Mast Cells in Early Rheumatoid Arthritis

 , , and
, , and
Abstract
:
1. Synovial Inflammation in Rheumatoid Arthritis
2. Emerging Role of Mast Cells in Synovial Inflammation
3. Synovial Mast Cells in Early RA
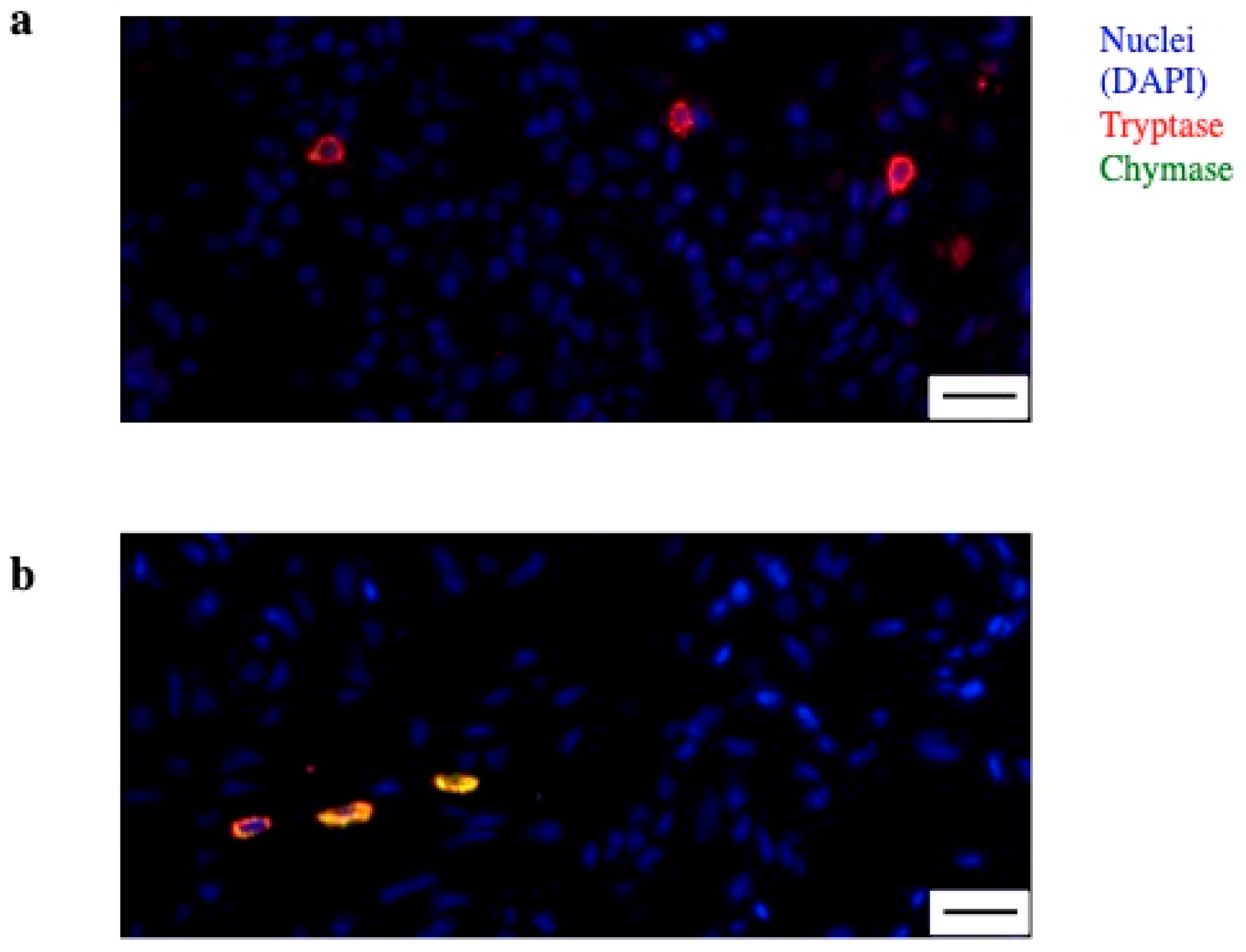
3.1. Synovial MC Heterogeneity
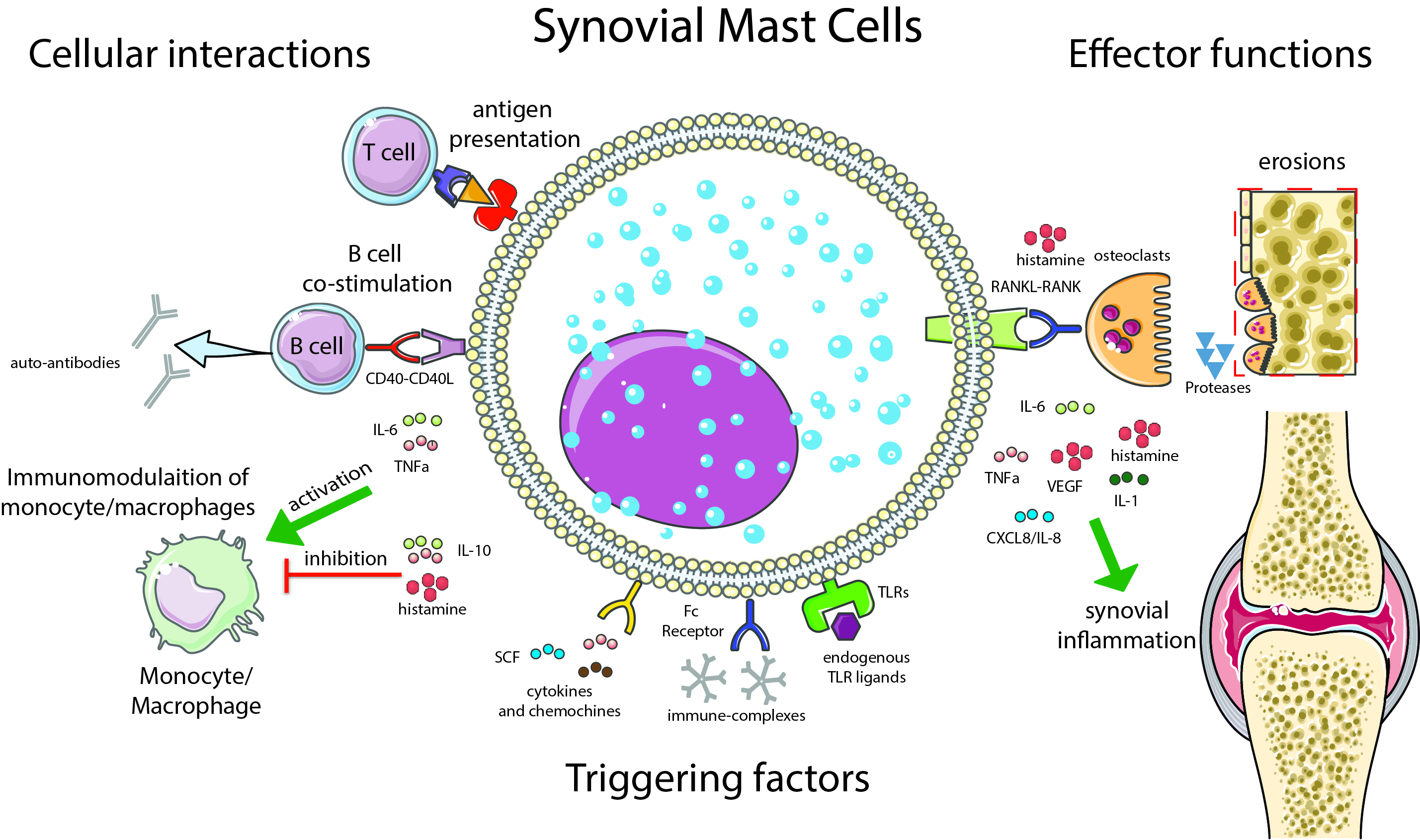
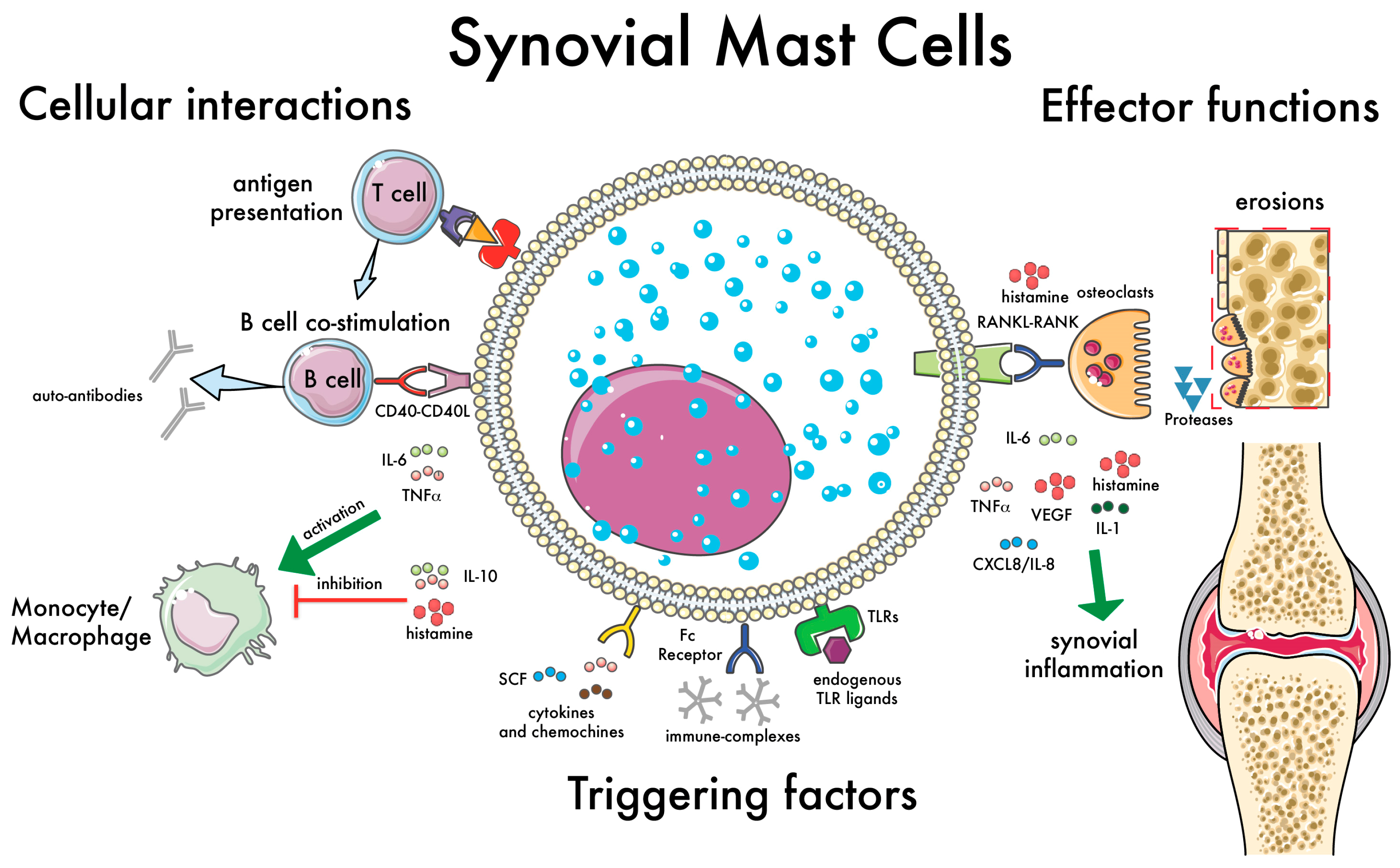
3.2. Synovial Mast Cells as Effector Cells in RA
3.3. Synovial Mast Cells and the Activation of Lymphocyte
3.4. Mast Cells as Immunomodulatory Cells in RA
3.5. Synovial Mast Cells as Markers of Disease Severity in Early and Established RA
4. Concluding Remarks and Future Work
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACPAs | anti-citrullinated protein antibodies |
| CRP C | reactive protein |
| DAPI | 4′,6-diamidino-2-phenylindole |
| ESR | erytrocyte sedimentation rate |
| FcεRI | Fc epsilon receptor I |
| MC | mast cell |
| MC_TC | mast cell expressing tryptase and chymase |
| MC_T | mast cell expressing tryptase |
| RANK | receptor activator of nuclear factor kappa-β |
| RANKL | receptor activator of nuclear factor kappa-β ligand |
| SCF | stem cell factor |
| sDMARDs | synthetic disease modifying anti-rheumatic drugs |
| TLR | toll-like receptors |
| TNFα | tumor necrosis factor alpha |
| VEGF | vascular endothelial growth factor |
References
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef] [Green Version]
- Pitzalis, C.; Kelly, S.; Humby, F. New learnings on the pathophysiology of RA from synovial biopsies. Curr. Opin. Rheumatol. 2013, 25, 334–344. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. The Pathogenesis of Rheumatoid Arthritis. N. Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef]
- Rivellese, F.; Lobasso, A.; Barbieri, L.; Liccardo, B.; De Paulis, A.; Rossi, F.W. Novel therapeutic approaches in Rheumatoid Arthritis: Role of Janus Kinases Inhibitors. Curr. Med. Chem. 2018, 25. [Google Scholar] [CrossRef] [PubMed]
- Burmester, G.R.; Pope, J.E. Novel treatment strategies in rheumatoid arthritis. Lancet (London, England) 2017, 389, 2338–2348. [Google Scholar] [CrossRef]
- McInnes, I.B.; Schett, G. Pathogenetic insights from the treatment of rheumatoid arthritis. Lancet (London, England) 2017, 389, 2328–2337. [Google Scholar] [CrossRef] [Green Version]
- Huizinga, T.W. Personalized medicine in rheumatoid arthritis: Is the glass half full or half empty. J. Intern. Med. 2014. [Google Scholar] [CrossRef]
- Willemze, A.; Trouw, L.A.; Toes, R.E.M.; Huizinga, T.W.J. The influence of ACPA status and characteristics on the course of RA. Nat. Rev. Rheumatol. 2012, 8, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Alivernini, S.; Galeazzi, M.; Peleg, H.; Tolusso, B.; Gremese, E.; Ferraccioli, G.; Naparstek, Y. Is ACPA positivity the main driver for rheumatoid arthritis treatment? Pros and cons. Autoimmun. Rev. 2017, 16, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Humby, F.; Lewis, M.; Ramamoorthi, N.; Hackney, J.A.; Barnes, M.R.; Bombardieri, M.; Setiadi, F.; Kelly, S.; Bene, F.; Dicicco, M.; et al. Synovial cellular and molecular signatures stratify clinical response to csDMARD therapy and predict radiographic progression in early rheumatoid arthritis patients. Ann Rheum Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Borriello, F.; Granata, F.; Varricchi, G.; Genovese, A.; Triggiani, M.; Marone, G. Immunopharmacological modulation of mast cells. Curr. Opin. Pharmacol. 2014, 17, 45–57. [Google Scholar] [CrossRef]
- Marone, G.; Borriello, F.; Varricchi, G.; Genovese, A.; Granata, F. Basophils: Historical reflections and perspectives. Chem. Immunol. Allergy 2014, 100, 172–192. [Google Scholar] [PubMed]
- Genovese, A.; Borgia, G.; Björck, L.; Petraroli, A.; de Paulis, A.; Piazza, M.; Marone, G. Immunoglobulin superantigen protein L induces IL-4 and IL-13 secretion from human Fc epsilon RI+ cells through interaction with the kappa light chains of IgE. J. Immunol. 2003, 170, 1854–1861. [Google Scholar] [CrossRef] [PubMed]
- Galdiero, M.R.; Varricchi, G.; Seaf, M.; Marone, G.; Levi-Schaffer, F.; Marone, G. Bidirectional Mast Cell–Eosinophil Interactions in Inflammatory Disorders and Cancer. Front. Med. 2017, 4, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [Green Version]
- Kurashima, Y.; Amiya, T.; Fujisawa, K.; Shibata, N.; Suzuki, Y.; Kogure, Y.; Hashimoto, E.; Otsuka, A.; Kabashima, K.; Sato, S.; et al. The Enzyme Cyp26b1 Mediates Inhibition of Mast-Cell Activation by Fibroblasts to Maintain Skin-Barrier Homeostasis. Immunity 2014, 40, 530–541. [Google Scholar] [CrossRef]
- Detoraki, A.; Staiano, R.I.; Granata, F.; Giannattasio, G.; Prevete, N.; de Paulis, A.; Ribatti, D.; Genovese, A.; Triggiani, M.; Marone, G. Vascular endothelial growth factors synthesized by human lung mast cells exert angiogenic effects. J. Allergy Clin. Immunol. 2009, 123, 1142–1149, 1149.e1–1149.e5. [Google Scholar] [CrossRef]
- Marone, G.; Varricchi, G.; Loffredo, S.; Granata, F. Mast cells and basophils in inflammatory and tumor angiogenesis and lymphangiogenesis. Eur. J. Pharmacol. 2016, 778, 146–151. [Google Scholar] [CrossRef]
- Douaiher, J.; Succar, J.; Lancerotto, L.; Gurish, M.F.; Orgill, D.P.; Hamilton, M.J.; Krilis, S.A.; Stevens, R.L. Development of Mast Cells and Importance of Their Tryptase and Chymase Serine Proteases in Inflammation and Wound Healing. Adv. Immunol. 2014, 122, 211–252. [Google Scholar] [Green Version]
- Varricchi, G.; Galdiero, M.R.; Marone, G.; Granata, F.; Borriello, F.; Marone, G. Controversial role of mast cells in skin cancers. Exp. Dermatol. 2017, 26, 11–17. [Google Scholar] [CrossRef]
- Giannou, A.D.; Marazioti, A.; Spella, M.; Kanellakis, N.I.; Apostolopoulou, H.; Psallidas, I.; Prijovich, Z.M.; Vreka, M.; Zazara, D.E.; Lilis, I.; et al. Mast cells mediate malignant pleural effusion formation. J. Clin. Investig. 2015, 125, 2317–2334. [Google Scholar] [CrossRef] [PubMed]
- Galdiero, M.R.; Varricchi, G.; Marone, G. The immune network in thyroid cancer. Oncoimmunology 2016, 5, e1168556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivellese, F.; Nerviani, A.; Rossi, F.W.; Marone, G.; Matucci-Cerinic, M.; de Paulis, A.; Pitzalis, C. Mast cells in rheumatoid arthritis: Friends or foes? Autoimmun. Rev. 2017, 16, 557–563. [Google Scholar] [CrossRef]
- De Paulis, A.; Marinò, I.; Ciccarelli, A.; De Crescenzo, G.; Concardi, M.; Verga, L.; Arbustini, E.; Marone, G. Human synovial mast cells: I. Ultrastructural in situ and in vitro immunologic characterization. Arthritis Rheum. 1996, 39, 1222–1233. [Google Scholar] [CrossRef] [PubMed]
- De Paulis, A.; Ciccarelli, A.; Marinò, I.; de Crescenzo, G.; Marinò, D.; Marone, G. Human synovial mast cells. II. Heterogeneity of the pharmacologic effects of antiinflammatory and immunosuppressive drugs. Arthritis Rheum. 1997, 40, 469–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noordenbos, T.; Yeremenko, N.; Gofita, I.; van de Sande, M.; Tak, P.P.; Caňete, J.D.; Baeten, D. Interleukin-17-positive mast cells contribute to synovial inflammation in spondylarthritis. Arthritis Rheum. 2012, 64, 99–109. [Google Scholar] [CrossRef] [PubMed]
- De Lange-Brokaar, B.J.E.; Kloppenburg, M.; Andersen, S.N.; Dorjée, a.L.; Yusuf, E.; Herb-van Toorn, L.; Kroon, H.M.; Zuurmond, A.M.; Stojanovic-Susulic, V.; Bloem, J.L.; et al. Characterization of synovial mast cells in knee osteoarthritis: Association with clinical parameters. Osteoarthr. Cartil. 2016, 24, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Suurmond, J.; Van Der Velden, D.; Kuiper, J.; Bot, I.; Toes, R.E.M. Mast cells in rheumatic disease. Eur. J. Pharmacol. 2016, 778, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Van der Velden, D.; Lagraauw, H.M.; Wezel, A.; Launay, P.; Kuiper, J.; Huizinga, T.W.J.; Toes, R.E.M.; Bot, I.; Stoop, J.N. Mast cell depletion in the preclinical phase of collagen-induced arthritis reduces clinical outcome by lowering the inflammatory cytokine profile. Arthritis Res. Ther. 2016, 18, 138. [Google Scholar] [CrossRef]
- Schubert, N.; Dudeck, J.; Liu, P.; Karutz, A.; Speier, S.; Maurer, M.; Tuckermann, J.; Dudeck, A. Mast cell promotion of T cell-driven antigen-induced arthritis despite being dispensable for antibody-induced arthritis in which T cells are bypassed. Arthritis Rheumatol. (Hoboken, N.J.) 2015, 67, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Rivellese, F.; Mauro, D.; Nerviani, A.; Pagani, S.; Fossati-Jimack, L.; Messemaker, T.; Kurreeman, F.A.S.; Toes, R.E.M.; Ramming, A.; Rauber, S.; et al. Mast cells in early rheumatoid arthritis associate with disease severity and support B cell autoantibody production. Ann. Rheum. Dis. 2018, 77, 1773–1781. [Google Scholar] [CrossRef]
- Crisp, A.J.; Chapman, C.M.; Kirkham, S.E.; Schiller, A.L.; Krane, S.M. Articular mastocytosis in rheumatoid arthritis. Arthritis Rheum. 1984, 27, 845–851. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, H.P.; Ilardi, C.; Engber, W.; Graziano, F.M. Quantitation of Human Synovial Mast Cells in Rheumatoid Arthritis and Other Rheumatic Diseases. Arthritis Rheum. 1984, 27, 852–856. [Google Scholar] [CrossRef]
- Malone, D.G.; Wilder, R.L.; Saavedra-Delgado, A.M.; Metcalfe, D.D. Mast cell numbers in rheumatoid synovial tissues. Correlations with quantitative measures of lymphocytic infiltration and modulation by antiinflammatory therapy. Arthritis Rheum. 1987, 30, 130–137. [Google Scholar] [CrossRef]
- Tetlow, L.C.; Woolley, D.E. Distribution, activation and tryptase/chymase phenotype of mast cells in the rheumatoid lesion. Ann. Rheum. Dis. 1995, 54, 549–555. [Google Scholar] [CrossRef]
- Gotis-Graham, I.; McNeil, H.P. Mast cell responses in rheumatoid synovium. Association of the MCTC subset with matrix turnover and clinical progression. Arthritis Rheum. 1997, 40, 479–489. [Google Scholar] [CrossRef] [Green Version]
- Gotis-Graham, I.; Smith, M.D.; Parker, a.; McNeil, H.P. Synovial mast cell responses during clinical improvement in early rheumatoid arthritis. Ann. Rheum. Dis. 1998, 57, 664–671. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, J.; Celis, R.; Usategui, A.; Ruiz-Esquide, V.; Faré, R.; Cuervo, A.; Sanmartí, R.; Pablos, J.L.; Cañete, J.D. Immunopathologic characterization of ultrasound-defined synovitis in rheumatoid arthritis patients in clinical remission. Arthritis Res. Ther. 2016, 18, 74. [Google Scholar] [CrossRef]
- Dwyer, D.F.; Barrett, N.A.; Austen, K.F.; Dwyer, D.F.; Barrett, N.A.; Austen, K.F.; Kim, E.Y.; Brenner, M.B.; Shaw, L.; Yu, B.; et al. Expression profiling of constitutive mast cells reveals a unique identity within the immune system. Nat. Immunol. 2016. [Google Scholar] [CrossRef]
- Motakis, E.; Guhl, S.; Ishizu, Y.; Itoh, M.; Kawaji, H.; de Hoon, M.; Lassmann, T.; Carninci, P.; Hayashizaki, Y.; Zuberbier, T.; et al. Redefinition of the human mast cell transcriptome by deep-CAGE sequencing. Blood 2014, 123, e58–e67. [Google Scholar] [CrossRef] [Green Version]
- Cildir, G.; Pant, H.; Lopez, A.F.; Tergaonkar, V. The transcriptional program, functional heterogeneity, and clinical targeting of mast cells. J. Exp. Med. 2017, 214, 2491–2506. [Google Scholar] [CrossRef]
- Galli, S.J.; Borregaard, N.; Wynn, T.A. Phenotypic and functional plasticity of cells of innate immunity: Macrophages, mast cells and neutrophils. Nat Immunol. 2011, 12, 1035–1044. [Google Scholar] [CrossRef]
- Bradding, P.; Arthur, G. Mast cells in asthma—state of the art. Clin. Exp. Allergy 2016, 46, 194–263. [Google Scholar] [CrossRef]
- Rossi, F.W.; Prevete, N.; Rivellese, F.; Lobasso, A.; Napolitano, F.; Granata, F.; Selleri, C.; de Paulis, A. HIV-1 Nef promotes migration and chemokine synthesis of human basophils and mast cells through the interaction with CXCR4. Clin. Mol. Allergy 2016, 14, 15. [Google Scholar] [CrossRef]
- Marone, G.; Varricchi, G.; Loffredo, S.; Galdiero, M.R.; Rivellese, F.; De Paulis, A. Are Basophils and Mast Cells Masters in HIV Infection? Int. Arch. Allergy Immunol. 2017, 171, 158–165. [Google Scholar] [CrossRef]
- Rossi, F.W.; Prevete, N.; Rivellese, F.; Napolitano, F.; Montuori, N.; Postiglione, L.; Selleri, C.; De Paulis, A. The Urokinase/Urokinase Receptor System in Mast Cells: Effects of its Functional Interaction with fMLF Receptors. Transl. Med. UniSa 2016, 15, 34–41. [Google Scholar]
- Okamura, Y.; Mishima, S.; Kashiwakura, J.I.; Sasaki-Sakamoto, T.; Toyoshima, S.; Kuroda, K.; Saito, S.; Tokuhashi, Y.; Okayama, Y. The dual regulation of substance P-mediated inflammation via human synovial mast cells in rheumatoid arthritis. Allergol. Int. 2017, 66, S9–S20. [Google Scholar] [CrossRef]
- Suurmond, J.; Rivellese, F.; Dorjée, a.L.; Bakker, A.M.; Rombouts, Y.J.P.C.; Rispens, T.; Wolbink, G.; Zaldumbide, A.; Hoeben, R.C.; Huizinga, T.W.J.; et al. Toll-like receptor triggering augments activation of human mast cells by anti-citrullinated protein antibodies. Ann. Rheum. Dis. 2015, 74, 1915–1923. [Google Scholar] [CrossRef]
- Sandler, C.; Lindstedt, K.a.; Joutsiniemi, S.; Lappalainen, J.; Juutilainen, T.; Kolah, J.; Kovanen, P.T.; Eklund, K.K. Selective activation of mast cells in rheumatoid synovial tissue results in production of TNF-alpha, IL-1beta and IL-1Ra. Inflamm. Res. 2007, 56, 230–239. [Google Scholar] [CrossRef]
- Olsson, N.; Ulfgren, A.K.; Nilsson, G. Demonstration of mast cell chemotactic activity in synovial fluid from rheumatoid patients. Ann. Rheum. Dis. 2001, 60, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Buckley, M.G.; Walters, C.; Wong, W.M.; Cawley, M.I.; Ren, S.; Schwartz, L.B.; Walls, A.F. Mast cell activation in arthritis: Detection of alpha- and beta-tryptase, histamine and eosinophil cationic protein in synovial fluid. Clin. Sci. (Lond). 1997, 93, 363–370. [Google Scholar] [CrossRef]
- Sawamukai, N.; Yukawa, S.; Saito, K.; Nakayamada, S.; Kambayashi, T.; Tanaka, Y. Mast cell-derived tryptase inhibits apoptosis of human rheumatoid synovial fibroblasts via rho-mediated signaling. Arthritis Rheum. 2010, 62, 952–959. [Google Scholar] [CrossRef] [Green Version]
- Arock, M. Mast cell differentiation: Still open questions? Blood 2016, 127, 373–374. [Google Scholar] [CrossRef]
- Lee, H.; Kashiwakura, J.; Matsuda, A.; Watanabe, Y.; Sakamoto-Sasaki, T.; Matsumoto, K.; Hashimoto, N.; Saito, S.; Ohmori, K.; Nagaoka, M.; et al. Activation of human synovial mast cells from rheumatoid arthritis or osteoarthritis patients in response to aggregated IgG through Fcγ receptor I and Fcγ receptor II. Arthritis Rheum. 2013, 65, 109–119. [Google Scholar] [CrossRef]
- Suurmond, J.; van Heemst, J.; van Heiningen, J.; Dorjée, A.L.; Schilham, M.W.; van der Beek, F.B.; Huizinga, T.W.J.; Schuerwegh, A.J.M.; Toes, R.E.M. Communication between human mast cells and CD4(+) T cells through antigen-dependent interactions. Eur. J. Immunol. 2013, 43, 1758–1768. [Google Scholar] [CrossRef]
- Suurmond, J.; Dorjée, A.L.; Huizinga, T.W.J.; Toes, R.E.M. Human mast cells co-stimulate T cells through a CD28-independent interaction. Eur. J. Immunol. 2016, 46, 1132–1141. [Google Scholar] [CrossRef]
- Rivellese, F.; Suurmond, J.; Habets, K.; Dorjée, A.L.; Ramamoorthi, N.; Townsend, M.J.; de Paulis, A.; Marone, G.; Huizinga, T.W.J.; Pitzalis, C.; et al. Ability of Interleukin-33- and Immune Complex-Triggered Activation of Human Mast Cells to Down-Regulate Monocyte-Mediated Immune Responses. Arthritis Rheumatol. 2015, 67, 2343–2353. [Google Scholar] [CrossRef]
- Shin, K.; Nigrovic, P.A.; Crish, J.; Boilard, E.; McNeil, H.P.; Larabee, K.S.; Adachi, R.; Gurish, M.F.; Gobezie, R.; Stevens, R.L.; et al. Mast Cells Contribute to Autoimmune Inflammatory Arthritis via Their Tryptase/Heparin Complexes. J. Immunol. 2008, 182, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.S.; Lax, A.; Liljeström, M.; Paakkari, I.; Ashammakhi, N.; Kovanen, P.T.; Konttinen, Y.T. Mast cells in atherosclerosis as a source of the cytokine RANKL. Clin. Chem. Lab. Med. 2006, 44, 672–674. [Google Scholar]
- Biosse-Duplan, M.; Baroukh, B.; Dy, M.; de Vernejoul, M.-C.; Saffar, J.-L. Histamine promotes osteoclastogenesis through the differential expression of histamine receptors on osteoclasts and osteoblasts. Am. J. Pathol. 2009, 174, 1426–1434. [Google Scholar] [CrossRef]
- Lotfi-Emran, S.; Ward, B.R.; Le, Q.T.; Pozez, A.L.; Manjili, M.H.; Woodfolk, J.; Schwartz, L.B. Human mast cells present antigen to autologous CD4 + T Cells. J. Allergy Clin. Immunol. 2017. [Google Scholar] [CrossRef]
- Suurmond, J.; Habets, K.L.L.; Dorjée, A.L.; Huizinga, T.W.; Toes, R.E.M. Expansion of Th17 Cells by Human Mast Cells Is Driven by Inflammasome-Independent IL-1β. J. Immunol. 2016, 197, 4473–4481. [Google Scholar] [CrossRef]
- Edwards, J.C.W.; Szczepański, L.; Szechiński, J.; Filipowicz-Sosnowska, A.; Emery, P.; Close, D.R.; Stevens, R.M.; Shaw, T. Efficacy of B-Cell–Targeted Therapy with Rituximab in Patients with Rheumatoid Arthritis. N. Engl. J. Med. 2004, 350, 2572–2581. [Google Scholar] [CrossRef] [Green Version]
- Kramm, H.; Hansen, K.E.; Gowing, E.; Bridges, A. Successful Therapy of Rheumatoid Arthritis with Rituximab: Renewed Interest in the Role of B Cells in the Pathogenesis of Rheumatoid Arthritis. J. Clin. Rheumatol. 2004, 10, 28–32. [Google Scholar] [CrossRef]
- Humby, F.; Bombardieri, M.; Manzo, A.; Kelly, S.; Blades, M.C.; Kirkham, B.; Spencer, J.; Pitzalis, C. Ectopic Lymphoid Structures Support Ongoing Production of Class-Switched Autoantibodies in Rheumatoid Synovium. PLoS Med. 2009, 6, e1. [Google Scholar] [CrossRef] [Green Version]
- Rosengren, S.; Wei, N.; Kalunian, K.C.; Zvaifler, N.J.; Kavanaugh, A.; Boyle, D.L. Elevated autoantibody content in rheumatoid arthritis synovia with lymphoid aggregates and the effect of rituximab. Arthritis Res. Ther. 2008, 10, R105. [Google Scholar] [CrossRef]
- Yeo, L.; Toellner, K.-M.; Salmon, M.; Filer, A.; Buckley, C.D.; Raza, K.; Scheel-Toellner, D. Cytokine mRNA profiling identifies B cells as a major source of RANKL in rheumatoid arthritis. Ann. Rheum. Dis. 2011, 70, 2022–2028. [Google Scholar] [CrossRef] [Green Version]
- Harre, U.; Jurdic, P.; Schett, G.; Georgess, D.; Bang, H.; Bozec, A.; Axmann, R.; Ossipova, E.; Jakobsson, P.-J.; Baum, W.; et al. Induction of osteoclastogenesis and bone loss by human autoantibodies against citrullinated vimentin. J. Clin. Investig. 2012, 122, 1791–1802. [Google Scholar] [CrossRef] [Green Version]
- Laurent, L.; Anquetil, F.; Clavel, C.; Ndongo-Thiam, N.; Offer, G.; Miossec, P.; Pasquali, J.-L.; Sebbag, M.; Serre, G. IgM rheumatoid factor amplifies the inflammatory response of macrophages induced by the rheumatoid arthritis-specific immune complexes containing anticitrullinated protein antibodies. Ann. Rheum. Dis. 2014, 1–7. [Google Scholar] [CrossRef]
- Clavel, C.; Nogueira, L.; Laurent, L.; Iobagiu, C.; Vincent, C.; Sebbag, M.; Serre, G. Induction of macrophage secretion of tumor necrosis factor alpha through Fcgamma receptor IIa engagement by rheumatoid arthritis-specific autoantibodies to citrullinated proteins complexed with fibrinogen. Arthritis Rheum. 2008, 58, 678–688. [Google Scholar] [CrossRef]
- Merluzzi, S.; Frossi, B.; Gri, G.; Parusso, S.; Tripodo, C.; Pucillo, C. Mast cells enhance proliferation of B lymphocytes and drive their differentiation toward IgA-secreting plasma cells. Blood 2010, 115, 2810–2817. [Google Scholar] [CrossRef] [Green Version]
- Mion, F.; D’Incà, F.; Danelli, L.; Toffoletto, B.; Guarnotta, C.; Frossi, B.; Burocchi, A.; Rigoni, A.; Gerdes, N.; Lutgens, E.; et al. Mast Cells Control the Expansion and Differentiation of IL-10–Competent B Cells. J. Immunol. 2014, 193, 4568–4579. [Google Scholar] [CrossRef] [Green Version]
- Carroll-Portillo, A.; Cannon, J.L.; Te Riet, J.; Holmes, A.; Kawakami, Y.; Kawakami, T.; Cambi, A.; Lidke, D.S. Mast cells and dendritic cells form synapses that facilitate antigen transfer for T cell activation. J. Cell Biol. 2015, 210, 851–864. [Google Scholar] [CrossRef] [Green Version]
- Mantri, C.K.; St. John, A.L. Immune synapses between mast cells and γδ T cells limit viral infection. J. Clin. Investig. 2018, 129, 1094–1108. [Google Scholar] [CrossRef]
- Galli, S.J.; Grimbaldeston, M.; Tsai, M. Immunomodulatory mast cells: Negative, as well as positive, regulators of immunity. Nat. Rev. Immunol. 2008, 8, 478–486. [Google Scholar] [CrossRef]
- Elenkov, I.J.; Webster, E.; Papanicolaou, D.A.; Fleisher, T.a.; Chrousos, G.P.; Wilder, R.L. Histamine potently suppresses human IL-12 and stimulates IL-10 production via H2 receptors. J. Immunol. 1998, 161, 2586–2593. [Google Scholar]
- Rivellese, F.; Suurmond, J.; de Paulis, A.; Marone, G.; Huizinga, T.W.J.; Toes, R.E.M. IgE and IL-33-mediated triggering of human basophils inhibits TLR4-induced monocyte activation. Eur. J. Immunol. 2014, 3045–3055. [Google Scholar] [CrossRef]
- Rossini, M.; Viapiana, O.; Zanoni, G.; Tripi, G.; Idolazzi, L.; Fracassi, E.; Adami, G.; Adami, S.; Gatti, D. Serum Levels of Tryptase Suggest That Mast Cells Might Have an Antiinflammatory Role in Rheumatoid Arthritis: Comment on the Article by Rivellese et al. Arthritis Rheumatol. 2016, 68, 769. [Google Scholar] [CrossRef]
- Rivellese, F.; De Paulis, A.; Marone, G.; Pitzalis, C.; Toes, R.E.M. Reply. Arthritis Rheumatol. 2016, 68, 769–770. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Year | Title | Disease Duration | Procedure (Joint Site) | Patients (n) | Main Results |
|---|---|---|---|---|---|
| Crisp et al. 1984 [32] | Articular mastocytosis in rheumatoid arthritis | Long-standing | Synoviectomy (wrist and knee) and joint replacement (knee) | 116 | Higher MC numbers in RA vs. healthy samples. Correlation with synovitis but not with ESR. Caveat treatment. |
| Godfrey et al. 1985 [33] | Mast Cells in Rheumatoid Arthritis and Other Rheumatic Diseases | Long-standing | Synoviectomy or joint replacement | 14 | Higher MC numbers in RA. Association with clinically active disease. |
| Malone et al. 1987 [34] | Mast cell numbers in rheumatoid synovial tissues. | Long-standing | Knee arthroscopy | 20 | Correlation with synovial inflammation. No attempt to look for clinical correlations. Reduction of MC numbers after IA steroids. |
| Tetlow & Woolley 1995 [35] | Distribution, activation and tryptase/chymase phenotype of mast cells in the rheumatoid | Long-standing | Joint replacement (knee) | 26 | MC present in all samples, but more abundant in 60% of patients. Higher prevalence of MC_T. No attempt to look for clinical correlations. |
| Gotis-Graham & McNeil 1997 [36] | Mast cell responses in rheumatoid synovium. Association of the MCTC subset with matrix turnover and clinical progression | Long-standing | Joint replacement or arthroscopy (knee) | 16 | Increase of MCs in RA vs. OA or healthy samples; MC_TC correlate with disease activity. |
| Gotis-Graham et al. 1998 [37] | Synovial mast cell responses during clinical improvement in early rheumatoid arthritis | Early (mean disease duration 8 months), sDMARD-naïve | Arthroscopy (knee) | 6 | Relative increase of MC_T was observed, in correlation with synovial inflammatory score. |
| Ramírez et al. 2016 [38] | Immunopathologic characterization of ultrasound-defined synovitis in rheumatoid arthritis patients in clinical remission. | Long standing; 20 patients in remission; 22 with active disease | US-guided synovial biopsies | 42 | MCs and B cells in patients in clinical remission associated with disease reactivation. |
| Rivellese et al. 2018 [31] | Mast cells in early rheumatoid arthritis associate with disease severity and support B cell autoantibody production | Early (<12 months), steroid and sDMARD-naive | US-guided synovial biopsies (60% small joints, i.e., MCPs or wrists) | 99 | Synovial MCs are associated with (i) synovial inflammation, (ii) lympho-myeloid infiltrate, (iii) systemic inflammation, (iv) autoantibodies, and (v) disease activity. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivellese, F.; Rossi, F.W.; Galdiero, M.R.; Pitzalis, C.; de Paulis, A. Mast Cells in Early Rheumatoid Arthritis. Int. J. Mol. Sci. 2019, 20, 2040. https://doi.org/10.3390/ijms20082040
Rivellese F, Rossi FW, Galdiero MR, Pitzalis C, de Paulis A. Mast Cells in Early Rheumatoid Arthritis. International Journal of Molecular Sciences. 2019; 20(8):2040. https://doi.org/10.3390/ijms20082040
Chicago/Turabian StyleRivellese, Felice, Francesca Wanda Rossi, Maria Rosaria Galdiero, Costantino Pitzalis, and Amato de Paulis. 2019. "Mast Cells in Early Rheumatoid Arthritis" International Journal of Molecular Sciences 20, no. 8: 2040. https://doi.org/10.3390/ijms20082040
APA StyleRivellese, F., Rossi, F. W., Galdiero, M. R., Pitzalis, C., & de Paulis, A. (2019). Mast Cells in Early Rheumatoid Arthritis. International Journal of Molecular Sciences, 20(8), 2040. https://doi.org/10.3390/ijms20082040