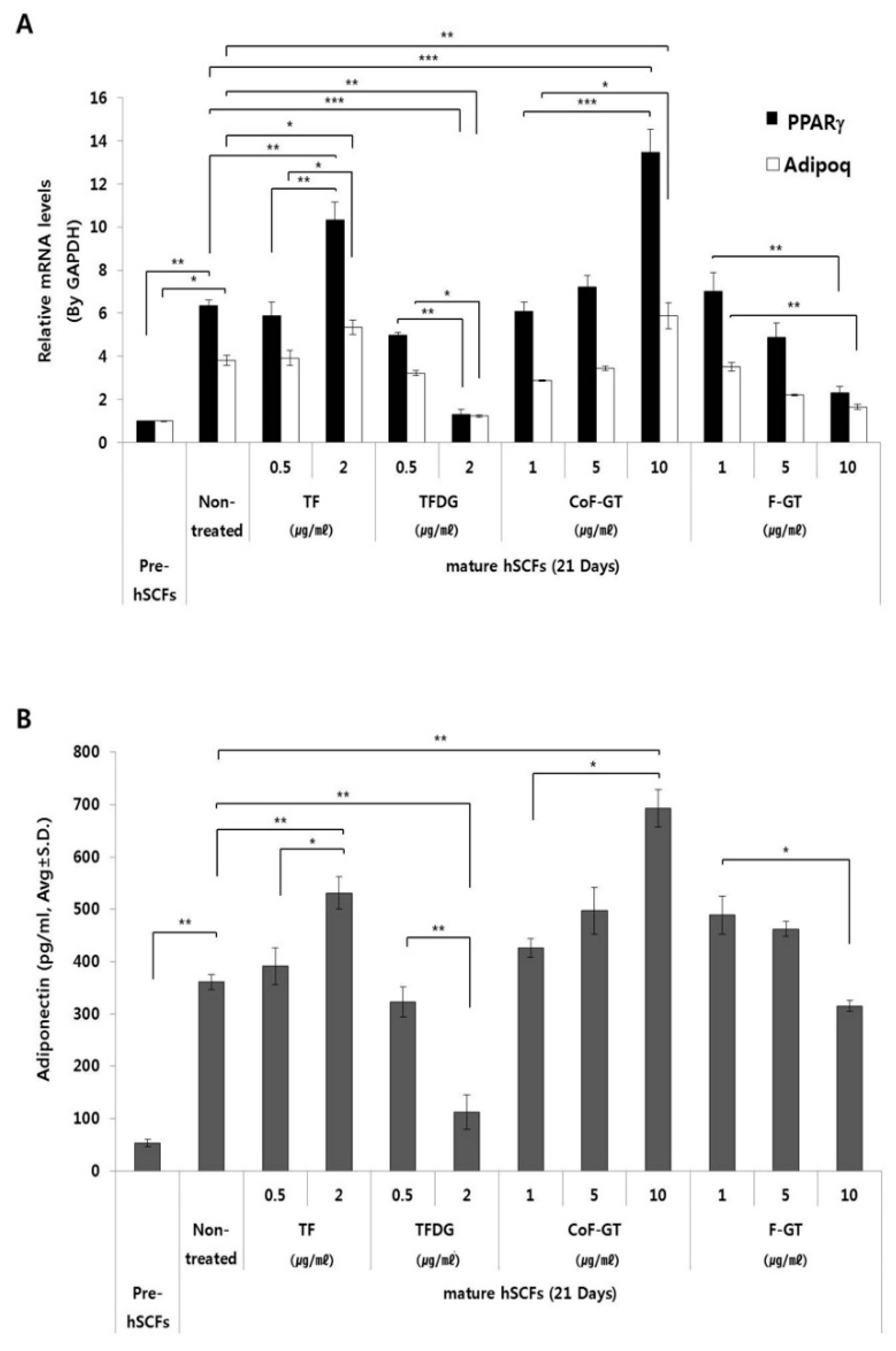
2.4. TF-Enriched Fraction Increases Expression of Lipogenesis-Related Genes in hSCF Cells
2.5. TF-Enriched Fraction Stimulates Adiponectin Secretion in hSCFs
3. Discussion
4. Materials and Methods
4.1. Reagents and Materials
4.2. Production of TF-Enriched CoF-GT and Conventionally Fermented F-GT Fractions
4.3. HPLC Analysis of CoF- and F-GT Fractions
4.4. Cell Culture and Differentiation
4.5. Cell Viability Assay
4.6. Analysis of Triglyceride (TG) Content by Oil Red O (ORO) Staining
4.7. Real-Time Quantitative Plymerase Chain Reaction (RT-qPCR)
4.8. Enzyme-Linked Immunosorbent Assay (ELISA) for Secreted Adiponectin
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADIPOQ | Adiponectin of human gene |
| APN | Adiponectin, secreted form mature adipocytes cytokine |
| CoF-GT | Co-fermented green tea fraction with high TF contents |
| F-GT | Conventionally fermented green tea |
| GC | Garland chrysanthemum |
| GT | Green tea |
| PPARγ | Peroxisome proliferator-activator receptor γ |
| TF | Theaflavin |
References
- Friedman, O. Changes associated with the aging face. Facial Plast. Surg. Clin. N. Am. 2005, 13, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Wollina, U.; Wetzker, R.; Abdel-Naser, M.B.; Kruglikov, I.L. Role of adipose tissue in facial aging. Clin. Interv. Aging 2017, 12, 2069–2076. [Google Scholar] [CrossRef] [Green Version]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Intrinsic and extrinsic factors in skin ageing: A review. Int. J. Cosmet. Sci. 2008, 30, 87–95. [Google Scholar] [CrossRef]
- Südel, K.M.; Venzke, K.; Mielke, H.; Breitenbach, U.; Mundt, C.; Jaspers, S.; Koop, U.; Sauermann, K.; Knussman-Hartig, E.; Moll, I.; et al. Novel aspects of intrinsic and extrinsic aging of human skin: Beneficial effects of soy extract. Photochem. Photobiol. 2005, 581–587. [Google Scholar] [CrossRef]
- Sotiropoulou, P.A.; Blanpain, C. Development and homeostasis of the skin epidermis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008383. [Google Scholar] [CrossRef]
- Kruglikov, I.L.; Scherer, P.E. Skin aging: Are adipocytes the next target? Aging (Albany NY) 2016, 8, 1457–1469. [Google Scholar] [CrossRef]
- Ezure, T.; Amano, S. Increment of subcutaneous adipose tissue is associated with decrease of elastic fibres in the dermal layer. Exp. Dermatol. 2015, 24, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Kruglikov, I.L.; Scherer, P.E. General theory of skin reinforcement. PLoS ONE 2017, 12, e0182865. [Google Scholar] [CrossRef]
- Pawlaczyk, M.; Lelonkiewicz, M.; Wieczorowski, M. Age-dependent biomechanical properties of the skin. Postepy Dermatol. Alergol. 2013, 30, 302–306. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.J.; Kim, Y.K.; Kim, J.E.; Kim, S.; Kim, M.K.; Park, C.H.; Chung, J.H. UV modulation of subcutaneous fat metabolism. J. Investig. Dermatol. 2011, 131, 1720–1726. [Google Scholar] [CrossRef] [PubMed]
- Lemieux, I. Energy partitioning in gluteal-femoral fat: does the metabolic fate of triglycerides affect coronary heart disease risk? Arterioscler. Thromb. Vasc. Biol. 2004, 24, 795–797. [Google Scholar] [CrossRef]
- Miranda, P.J.; De Fronzo, R.A.; Califf, R.M.; Guyton, J.R. Metabolic syndrome: Definition, pathophysiology, and mechanisms. Am. Heart J. 2005, 1, 33–45. [Google Scholar] [CrossRef]
- Petrofsky, J.S.; Prowse, M.; Lohman, E. The influence of ageing and diabetes on skin and subcutaneous fat thickness in different regions of the body. J. Appl. Res. 2008, 8, 55–62. [Google Scholar]
- Mukhtar, H.; Ahmad, N. Tea polyphenols: Prevention of cancer and optimizing health. Am. J. Clin. Nutr. 2000, 71, 1698S–1702S. [Google Scholar] [CrossRef]
- Tanaka, T.; Matsuo, Y.; Kouno, I. Chemistry of secondary polyphenols produced during processing of tea and selected foods. Int. J. Mol. Sci. 2010, 11, 14–40. [Google Scholar] [CrossRef]
- Lee, L.S.; Kim, Y.C.; Park, J.D.; Kim, Y.B.; Kim, S.H. Changes in major polyphenolic compounds of tea (Camellia sinensis) leaves during the production of black tea. Food Sci. Biotechnol. 2016, 25, 1523–1527. [Google Scholar] [CrossRef]
- Lin, C.L.; Huang, H.C.; Lin, J.K. Theaflavins attenuate hepatic lipid accumulation through activating AMPK in human HepG2 cells. J. Lipid Res. 2007, 48, 2334–2343. [Google Scholar] [CrossRef]
- Pan, H.; Gao, Y.; Tu, Y. Mechanisms of body weight reduction by black tea polyphenols. Molecules 2016, 21, 1659. [Google Scholar] [CrossRef]
- Yamashita, Y.; Wang, L.; Wang, L.; Tanaka, Y.; Zhang, T.; Ashida, H. Oolong, black and pu-erh tea suppresses adiposity in mice via activation of AMP-activated protein kinas. Food Funct. 2014, 5, 2420–2429. [Google Scholar] [CrossRef]
- Subramanian, N.; Venkatesh, P.; Ganguli, S.; Sinkar, V.P. Role of polyphenol oxidase and peroxidase in the generation of black tea theaflavins. J. Agric. Food Chem. 1999, 47, 2571–2578. [Google Scholar] [CrossRef]
- Li, S.; Lo, C.Y.; Pan, M.H.; Lai, C.S.; Ho, C.T. Black tea: Chemical analysis and stability. Food Funct. 2013, 4, 10–18. [Google Scholar] [CrossRef]
- Hamdaoui, M.H.; Snoussi, C.; Dhaouadi, K.; Fattouch, S.; Ducroc, R.; Le Gall, M.; Bado, A. Tea decoctions prevent body weight gain in rats fed high-fat diet; black tea being more efficient than green tea. J. Nutr. Intermed. Metab. 2016, 6, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Nomura, M.; Ma, W.Y.; Huang, C.; Yang, C.S.; Bowden, G.T.; Miyamoto, K.I.; Dong, Z. Inhibition of ultraviolet B-induced AP-1 activation by theaflavins from black tea. Mol. Carcinog. 2000, 28, 148–155. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Ohguchi, K.; Itoh, T.; Nozawa, Y.; Akao, Y. Effects of theaflavins on melanin biosynthesis in mouse B16 melanoma cells. Biosci. Biotechnol. Biochem. 2009, 73, 1429–1431. [Google Scholar] [CrossRef]
- Kim, Y.C.; Choi, S.Y.; Park, E.Y. Anti-melanogenic effects of black, green, and white tea extracts on immortalized melanocytes. J. Vet. Sci. 2015, 16, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Pou, K.J. Fermentation: The key step in the processing of black tea. J. Biosyst. Eng. 2016, 41, 85–92. [Google Scholar] [CrossRef]
- Obanda, M.; Owuor, P.O.; Mang’oka, R. Changes in the chemical and sensory quality parameters of black tea due to variations of fermentation time and temperature. Food Chem. 2001, 75, 395–404. [Google Scholar] [CrossRef]
- Torre-Villalvazo, I.; Bunt, A.E.; Alemán, G.; Marquez-Mota, C.C.; Diaz-Villaseñor, A.; Noriega, L.G.; Estrada, I.; Figueroa-Juárez, E.; Tovar-Palacio, C.; Rodriguez-López, L.A.; et al. Adiponectin synthesis and secretion by subcutaneous adipose tissue is impaired during obesity by endoplasmic reticulum stress. J. Cell. Biochem. 2018, 119, 5970–5984. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Tea polyphenols for health promotion. Life Sci. 2007, 81, 519–533. [Google Scholar] [CrossRef] [Green Version]
- Chacko, S.M.; Thambi, P.T.; Kuttan, R.; Nishigaki, I. Beneficial effects of green tea: A literature review. Chin. Med. 2010, 5, 13. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Tea and health: Studies in humans. Curr. Pharm. Des. 2013, 19, 6141–6147. [Google Scholar] [CrossRef]
- Meydani, M.; Hasan, S.T. Dietary polyphenols and obesity. Nutrients 2010, 2, 737–751. [Google Scholar] [CrossRef]
- Wang, S.; Moustaid-Moussa, N.; Chen, L.; Mo, H.; Shastri, A.; Su, R.; Bapat, P.; Kwun, I.; Shen, C.L. Novel insights of dietary polyphenols and obesity. J. Nutr. Biochem. 2014, 25, 1–8. [Google Scholar] [CrossRef]
- Sajilata, M.G.; Bajaj, P.R.; Singhal, R.S. Tea polyphenols as nutraceuticals. Compr. Rev. Food Sci. Food Saf. 2008, 7, 229–254. [Google Scholar] [CrossRef]
- Higdon, J.V.; Frei, B. Tea catechins and polyphenols: Health effects, metabolism, and antioxidant functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef]
- Lin, J.K.; Lin-Shiau, S.Y. Mechanisms of hypolipidemic and anti-obesity effects of tea and tea polyphenols. Mol. Nutr. Food Res. 2006, 50, 211–217. [Google Scholar] [CrossRef]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef]
- Kraemer, F.B.; Shen, W.J. Hormone-sensitive lipase: Control of intracellular tri-(di-)acylglycerol and cholesteryl ester hydrolysis. J. Lipid Res. 2002, 43, 1585–1594. [Google Scholar] [CrossRef]
- Oikarinen, A. Connective tissue and aging. Int. J. Cosmet. Sci. 2004, 26, 107. [Google Scholar] [CrossRef]
- Madison, K.C. Barrier function of the skin: “la raison d’être” of the epidermis. J. Investig. Dermatol. 2003, 121, 231–241. [Google Scholar] [CrossRef]
- Kim, S.O.; Han, Y.; Ahn, S.; An, S.; Shin, J.C.; Choi, H.; Kim, H.J.; Park, N.H.; Kim, Y.J.; Jin, S.H.; et al. Kojyl cinnamate esters are peroxisome proliferator-activated receptor α/γ dual agonists. Bioorg. Med. Chem. 2018, 26, 5654–5663. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, P.J.; Rha, C.-S.; Kim, S.T. Theaflavin-Enriched Fraction Stimulates Adipogenesis in Human Subcutaneous Fat Cells. Int. J. Mol. Sci. 2019, 20, 2034. https://doi.org/10.3390/ijms20082034
Park PJ, Rha C-S, Kim ST. Theaflavin-Enriched Fraction Stimulates Adipogenesis in Human Subcutaneous Fat Cells. International Journal of Molecular Sciences. 2019; 20(8):2034. https://doi.org/10.3390/ijms20082034
Chicago/Turabian StylePark, Phil June, Chan-Su Rha, and Sung Tae Kim. 2019. "Theaflavin-Enriched Fraction Stimulates Adipogenesis in Human Subcutaneous Fat Cells" International Journal of Molecular Sciences 20, no. 8: 2034. https://doi.org/10.3390/ijms20082034
APA StylePark, P. J., Rha, C.-S., & Kim, S. T. (2019). Theaflavin-Enriched Fraction Stimulates Adipogenesis in Human Subcutaneous Fat Cells. International Journal of Molecular Sciences, 20(8), 2034. https://doi.org/10.3390/ijms20082034