The Human Isoform of RNA Polymerase II Subunit hRPB11bα Specifically Interacts with Transcription Factor ATF4

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Yeast Two-Hybrid Screening
2.2. hRPB11bα Specifically Interact with ATF4 In Vitro
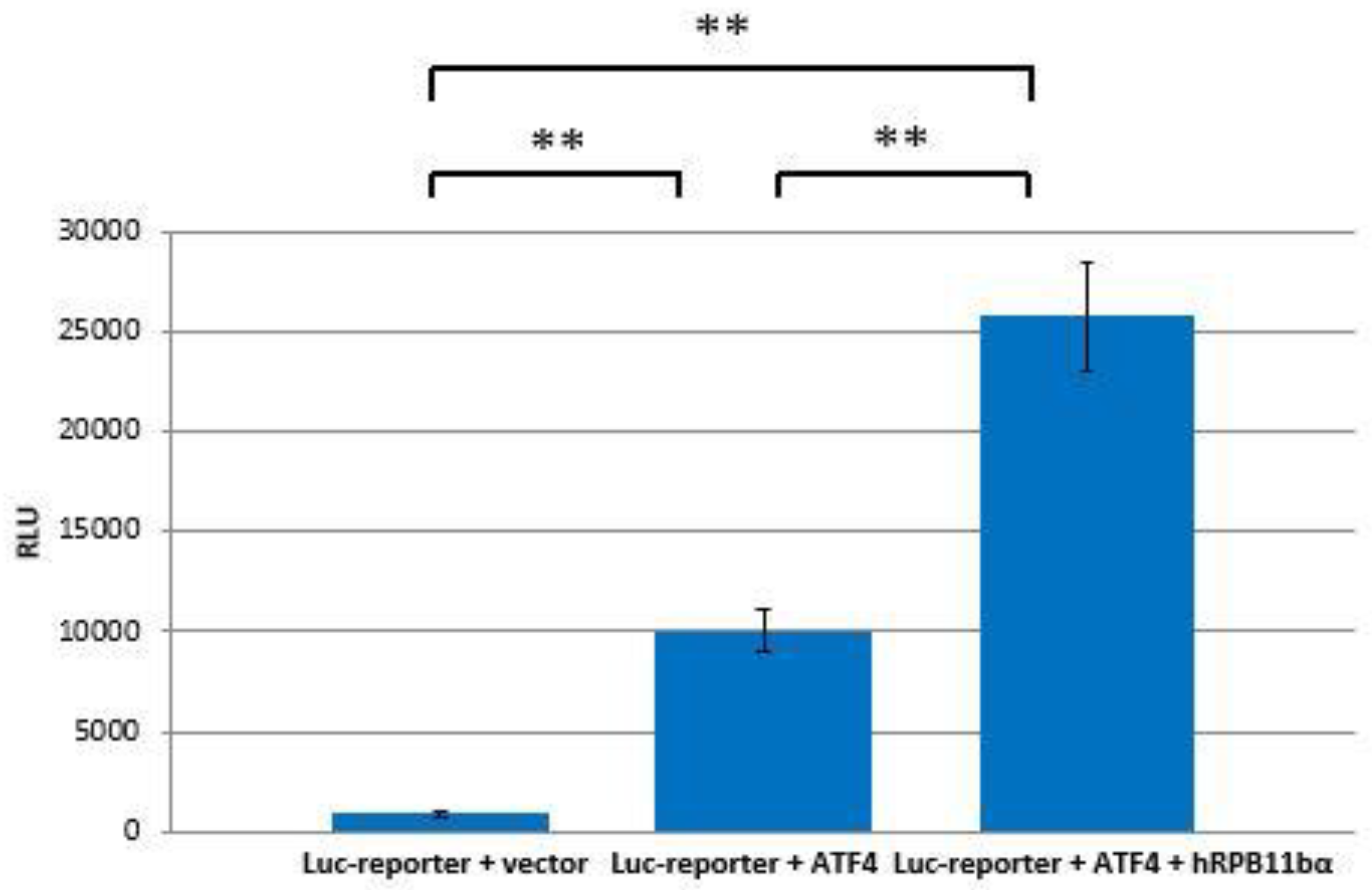
2.3. RPB11bα Enhances ATF4-Mediated Transactivation
3. Discussion
4. Materials and Methods
4.1. Yeast Two-Hybrid Screen
4.2. Liquid ß-Galactosidase Assay
4.3. Proteins Coexpression and Purification
4.4. Cell Culture and Transfections
4.5. Plasmids
4.6. Luciferase Assay
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ATF/CREB | activating transcription factor/cAMP response element binding protein |
| b-Zip | basic region-leucine zipper domain in ATF/CREB family of proteins |
| EST | expressed sequence tag |
| hEIF3 | 13-subunits factor initiation of transcription #3 of Homo sapiens (human eukaryotic initiation factor 3) |
| RLU | relative light units |
| RNAPII | RNA polymerase II |
| YTH | yeast two-hybrid system |
References
- Cramer, P.; Bushnell, D.A.; Fu, J.; Gnatt, A.L.; Maier-Davis, B.; Thompson, N.E.; Burgess, R.R.; Edwards, A.M.; David, P.R.; Kornberg, R.D. Architecture of RNA polymerase II and implications for the transcription mechanism. Science 2000, 288, 640–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bushnell, D.A.; Kornberg, R.D. Complete, 12-subunit RNA polymerase II at 4.1-A resolution: Implications for the initiation of transcription. Proc. Natl. Acad. Sci. USA 2003, 100, 6969–6973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armache, K.J.; Kettenberger, H.; Cramer, P. Architecture of initiation-competent 12-subunit RNA polymerase II. Proc. Natl. Acad. Sci. USA 2003, 100, 6964–6968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brueckner, F.; Armache, K.J.; Cheung, A.; Damsma, G.E.; Kettenberger, H.; Lehmann, E.; Sydow, J.; Cramer, P. Structure-function studies of the RNA polymerase II elongation complex. Acta Crystallogr. D Biol. Crystallogr. 2009, 65, 112–120. [Google Scholar] [CrossRef]
- He, Y.; Fang, J.; Taatjes, D.J.; Nogales, E. Structural visualization of key steps in human transcription initiation. Nature 2013, 495, 481–486. [Google Scholar] [CrossRef] [Green Version]
- Bernecky, C.; Herzog, F.; Baumeister, W.; Plitzko, J.M.; Cramer, P. Structure of transcribing mammalian RNA polymerase II. Nature 2016, 529, 551–555. [Google Scholar] [CrossRef]
- Plaschka, C.; Hantsche, M.; Dienemann, C.; Burzinski, C.; Plitzko, J.; Cramer, P. Transcription initiation complex structures elucidate DNA opening. Nature 2016, 533, 353–358. [Google Scholar] [CrossRef]
- He, Y.; Yan, C.; Fang, J.; Inouye, C.; Tjian, R.; Ivanov, I.; Nogales, E. Nearatomic resolution visualization of human transcription promoter opening. Nature 2016, 533, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Robinson, P.J.; Trnka, M.J.; Bushnell, D.A.; Davis, R.E.; Mattei, P.J.; Burlingame, A.L.; Kornberg, R.D. Structure of a Complete Mediator-RNA Polymerase II Pre-Initiation Complex. Cell 2016, 166, 1411–1422. [Google Scholar] [CrossRef] [Green Version]
- Vos, S.M.; Farnung, L.; Urlaub, H.; Cramer, P. Structure of paused transcription complex Pol II-DSIF-NELF. Nature 2018, 560, 601–606. [Google Scholar] [CrossRef]
- Vos, S.M.; Farnung, L.; Boehning, M.; Wigge, C.; Linden, A.; Urlaub, H.; Cramer, P. Structure of activated transcription complex Pol II-DSIF-PAF-SPT6. Nature 2018, 560, 607–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farnung, L.; Vos, S.M.; Cramer, P. Structure of transcribing RNA polymerase II-nucleosome complex. Nat. Commun. 2018, 9, 5432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shpakovski, G.V.; Acker, J.; Wintzerith, M.; Lacroix, J.F.; Thuriaux, P.; Vigneron, M. Four subunits that are shared by the three classes of RNA polymerase are functionally interchangeable between Homo sapiens and Saccharomyces cerevisiae. Mol. Cell. Biol. 1995, 15, 4702–4710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKune, K.; Moore, P.A.; Hull, M.W.; Woychik, N.A. Six human RNA polymerase subunits functionally substitute for their yeast counterparts. Mol. Cell. Biol. 1995, 15, 6895–6900. [Google Scholar] [CrossRef] [Green Version]
- Schaller, S.; Grandemange, S.; Shpakovski, G.V.; Golemis, E.A.; Kedinger, C.; Vigneron, M. Interactions between the full complement of human RNA polymerase II subunits. FEBS Lett. 1999, 461, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M.; Ishiguro, A.; Ishihama, A. RNA polymerase II subunits 2; 3; and 11 form a core subassembly with DNA binding activity. J. Biol. Chem. 1997, 272, 25851–25855. [Google Scholar] [CrossRef] [Green Version]
- Tan, Q.; Linask, K.L.; Ebright, R.H.; Woychik, A. Activation mutants in yeast RNA polymerase II subunit RPB3 provide evidence for a structurally conserved surface required for activation in eukaryotes and bacteria. Genes Dev. 2000, 14, 339–348. [Google Scholar]
- Eyboulet, F.; Wydau-Dematteis, S.; Eychenne, T.; Alibert, O.; Neil, H.; Boschiero, C.; Nevers, M.C.; Volland, H.; Cornu, D.; Redeker, V.; et al. Mediator independently orchestrates multiple steps of preinitiation complex assembly in vivo. Nucleic Acids Res. 2015, 43, 9214–9231. [Google Scholar] [CrossRef]
- Proshkina, G.M.; Shpakovskiĭ, G.V. The functional interaction of an RNA polymerase II Rpb11 subunit with the Med18 subunit (Srb5) of the Saccharomyces cerevisiae Mediator complex. Bioorg. Khim. 2009, 35, 572–576. [Google Scholar] [CrossRef]
- Grandemange, S.; Schalle, S.; Yamano, S.; Du Manoir, S.; Shpakovski, G.V.; Mattei, M.G.; Kedinger, C.; Vigneron, M. A human RNA polymerase II subunit is encoded by a recently generated multigene family. BMC Mol. Biol. 2001, 2, 14. [Google Scholar] [CrossRef]
- Shpakovskiĭ, D.G.; Shematorova, E.K.; Shpakovskiĭ, G.V. New genes on human chromosome 7: Bioinformatic analysis of a gene cluster from the POLR2J family. Bioorg. Khim. 2004, 30, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Durrin, L.K.; Krontiris, T.G. The thymocyte-specific MAR binding protein, SATB1, interacts in vitro with a novel variant of DNA-directed RNA polymerase II, subunit 11. Genomics 2002, 79, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Benga, W.J.; Grandemange, S.; Shpakovski, G.V.; Shematorova, E.K.; Kedinger, C.; Vigneron, M. Distinct regions of RPB11 are required for heterodimerization with RPB3 in human and yeast RNA polymerase II. Nucleic. Acids. Res. 2005, 33, 3582–3590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shematorova, E.K.; Shpakovskiĭ, D.G.; Shpakovskiĭ, G.V. PMS2 and POLR2J gene families as molecular markers of the higher primate evolution. Genetika 2010, 46, 1254–1257. [Google Scholar] [CrossRef] [PubMed]
- Proshkin, S.A.; Shematorova, E.K.; Souslova, E.A.; Proshkina, G.M.; Shpakovski, G.V. A minor isoform of the human RNA polymerase II subunit hRPB11 (POLR2J) interacts with several components of the translation initiation factor eIF3. Biochemistry 2011, 76, 976–980. [Google Scholar] [CrossRef] [PubMed]
- Shematorova, E.K.; Shpakovskiĭ, D.G.; Shpakovskiĭ, G.V. Novel complexes of gene expression and their role in the appearance and evolution of the genus Homo. Tsitologiia 2013, 55, 172–177. [Google Scholar] [CrossRef]
- Liang, G.; Hai, T. Characterization of human activating transcription factor 4; a transcriptional activator that interacts with multiple domains of cAMP-responsive element-binding protein (CREB)-binding protein. J. Biol. Chem. 1997, 272, 24088–24095. [Google Scholar] [CrossRef] [Green Version]
- Rutkowski, D.T.; Kaufman, R.J. All roads lead to ATF4. Dev. Cell 2003, 4, 442–444. [Google Scholar] [CrossRef] [Green Version]
- Pakos-Zebrucka, K.; Koryga, I.; Mnich, K.; Ljujic, M.; Samali, A.; Gorman, A.M. The integrated stress response. EMBO Rep. 2016, 17, 1374–1395. [Google Scholar] [CrossRef] [Green Version]
- Golemis, E.A.; Serebriiskii, I.; Finley, R.L., Jr.; Kolonin, M.G.; Gyuris, J.; Brent, R. Interaction Trap/Two Hybrid System to Identify Interacting Proteins. In Current Protocols in Protein Science; Wiley Press: New York, NY, USA, 2009; pp. 1921–19235. [Google Scholar]
- Reynolds, A.; Lundblad, V.; Dorris, D.; Keaveney, M. Current Protocols in Molecular Biology; John Wiley & Sons Press: New York, NY, USA, 1997; pp. 1361–1366. [Google Scholar]
- Ebright, R.H.; Busby, S. The E. coli RNA polymerase alpha subunit: Structure and Function. Curr. Opin. Genet. Dev. 1995, 5, 197–203. [Google Scholar] [CrossRef]
- De Angelis, R.; Iezzi, S.; Bruno, T.; Corbi, N.; Di Padova, M.; Floridi, A.; Fanciulli, M.; Passananti, C. Functional interaction of the subunit 3 of RNA polymerase II (RPB3) with transcription factor-4 (ATF4). FEBS Lett. 2003, 547, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Corbi, N.; Di Padova, M.; De Angelis, R.; Bruno, T.; Libri, V.; Iezzi, S.; Floridi, A.; Fanciulli, M.; Passananti, C. The alpha-like RNA polymerase II core subunit 3 (RPB3) is involved in tissue-specific transcription and muscle differentiation via interaction with the myogenic factor myogenin. FASEB J. 2002, 12, 1639–1641. [Google Scholar] [CrossRef] [PubMed]
- Fanciulli, M.; Bruno, T.; Di Padova, M.; De Angelis, R.; Iezzi, S.; Iacobini, C.; Floridi, A.; Passananti, C. Identification of a novel partner of RNA polymerase II subunit 11; Che-1; which interacts with and affects the growth suppression function of Rb. FASEB J. 2000, 7, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Thiaville, M.M.; Dudenhausen, E.E.; Awad, K.S.; Gjymishka, A.; Zhong, C.; Kilberg, M.S. Activated transcription via mammalian amino acid response elements does not require enhanced recruitment of the Mediator complex. Nucleic Acids Res. 2008, 36, 5571–5580. [Google Scholar] [CrossRef]
- Asturias, F.J.; Jiang, Y.W.; Myers, L.C.; Gustafsson, C.M.; Kornberg, R.D. Conserved structures of Mediator and RNA polymerase II holoenzyme. Science 1999, 283, 985–987. [Google Scholar] [CrossRef]
- Plaschka, C.; Larivière, L.; Wenzeck, L.; Seizl, M.; Hemann, M.; Tegunov, D.; Petrotchenko, E.V.; Borchers, C.H.; Baumeister, W.; Herzog, F.; et al. Architecture of the RNA polymerase II-Mediator core initiation complex. Nature 2015, 518, 376–380. [Google Scholar] [CrossRef]
- Robinson, P.J.; Trnka, M.J.; Pellarin, R.; Greenberg, C.H.; Bushnell, D.A.; Davis, R.; Burlingame, A.L.; Sali, A.; Kornberg, R.D. Molecular architecture of the yeast Mediator complex. eLife 2015. [Google Scholar] [CrossRef]
- Sato, S.; Tomomori-Sato, C.; Tsai, K.-L.; Yu, X.; Sardiu, M.; Saraf, A.; Washburn, M.P.; Florens, L.; Asturias, F.J.; Conaway, R.C.; et al. Role for the MED21-MED7 Hinge in Assembly of the Mediator-RNA Polymerase II Holoenzyme. J. Biol. Chem. 2016, 291, 26886–26898. [Google Scholar] [CrossRef] [Green Version]
- Kudo, K.; Xi, Y.; Wang, Y.; Song, B.; Chu, E.; Ju, J.; Russo, J.J.; Ju, J. Translational control analysis by translationally active RNA capture/microarray analysis (TrIP–Chip). Nucleic Acids Res. 2010, 38, e104. [Google Scholar] [CrossRef] [Green Version]
- Watson, M.A.; Buckholz, R.; Weiner, M.P. Vectors encoding alternative antibiotic resistance for use in the yeast two-hybrid system. Biotechniques 1996, 21, 255–259. [Google Scholar] [CrossRef]
- Serebriiskii, I.G.; Toby, G.G.; Finley, R.L., Jr.; Golemis, E.A. Genomic analysis utilizing the yeast two-hybrid system. Methods Mol. Biol. 2001, 175, 415–454. [Google Scholar] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; CSHL Press: Cold Spring Harbor, NY, USA, 1989; Volume 1–3. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Proshkin, S.A.; Shematorova, E.K.; Shpakovski, G.V. The Human Isoform of RNA Polymerase II Subunit hRPB11bα Specifically Interacts with Transcription Factor ATF4. Int. J. Mol. Sci. 2020, 21, 135. https://doi.org/10.3390/ijms21010135
Proshkin SA, Shematorova EK, Shpakovski GV. The Human Isoform of RNA Polymerase II Subunit hRPB11bα Specifically Interacts with Transcription Factor ATF4. International Journal of Molecular Sciences. 2020; 21(1):135. https://doi.org/10.3390/ijms21010135
Chicago/Turabian StyleProshkin, Sergey A., Elena K. Shematorova, and George V. Shpakovski. 2020. "The Human Isoform of RNA Polymerase II Subunit hRPB11bα Specifically Interacts with Transcription Factor ATF4" International Journal of Molecular Sciences 21, no. 1: 135. https://doi.org/10.3390/ijms21010135
APA StyleProshkin, S. A., Shematorova, E. K., & Shpakovski, G. V. (2020). The Human Isoform of RNA Polymerase II Subunit hRPB11bα Specifically Interacts with Transcription Factor ATF4. International Journal of Molecular Sciences, 21(1), 135. https://doi.org/10.3390/ijms21010135