Expression and Function of Transient Receptor Potential Ankyrin 1 Ion Channels in the Caudal Nucleus of the Solitary Tract



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
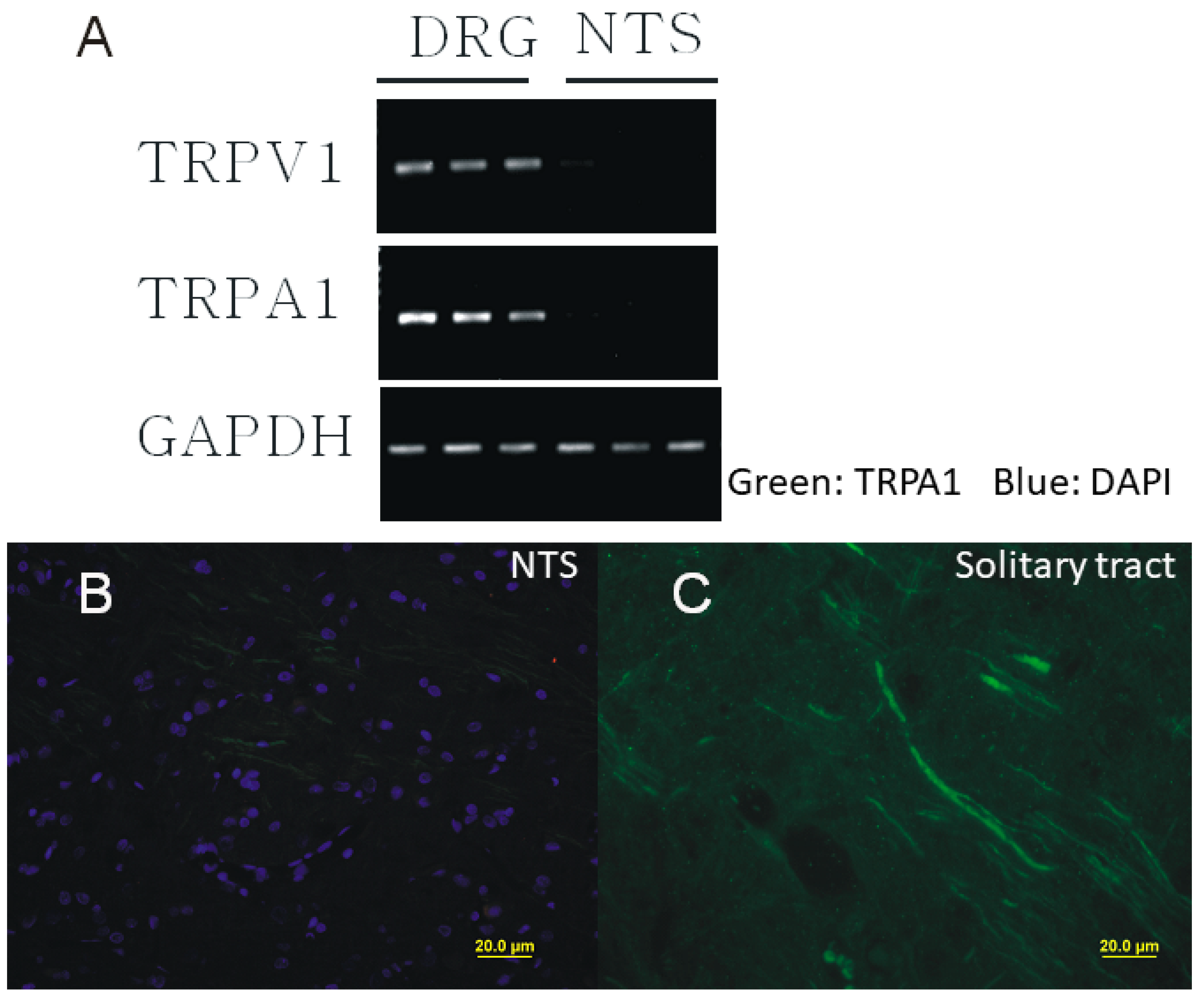
2.1. Presynaptic Expression of TRPA1 in the Caudal NTS
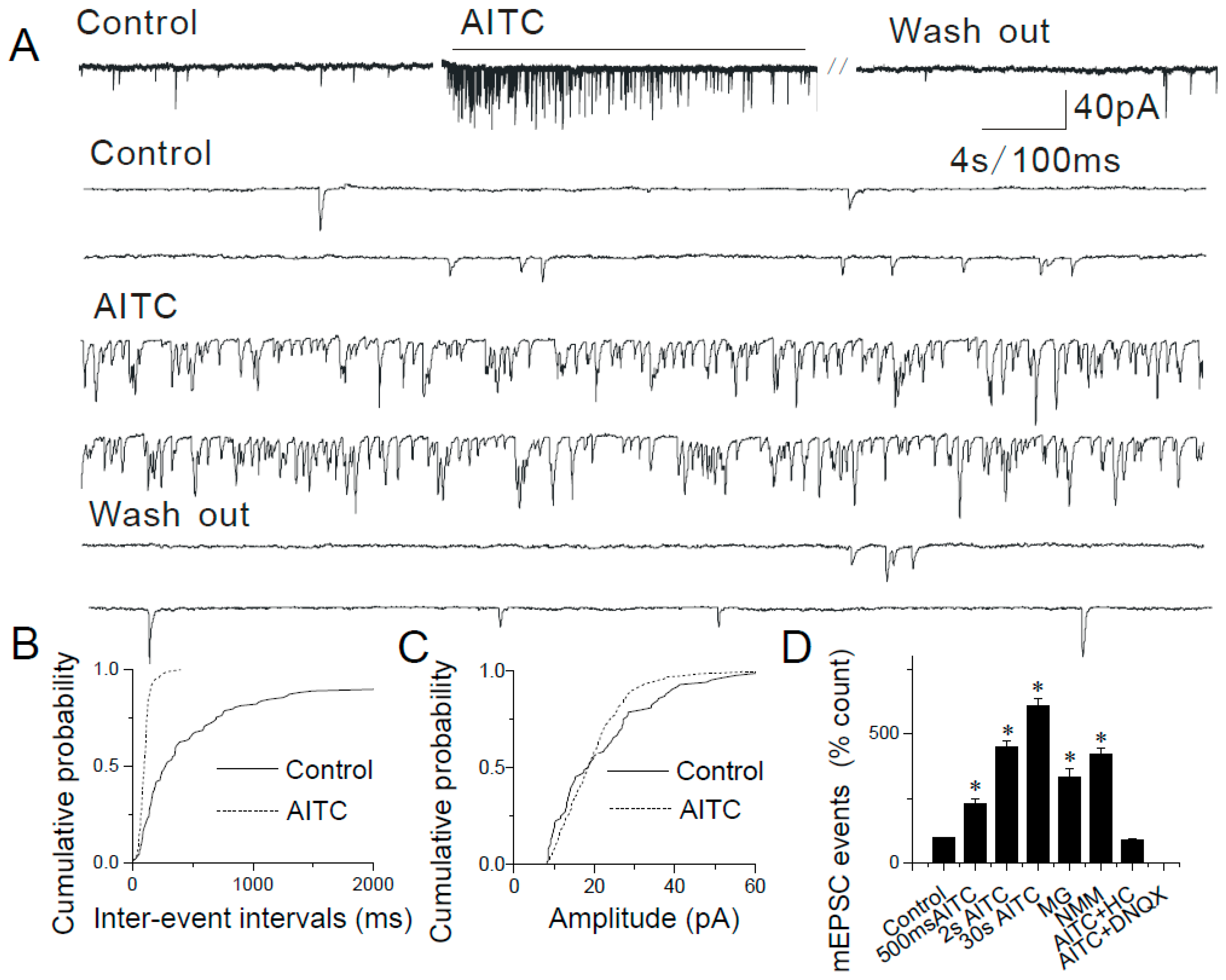
2.2. Modulation of Synaptic Transmission by AITC in the Caudal NTS
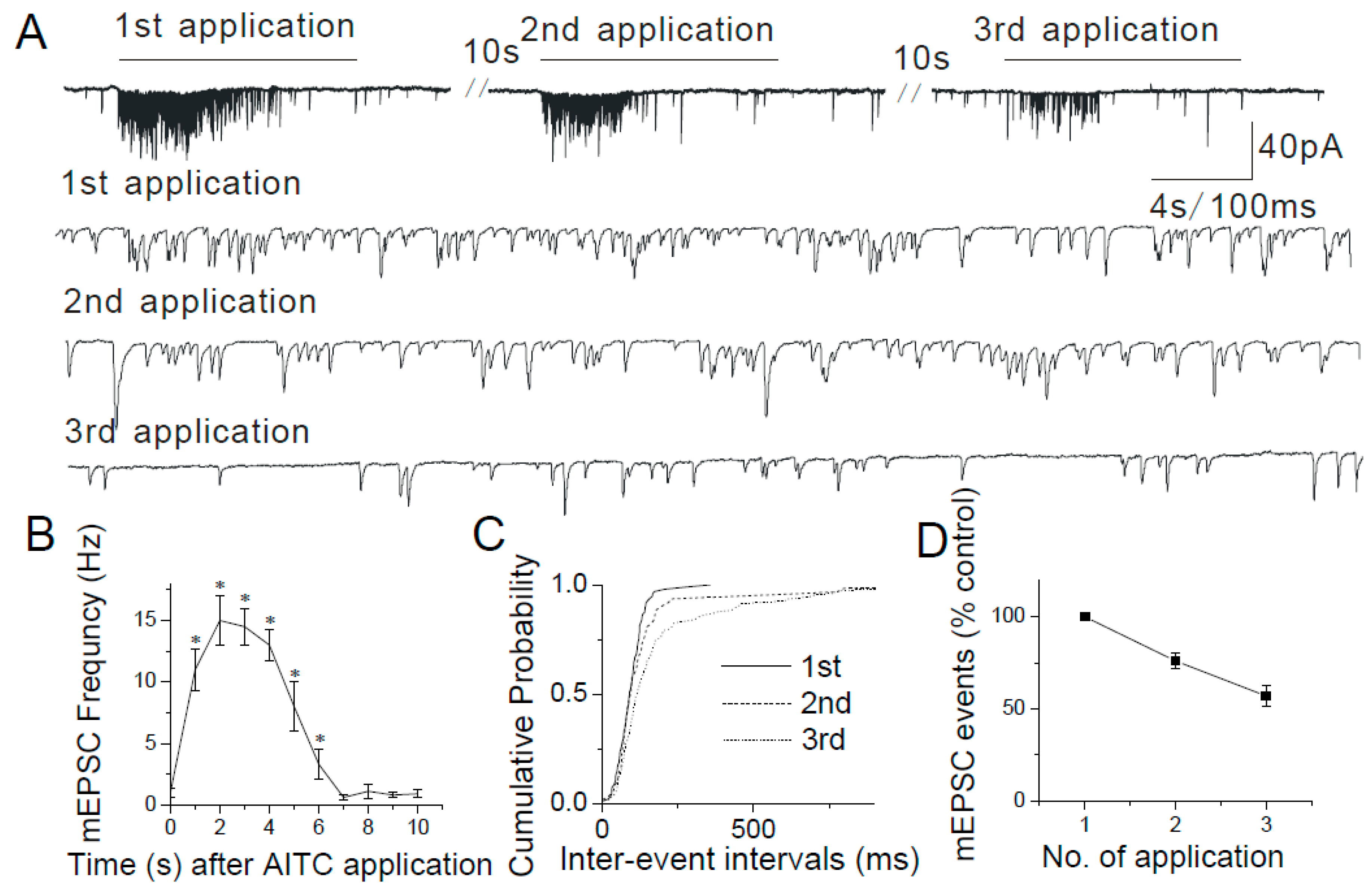
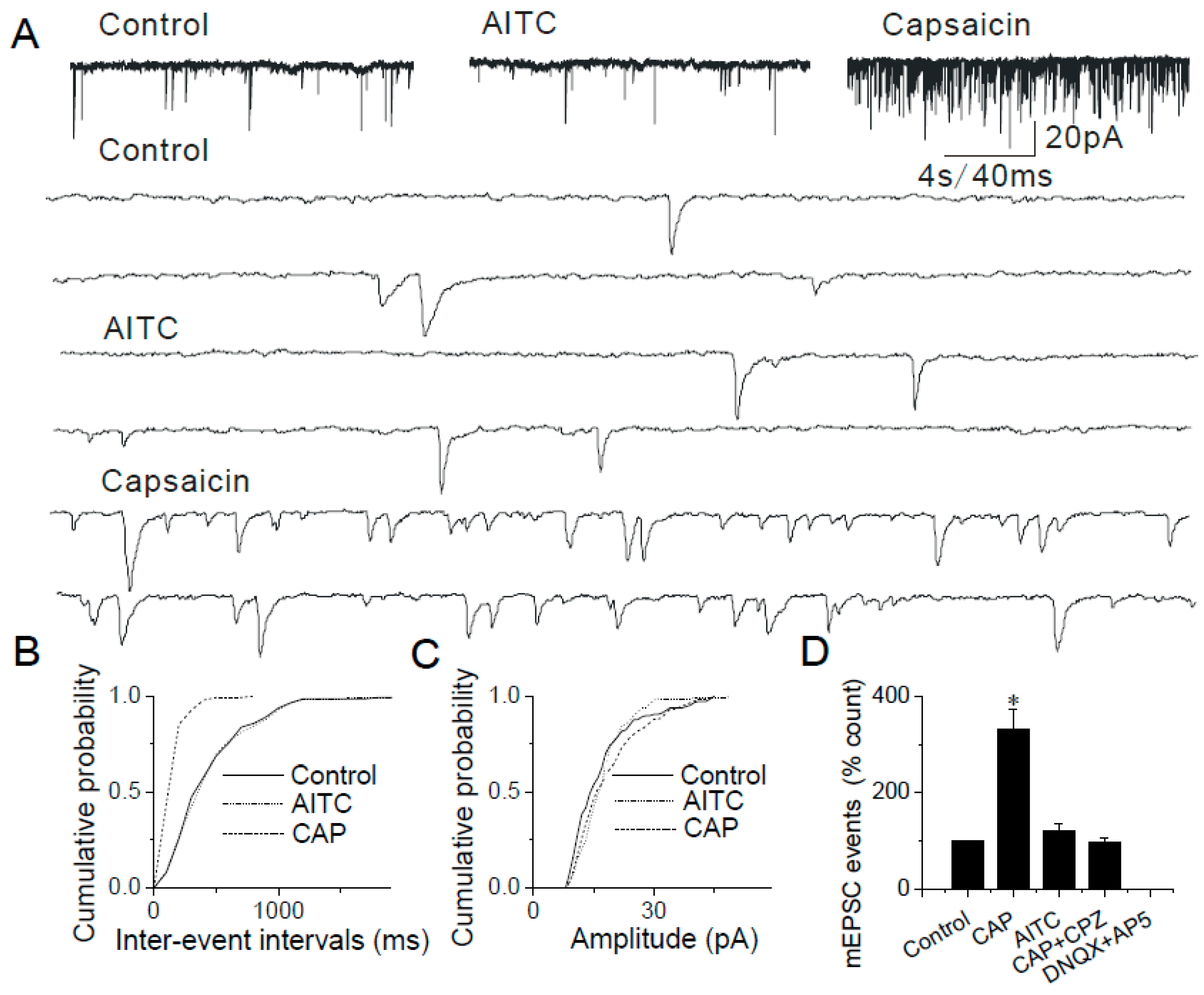
2.3. Changes in mEPSCs in Response to Continuous and Repeated Application of AITC
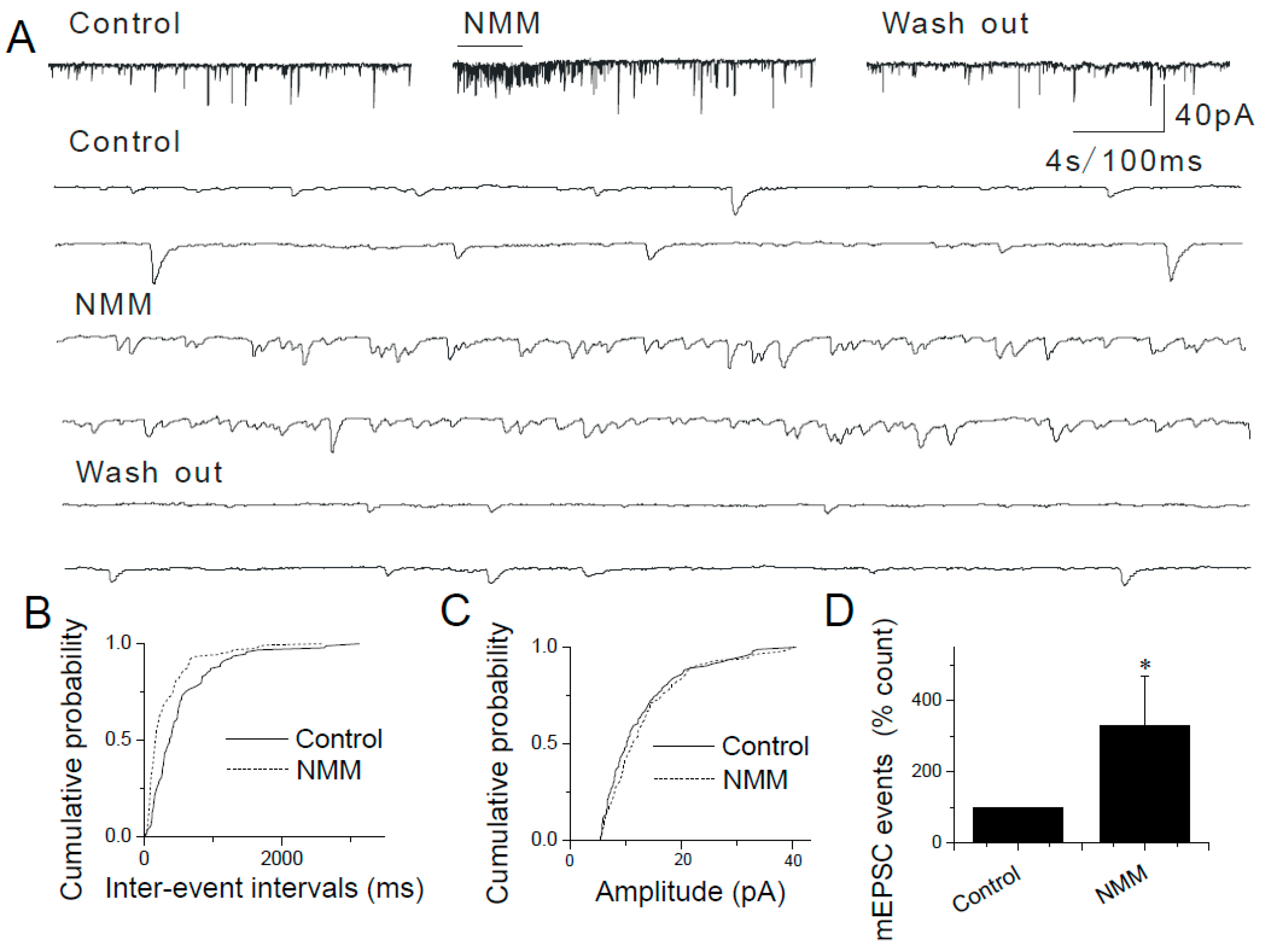
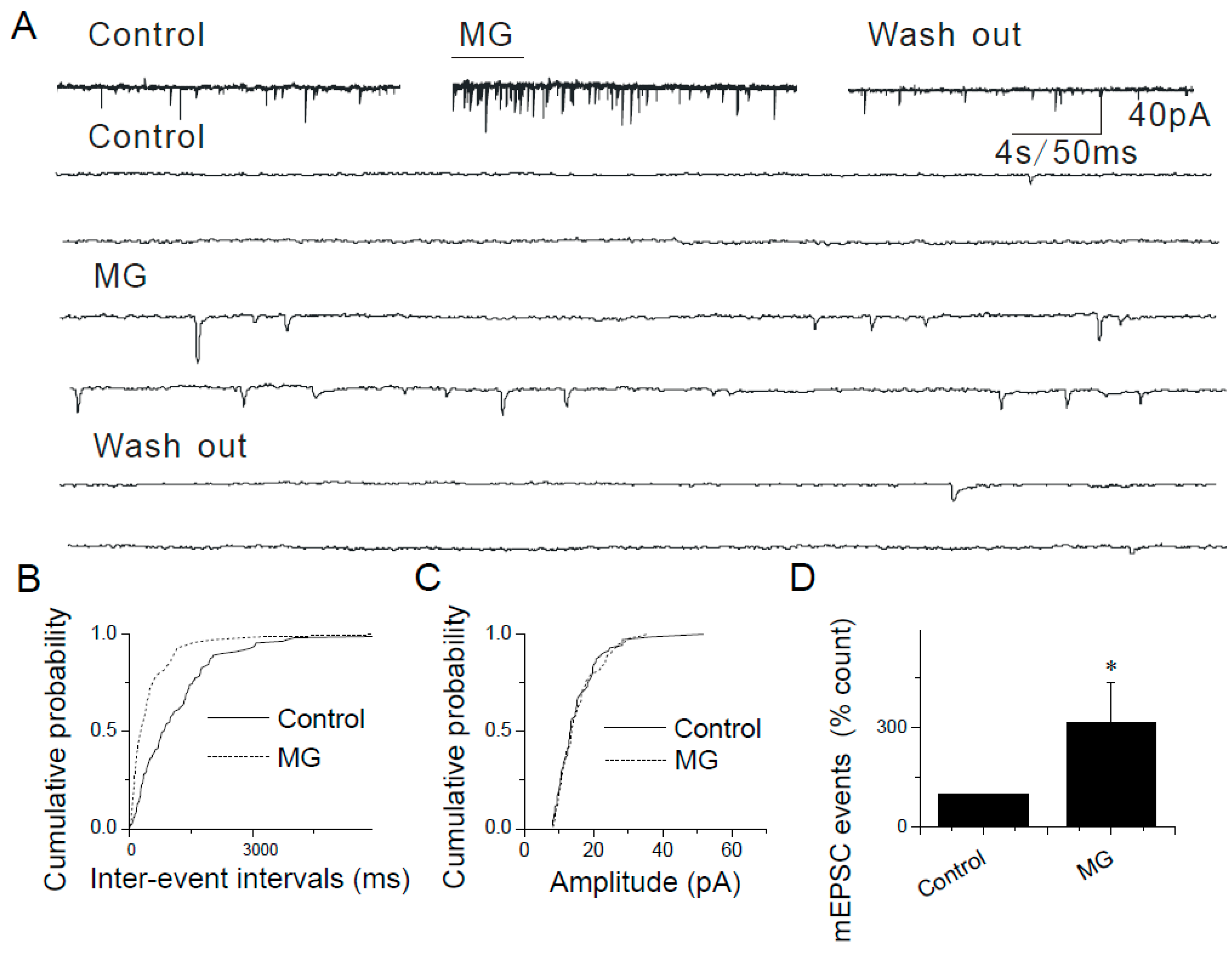
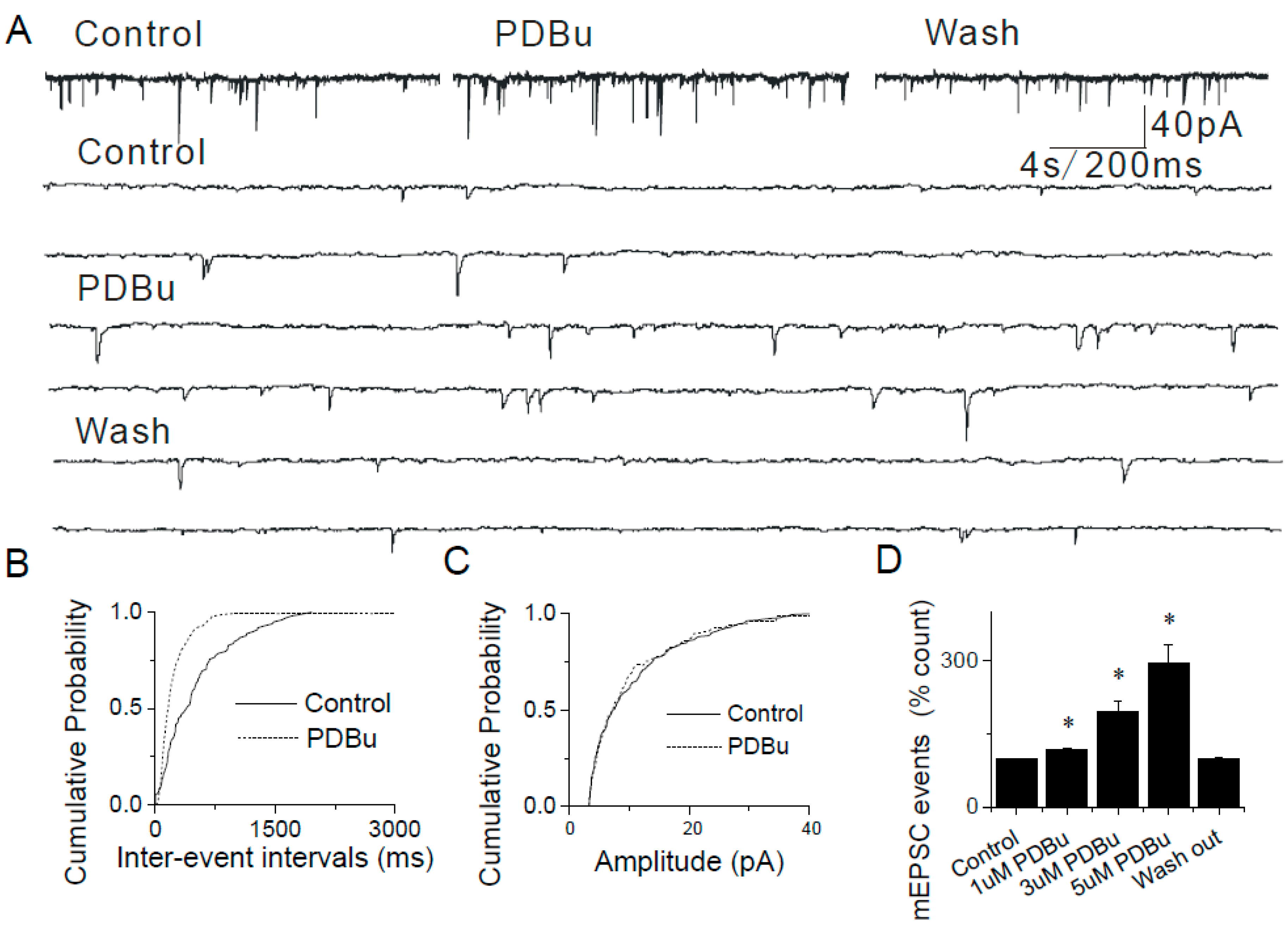
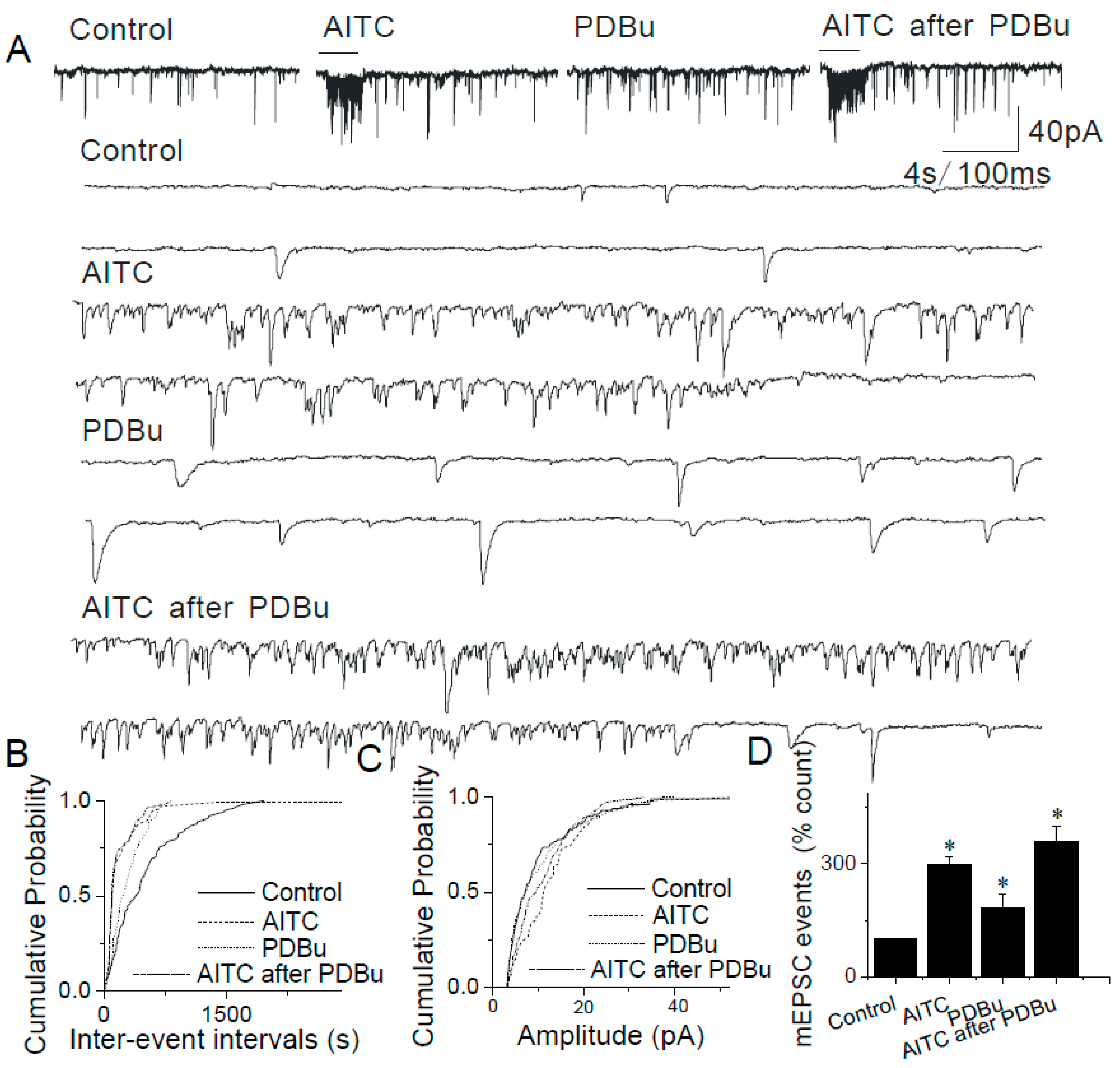
2.4. The Lack of Sensitization of Presynaptic TRPA1 by PDBu in the Caudal NTS
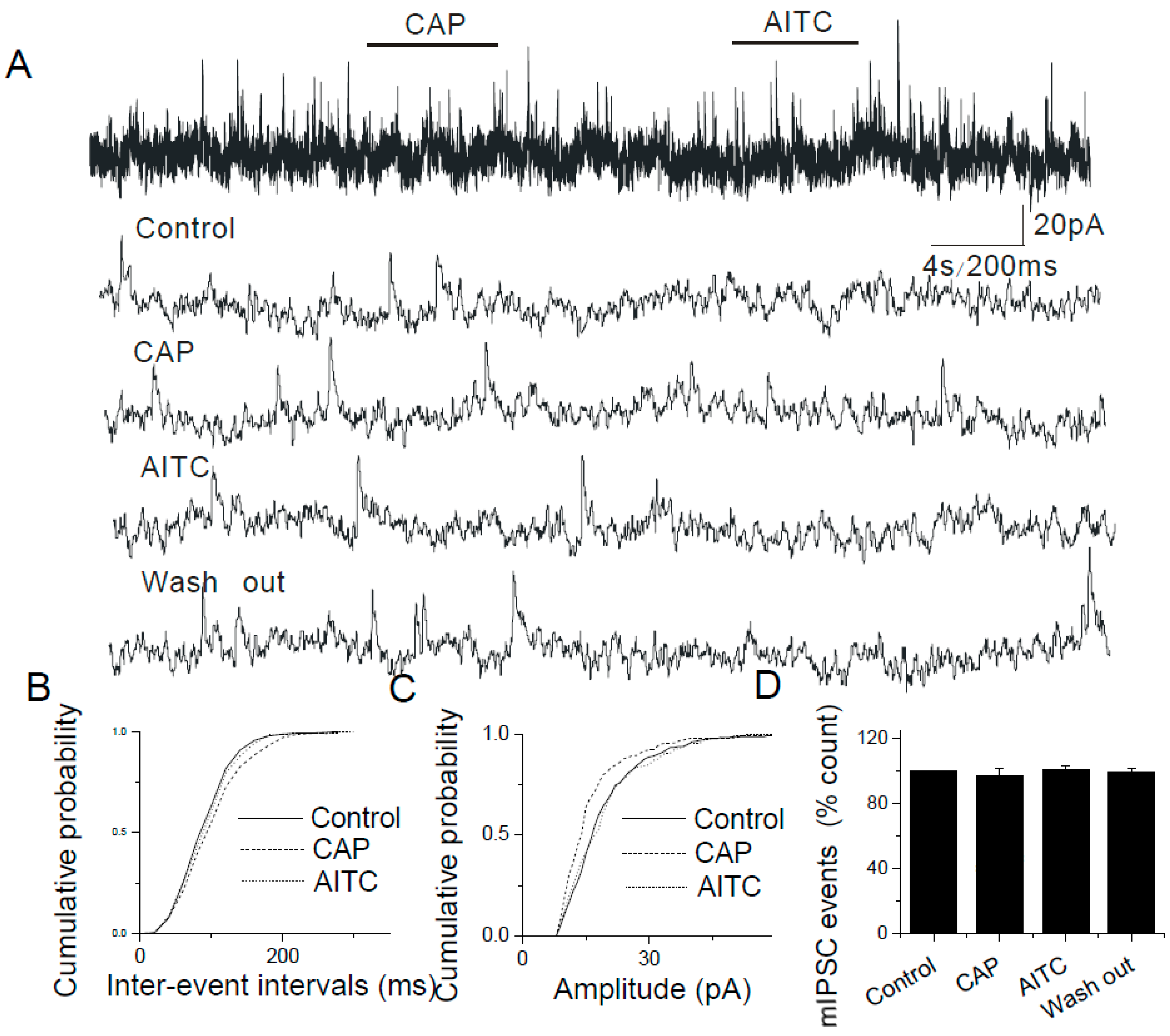
2.5. The Lack of Modulatory Effects of AITC on Inhibitory Synapses in the Caudal NTS
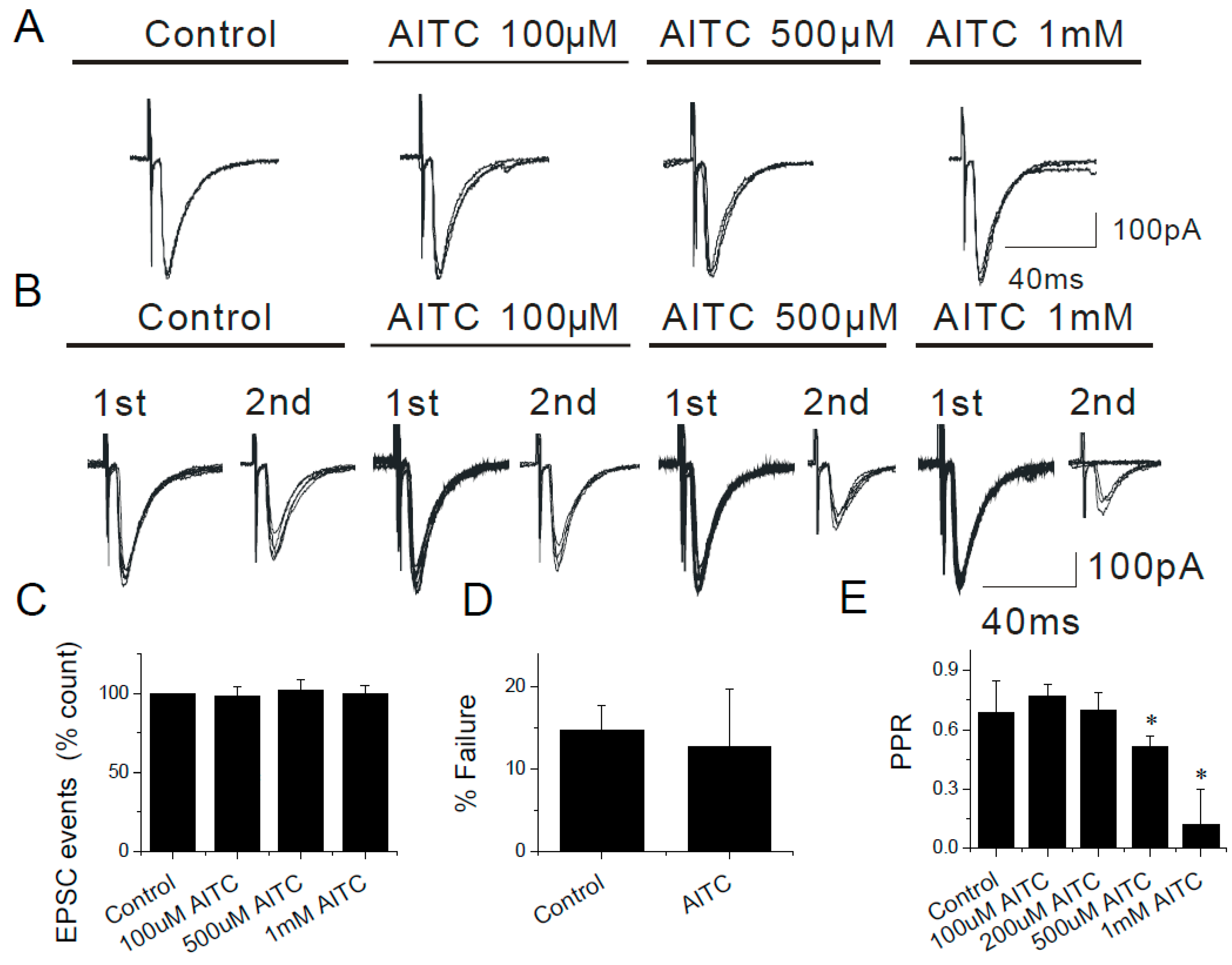
2.6. Modulation of Evoked EPSCs by AITC in the Caudal NTS
2.7. Data from TRPA1 Knockout Mice Support the Modulatory Role of TRPA1 in the Caudal NTS
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Preparation of Brainstem Slices
4.3. Electrophysiological Patch-Clamp Recordings
4.4. Immunohistochemistry and Peptide Absorption Studies
4.5. Total RNA Extraction and RT-PCR
4.6. Data Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Ionescu, D.A.; Lugoji, G.; Radula, D. The nucleus of the solitary tract: A review of its anatomy and functions, with emphasis on its role in a putative central-control of brain-capillaries permeability. Neurol. Psychiatr. 1986, 24, 69–85. [Google Scholar]
- McAllen, R.M.; Spyer, K.M. The location of cardiac vagal preganglionic motoneurones in the medulla of the cat. J. Physiol. 1976, 258, 187–204. [Google Scholar] [CrossRef]
- Miura, M.; Reis, D.J. Termination and secondary projections of carotid sinus nerve in the cat brain stem. Am. J. Physiol. 1969, 217, 142–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckstead, R.M.; Norgren, R. An autoradiographic examination of the central distribution of the trigeminal, facial, glossopharyngeal, and vagal nerves in the monkey. J. Comp. Neurol. 1979, 184, 455–472. [Google Scholar] [CrossRef] [PubMed]
- Gamboa-Esteves, F.O.; Tavares, I.; Almeida, A.; Batten, T.F.; McWilliam, P.N.; Lima, D. Projection sites of superficial and deep spinal dorsal horn cells in the nucleus tractus solitarii of the rat. Brain Res. 2001, 921, 195–205. [Google Scholar] [CrossRef]
- Hamilton, R.B.; Norgren, R. Central projections of gustatory nerves in the rat. J. Comp. Neurol. 1984, 222, 560–577. [Google Scholar] [CrossRef]
- Kalia, M.; Sullivan, J.M. Brainstem projections of sensory and motor components of the vagus nerve in the rat. J. Comp. Neurol. 1982, 211, 248–265. [Google Scholar] [CrossRef]
- Menetrey, D.; Basbaum, A.I. Spinal and trigeminal projections to the nucleus of the solitary tract: A possible substrate for somatovisceral and viscerovisceral reflex activation. J. Comp. Neurol. 1987, 255, 439–450. [Google Scholar] [CrossRef]
- Travers, S.P.; Pfaffmann, C.; Norgren, R. Convergence of lingual and palatal gustatory neural activity in the nucleus of the solitary tract. Brain Res. 1986, 365, 305–320. [Google Scholar] [CrossRef]
- Boscan, P.; Pickering, A.E.; Paton, J.F. The nucleus of the solitary tract: An integrating station for nociceptive and cardiorespiratory afferents. Exp. Physiol. 2002, 87, 259–266. [Google Scholar] [CrossRef]
- Feng, L.; Uteshev, V.V. Projection target-specific action of nicotine in the caudal nucleus of the solitary tract. J. Neurosci. Res. 2014, 92, 1560–1572. [Google Scholar] [CrossRef]
- Ricardo, J.A.; Koh, E.T. Anatomical evidence of direct projections from the nucleus of the solitary tract to the hypothalamus, amygdala, and other forebrain structures in the rat. Brain Res. 1978, 153, 1–26. [Google Scholar] [CrossRef]
- Spyer, K.M.; Donoghue, S.; Felder, R.B.; Jordan, D. Processing of afferent inputs in cardiovascular control. Clin. Exp. Hypertens. A 1984, 6, 173–184. [Google Scholar] [CrossRef]
- Wiertelak, E.P.; Roemer, B.; Maier, S.F.; Watkins, L.R. Comparison of the effects of nucleus tractus solitarius and ventral medial medulla lesions on illness-induced and subcutaneous formalin-induced hyperalgesias. Brain Res. 1997, 748, 143–150. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Andresen, M.C.; Hofmann, M.E.; Fawley, J.A. The unsilent majority-TRPV1 drives “spontaneous” transmission of unmyelinated primary afferents within cardiorespiratory NTS. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R1207–R1216. [Google Scholar] [CrossRef]
- Bailey, T.W.; Jin, Y.H.; Doyle, M.W.; Andresen, M.C. Vanilloid-sensitive afferents activate neurons with prominent A-type potassium currents in nucleus tractus solitarius. J. Neurosci. 2002, 22, 8230–8237. [Google Scholar] [CrossRef]
- Doyle, M.W.; Bailey, T.W.; Jin, Y.H.; Andresen, M.C. Vanilloid receptors presynaptically modulate cranial visceral afferent synaptic transmission in nucleus tractus solitarius. J. Neurosci. 2002, 22, 8222–8229. [Google Scholar] [CrossRef]
- Sun, B.; Bang, S.I.; Jin, Y.H. Transient receptor potential A1 increase glutamate release on brain stem neurons. Neuroreport 2009, 20, 1002–1006. [Google Scholar] [CrossRef]
- Smith, D.V.; Uteshev, V.V. Heterogeneity of nicotinic acetylcholine receptor expression in the caudal nucleus of the solitary tract. Neuropharmacology 2008, 54, 445–453. [Google Scholar] [CrossRef]
- Cattaruzza, F.; Spreadbury, I.; Miranda-Morales, M.; Grady, E.F.; Vanner, S.; Bunnett, N.W. Transient receptor potential ankyrin-1 has a major role in mediating visceral pain in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G81–G91. [Google Scholar] [CrossRef]
- Kimball, E.S.; Prouty, S.P.; Pavlick, K.P.; Wallace, N.H.; Schneider, C.R.; Hornby, P.J. Stimulation of neuronal receptors, neuropeptides and cytokines during experimental oil of mustard colitis. Neurogastroenterol. Motil. 2007, 19, 390–400. [Google Scholar] [CrossRef]
- Vermeulen, W.; De Man, J.G.; De Schepper, H.U.; Bult, H.; Moreels, T.G.; Pelckmans, P.A.; De Winter, B.Y. Role of TRPV1 and TRPA1 in visceral hypersensitivity to colorectal distension during experimental colitis in rats. Eur. J. Pharmacol. 2013, 698, 404–412. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Zuo, X.; Zhen, Y.; Yu, Y.; Gao, L. Transient receptor potential ankyrin-1 participates in visceral hyperalgesia following experimental colitis. Neurosci. Lett. 2008, 440, 237–241. [Google Scholar] [CrossRef]
- Engel, M.A.; Leffler, A.; Niedermirtl, F.; Babes, A.; Zimmermann, K.; Filipović, M.R.; Izydorczyk, I.; Eberhardt, M.; Kichko, T.I.; Mueller-Tribbensee, S.M.; et al. TRPA1 and substance P mediate colitis in mice. Gastroenterology 2011, 141, 1346–1358. [Google Scholar] [CrossRef]
- Kondo, T.; Sakurai, J.; Miwa, H.; Noguchi, K. Activation of p38 MAPK through transient receptor potential A1 in a rat model of gastric distension-induced visceral pain. Neuroreport 2013, 24, 68–72. [Google Scholar] [CrossRef]
- Laird, J.M.; Martinez-Caro, L.; Garcia-Nicas, E.; Cervero, F. A new model of visceral pain and referred hyperalgesia in the mouse. Pain 2001, 92, 335–342. [Google Scholar] [CrossRef]
- Kwan, K.Y.; Glazer, J.M.; Corey, D.P.; Rice, F.L.; Stucky, C.L. TRPA1 modulates mechanotransduction in cutaneous sensory neurons. J. Neurosci. 2009, 29, 4808–4819. [Google Scholar] [CrossRef] [Green Version]
- Sotomayor, M.; Corey, D.P.; Schulten, K. In search of the hair-cell gating spring elastic properties of ankyrin and cadherin repeats. Structure 2005, 13, 669–682. [Google Scholar] [CrossRef]
- Story, G.M.; Peier, A.M.; Reeve, A.J.; Eid, S.R.; Mosbacher, J.; Hricik, T.R.; Earley, T.J.; Hergarden, A.C.; Andersson, D.A.; Hwang, S.W.; et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef]
- Jordt, S.E.; Bautista, D.M.; Chuang, H.H.; McKemy, D.D.; Zygmunt, P.M.; Högestätt, E.D.; Meng, I.D.; Julius, D. Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature 2004, 427, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Zurborg, S.; Yurgionas, B.; Jira, J.A.; Caspani, O.; Heppenstall, P.A. Direct activation of the ion channel TRPA1 by Ca2+. Nat. Neurosci. 2007, 10, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef]
- Hinman, A.; Chuang, H.H.; Bautista, D.M.; Julius, D. TRP channel activation by reversible covalent modification. Proc. Natl. Acad. Sci. USA 2006, 103, 19564–19568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista, D.M.; Pellegrino, M.; Tsunozaki, M. TRPA1: A gatekeeper for inflammation. Annu. Rev. Physiol. 2013, 75, 181–200. [Google Scholar] [CrossRef]
- Hondoh, A.; Ishida, Y.; Ugawa, S.; Ueda, T.; Shibata, Y. Distinct expression of cold receptors (TRPM8 and TRPA1) in the rat nodose-petrosal ganglion complex. Brain Res. 2010, 1319, 60–69. [Google Scholar] [CrossRef]
- Katsura, H.; Tsuzuki, K.; Noguchi, K.; Sakagami, M. Differential expression of capsaicin-, menthol-, and mustard oil-sensitive receptors in naive rat geniculate ganglion neurons. Chem. Senses 2006, 31, 681–688. [Google Scholar] [CrossRef]
- Cavanaugh, D.J.; Chesler, A.T.; Jackson, A.C.; Sigal, Y.M.; Yamanaka, H.; Grant, R.; O’Donnell, D.; Nicoll, R.A.; Shah, N.M.; Julius, D.; et al. Trpv1 reporter mice reveal highly restricted brain distribution and functional expression in arteriolar smooth muscle cells. J. Neurosci. 2011, 31, 5067–5177. [Google Scholar] [CrossRef]
- Mishra, S.K.; Tisel, S.M.; Orestes, P.; Bhangoo, S.K.; Hoon, M.A. TRPV1-lineage neurons are required for thermal sensation. EMBO J. 2011, 30, 582–593. [Google Scholar] [CrossRef]
- Macpherson, L.J.; Dubin, A.E.; Evans, M.J.; Marr, F.; Schultz, P.G.; Cravatt, B.F.; Patapoutian, A. Noxious compounds activate TRPA1 ion channels through covalent modification of cysteines. Nature 2007, 445, 541–545. [Google Scholar] [CrossRef]
- Raisinghani, M.; Zhong, L.; Jeffry, J.A.; Bishnoi, M.; Pabbidi, R.M.; Pimentel, F.; Cao, D.S.; Evans, M.S.; Premkumar, L.S. Activation characteristics of transient receptor potential ankyrin 1 and its role in nociception. Am. J. Physiol. Cell Physiol. 2011, 301, C587–C600. [Google Scholar] [CrossRef] [Green Version]
- Brose, N.; Rosenmund, C. Move over protein kinase C, you’ve got company: Alternative cellular effectors of diacylglycerol and phorbol esters. J. Cell Sci. 2002, 115, 4399–4411. [Google Scholar] [CrossRef]
- Cao, D.S.; Yu, S.Q.; Premkumar, L.S. Modulation of transient receptor potential Vanilloid 4-mediated membrane currents and synaptic transmission by protein kinase C. Mol. Pain 2009, 5, 5. [Google Scholar] [CrossRef]
- Lou, X.; Korogod, N.; Brose, N.; Schneggenburger, R. Phorbol esters modulate spontaneous and Ca2+-evoked transmitter release via acting on both Munc13 and protein kinase C. J. Neurosci. 2008, 28, 8257–8267. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.H.; McDougall, S.J.; Fawley, J.A.; Smith, S.M.; Andresen, M.C. Primary afferent activation of thermosensitive TRPV1 triggers asynchronous glutamate release at central neurons. Neuron 2010, 65, 657–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanotto, K.L.; Iodi Carstens, M.; Carstens, E. Cross-desensitization of responses of rat trigeminal subnucleus caudalis neurons to cinnamaldehyde and menthol. Neurosci. Lett. 2008, 430, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Son, J.Y.; Kim, T.H.; Paik, S.K.; Dai, Y.; Noguchi, K.; Ahn, D.K.; Bae, Y.C. Expression of transient receptor potential ankyrin 1 (TRPA1) in the rat trigeminal sensory afferents and spinal dorsal horn. J. Comp. Neurol. 2010, 518, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Fukuoka, T.; Obata, K.; Yamanaka, H.; Dai, Y.; Tokunaga, A.; Noguchi, K. Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with adelta/c-fibers and colocalization with trk receptors. J. Comp. Neurol. 2005, 493, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, M.; Nakatsuka, T.; Fujita, T.; Kuroda, Y.; Kumamoto, E. Activation of TRPA1 channel facilitates excitatory synaptic transmission in substantia gelatinosa neurons of the adult rat spinal cord. J. Neurosci. 2007, 27, 4443–4451. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, T.; Ohbuchi, T.; Saito, T.; Sudo, Y.; Fujihara, H.; Minami, K.; Nagatomo, T.; Uezono, Y.; Ueta, Y. Allyl isothiocyanates and cinnamaldehyde potentiate miniature excitatory postsynaptic inputs in the supraoptic nucleus in rats. Eur. J. Pharmacol. 2011, 655, 31–37. [Google Scholar] [CrossRef]
- Sikand, P.; Premkumar, L.S. Potentiation of glutamatergic synaptic transmission by protein kinase C-mediated sensitization of TRPV1 at the first sensory synapse. J. Physiol. 2007, 581, 631–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffry, J.A.; Yu, S.Q.; Sikand, P.; Parihar, A.; Evans, M.S.; Premkumar, L.S. Selective targeting of TRPV1 expressing sensory nerve terminals in the spinal cord for long lasting analgesia. PLoS ONE 2009, 4, e7021. [Google Scholar] [CrossRef]
- Grace, M.S.; Belvisi, M.G. TRPA1 receptors in cough. Pulm. Pharmacol. Ther. 2011, 24, 286–288. [Google Scholar] [CrossRef]
- Ramsey, I.S.; Delling, M.; Clapham, D.E. An introduction to TRP channels. Annu. Rev. Physiol. 2006, 68, 619–647. [Google Scholar] [CrossRef]
- Sadofsky, L.R.; Boa, A.N.; Maher, S.A.; Birrell, M.A.; Belvisi, M.G.; Morice, A.H. TRPA1 is activated by direct addition of cysteine residues to the N-hydroxysuccinyl esters of acrylic and cinnamic acids. Pharmacol. Res. 2011, 63, 30–36. [Google Scholar] [CrossRef]
- Andersson, D.A.; Gentry, C.; Moss, S.; Bevan, S. Transient receptor potential A1 is a sensory receptor for multiple products of oxidative stress. J. Neurosci. 2008, 28, 2485–2494. [Google Scholar] [CrossRef]
- Bessac, B.F.; Sivula, M.; von Hehn, C.A.; Escalera, J.; Cohn, L.; Jordt, S.E. TRPA1 is a major oxidant sensor in murine airway sensory neurons. J. Clin. Investig. 2008, 118, 1899–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obata, K.; Katsura, H.; Mizushima, T.; Yamanaka, H.; Kobayashi, K.; Dai, Y.; Fukuoka, T.; Tokunaga, A.; Tominaga, M.; Noguchi, K. TRPA1 induced in sensory neurons contributes to cold hyperalgesia after inflammation and nerve injury. J. Clin. Investig. 2005, 115, 2393–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwan, K.Y.; Allchorne, A.J.; Vollrath, M.A.; Christensen, A.P.; Zhang, D.S.; Woolf, C.J.; Corey, D.P. TRPA1 contributes to cold, mechanical, and chemical nociception but is not essential for hair-cell transduction. Neuron 2006, 50, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Kerstein, P.C.; del Camino, D.; Moran, M.M.; Stucky, C.L. Pharmacological blockade of TRPA1 inhibits mechanical firing in nociceptors. Mol. Pain 2009, 5, 19. [Google Scholar] [CrossRef]
- Bautista, D.M.; Jordt, S.E.; Nikai, T.; Tsuruda, P.R.; Read, A.J.; Poblete, J.; Yamoah, E.N.; Basbaum, A.I.; Julius, D. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell 2006, 124, 1269–1282. [Google Scholar] [CrossRef]
- Christianson, J.A.; Bielefeldt, K.; Altier, C.; Cenac, N.; Davis, B.M.; Gebhart, G.F.; High, K.W.; Kollarik, M.; Randich, A.; Undem, B.; et al. Development, plasticity and modulation of visceral afferents. Brain Res. Rev. 2009, 60, 171–186. [Google Scholar] [CrossRef]
- Blackshaw, L.A.; Brierley, S.M.; Hughes, P.A. TRP channels: New targets for visceral pain. Gut 2010, 59, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.P.; Pelayo, J.C.; Cattaruzza, F.; Kuo, Y.M.; Gai, G.; Chiu, J.V.; Bron, R.; Furness, J.B.; Grady, E.F.; Bunnett, N.W. Transient receptor potential ankyrin 1 is expressed by inhibitory motoneurons of the mouse intestine. Gastroenterology 2011, 141, 565–575.e4. [Google Scholar] [CrossRef] [PubMed]
- Premkumar, L.S.; Abooj, M. TRP channels and analgesia. Life Sci. 2013, 92, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Moran, M.M.; McAlexander, M.A.; Biro, T.; Szallasi, A. Transient receptor potential channels as therapeutic targets. Nat. Rev. Drug Discov. 2011, 10, 601–620. [Google Scholar] [PubMed]
- Hori, Y.; Endo, K.; Takahashi, T. Long-lasting synaptic facilitation induced by serotonin in superficial dorsal horn neurones of the rat spinal cord. J. Physiol. 1996, 492, 867–876. [Google Scholar] [CrossRef]
- Malenka, R.C.; Madison, D.V.; Nicoll, R.A. Potentiation of synaptic transmission in the hippocampus by phorbol esters. Nature 1986, 321, 175–177. [Google Scholar] [CrossRef]
- Premkumar, L.S.; Ahern, G.P. Induction of vanilloid receptor channel activity by protein kinase C. Nature 2000, 408, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Kalappa, B.I.; Feng, L.; Kem, W.R.; Gusev, A.G.; Uteshev, V.V. Mechanisms of facilitation of synaptic glutamate release by nicotinic agonists in the nucleus of the solitary tract. Am. J. Physiol. Cell Physiol. 2011, 301, C347–C361. [Google Scholar] [CrossRef] [PubMed] [Green Version]










© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, L.; Uteshev, V.V.; Premkumar, L.S. Expression and Function of Transient Receptor Potential Ankyrin 1 Ion Channels in the Caudal Nucleus of the Solitary Tract. Int. J. Mol. Sci. 2019, 20, 2065. https://doi.org/10.3390/ijms20092065
Feng L, Uteshev VV, Premkumar LS. Expression and Function of Transient Receptor Potential Ankyrin 1 Ion Channels in the Caudal Nucleus of the Solitary Tract. International Journal of Molecular Sciences. 2019; 20(9):2065. https://doi.org/10.3390/ijms20092065
Chicago/Turabian StyleFeng, Lin, Victor V. Uteshev, and Louis S. Premkumar. 2019. "Expression and Function of Transient Receptor Potential Ankyrin 1 Ion Channels in the Caudal Nucleus of the Solitary Tract" International Journal of Molecular Sciences 20, no. 9: 2065. https://doi.org/10.3390/ijms20092065
APA StyleFeng, L., Uteshev, V. V., & Premkumar, L. S. (2019). Expression and Function of Transient Receptor Potential Ankyrin 1 Ion Channels in the Caudal Nucleus of the Solitary Tract. International Journal of Molecular Sciences, 20(9), 2065. https://doi.org/10.3390/ijms20092065