Evaluation of Residual Human-Induced Pluripotent Stem Cells in Human Chondrocytes by Cell Type-Specific Glycosphingolipid Glycome Analysis Based on the Aminolysis-SALSA Technique

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
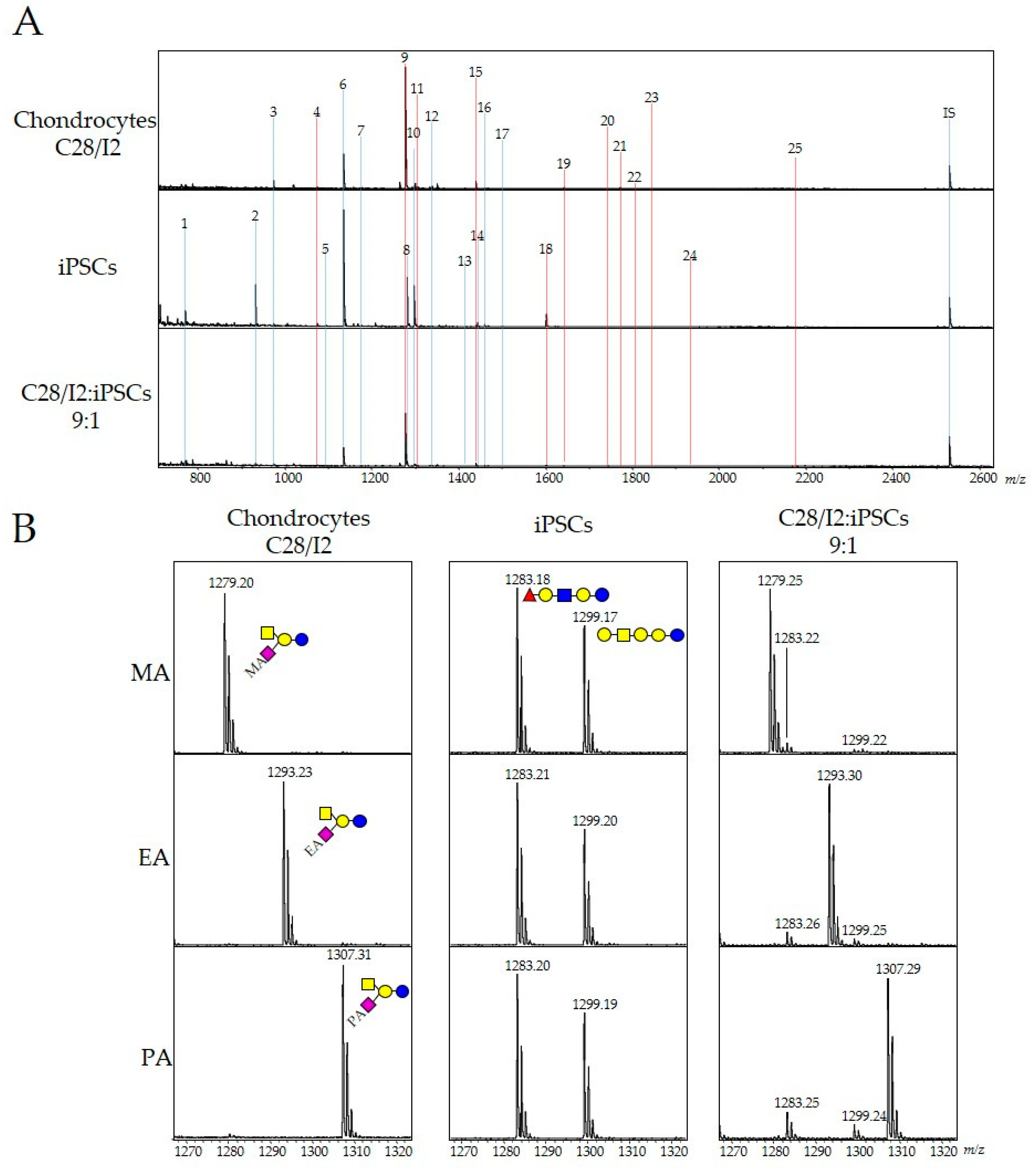
2.1. Detection of Undifferentiated iPSCs with Human Chondrocytes by GSL-Glycome Analysis
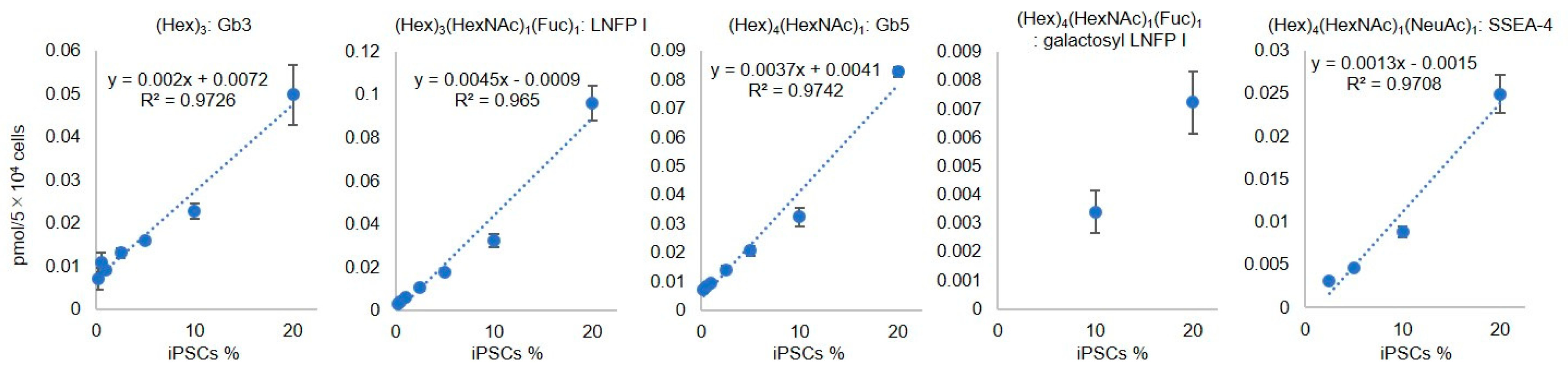
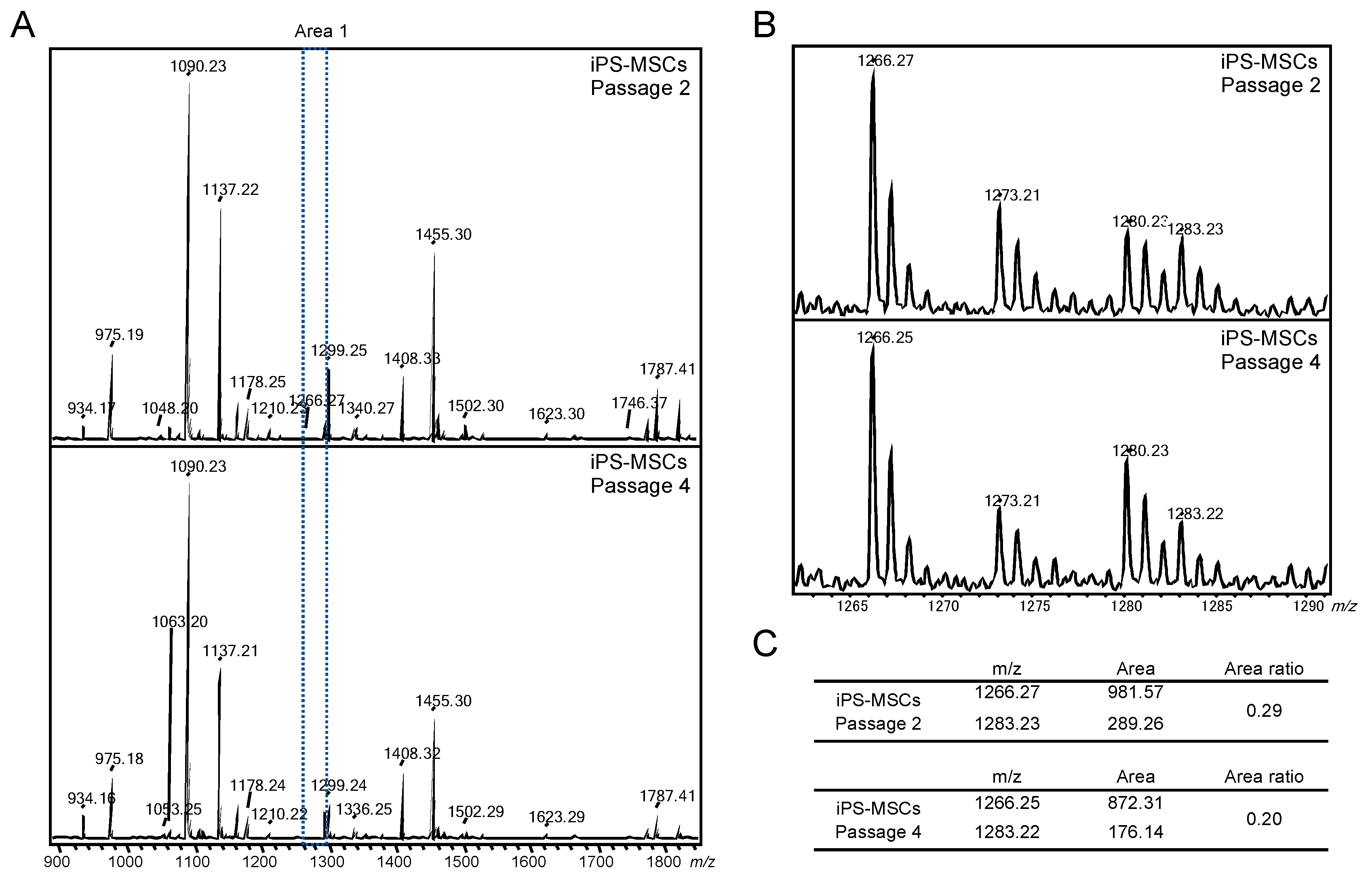
2.2. Evaluation of Residual iPSCs among Human Chondrocytes by Cell Type-Specific GSL-Glycome Analysis
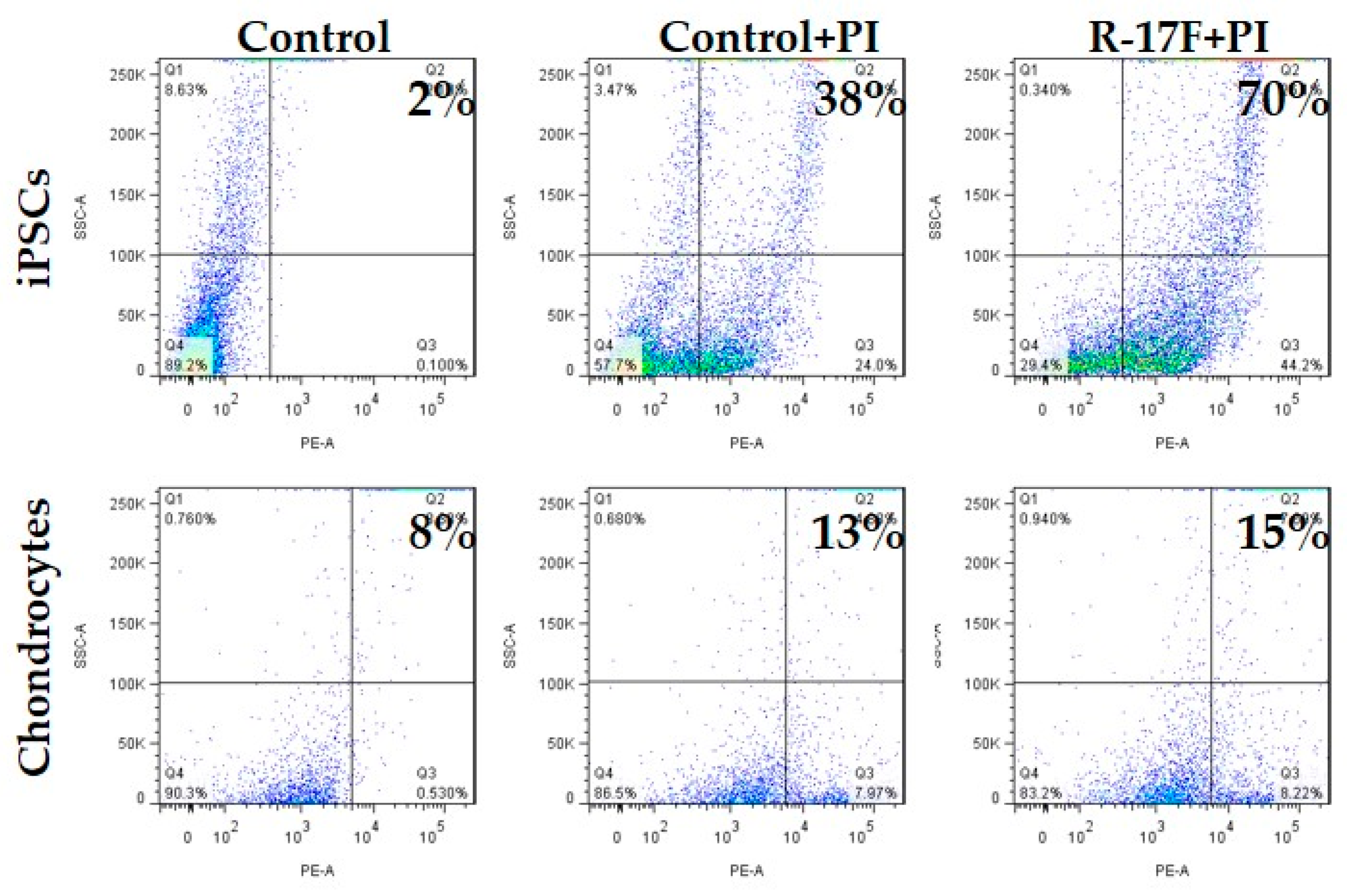
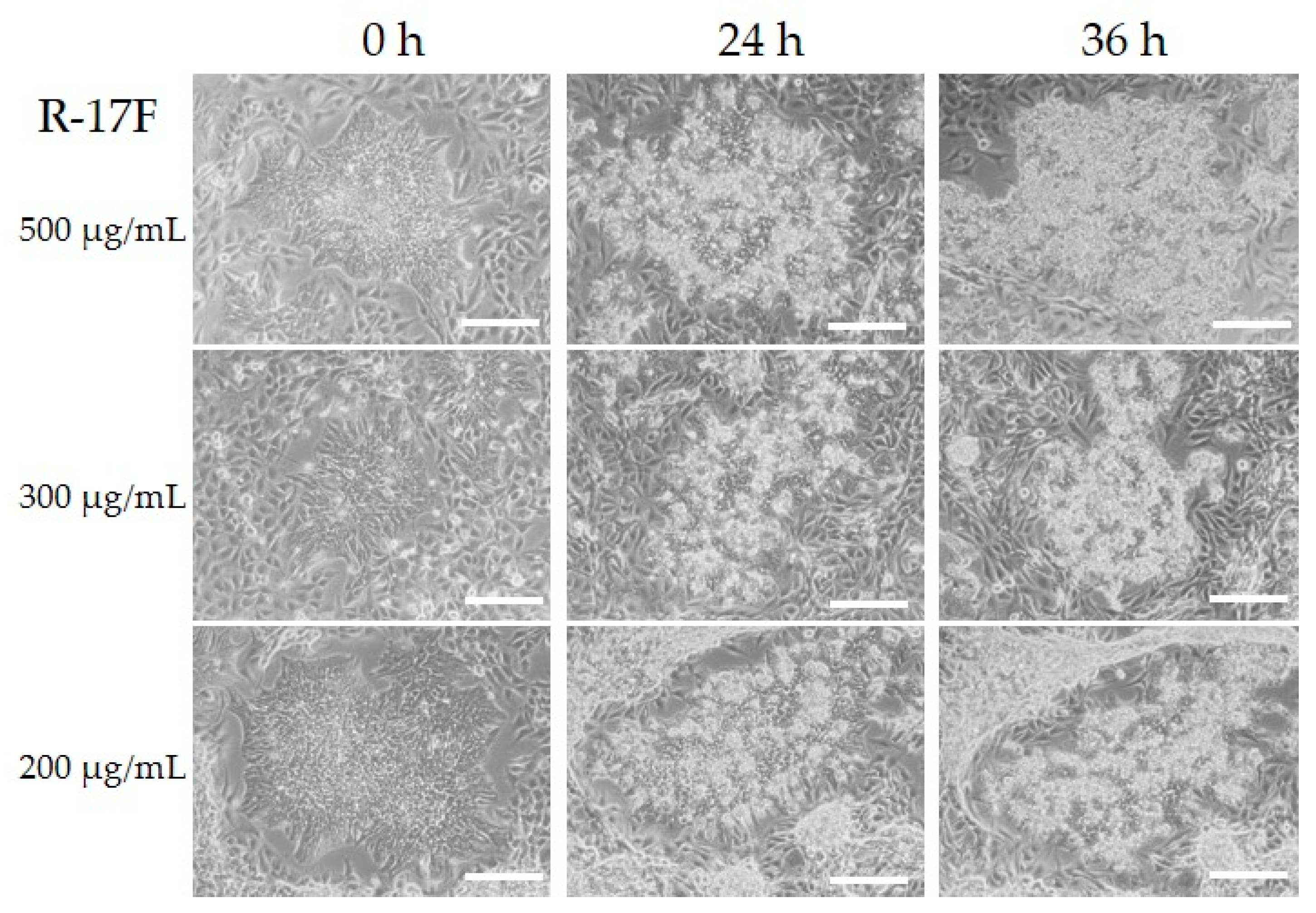
2.3. Evaluation of R-17F-Induced Cytotoxicity Toward iPSCs by GSL-Glycome Analysis
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Materials
4.2. Cell Culture
4.3. GSL-Glycome Analysis of iPSCs Co-Cultured with Chondrocytes
4.4. Flow Cytometry Analysisof the Cytotoxic Activity of R-17F
4.5. GSL-Glycome Analysis of the Cytotoxic Activity of R-17F in Chondrocytes Co-Cultured with iPSCs
4.6. In Vitro Differentiation of iPSCs into iPSC-Derived MSCs
4.7. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Brittberg, M.; Lindahl, A.; Nilsson, A.; Ohlsson, C.; Isaksson, O.; Peterson, L. Treatment of deep cartilage defects in the knee with autologous chondrocyte transplantation. N. Engl. J. Med. 1994, 331, 889–895. [Google Scholar] [CrossRef]
- Roberts, S.; Menage, J.; Sandell, L.J.; Evans, E.H.; Richardson, J.B. Immunohistochemical study of collagen types I and II and procollagen IIA in human cartilage repair tissue following autologous chondrocyte implantation. Knee 2009, 16, 398–404. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, A.; Morioka, M.; Yahara, Y.; Okada, M.; Kobayashi, T.; Kuriyama, S.; Matsuda, S.; Tsumaki, N. Generation of scaffoldless hyaline cartilaginous tissue from human iPSCs. Stem Cell Rep. 2015, 4, 404–418. [Google Scholar] [CrossRef] [Green Version]
- Kawamata, S.; Kanemura, H.; Sakai, N.; Takahashi, M.; Go, M.J. Design of a Tumorigenicity Test for Induced Pluripotent Stem Cell (iPSC)-Derived Cell Products. J. Clin. Med. 2015, 4, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Nagashima, T.; Shimizu, K.; Matsumoto, R.; Honda, H. Selective Elimination of Human Induced Pluripotent Stem Cells Using Medium with High Concentration of L-Alanine. Sci. Rep. 2018, 8, 12427. [Google Scholar] [CrossRef] [Green Version]
- Tateno, H.; Onuma, Y.; Ito, Y.; Minoshima, F.; Saito, S.; Shimizu, M.; Aiki, Y.; Asashima, M.; Hirabayashi, J. Elimination of tumorigenic human pluripotent stem cells by a recombinant lectin-toxin fusion protein. Stem Cell Rep. 2015, 4, 811–820. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Lee, A.S.; Volkmer, J.P.; Sahoo, D.; Nag, D.; Mosley, A.R.; Inlay, M.A.; Ardehali, R.; Chavez, S.L.; Pera, R.R.; et al. An antibody against SSEA-5 glycan on human pluripotent stem cells enables removal of teratoma-forming cells. Nat. Biotechnol. 2011, 29, 829–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, S.; Nakao, H.; Kawabe, K.; Nonaka, M.; Toyoda, H.; Takishima, Y.; Kawabata, K.; Yamaguchi, T.; Furue, M.K.; Taki, T.; et al. A Cytotoxic Antibody Recognizing Lacto-N-fucopentaose I (LNFP I) on Human Induced Pluripotent Stem (hiPS) Cells. J. Biol. Chem. 2015, 290, 20071–20085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, J.I.; Okada, K.; Shinohara, Y. Glycomics of human embryonic stem cells and human induced pluripotent stem cells. Glycoconj. J. 2017, 34, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.J.; Kuo, H.H.; Lin, C.H.; Chen, Y.Y.; Yang, B.C.; Cheng, Y.Y.; Yu, A.L.; Khoo, K.H.; Yu, J. Switching of the core structures of glycosphingolipids from globo- and lacto- to ganglio-series upon human embryonic stem cell differentiation. Proc. Natl. Acad. Sci. USA 2010, 107, 22564–22569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homan, K.; Hanamatsu, H.; Furukawa, J.I.; Okada, K.; Yokota, I.; Onodera, T.; Iwasaki, N. Alteration of the Total Cellular Glycome during Late Differentiation of Chondrocytes. Int. J. Mol. Sci. 2019, 20, 3546. [Google Scholar] [CrossRef] [Green Version]
- Fujitani, N.; Takegawa, Y.; Ishibashi, Y.; Araki, K.; Furukawa, J.; Mitsutake, S.; Igarashi, Y.; Ito, M.; Shinohara, Y. Qualitative and quantitative cellular glycomics of glycosphingolipids based on rhodococcal endoglycosylceramidase-assisted glycan cleavage, glycoblotting-assisted sample preparation, and matrix-assisted laser desorption ionization tandem time-of-flight mass spectrometry analysis. J. Biol. Chem. 2011, 286, 41669–41679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanamatsu, H.; Nishikaze, T.; Miura, N.; Piao, J.; Okada, K.; Sekiya, S.; Iwamoto, S.; Sakamoto, N.; Tanaka, K.; Furukawa, J.I. Sialic Acid Linkage Specific Derivatization of Glycosphingolipid Glycans by Ring-Opening Aminolysis of Lactones. Anal. Chem. 2018, 90, 13193–13199. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Gulbranson, D.R.; Hou, Z.; Bolin, J.M.; Ruotti, V.; Probasco, M.D.; Smuga-Otto, K.; Howden, S.E.; Diol, N.R.; Propson, N.E.; et al. Chemically defined conditions for human iPSC derivation and culture. Nat. Methods 2011, 8, 424–429. [Google Scholar] [CrossRef] [Green Version]
- Guzzo, R.M.; Gibson, J.; Xu, R.H.; Lee, F.Y.; Drissi, H. Efficient differentiation of human iPSC-derived mesenchymal stem cells to chondroprogenitor cells. J. Cell. Biochem. 2013, 114, 480–490. [Google Scholar] [CrossRef]
- Gropp, M.; Shilo, V.; Vainer, G.; Gov, M.; Gil, Y.; Khaner, H.; Matzrafi, L.; Idelson, M.; Kopolovic, J.; Zak, N.B.; et al. Standardization of the teratoma assay for analysis of pluripotency of human ES cells and biosafety of their differentiated progeny. PLoS ONE 2012, 7, e45532. [Google Scholar] [CrossRef]
- Cho, S.J.; Kim, S.Y.; Jeong, H.C.; Cheong, H.; Kim, D.; Park, S.J.; Choi, J.J.; Kim, H.; Chung, H.M.; Moon, S.H.; et al. Repair of Ischemic Injury by Pluripotent Stem Cell Based Cell Therapy without Teratoma through Selective Photosensitivity. Stem Cell Rep. 2015, 5, 1067–1080. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, T.; Yasuda, S.; Kusakawa, S.; Hirata, N.; Kanda, Y.; Suzuki, K.; Takahashi, M.; Nishikawa, S.; Kawamata, S.; Sato, Y. Highly sensitive in vitro methods for detection of residual undifferentiated cells in retinal pigment epithelial cells derived from human iPS cells. PLoS ONE 2012, 7, e37342. [Google Scholar] [CrossRef] [Green Version]
- Kuang, Y.; Miki, K.; Parr, C.J.C.; Hayashi, K.; Takei, I.; Li, J.; Iwasaki, M.; Nakagawa, M.; Yoshida, Y.; Saito, H. Efficient, Selective Removal of Human Pluripotent Stem Cells via Ecto-Alkaline Phosphatase-Mediated Aggregation of Synthetic Peptides. Cell Chem. Biol. 2017, 24, 685–694.e684. [Google Scholar] [CrossRef] [Green Version]
- Ben-David, U.; Gan, Q.F.; Golan-Lev, T.; Arora, P.; Yanuka, O.; Oren, Y.S.; Leikin-Frenkel, A.; Graf, M.; Garippa, R.; Boehringer, M.; et al. Selective elimination of human pluripotent stem cells by an oleate synthesis inhibitor discovered in a high-throughput screen. Cell Stem Cell 2013, 12, 167–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, J.; Sakai, S.; Yokota, I.; Okada, K.; Hanamatsu, H.; Kobayashi, T.; Yoshida, Y.; Higashino, K.; Tamura, T.; Igarashi, Y.; et al. Quantitative GSL-glycome analysis of human whole serum based on an EGCase digestion and glycoblotting method. J. Lipid Res. 2015, 56, 2399–2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Glycan Composition | Class | m/z | Chondrocytes (pmol/5 × 104) | iPSCs (pmol/5 × 104) | Relative Amount iPSCs | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | (Hex)2 | Neutral | 772.39 | 0.06 | ± | 0.01 | 0.22 | ± | 0.02 | 3.6 |
| 2 | (Hex)3 | Neutral | 934.47 | 0.04 | ± | 0.01 | 0.80 | ± | 0.08 | 21.2 |
| 3 | (Hex)2(HexNAc)1 | Neutral | 975.5 | 0.19 | ± | 0.04 | 0.06 | ± | 0.00 | 0.3 |
| 5 | (Hex)4 | Neutral | 1096.53 | 0.00 | ± | 0.00 | 0.02 | ± | 0.01 | INF |
| 6 | (Hex)3(HexNAc)1 | Neutral | 1137.56 | 0.99 | ± | 0.18 | 2.93 | ± | 0.32 | 3.0 |
| 7 | (Hex)2(HexNAc)2 | Neutral | 1178.59 | 0.04 | ± | 0.01 | 0.04 | ± | 0.00 | 1.1 |
| 8 | (Hex)3(HexNAc)1(Fuc)1 | Neutral | 1283.63 | 0.00 | ± | 0.00 | 1.36 | ± | 0.18 | INF |
| 10 | (Hex)4(HexNAc)1 | Neutral | 1299.67 | 0.01 | ± | 0.01 | 1.15 | ± | 0.13 | 174.5 |
| 12 | (Hex)3(HexNAc)2 | Neutral | 1340.67 | 0.09 | ± | 0.02 | 0.03 | ± | 0.00 | 0.3 |
| 13 | (Hex)3(HexNAc)1(Fuc)2 | Neutral | 1429.7 | 0.00 | ± | 0.00 | 0.01 | ± | 0.01 | INF |
| 14 | (Hex)4(HexNAc)1(Fuc)1 | Neutral | 1445.71 | 0.00 | ± | 0.00 | 0.14 | ± | 0.01 | INF |
| 16 | (Hex)5(HexNAc)1 | Neutral | 1461.71 | 0.01 | ± | 0.01 | 0.07 | ± | 0.01 | 10.5 |
| 17 | (Hex)4(HexNAc)2 | Neutral | 1502.72 | 0.01 | ± | 0.01 | 0.02 | ± | 0.01 | 1.5 |
| 4 | (Hex)2(α2,3NeuAc)1 | Ganglioside | 1076.57 | 0.04 | ± | 0.01 | 0.07 | ± | 0.01 | 1.7 |
| 9 | (Hex)2(HexNAc)1(α2,3NeuAc)1 | Ganglioside | 1279.67 | 4.25 | ± | 0.87 | 0.00 | ± | 0.00 | 0 |
| 11 | (Hex)2(HexNAc)1(α2,6NeuAc)1 | Ganglioside | 1307.7 | 0.07 | ± | 0.01 | 0.00 | ± | 0.00 | 0 |
| 15 | (Hex)3(HexNAc)1(α2,3NeuAc)1 | Ganglioside | 1441.73 | 0.27 | ± | 0.05 | 0.06 | ± | 0.01 | 0.2 |
| 18 | (Hex)4(HexNAc)1(α2,3NeuAc)1 | Ganglioside | 1603.78 | 0.00 | ± | 0.00 | 0.46 | ± | 0.05 | INF |
| 19 | (Hex)3(HexNAc)2(α2,3NeuAc)1 | Ganglioside | 1644.81 | 0.05 | ± | 0.01 | 0.00 | ± | 0.00 | 0 |
| 20 | (Hex)3(HexNAc)1(α2,3NeuAc)2 | Ganglioside | 1745.86 | 0.01 | ± | 0.01 | 0.00 | ± | 0.00 | 0 |
| 21 | (Hex)3(HexNAc)1(α2,3NeuAc)1(α2,6NeuAc)1 | Ganglioside | 1773.88 | 0.06 | ± | 0.01 | 0.00 | ± | 0.00 | 0 |
| 22 | (Hex)4(HexNAc)2(α2,3NeuAc)1 | Ganglioside | 1806.85 | 0.02 | ± | 0.00 | 0.00 | ± | 0.00 | 0 |
| 23 | (Hex)3(HexNAc)3(α2,3NeuAc)1 | Ganglioside | 1847.87 | 0.01 | ± | 0.01 | 0.00 | ± | 0.00 | 0 |
| 24 | (Hex)4(HexNAc)1(α2,3NeuAc)1(α2,6NeuAc)1 | Ganglioside | 1935.87 | 0.00 | ± | 0.00 | 0.03 | ± | 0.00 | INF |
| 25 | (Hex)5(HexNAc)3(α2,3NeuAc)1 | Ganglioside | 2172 | 0.02 | ± | 0.01 | 0.00 | ± | 0.00 | 0 |
| Total | - | - | 6.25 | ± | 1.20 | 7.45 | ± | 0.83 | 1.2 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miyazaki, T.; Hanamatsu, H.; Xu, L.; Onodera, T.; Furukawa, J.-i.; Homan, K.; Baba, R.; Kawasaki, T.; Iwasaki, N. Evaluation of Residual Human-Induced Pluripotent Stem Cells in Human Chondrocytes by Cell Type-Specific Glycosphingolipid Glycome Analysis Based on the Aminolysis-SALSA Technique. Int. J. Mol. Sci. 2020, 21, 231. https://doi.org/10.3390/ijms21010231
Miyazaki T, Hanamatsu H, Xu L, Onodera T, Furukawa J-i, Homan K, Baba R, Kawasaki T, Iwasaki N. Evaluation of Residual Human-Induced Pluripotent Stem Cells in Human Chondrocytes by Cell Type-Specific Glycosphingolipid Glycome Analysis Based on the Aminolysis-SALSA Technique. International Journal of Molecular Sciences. 2020; 21(1):231. https://doi.org/10.3390/ijms21010231
Chicago/Turabian StyleMiyazaki, Takuji, Hisatoshi Hanamatsu, Liang Xu, Tomohiro Onodera, Jun-ichi Furukawa, Kentaro Homan, Rikiya Baba, Toshisuke Kawasaki, and Norimasa Iwasaki. 2020. "Evaluation of Residual Human-Induced Pluripotent Stem Cells in Human Chondrocytes by Cell Type-Specific Glycosphingolipid Glycome Analysis Based on the Aminolysis-SALSA Technique" International Journal of Molecular Sciences 21, no. 1: 231. https://doi.org/10.3390/ijms21010231
APA StyleMiyazaki, T., Hanamatsu, H., Xu, L., Onodera, T., Furukawa, J. -i., Homan, K., Baba, R., Kawasaki, T., & Iwasaki, N. (2020). Evaluation of Residual Human-Induced Pluripotent Stem Cells in Human Chondrocytes by Cell Type-Specific Glycosphingolipid Glycome Analysis Based on the Aminolysis-SALSA Technique. International Journal of Molecular Sciences, 21(1), 231. https://doi.org/10.3390/ijms21010231