The Possible Role of Non-Structural Carbohydrates in the Regulation of Tree Hydraulics

 , , ,
, , , 
Abstract
:1. Introduction
2. Plant Hydraulics and Drought Stress
3. Stem NSCs Dynamics and the Maintenance of Hydraulic Function under Drought
4. Stem NSCs and the Post-Drought Recovery of Xylem Function
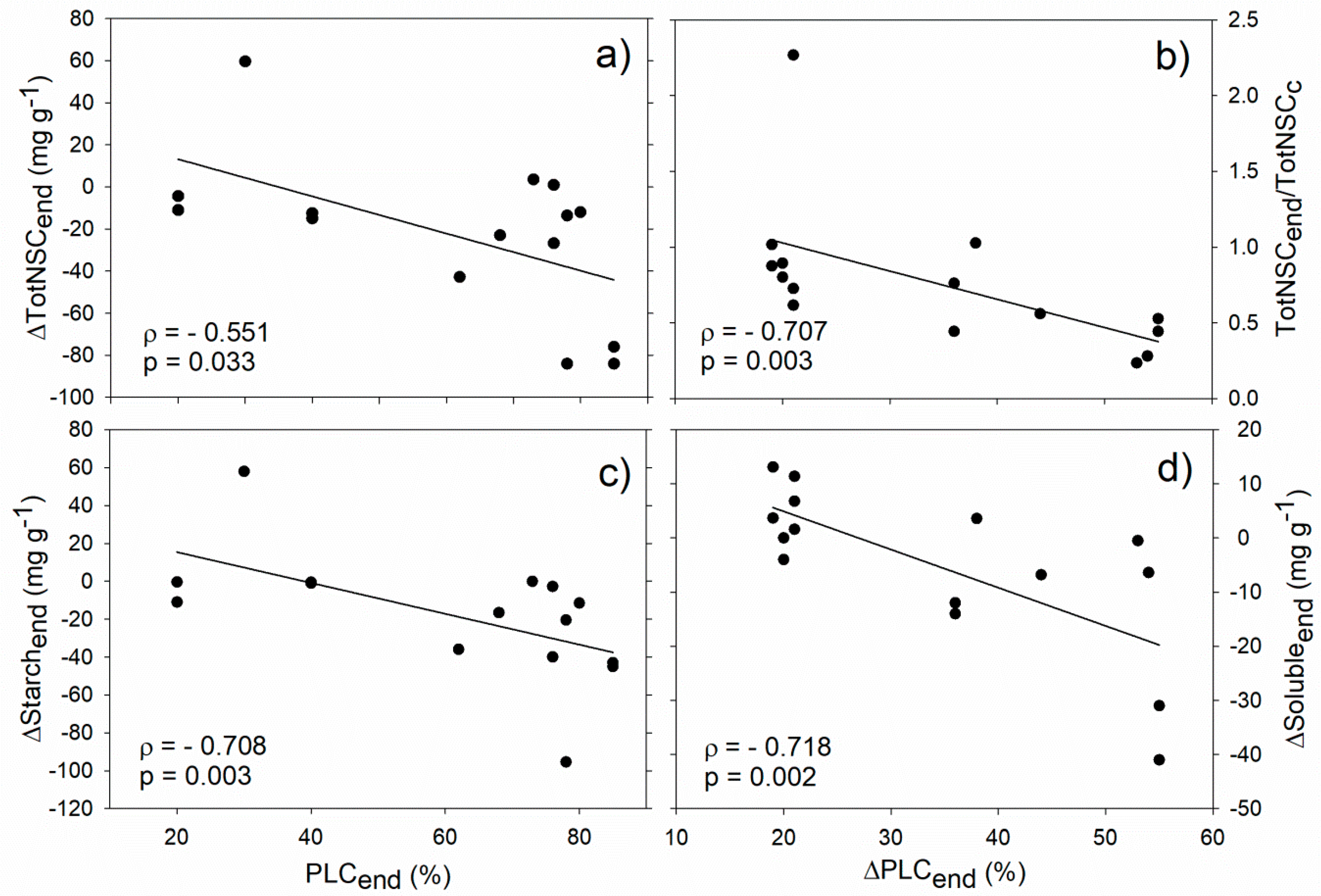
5. NSC-PLC Relationships: A Survey from Currently Available Data
6. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Conflicts of Interest
Abbreviations
| PLC | Percentage loss of hydraulic conductance |
| NSC | Non-Structural Carbohydrates |
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Solubleend | Starchend | TotNSCend | ΔSolubleend | ΔStarchend | ΔTotNSCend | Solubleend/Solublec | Starchend/Starchc | TotNSCend/TotNSCc | |
|---|---|---|---|---|---|---|---|---|---|
| ΔPLCend | −0.273 | −0.196 | −0.324 | −0.718 ** | −0.288 | 0.417 | −0.592 * | 0.270 | −0.707 ** |
| PLCend | 0.167 | 0.216 | 0.088 | 0 | −0.708 ** | −0.551 * | −0.048 | −0.142 | −0.473 (*) |
| PLCrec_% | / | / | / | −0.555 * | 0.409 | 0.254 | −0.306 | −0.348 | 0.086 |
| Solubleend | Starchend | TotNSCend | ΔSolublerec | ΔStarchrec | ΔTotNSCrec | Solublerec | Starchrec | Totrec | Solublerec/Solubleend | Starchrec/Starchend | TotNSCrec/TotNSCend | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ΔPLCrec | 0.027 | 0.067 | 0.160 | −0.287 | −0.172 | 0 | −0.067 | −0.009 | 0.297 | −0.201 | 0.160 | 0.276 |
| PLCrec_% | 0.054 | −0.284 (*) | −0.062 | −0.397 * | −0.131 | −0.065 | −0.170 | −0.324 (*) | −0.026 | −0.181 | 0.249 | −0.034 |
References
- Raven, J.A. Selection pressures on stomatal evolution. New Phytol. 2002, 153, 371–386. [Google Scholar] [CrossRef] [Green Version]
- Dai, A. Drought under global warming: A review. Wiley Interdiscip. Rev. Clim. Chang. 2011, 2, 45–65. [Google Scholar] [CrossRef] [Green Version]
- Adams, H.D.; Zeppel, M.J.B.; Anderegg, W.R.L.; Hartmann, H.; Landhausser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Field, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland, L.; Da Costa, A.C.L.; Galbraith, D.R.; Oliveira, R.S.; Binks, O.J.; Oliveira, A.A.R.; Pullen, A.M.; Doughty, C.E.; Metcalfe, D.B.; Vasconcelos, S.S.; et al. Death from drought in tropical forests in triggered by hydraulics not carbon starvation. Nature 2015, 528, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, H.; Trumbore, S. Understanding the roles of non-structural carbohydrates in forest trees—From what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef] [Green Version]
- Deslauriers, A.; Beaulieu, M.; Balducci, L.; Giovannelli, A.; Gagnon, M.J.; Rossi, S. Impact of warming and drought on carbon balance related to wood formation in black spruce. Ann. Bot. 2014, 114, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Tomasella, M.; Häberle, K.-H.; Nardini, A.; Hesse, B.; Machlet, A.; Matyssek, R. Post-drought hydraulic recovery is accompanied by non-structural carbohydrate depletion in the stem wood of Norway spruce saplings. Sci. Rep. 2017, 7, 14308. [Google Scholar] [CrossRef]
- Hoch, G.; Richter, A.; Körner, C. Non-structural carbon compounds in temperate forest trees. Plant Cell Environ. 2003, 26, 1067–1081. [Google Scholar] [CrossRef]
- Lloret, F.; Sapes, G.; Rosas, T.; Galieno, L.; Saura-Mas, S.; Sala, A.; Martínez-Vilalta, J. Non-structural carbohydrate dynamics associated with drought-induced die-off in woody species of a shrubland community. Ann. Bot. 2018, 121, 1383–1396. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.G.; Beerling, D.J.; Breshears, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Sevanto, S. Phloem transport and drought. J. Exp. Bot. 2014, 65, 1751–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevanto, S.; McDowell, N.G.; Dickman, I.T.; Pangle, R.; Pockman, W.T. How do trees die? A test of the hydraulic failure and carbon starvation hypotheses. Plant Cell Environ. 2014, 37, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Zeppel, M.J.B.; Anderegg, W.R.L.; Bloemen, J.; De Kauwe, M.G.; Hudson, P.; Ruehr, N.K.; Powell, T.L.; Von Arx, G.; Nardini, A. Xylem embolism refilling and resilience against drought-induced mortality in woody plants: Processes and trade-offs. Ecol. Res. 2018, 33, 839–855. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Bowman, D.J.M.S.; Nichols, S.; Delzon, S.; Burlett, R. Xylem function and growth rate interact to determine recovery rates after exposure to extreme water deficit. New Phytol. 2010, 188, 533–542. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef] [Green Version]
- Huntingford, C.; Atkin, O.K.; Martinez-de la Torre, A.; Mercado, L.M.; Heskel, M.A.; Harper, A.B.; Bloomfield, K.J.; O’Sullivan, O.S.; Reich, P.B.; Wythers, K.R.; et al. Implications of improved representations of plant respiration in a changing climate. Nat. Commun. 2017, 8, 1602. [Google Scholar] [CrossRef]
- Tomasella, M.; Nardini, A.; Hesse, B.D.; Machlet, A.; Matyssek, R.; Häberle, K.-H. Close to the edge: Effects of repeated severe drought on stem hydraulics and non-structural carbohydrates in European beech saplings. Tree Physiol. 2019, 39, 717–728. [Google Scholar] [CrossRef]
- Secchi, F.; Zwieniecki, M.A. Sensing embolism in xylem vessels: The role of sucrose as a trigger for refilling. Plant Cell Environ. 2011, 34, 514–524. [Google Scholar] [CrossRef]
- Nardini, A.; Savi, T.; Trifilò, P.; Lo Gullo, M.A. Drought stress and the recovery from xylem embolism in woody plants. In Progress in Botany; Cánovas, F.M., Luettge, U., Matyssek, R., Eds.; Springer: Berlin, Germany, 2018; Volume 79, pp. 137–231. [Google Scholar]
- McDowell, N.G.; Nardini, A.; Brodribb, T.J. Hydraulics in the 21st century. New Phytol. 2019, 224, 537–542. [Google Scholar] [CrossRef] [Green Version]
- Tyree, M.T.; Zimmermann, M.H. Xylem Structure and the Ascent of Sap, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Dixon, H.H.; Joly, J. On the ascent of sap. Ann. Bot. 1894, 8, 468–470. [Google Scholar] [CrossRef]
- Tyree, M.T. Plant hydraulics: The ascent of water. Nature 2003, 423, 923. [Google Scholar] [CrossRef] [PubMed]
- Hacke, U.G.; Sperry, J.S.; Pockman, W.T.; Davis, S.D.; McCulloh, K.A. Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 2001, 126, 457–461. [Google Scholar] [CrossRef]
- Choat, B.; Cobb, A.R.; Jansen, S. Structure and function of bordered pits: New discoveries and impacts on whole-plant hydraulic function. New Phytol. 2008, 177, 608–626. [Google Scholar] [CrossRef] [PubMed]
- Tyree, M.T.; Sperry, J.S. Vulnerability of xylem to cavitation and embolism. Annu. Rev. Plant Physiol. 1989, 40, 19–38. [Google Scholar] [CrossRef]
- Vargaftik, N.B.; Volkov, B.N.; Voljak, L.D. International tables of the surface tension of water. J. Phys. Chem. Ref. Data 1983, 12, 817–820. [Google Scholar] [CrossRef] [Green Version]
- Mayr, S.; Améglio, T. Freezing stress in tree xylem. Prog. Bot. 2016, 77, 381–414. [Google Scholar]
- Sperry, J.S.; Hacke, U.G.; Pittermann, J. Size and function in conifer tracheids and angiosperm vessels. Am. J. Bot. 2006, 93, 1490–1500. [Google Scholar] [CrossRef] [Green Version]
- Sperry, J.S.; Hacke, U.G.; Feild, T.S.; Sano, Y.; Sikkema, E.H. Hydraulic consequences of vessel evolution in angiosperms. Int. J. Plant Sci. 2007, 168, 1127–1139. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.; Rahman, M.; Bräuning, A. Long-term wood anatomical time series of two ecologically contrasting tropical tree species reveal differential hydraulic adjustment to climatic stress. Agric. For. Meteorol. 2019, 265, 412–423. [Google Scholar] [CrossRef]
- Hillabrand, R.M.; Hacke, U.G.; Lieffers, V.J. Drought-induced xylem pit membrane damage in aspen and balsam poplar. Plant Cell Environ. 2016, 39, 2210–2220. [Google Scholar] [CrossRef] [PubMed]
- Nardini, A.; Salleo, S.; Jansen, S. More than just a vulnerable pipeline: Xylem physiology in the light of ion-mediated regulation of plant water transport. J. Exp. Bot. 2011, 63, 4701–4718. [Google Scholar] [CrossRef] [PubMed]
- Spicer, R. Symplasmic networks in secondary vascular tissues: Parenchyma distribution and activity supporting long-distance transport. J. Exp. Bot. 2014, 65, 1829–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kedrov, G.B. Functioning wood. Wulfenia 2002, 19, 57–95. [Google Scholar]
- Höll, W. Distribution, fluctuation and metabolism of food reserves in the wood of trees. In Cell and Molecular Biology of Wood Formation; Savidge, R., Barnett, J., Napier, R., Eds.; BIOS Scientific Publishers: Oxford, MS, USA, 2000; pp. 347–362. [Google Scholar]
- Plavcová, L.; Hoch, G.; Morris, H.; Ghiasi, S.; Jansen, S. The amount of parenchyma and living fibers affects storage of nonstructural carbohydrates in young stems and roots of temperate trees. Am. J. Bot. 2016, 103, 603–612. [Google Scholar] [CrossRef] [Green Version]
- Sauter, J.J.; Iten, W.I.; Zimmermann, M.H. Studies on the release of sugar into the vessels of the sugar maple (Acer saccharum). Can. J. Bot. 1973, 51, 1–8. [Google Scholar] [CrossRef]
- Brodersen, C.R.; McElrone, A.J.; Choat, B.; Matthews, M.A.; Shackel, K.A. The dynamics of embolism repair in xylem: In vivo visualizations using high-resolution computed tomography. Plant Physiol. 2010, 154, 1088–1095. [Google Scholar] [CrossRef] [Green Version]
- Salleo, S.; Lo Gullo, M.A.; Trifilò, P.; Nardini, A. New evidence for a role of vessel-associated cells and phloem in the rapid xylem refilling of cavitated stems of Laurus nobilis L. Plant Cell Environ. 2004, 27, 1065–1076. [Google Scholar] [CrossRef]
- Secchi, F.; Pagliarani, C.; Zwieniecki, M.A. The functional role of xylem parenchyma cells and aquaporins during recovery from severe water stress. Plant Cell Environ. 2017, 40, 858–871. [Google Scholar] [CrossRef]
- Meinzer, F.C.; James, S.A.; Goldstein, G.; Woodruff, D. Whole-tree water transport scales with sapwood capacitance on tropical forest canopy trees. Plant Cell Environ. 2017, 26, 1147–1155. [Google Scholar] [CrossRef] [Green Version]
- Sakr, S.; Alves, G.; Morillon, R.; Maurel, K.; Decourteix, M.; Guilliot, A.; Fleurat-Lessard, P.; Julien, J.-L.; Chrispeels, M.J. Plasma membrane aquaporins are involved in winter embolism recovery in walnut tree. Plant Physiol. 2003, 133, 630–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rioux, D.; Nicole, M.; Simard, M.; Ouellette, G.B. Immunocytochemical evidence that secretion of pectin occurs during gel (gum) and tylosis formation in trees. Phytopathology 1998, 88, 494–505. [Google Scholar] [CrossRef] [PubMed]
- De Boer, A.H.; Volkov, V. Logistics of water and salt transport through the plant: Structure and functioning of the xylem. Plant Cell Environ. 2003, 265, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Morris, H.; Plavcova, L.; Goral, M.; Klepsch, M.M.; Kotowska, M.; Schenk, H.J.; Jansen, S. Vessel-associated cells in angiosperm xylem: Highly specialized living cells at the symplast-apoplast boundary. Am. J. Bot. 2018, 105, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Améglio, T.; Devourteix, M.; Alves, G.; Valentin, V.; Sakr, S.; Julien, J.L.; Pétel, G.; Guilliot, A.; Lacointe, A. Temperature effects on xylem sap osmolarity in walnut trees: Evidence for a vitalistic model of winter embolism repair. Tree Physiol. 2004, 24, 785–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.-Y.; Han, S.-J.; Zhang, J.-H.; Wang, M.; Yin, X.-H.; Fang, L.-D.; Yang, D.; Hao, G.-Y. The interaction between nonstructural carbohydrate reserves and xylem hydraulics in Korean pine trees across an altitudinal gradient. Tree Physiol. 2018, 38, 1792–1804. [Google Scholar] [CrossRef]
- Mayr, S.; Schmid, P.; Laur, J.; Rosner, S.; Charra-Vaskou, K.; Damon, B.; Hacke, U.G. Uptake of water via branches helps timberline conifers refill embolized xylem in late winter. Plant Physiol. 2014, 164, 1731–1740. [Google Scholar] [CrossRef] [Green Version]
- Lintunen, A.; Mayr, S.; Salmon, Y.; Cochard, H.; Höltta, T. Drivers of apoplastic freezing in gymnosperm and angiosperm branches. Ecol. Evol. 2018, 8, 333–343. [Google Scholar] [CrossRef]
- Chen, T.H.; Murata, N. Enhancement of tolerance of abiotic stresses by metabolic engineering of betaines and other compatible solutes. Curr. Opin. Plant Biol. 2002, 5, 250–257. [Google Scholar] [CrossRef]
- O’Brien, M.J.; Leuzinger, S.; Philipson, C.D.; Hector, T.J. Drought survival of tropical tree seedlings enhanced by non-structural carbohydrate levels. Nat. Clim. Chang. 2014, 4, 710–714. [Google Scholar] [CrossRef]
- Sala, A.; Woodruff, D.R.; Meinzer, F.C. Carbon dynamics in tree: Feast or famine? Tree Phyisol. 2012, 32, 764–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittmann, C.; Pfanz, H. Antitranspirant functions of stem periderms and their influence on corticular photosynthesis under drought stress. Trees 2008, 22, 187–196. [Google Scholar] [CrossRef]
- Vandegehuchte, M.M.; Bloemen, J.; Vergeynst, L.L.; Steppe, K. Woody tissue photosynthesis in trees: Salve on the wounds of drought? New Phytol. 2015, 208, 998–1002. [Google Scholar] [CrossRef] [PubMed]
- Eyles, A.; Pinkard, E.A.; O’Grady, A.P.; Worledge, D.; Warren, C.R. Role of corticular photosynthesis following defoliation in Eucaliptus globulus. Plant Cell Environ. 2009, 32, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, N.; Egerton, J.J.G.; Lovelock, C.E.; Ball, M.C. Light-dependent maintenance of hydraulic function in mangrove branches: Do xylary chloroplasts play a role in embolism repair? New Phytol. 2012, 195, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Bloemen, J.; Vergeynst, L.L.; Overlaet-Michelis, L.; Steppe, K. How important is woody tissue photosynthesis in poplar during drought stress? Trees 2016, 30, 63–72. [Google Scholar] [CrossRef]
- De Baerdemaeker, N.J.F.; Salomon, R.L.; De Roo, L.; Steppe, K. Sugars from woody tissue photosynthesis reduce vulnerability to cavitation. New Phytol. 2017, 216, 720–727. [Google Scholar] [CrossRef] [Green Version]
- Schenk, H.J.; Steppe, K.; Jansen, S. Nanobubbles: A new paradigm for air-seeding in xylem. Trends Plant Sci. 2015, 20, 199–205. [Google Scholar] [CrossRef]
- Schenk, H.J.; Espino, S.; Romo, D.M.; Nima, N.; Do, A.Y.T.; Michaud, J.M.; Papahadjopoulos-Sternberg, B.; Yang, J.; Zuo, Y.Y.; Steppe, K.; et al. Xylem surfactants introduce a new element to the cohesion-tension theory. Plant Physiol. 2017, 173, 1177–1196. [Google Scholar] [CrossRef] [Green Version]
- Secchi, F.; Zwieniecki, M.A. Down-regulation of plasma intrinsic protein1 aquaporin in poplar trees is detrimental to recovery from embolism. Plant Physiol. 2014, 164, 1789–1799. [Google Scholar] [CrossRef] [Green Version]
- Pagliarani, C.; Casolo, V.; Beiragi, M.A.; Cavalletto, S.; Siciliano, I.; Schubert, A.; Gullino, M.L.; Zwieniecki, M.A.; Secchi, F. Priming xylem for stress recovery depends on coordinated activity of sugar metabolic pathways and changes in xylem sap pH. Plant Cell Environ. 2019, 42, 1775–1787. [Google Scholar] [CrossRef] [Green Version]
- Nardini, A.; Lo Gullo, M.A.; Salleo, S. Refilling of embolized conduits: Is it a matter of phloem unloading? Plant Sci. 2011, 180, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Bucci, S.J.; Scholz, F.G.; Goldstein, G.; Meinzer, F.C.; Sternberg, L.D.L. Dynamic changes in hydraulic conductivity in petioles of two savanna tree species: Factors and mechanisms contributing to the refilling of embolized vessels. Plant Cell Environ. 2003, 26, 1633–1645. [Google Scholar] [CrossRef]
- Salleo, S.; Trifilò, P.; Esposito, S.; Nardini, A.; Lo Gullo, M.A. Starch-to-sugar conversion in wood parenchyma of field-growing laurus nobilis plants: A component if the signal pathway for embolims repair? Funct. Plant Biol. 2009, 36, 815–825. [Google Scholar] [CrossRef]
- Secchi, F.; Zwieniecki, M.A. Analysis of xylem sap from functional (nonembolized) and non-functional (embolized) vessels of Populus nigra: Chemistry of refilling. Plant Physiol. 2012, 160, 955–964. [Google Scholar] [CrossRef] [Green Version]
- Sharp, R.G.; Davies, W.J. Variability among species in the apoplastic pH signalling response to drying soils. J. Exp. Bot. 2009, 60, 4363–4370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Losso, A.; Beikircher, B.; Dämon, B.; Kikuta, S.; Schmid, P.; Mayr, S. Xylem sap surface tension may be crucial for hydraulic safety. Plant Physiol. 2017, 175, 1135–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Secchi, F.; Zwieniecki, M.A. Accumulation of sugars in the xylem apoplast observed under water stress conditions is controlled by xylem pH. Plant Cell Environ. 2016, 39, 2350–2360. [Google Scholar] [CrossRef]
- Secchi, F.; Gilbert, M.E.; Zwieniecki, M.A. Transcriptome response to embolism formation in stems of Populus trichocarpa provides insight into signaling and the biology of refilling. Plant Physiol. 2011, 157, 1419–1429. [Google Scholar] [CrossRef] [Green Version]
- Perrone, I.; Pagliarani, C.; Lovisolo, C.; Chitarra, W.; Roman, F.; Schubert, A. Recovery from water stress affects grape leaf petiole transcriptome. Planta 2012, 235, 1383–1396. [Google Scholar] [CrossRef]
- Secchi, F.; Zwieniecki, M.A. Patterns in PIP gene expression in Populus trichocarpa during recovery from xylem embolism suggest a major role for the PIP1 aquaporin subfamily as moderators of refilling process. Plant Cell Environ. 2010, 33, 1285–1297. [Google Scholar] [CrossRef] [PubMed]
- Chitarra, W.; Balestrini, R.; Vitali, M.; Pagliarani, C.; Perrone, I.; Schubert, A. Gene expression in vessel-associated cells upon xylem embolism repair in Vitis vinifera L. Planta 2014, 239, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Kempa, S.; Krasensky, J.; Dal Santo, S.; Kopka, J.; Jonak, C. A central role of abscisic acid in stress-regulated carbohydrate metabolism. PLoS ONE 2008, 3, e3935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunetti, C.; Gori, A.; Marino, G.; Latini, P.; Sobolev, A.P.; Nardini, A.; Haworth, M.; Giovannelli, A.; Capitani, D.; Loreto, F.; et al. Dynamic changes in ABA content in water-stressed Populus nigra: Effects of carbon fixation and soluble carbohydrates. Ann. Bot. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochard, H.; Lemoine, D.; Ameglio, T.; Granier, A. Mechanisms of xylem recovery from winter embolism in Fagus sylvatica. Tree Physiol. 2001, 21, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roach, M.; Arrivault, S.; Mahboubi, A.; Krohn, N.; Sulpice, R.; Stitt, M.; Niittylä, T. Spatially resolved metabolic analysis reveals a central role for transcriptional control in carbon allocation to wood. J. Exp. Bot. 2017, 68, 3529–3539. [Google Scholar] [CrossRef]
- Mahboubi, A.; Niittylä, T. Sucrose transport and carbon fluxes during wood formation. Physiol. Plant. 2018, 164, 67–81. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sauter, J.J. Xylem dysfunction during winter and recovery of hydraulic conductivity in diffuse-porous and ring-porous trees. Oecologia 1996, 105, 435–439. [Google Scholar] [CrossRef]
- Yang, S.J.; Zhang, Y.J.; Sun, M.; Goldstein, G.; Cao, K.F. Recovery of diurnal depression of leaf hydraulic conductance in a subtropical woody bamboo species: Embolism refilling by nocturnal root pressure. Tree Physiol. 2012, 32, 414–422. [Google Scholar] [CrossRef] [Green Version]
- Ewers, F.W.; Ameglio, F.W.; Cochard, H.; Beaujard, F.; Martignac, M.; Vandame, M.; Bodet, C.; Cruiziat, P. Seasonal variation in xylem pressure of walnut trees: Root and stem pressures. Tree Physiol. 2001, 21, 1123–1132. [Google Scholar] [CrossRef]
- Améglio, T.; Bodet, C.; Lacointe, A.; Cochard, H. Winter embolism, mechanisms of xylem hydraulic conductivity recovery and springtime growth patterns in walnut and peach trees. Tree Physiol. 2002, 22, 1211–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodersen, C.R.; McElrone, A.J. Maintenance of xylem network transport capacity: A review of embolism repair in vascular plants. Front. Plant Sci. 2013, 4, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, G.Y.; Wheeler, J.K.; Holbrook, N.M.; Goldstein, G. Investigating xylem embolism formation, refilling and water storage in tree trunks using frequency domain reflectometry. J. Exp. Bot. 2013, 64, 2321–2332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westhoff, M.; Schneider, H.; Zimmermann, D.; Mimietz, S.; Stinzing, A.; Wegner, L.; Kaiser, W.; Krohne, G.; Shirley, S.; Jakob, P.; et al. The mechanisms of refilling of xylem conduits and bleeding of tall birch during spring. Plant Biol. 2008, 10, 604–623. [Google Scholar] [CrossRef]
- Améglio, T.; Ewers, F.W.; Cochard, H.; Martignac, M.; Vandame, M.; Bodet, C.; Cruiziat, P. Winter stem pressures in walnut trees: Effects of carbohydrates, cooling and freezing. Tree Physiol. 2001, 21, 387–394. [Google Scholar] [CrossRef] [Green Version]
- Brodersen, C.R.; Knipfer, T.; McElrone, A.J. In vivo visualization of the final stages of xylem vessel refilling in grapevine (Vitis vinifera) stems. New Phytol. 2018, 217, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Knipfer, T.; Cuneo, I.F.; Brodersen, C.R.; McElrone, A.J. In situ visualization of the dynamics in xylem embolism formation and removal in the absence of root pressure: A study on excised grapevine stems. Plant Physiol. 2016, 171, 1024–1036. [Google Scholar] [CrossRef] [Green Version]
- Heizmann, U.; Kreuzwieser, J.; Schnitzler, J.P.N.; Bruggemann, N.; Rennenberg, H. Assimilate transport in the xylem sap of pedunculated oak (Quercus robur) saplings. Plant Biol. 2001, 3, 132–138. [Google Scholar] [CrossRef]
- Fromard, L.; Babin, V.; Fleurat-Lessard, P.; Fromont, J.C.; Serrano, R.; Bonnemain, J.L. Control of vascular sap pH by the vessel-associated cells in woody species. Plant Physiol. 1995, 108, 913–918. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, K.; Saiki, S.-T.; Yazaki, K.; Ogasa, M.Y.; Shirai, M.; Nakano, T.; Yoshimura, J.; Ishida, A. The dynamics of carbon stored in xylem sapwood to drought-induced hydraulic stress in mature trees. Sci. Rep. 2016, 6, 24513. [Google Scholar] [CrossRef] [Green Version]
- Losso, A.; Nardini, A.; Dämon, B.; Mayr, S. Xylem sap chemistry: Seasonal changes in timberline conifers Pinus cembra, Picea abies, Larix decidua. Biol. Plant. 2018, 62, 157–165. [Google Scholar] [CrossRef]
- Oroian, M.; Ropciuc, S.; Amariei, S.; Gutt, G. Correlations between density viscosity, surface tension and ultrasonic velocity of different mono- and di-saccharides. J. Mol. Liq. 2015, 207, 145–151. [Google Scholar] [CrossRef]
- Beattie, J.K.; Djerdjev, A.M.; Gray-Weale, A.; Kallay, N.; Lützenkirchen, J.; Preočanin, T.; Selmani, A. pH and the surface tension of water. J. Colloid Interface Sci. 2014, 422, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Christensen-Dalsgaard, K.K.; Tyree, M.T.; Mussone, P.G. Surface tension phenomena in the xylem sap of three diffuse porous temperate tree species. Tree Physiol. 2011, 31, 361–368. [Google Scholar] [CrossRef]
- Salleo, S.; Lo Gullo, M.A.; De Paoli, D.; Zippo, M. Xylem recovery from cavitation-induced embolism plants of Laurus nobilis: A possible mechanism. New Phytol. 1996, 132, 47–56. [Google Scholar] [CrossRef]
- Zwieniecki, M.A.; Hutyra, L.; Thompson, M.V.; Holbrook, N.M. Dynamic changes in petiole specific conductivity in red maple (Acer rubrum L.), tulip tree (Liriodendron tulipifera L.) and northern fox grape (Vitis labrusca). Plant Cell Environ. 2000, 23, 407–414. [Google Scholar] [CrossRef] [Green Version]
- Salleo, S.; Trifilo, P.; Lo Gullo, M.A. Phloem as a possible major determinant of rapid cavitation reversal in stems of Laurus nobilis (laurel). Funct. Plant Biol. 2006, 33, 1063–1074. [Google Scholar] [CrossRef]
- Liu, J.; Gu, L.; Yu, Y.; Huang, P.; Wu, Z.; Zhang, Q.; Qian, Y.; Wan, X.; Sun, Z. Corticular photosynthesis drives bark water uptake to refill embolized vessels in dehydrated branches of Salix matsudana. Plant Cell Environ. 2019, 42, 2584–2596. [Google Scholar] [CrossRef]
- Martorell, S.; Diaz-Espejo, A.; Medrano, H.; Ball, M.C.; Choat, B. Rapid hydraulic recovery of Eucalyptus pauciflora after drought: Link-ages between stem hydraulics and leaf gas exchange. Plant Cell Environ. 2014, 37, 617–626. [Google Scholar] [CrossRef]
- Trifilò, P.; Casolo, V.; Raimondo, F.; Petrussa, E.; Boscutti, F.; Lo Gullo, M.A.; Nardini, A. Effects of prolonged drought on stem non-structural carbohydrates content and post-drought hydraulic recovery in Laurus noblis L.: The possible link between carbon starvation and hydraulic failure. Plant Physiol. Biochem. 2017, 120, 232–241. [Google Scholar] [CrossRef]
- Wheeler, J.K.; Huggert, B.A.; Tofte, A.N.; Rockwell, F.E.; Holbrook, N.M. Cutting xylem under tension or supersaturated with gas can generate PLC and the appearance of rapid recovery from embolism. Plant Cell Environ. 2013, 36, 1938–1949. [Google Scholar] [CrossRef] [PubMed]
- Trifilò, P.; Raimondo, F.; Lo Gullo, M.A.; Barbera, P.M.; Salleo, S.; Nardini, A. Relax and refill: Xylem rehydration prior to hydraulic measurements favours embolism repair in stems and generates artificially low PLC values. Plant Cell Environ. 2014, 37, 2491–2499. [Google Scholar] [CrossRef] [PubMed]
- Torres-Ruiz, J.M.; Jansen, S.; Choat, B.; McElrone, A.J.; Cochard, H.; Brodribb, T.J.; Badel, E.; Burlett, R.; Bouche, P.S.; Brodersen, C.R.; et al. Direct X-Ray microtomography observations confirm the induction of embolism upon xylem cutting under tension. Plant Physiol. 2015, 167, 40–43. [Google Scholar] [CrossRef] [Green Version]
- Cochard, H.; Delzon, S. Hydraulic failure and repair are not routine in trees. Ann. For. Sci. 2013, 70, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Holbrook, N.M.; Ahrens, E.T.; Burns, M.J.; Zwieniecki, M.A. In vivo observation of cavitation and embolism repair using magnetic resonance imaging. Plant Physiol. 2001, 126, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knipfer, T.; Eustis, A.; Brodersen, C.; Walker, A.M.; McElrone, A.J. Grapevine species from varied native habitats exhibit differences ibn embolism formation/repair associated with leaf gas exchange and root pressure. Plant Cell Environ. 2015, 38, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Zwieniecki, M.A.; Melcher, P.J.; Ahrens, E.T. Analysis of spatial and temporal dynamics of xylem refilling in Acer rubrum L. using magnetic resonance imaging. Front. Plant Sci. 2013, 4, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clearwater, M.J.; Clark, C.J. In vivo magnetic resonance imaging of xylem vessel contents in woody lianas. Plant Cell Environ. 2003, 26, 1205–1214. [Google Scholar] [CrossRef] [Green Version]
- Charrier, G.; Torres-Ruiz, J.M.; Badel, E.; Burlett, R.; Choat, B.; Cochard, H.; Delmas, E.L.; Domec, J.-C.; Jansen, S.; King, A.; et al. Evidence for hydraulic vulnerability segmentation and lack of xylem refilling under tension. Plant Physiol. 2016, 172, 1657–1668. [Google Scholar] [CrossRef]
- Choat, B.; Brodersen, C.R.; McElrone, A.J. Synchrotron X-ray microtomography of xylem embolism in Sequoia sempervirens saplings during cycles of drought and recovery. New Phytol. 2015, 205, 1095–1105. [Google Scholar] [CrossRef]
- Petruzzellis, F.; Pagliarani, C.; Savi, T.; Losso, A.; Cavalletto, S.; Tromba, G.; Dullin, C.; Bär, A.; Ganthaler, A.; Miotto, A.; et al. The pitfalls of in vivo imaging techniques: Evidence for cellular damage caused by synchrotron X-ray computed micro-tomography. New Phytol. 2018, 220, 104–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laur, J.; Hacke, U. Exploring Picea glauca aquaporins in the context of needle water uptake and xylem refilling. New Phytol. 2014, 203, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Eller, C.B.; Lima, A.L.; Oliveira, R.S. Foliar uptake of fog water and transport belowground alleviates drought effects in the cloud forest tree species, Drimys brasiliensis (Winteraceae). New Phytol. 2003, 199, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Earles, J.M.; Sperling, O.; Silva, L.C.R.; Brodersen, C.R.; North, M.P.; Zwieniecki, M.A. Bark water uptake promotes localized hydraulic recovery in coastal redwood crown. Plant Cell Environ. 2016, 39, 320–328. [Google Scholar] [CrossRef]
- Yu, Y.C.; Liu, J.X.; Sun, Z.Y. Transcriptome profiling in Salix matsudana during refilling xylem vessels after embolism. Biol. Plant. 2019, 63, 425–431. [Google Scholar] [CrossRef]
- Tomasella, M.; Casolo, V.; Aichner, N.; Petruzzellis, F.; Savi, T.; Trifilò, P.; Nardini, A. Non-structural carbohydrate and hydraulic dynamics during drought and recovery in Fraxinus ornus and Ostrya carpinifolia saplings. Plant Physiol. Biochem. 2019, 145. [Google Scholar] [CrossRef]
- Trifilò, P.; Kiorapostolou, N.; Petruzzellis, F.; Vitti, S.; Petit, G.; Lo Gullo, M.; Nardini, A.; Casolo, V. Hydraulic recovery from xylem embolism in excised branches of twelve woody species: Relationships with parenchyma cells and non-structural carbohydrates. Plant Physiol. Biochem. 2019, 139, 513–520. [Google Scholar] [CrossRef]
- Savi, T.; Casolo, V.; Luglio, J.; Bertuzzi, S.; Trifilò, P.; Lo Gullo, M.A.; Nardini, A. Species-specific reversal of stem xylem embolism after a prolonged drought correlates to endpoint concentration of soluble sugars. Plant Physiol. Biochem. 2016, 106, 198–207. [Google Scholar] [CrossRef]
- Beikircher, B.; Mittmann, C.; Mayr, S. Prolonged soil frost affects hydraulics and phenology of apple trees. Front. Plant Sci. 2016, 7, 867. [Google Scholar] [CrossRef] [Green Version]
- Beikircher, B.; Mayr, S. Annual patterns of xylem embolism in high-yield apple cultivars. Funct. Plant Biol. 2017, 44, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Mayr, S.; Schmid, P.; Rosner, S. Winter embolism and recovery in the conifer schrub Pinus mugo L. Forests 2019, 10, 941. [Google Scholar] [CrossRef] [Green Version]
- Tixier, A.; Gambetta, G.A.; Orozco, J.; Zwieniecki, M.A. Non-structural carbohydrates in dormant woody perennials; the tale of winter survival and spring arrival. Front. For. Glob. Chang. 2019, 2, 18. [Google Scholar] [CrossRef] [Green Version]
- Landhausser, S.M.; Chow, P.S.; Dickman, L.T.; Furze, M.E.; Kuhlman, I.; Schmid, S.; Wiesbauer, J.; Wild, B.; Gleixner, G.; Hartmann, H.; et al. Standardized protocols and procedures can precisely and accurately quantify non-structural carbohydrates. Tree Physiol. 2018, 38, 1764–1778. [Google Scholar] [CrossRef] [PubMed]
- Tixier, A.; Orozco, J.; Roxas, A.A.; Earles, J.M.; Zwieniecki, M.A. Diurnal variation in nonstructural carbohydrate storage in trees: Remobilization and vertical mixing. Plant Physiol. 2018, 178, 1602–1613. [Google Scholar] [CrossRef] [Green Version]
- Earles, J.M.; Knipfer, T.; Tixier, A.; Orozco, J.; Reyes, C.; Zwieniecki, M.A.; Brodersen, C.R.; McElrone, A.J. In vivo quantification of plant starch reserves at micrometer resolution using X-ray microCT imaging and machine learning. New Phytol. 2018, 218, 1260–1269. [Google Scholar] [CrossRef] [Green Version]



| Species | Type of Sample | Embolism Induction | Rehydration Type | PLC before Recovery | NSC at Peak Embolism | NSC at Drought Relief | Recovery Duration | Hydraulic Recovery | Citation |
|---|---|---|---|---|---|---|---|---|---|
| Fagus sylvatica | Pot | SD | SR | 85% | St decrease, SS increase in W and B | St increase, SS decrease in W and B | One week | No | [18] |
| Fraxinus ornus | Pot | SD | SR | 76% | SS and Tot depleted only in B | Not changed | One day | Yes | [119] |
| 78% | St and Tot depleted in B and W | Not changed | No | ||||||
| Hibiscus glaber | Field | SD | SR | 70% | SS increase and St decrease | SS decrease and St increase | One week | Yes | [93] |
| Ligustrum micranthum | 40% | SS increase and St decrease | Not changed | One week | Yes | ||||
| Populus tremula x alba | Pot | SD | SR | 80% | SS increase in xylem sap St decrease, Glu increase in W | SS decrease in xylem sap Glu decrease in W | One week | Yes | [64] |
| Laurus nobilis | Pot | SD | SR | 23% | Not changed | Not changed | One day | Yes | [103] |
| 34% | Not changed | Not changed | One day | Yes | |||||
| One week | Yes | ||||||||
| Arbutus unedo | CB | BD | B/L WU | ~50% | N.A. | St and SS decrease | One hour | Yes | [120] |
| Ceratonia siliqua | St increase, SS decrease | Yes | |||||||
| Cercis siliquastrum | St and SS decrease | Yes | |||||||
| Eucalyptus camaldulensis | St increase, SS decrease | No | |||||||
| Laurus nobilis | St increase, SS decrease | Yes | |||||||
| Morus alba | St increase, SS decrease | Yes | |||||||
| Myrtus communis | St increase, SS decrease | Yes | |||||||
| Nerium oleander | St decrease, SS increase | No | |||||||
| Olea europea | St and SS decrease | Yes | |||||||
| Phillyrea latifolia | St and SS increase | No | |||||||
| Pistacia lentiscus | St and SS increase | No | |||||||
| Quercus ilex | St and SS not changed | Yes | |||||||
| Salix matsudana | CB | BD | B/L WU | NA | N.A. | SS increase, St decrease in B and W | 6 hours | Yes | [101] |
| Quercus pubescens | Pot | SD | SR | 73% | Increase in SS St not changed | N.A. | 5 days | Yes | [121] |
| Prunus mahleb | SD | SR | 30% | SS not changed Increase in St | N.A. | No | |||
| Robinia pseudoacacia | SD | SR | 68% | Decrease in St and SS | N.A. | No | |||
| Ailanthus altissima | SD | SR | 62% | Decrease in St and SS | N.A. | Yes | |||
| Malus domestica var. Golden delicious | Field | FT + FD | SR | 70% | St not changed in W and B | St decrease in W and B, more pronounced in W | 1 to 3 months | Yes | [122] |
| Malus domestica (4 cultivars) | Field | FT + FD | SR | 20–80% | N.A. | St increase in W and B | Several weeks | Yes | [123] |
| Picea abies | Field | FT + FD | B/L WU | 43% | SS not changed Very low, constant St in W and B | St increase in phloem and needles SS not changed Very low St in W | 1 month | Yes | [50] |
| SD | SR | 30% | St increase in phloem | St decrease in phloem | 3 months | Yes | |||
| Picea abies | Pot | SD | SR | 20% | St depletion in B Tot not changed in B and W | 30% depletion of Tot in W | One week | Yes | [8] |
| Pinus mugo | Field | FT + FD | B/L WU | 40% | St and SS not changed | St increase only in phloem and needles SS not changed | One month | Yes | [124] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomasella, M.; Petrussa, E.; Petruzzellis, F.; Nardini, A.; Casolo, V. The Possible Role of Non-Structural Carbohydrates in the Regulation of Tree Hydraulics. Int. J. Mol. Sci. 2020, 21, 144. https://doi.org/10.3390/ijms21010144
Tomasella M, Petrussa E, Petruzzellis F, Nardini A, Casolo V. The Possible Role of Non-Structural Carbohydrates in the Regulation of Tree Hydraulics. International Journal of Molecular Sciences. 2020; 21(1):144. https://doi.org/10.3390/ijms21010144
Chicago/Turabian StyleTomasella, Martina, Elisa Petrussa, Francesco Petruzzellis, Andrea Nardini, and Valentino Casolo. 2020. "The Possible Role of Non-Structural Carbohydrates in the Regulation of Tree Hydraulics" International Journal of Molecular Sciences 21, no. 1: 144. https://doi.org/10.3390/ijms21010144
APA StyleTomasella, M., Petrussa, E., Petruzzellis, F., Nardini, A., & Casolo, V. (2020). The Possible Role of Non-Structural Carbohydrates in the Regulation of Tree Hydraulics. International Journal of Molecular Sciences, 21(1), 144. https://doi.org/10.3390/ijms21010144