Physical Mapping of Pm57, a Powdery Mildew Resistance Gene Derived from Aegilops searsii

 ,
,
Abstract
:1. Introduction
2. Results
2.1. RNA-seq Quantity, Sequence Assembly, and Differential Expression Gene (DEGs) Analysis
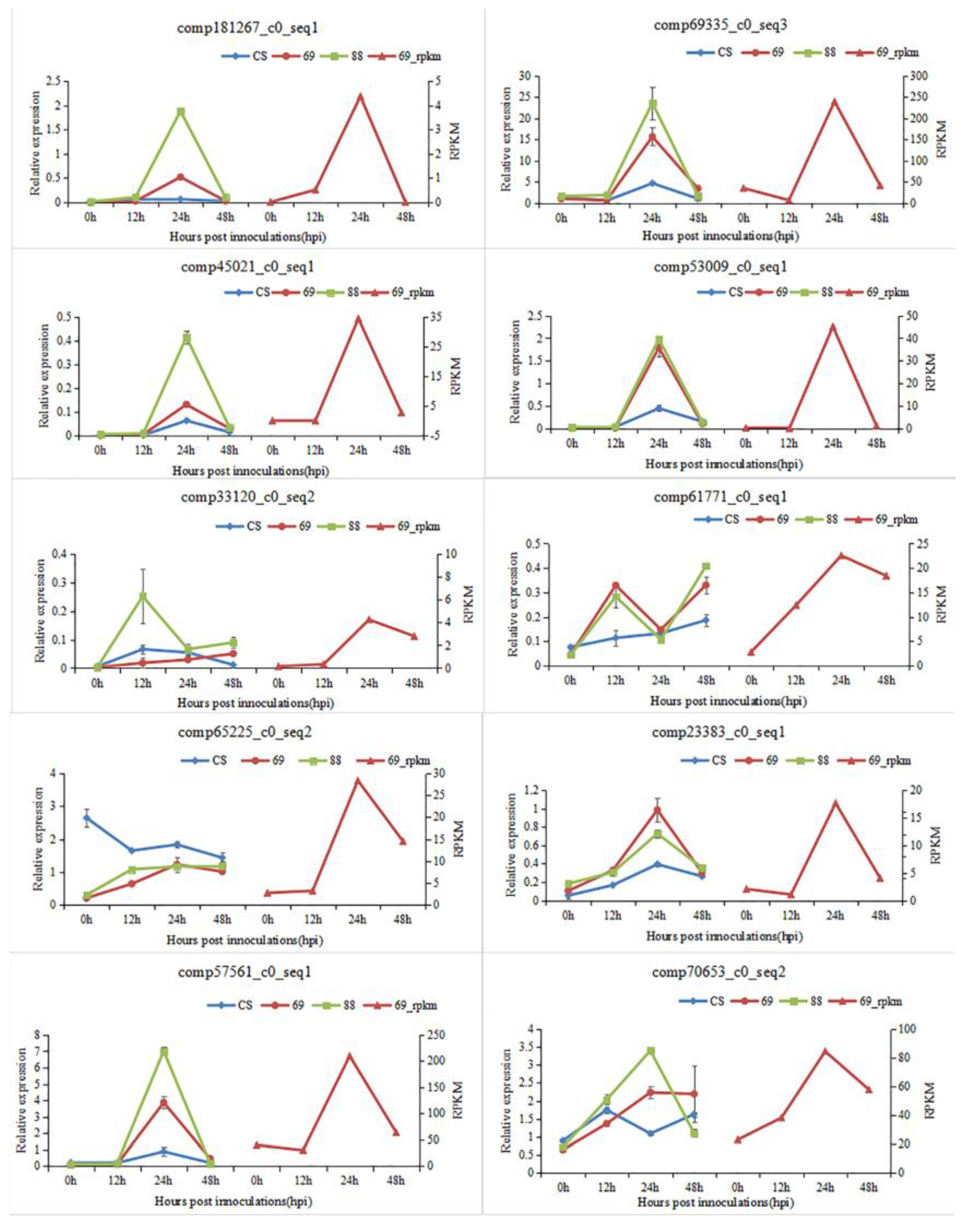
2.2. Validation of DEGs by Quantitative RT-PCR (qRT-PCR)
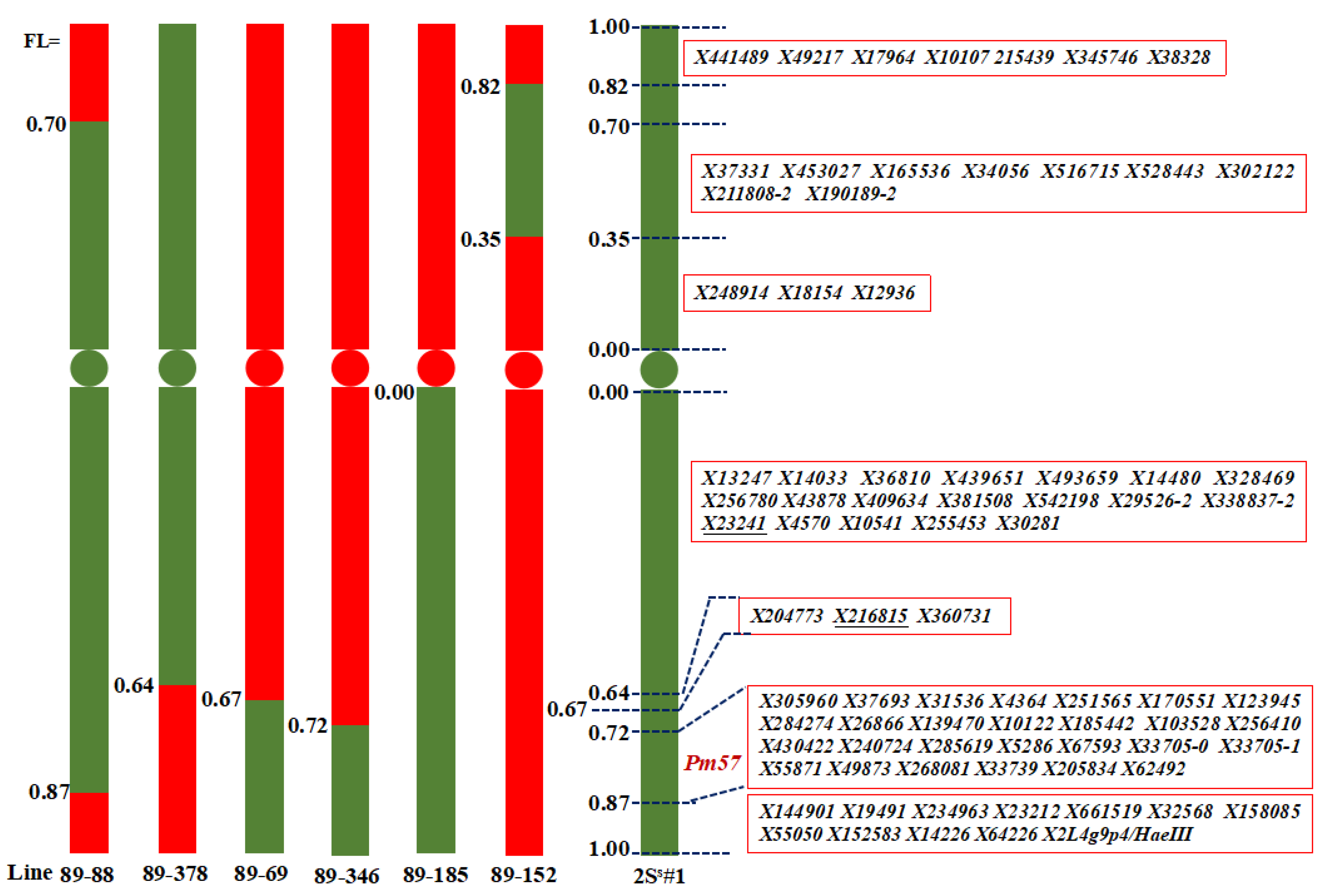
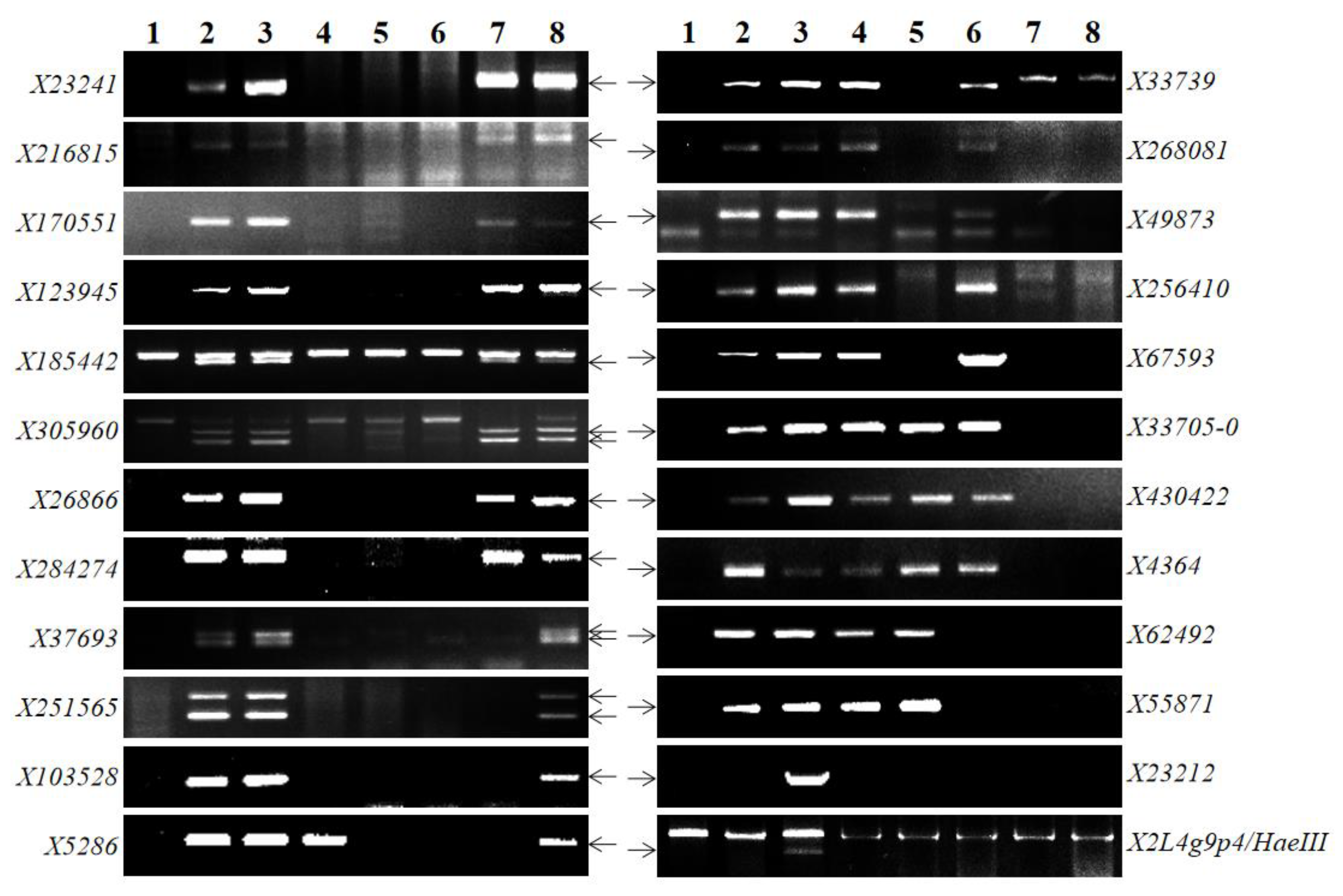
2.3. Chromosome Fraction Length Interval Assignment of Chromosome 2Ss#1-Specific Markers
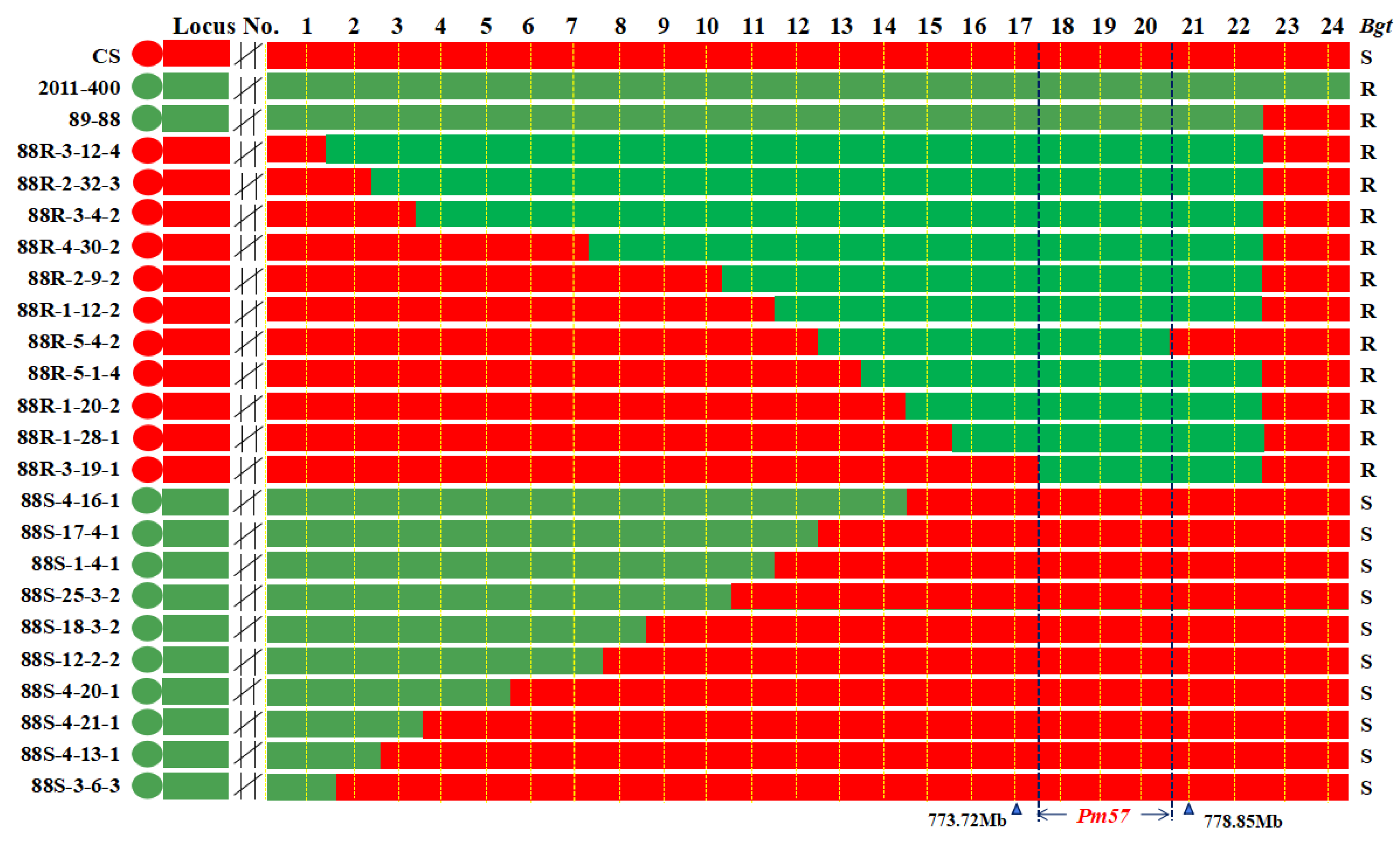
2.4. Development of CS-Ae. searsii Disomic 2Ss#1 Recombinants and Physical Mapping of Pm57
2.5. Comparative Synteny and Genes in Pm57 Candidate Region
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Construction of cDNA Libraries for Illumina Sequencing
4.3. RNA-seq Data Analysis
4.4. Validations of RNA-seq Data by Quantitative RT-PCR
4.5. Molecular Marker Analysis
4.6. Powdery Mildew Response Assay
4.7. Identification of CS-Ae. searsii 2Ss#1 Recombinants and Physical Mapping of Pm57
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Bgt | Blumeria graminis f. sp. tritici |
| FL | fraction length |
| CNL | coiled-coil nucleotide-binding site-leucine-rich repeat |
| DEG | differential expression gene |
| SFL | short-arm fraction length |
| LFL | long-arm fraction length |
| RPKM | reads per kilobase per million mapped reads |
| IT | infection types |
| VIGS | virus-induced gene silencing |
References
- Fried, P.; Mackenzie, D.; Nelson, R. Yield loss caused by Erysiphe graminis f. sp. tritici on single culms of “Chancellor” wheat and four multilines. J. Plant Dis. Prot. 1981, 88, 256–264. [Google Scholar]
- Conner, R.; Kuzyk, A.; Su, H. Impact of powdery mildew on the yield of soft white spring wheat cultivars. Can. J. Plant Sci. 2003, 83, 725–728. [Google Scholar] [CrossRef]
- Morgounov, A.; Tufan, H.; Sharma, R.; Akin, B.; Bagci, A.; Braun, H.; Kaya, Y.; Keser, M.; Payne, T.; Sonder, K.; et al. Global incidence of wheat rusts and powdery mildew during 1969–2010 and durability of resistance of winter wheat variety Bezostaya 1. Eur. J. Plant Pathol. 2012, 132, 323–340. [Google Scholar] [CrossRef]
- Bennett, F. Resistance to powdery mildew in wheat: A review of its use in agriculture and breeding programs. Plant Pathol. 1984, 33, 279–300. [Google Scholar] [CrossRef]
- Wang, Z.L.; Li, L.H.; He, Z.H.; Duan, X.Y.; Zhou, Y.L.; Chen, X.M.; Lillemo, M.; Singh, R.P.; Wang, H.; Xia, X.C. Seedling and adult plant resistance to powdery mildew in Chinese bread wheat cultivars and lines. Plant Dis. 2005, 89, 457–463. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, R.; Dubcovsky, J.; Rogers, W.; Morris, C. Catalogue of gene symbols for wheat. 2017. Supplement. Ann. Wheat Newsl. 2017, 53, 107–128. [Google Scholar]
- Liu, W.X.; Koo, D.; Xia, Q.; Li, C.X.; Bai, F.Q.; Song, Y.L.; Friebe, B.; Gill, B.S. Homoeologous recombination-based transfer and molecular cytogenetic mapping of powdery mildew-resistant gene Pm57 from Aegilops searsii into wheat. Theor. Appl. Genet. 2017, 130, 841–848. [Google Scholar] [CrossRef]
- Zou, S.; Wang, H.; Li, Y.; Kong, Z.; Tang, D.Z. The NB-LRR gene Pm60 confers powdery mildew resistance in wheat. New Phytol. 2018, 218, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.; Li, G.; Cowger, C.; Carver, B.; Xu, X.Y. Characterization of Pm63, a powdery mildew resistance gene in Iranian landrace PI 628024. Theor. Appl. Genet. 2019, 132, 1137–1144. [Google Scholar] [CrossRef]
- Zhang, D.; Zhu, K.; Dong, L.; Liang, Y.; Li, G.Q.; Fang, T.L.; Guo, G.H.; Wu, Q.H.; Xie, J.Z.; Chen, Y.X. Wheat powdery mildew resistance gene Pm64 derived from wild emmer (Triticum turgidum var. dicoccoides) is tightly linked in repulsion with stripe rust resistance gene Yr5. Crop. J. 2018, 7, 761–770. [Google Scholar] [CrossRef]
- Li, G.Q.; Cowger, C.; Wang, X.W.; Carver, B.F.; Xu, X.Y. Characterization of Pm65, a new powdery mildew resistance gene on chromosome 2AL of a facultative wheat cultivar. Theor. Appl. Genet. 2019, 132, 2625–2632. [Google Scholar] [CrossRef]
- Zeller, F.; Fuchs, E. Cytology and disease resistance of 1A/1R and some 1B/1R wheat-rye translocation cultivars. J. Plant Dis. Prot. 1983, 90, 285–296. [Google Scholar]
- Chen, P.; Qi, L.; Zhou, B.; Zhang, S.; Liu, D.J. Development and molecular cytogenetic analysis of wheat–Haynaldia villosa 6VS/6AL translocation lines specifying resistance to powdery mildew. Theor. Appl. Genet. 1995, 91, 1125–1128. [Google Scholar] [CrossRef]
- Luo, P.; Zhang, H.; Shu, K.; Wu, X.; Zhang, H.Q.; Ren, Z.L. The physiological genetic effects of 1BL/1RS translocated chromosome in “stay green” wheat cultivar CN17. Can. J. Plant Sci. 2009, 89, 1–10. [Google Scholar] [CrossRef]
- Xing, L.; Hu, P.; Liu, J.; Witek, K.; Zhou, S.; Xu, J.; Zhou, W.; Gao, L.; Huang, Z.; Zhang, R.; et al. Pm21 from Haynaldia villosa encodes a CC-NBS-LRR protein conferring powdery mildew resistance in wheat. Mol. Plant 2018, 11, 874–878. [Google Scholar] [CrossRef] [Green Version]
- Johal, G.S.; Briggs, S.P. Reductase activity encoded by the HM1 disease resistance gene in maize. Science 1992, 258, 985–987. [Google Scholar] [CrossRef]
- Kourelis, J.; van der Hoo, R. Defended to the nines: 25 years of resistance gene cloning identifies nine mechanisms for R protein function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.B.; Brommonschenkel, S.H.; Chunwongse, J.; Frary, A.; Ganal, M.W.; Spivey, R.; Wu, T.; Earle, E.D.; Tanksley, S.D. Map-based cloning of a protein kinase gene conferring disease resistance in tomato. Science 1993, 262, 1432–1436. [Google Scholar] [CrossRef]
- Bent, A.F.; Kunkel, B.N.; Dahlbeck, D.; Brown, K.L.; Schmidt, R.; Giraudat, J.; Leung, J.; Staskawicz, B.J. RPS2 of Arabidopsis thaliana: A leucine-rich repeat class of plant disease resistance genes. Science 1994, 265, 1856–1860. [Google Scholar] [CrossRef]
- Song, W.Y.; Wang, G.L.; Chen, L.L.; Kim, H.S.; Pi, L.Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.X.; Zhu, L.H. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [Green Version]
- Staskawicz, B.J.; Ausubel, F.M.; Baker, B.J.; Ellis, J.G.; Jones, J.D. Molecular genetics of plant disease resistance. Science 1995, 268, 661–667. [Google Scholar] [CrossRef] [Green Version]
- Periyannan, S. Sustaining global agriculture through rapid detection and deployment of genetic resistance to deadly crop diseases. New Phytol. 2018, 219, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Keller, B.; Feuillet, C.; Yahiaoui, N. Map-based isolation of disease resistance genes from bread wheat: Cloning in a supersize genome. Genet. Res. 2005, 85, 93–100. [Google Scholar] [CrossRef]
- Bevan, M.W.; Uauy, C.; Wulff, B.B.; Zhou, J.; Krasileva, K.; Clark, M.D. Genomic innovation for crop improvement. Nature 2017, 543, 346–354. [Google Scholar] [CrossRef]
- Sears, E.R. Genetic control of chromosome pairing in wheat. Ann. Rev. Genet. 1976, 10, 31–51. [Google Scholar] [CrossRef]
- Martinez-Perez, E.; Moore, G. To check or not to check? The application of meiotic studies to plant breeding. Curr. Opin. Plant Biol. 2008, 11, 222–227. [Google Scholar] [CrossRef]
- Able, J.A.; Crismani, W.; Boden, S.A. Understanding meiosis and the implications for crop improvement. Funct. Plant Biol. 2009, 36, 575–588. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Liu, S.; Yeh, C.T.; Tang, H.M.; Nettleton, D.; Schnable, P.S. Gene mapping via bulked segregant RNA-seq (BSR-Seq). PLoS ONE 2012, 7, e36406. [Google Scholar] [CrossRef] [Green Version]
- Ling, H.; Zhao, S.; Liu, D.; Wang, J.Y.; Sun, H.; Zhang, C.; Fan, H.J.; Li, D.; Dong, L.L.; Tao, Y. Draft genome of the wheat A-genome progenitor Triticum urartu. Nature 2013, 496, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Jia, J.; Zhao, S.; Kong, X.; Li, Y.R.; Zhao, G.Y.; He, W.M.; Appels, R.; Pfeifer, M.; Tao, Y.; Zhang, Y.X. Aegilops tauschii draft genome sequence reveals a gene repertoire for wheat adaptation. Nature 2013, 496, 91–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.C.; Zheng, H.Y.; Li, Y.W.; Li, H.J.; Liu, X.; Qin, H.J.; Dong, L.L.; Wang, D.W. Coexpression network analysis of the genes regulated by two types of resistance responses to powdery mildew in wheat. Sci. Rep. 2016, 6, 23805. [Google Scholar] [CrossRef] [PubMed]
- Avni, R.; Nave, M.; Barad, O.; Baruch, K.; Twardziok, S.O.; Gundlach, H.; Hale, I.; Mascher, M.; Spannagl, M.; Wiebe, K. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 2017, 357, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Thind, A.; Wicker, T.; Šimková, H.; Fossati, D.; Moullet, O.; Brabant, C.; Vrana, J.; Dolezel, J.; Krattinger, S.G. Rapid cloning of genes in hexaploid wheat using cultivar-specific long-range chromosome assembly. Nat. Biotechnol. 2017, 35, 793–796. [Google Scholar] [CrossRef]
- Keller, B.; Wicker, T.; Krattinger, S.G. Advances in wheat and pathogen genomics: Implications for disease control. Ann. Rev. Phytopathol. 2018, 56, 67–87. [Google Scholar] [CrossRef] [Green Version]
- Yahiaoui, N.; Srichumpa, P.; Dudler, R.; Keller, B. Genome analysis at different ploidy levels allows cloning of the powdery mildew resistance gene Pm3b from hexaploid wheat. Plant J. 2004, 37, 528–538. [Google Scholar] [CrossRef]
- Hurni, S.; Brunner, S.; Buchmann, G.; Herren, G.; Jordan, T.; Krukowski, P.; Wicker, T.; Yahiaoui, N.; Mago, R.; Keller, B. Rye Pm8 and wheat Pm3 are orthologous genes and show evolutionary conservation of resistance function against powdery mildew. Plant J. 2013, 76, 957–969. [Google Scholar] [CrossRef]
- Jupe, F.; Witek, K.; Verweij, W.; Sliwka, J.; Pritchard, L.; Etherington, G.; Maclean, D.; Cock, P.J.; Leggett, R.M.; Bryan, G.J. Resistance gene enrichment sequencing (RenSeq) enables reannotation of the NB-LRR gene family from sequenced plant genomes and rapid mapping of resistance loci in segregating populations. Plant J. 2013, 76, 530–544. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Martin, J.; Steuernagel, B.; Ghosh, S.; Herren, G.; Hurni, S.; Adamski, N.; Vrana, J.; Kubalakova, M.; Krattinger, S.M.; Wicker, T. Rapid gene isolation in barley and wheat by mutant chromosome sequencing. Genome Biol. 2016, 17, 221. [Google Scholar] [CrossRef] [Green Version]
- Witek, K.; Jupe, F.; Witek, A.; Baker, D.; Clark, M.D.; Jones, J.D. Accelerated cloning of a potato late blight–resistance gene using RenSeq and SMRT sequencing. Nat. Biotechnol. 2016, 34, 656–660. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Zhu, S.; Zhao, R.; Jiang, Z.; Ji, Y.; Ji, J.; Qiu, D.; Li, H.; Bie, T. Pm21, encoding a typical CC-NBS-LRR protein, confers broad-spectrum resistance to wheat powdery mildew disease. Mol. Plant 2018, 11, 879–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, M.; Wang, X.; Peng, H.; Yao, Y.; Xie, C.J.; Han, Y.; Ni, Z.F.; Sun, Q.X. Transcriptome comparison of susceptible and resistant wheat in response to powdery mildew infection. Genom. Proteom. Bioinform. 2012, 10, 94–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boni, R.; Chauhan, H.; Hensel, G.; Roulin, A.; Sucher, J.; Kumlehn, J.; Brunner, S.; Krattinger, S.G.; Keller, B. Pathogen-inducible Ta-Lr34res expression in heterologous barley confers disease resistance without negative pleiotropic effects. Plant Biotechnol. J. 2017, 16, 245–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, A.; Xing, L.; Wang, X.; Yang, X.; Wang, W.; Sun, Y.; Qian, C.; Ni, J.L.; Chen, Y.P.; Liu, D.J.; et al. Serine/threonine kinase gene Stpk-V, a key member of powdery mildew resistance gene Pm21, confers powdery mildew resistance in wheat. Proc. Natl. Acad. Sci. USA 2011, 108, 7727–7732. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Li, Y.; Fei, F.; Wang, Z.; Wang, W.; Cao, A.; Liu, Y.; Han, S.; Xing, L.; Wang, H.Y. E3 ubiquitin ligase gene CMPG1–V from Haynaldia villosa L. contributes to powdery mildew resistance in common wheat (Triticum aestivum L.). Plant J. 2015, 84, 154–168. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Lin, Z.; Liu, C.; Wang, K.; Du, L.P.; Ye, X.G. Development and comparative genomic mapping of Dasypyrum villosum 6V#4S-specific PCR markers using transcriptome data. Theor. Appl. Genet. 2017, 130, 2057–2068. [Google Scholar]
- Wang, K.; Lin, Z.; Wang, L.; Wang, K.; Shi, Q.H.; Du, L.P.; Ye, X.G. Development of a set of PCR markers specific to Aegilops longissima chromosome arms and application in breeding a translocation line. Theor. Appl. Genet. 2018, 131, 13–25. [Google Scholar] [CrossRef]
- Feldman, M.; Kislev, M. Aegilops searsii, a new species of section Sitopsisi (Platystachys). Isr. J. Plant Sci. 1977, 26, 190–201. [Google Scholar]
- Grabherr, M.; Haas, B.; Yassour, M.; Levin, J.; Thompson, D.A.; Amit, I.; Adicoins, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Sears, E.R. Genetics society of canada award of excellence lecture an induced mutant with homoeologous pairing in common wheat. Can. J. Genet. Cytol. 1977, 19, 585–593. [Google Scholar] [CrossRef]
- Friebe, B.; Tuleen, N.A.; Gill, B.S. Standard karyotype of Triticum searsii and its relationship with other S-genome species and common wheat. Theor. Appl. Genet. 1995, 91, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Dodds, P.; Rathjen, J. Plant immunity: Towards an integrated view of plant–pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.; Dang, J. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekhwal, M.; Li, P.; Lam, I.; Wang, X.; Cloutier, S.; You, F. Disease resistance gene analogs (RGAs) in plants. Int. J. Mol. Sci. 2015, 16, 19248–19290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raupp, W.; Friebe, B.; Gill, B. Suggested guidelines for the nomenclature and abbreviation of the genetic stocks of wheat and its relatives. Wheat Inf. Serv. 1995, 81, 50–55. [Google Scholar]
- Li, H.H.; Dong, Z.J.; Ma, C.; Tian, X.B.; Xiang, Z.G.; Xia, Q.; Ma, P.T.; Liu, W.X. Discovery of powdery mildew resistance gene candidates from Aegilops biuncialis chromosome 2Mb based on transcriptome sequencing. PLoS ONE 2019, 14, e0220089. [Google Scholar] [CrossRef] [Green Version]
- Osuna-Cruz, C.; Paytuvi-Gallart, A.; Di Donato, A.; Sundesha, V.; Andolfo, G.; Cigliano, R.; Sanseverino, W.; Ercolano, M.R. PRGdb 3.0: A comprehensive platform for prediction and analysis of plant disease resistance genes. Nucleic Acids Res. 2017, 46, D1197–D1201. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Li, H.H.; Dong, Z.J.; Ma, C.; Tian, X.B.; Qi, Z.J.; Wu, N.; Friebe, B.; Xiang, Z.G.; Xia, Q.; Liu, W.X. Physical mapping of stem rust resistance gene Sr52 from Dasypyrum villosum based on ph1b-induced homoeologous recombination. Int. Mol. Sci. 2019, 20, 4887. [Google Scholar] [CrossRef] [Green Version]
- Shi, Q.; Zhang, X.; Duan, X. Identification of isolates of Blumeria graminis f. sp. tritici. Sci. Agric. Sin. 1987, 20, 64–70. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
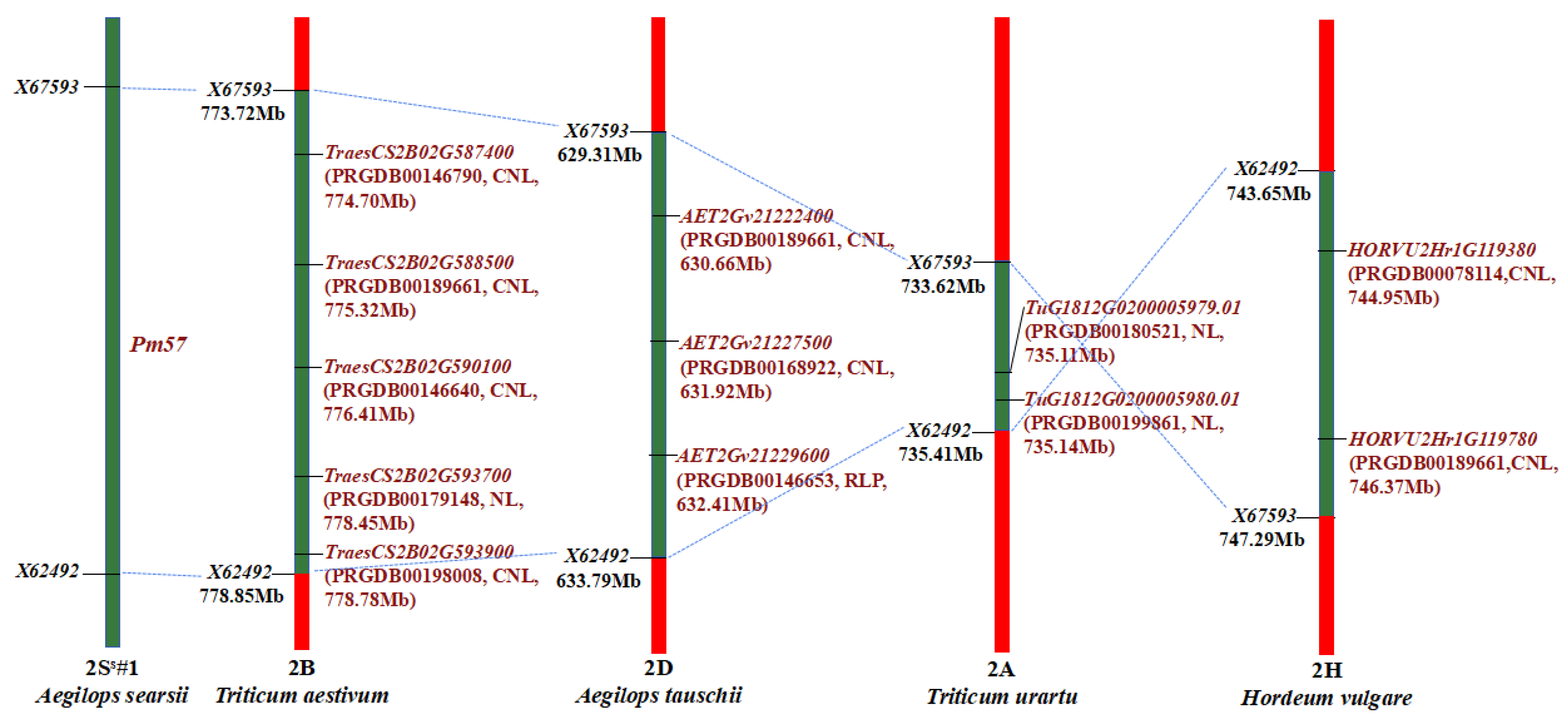{kind=link}
| WGRC 1 Accession Number | Description | Fraction Length Interval Harboring 2Ss#1 Segment | Reference |
|---|---|---|---|
| TA3808 | CS | - 2 | - 3 |
| TA3809 | CS ph1b mutant | - 2 | Sears (1977) [50] |
| TA3581 (2011-400) | CS-Ae. searsii disomic 2Ss#1 addition line | - 2 | Friebe et al. (1995) [51] |
| TA5109 (89-69) | CS-Ae. searsii T2BS.2BL-2Ss#1L recombinant line | LFL0.67-1.00 | Liu et al. (2017) [7] |
| 89-88 | CS-Ae. searsii Ti2AS-2Ss#1S.2Ss#1L-2AL recombinant line | SFL0.70-LFL0.87 | Liu et al. (2017) [7] |
| 89-152 4 | CS-Ae. searsii TiW?S-2Ss#1S-W?S.W?L recombinant line | SFL0.35-0.82 | Liu et al. (2017) [7] |
| 89-185 | CS-Ae. searsii ditelosomic 2Ss#1L addition line | LFLC-1.00 | Liu et al. (2017) [7] |
| 89-346 | CS-Ae. searsii T2BS.2BL-2Ss#1L recombinant line | LFL0.72-1.00 | Liu et al. (2017) [7] |
| 89-378 4 | CS-Ae. searsii T2Ss#1S.2Ss#1L-2W? recombinant line | SFL1.00-LFL0.64 | Liu et al. (2017) [7] |
| Locus 1 No. | Marker Name | Chromosome Bin | Genomic Position (Mb) 2 |
|---|---|---|---|
| 1 | X23241 | LFLC-0.64 | 488.87 |
| 2 | X216815 | LFC0.64-0.67 | 658.61 |
| 3 | X170551 | LFL0.72-0.87 | 320.50 |
| 4 | X123945 | LFL0.72-0.87 | 622.52 |
| 5 | X185442 | LFL0.72-0.87 | 733.47 |
| 6 | X305960 | LFL0.72-0.87 | 744.52 |
| 7 | X26866 | LFL0.72-0.87 | 752.95 |
| 8 | X284274 | LFL0.72-0.87 | 748.97 |
| 9 | X37693 | LFL0.72-0.87 | 754.92 |
| 10 | X251565 | LFL0.72-0.87 | 777.64 |
| 11 | X103528 | LFL0.72-0.87 | 762.50 |
| 12 | X5286/X315363 | LFL0.72-0.87 | 765.05 |
| 13 | X33739 | LFL0.72-0.87 | 766.23 |
| 14 | X268081 | LFL0.72-0.87 | 768.61 |
| 15 | X49873 | LFL0.72-0.87 | 770.18 |
| 16 | X256410 | LFL0.72-0.87 | 771.04 |
| 17 | X67593 | LFL0.72-0.87 | 773.72 |
| 18 | X33705 | LFL0.72-0.87 | 775.02 |
| 19 | X430422/X285619/X205834/X2407243 | LFL0.72-0.87 | 775.33 |
| 20 | X4364 | LFL0.72-0.87 | 778.33 |
| 21 | X62492 | LFL0.72-0.87 | 778.85 |
| 22 | X55871 | LFL0.72-0.87 | 779.34 |
| 23 | X23212 | LFL0.87-1.00 | 783.60 |
| 24 | X2L4g9p4 | LFL0.87-1.00 | 788.66 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Z.; Tian, X.; Ma, C.; Xia, Q.; Wang, B.; Chen, Q.; Sehgal, S.K.; Friebe, B.; Li, H.; Liu, W. Physical Mapping of Pm57, a Powdery Mildew Resistance Gene Derived from Aegilops searsii. Int. J. Mol. Sci. 2020, 21, 322. https://doi.org/10.3390/ijms21010322
Dong Z, Tian X, Ma C, Xia Q, Wang B, Chen Q, Sehgal SK, Friebe B, Li H, Liu W. Physical Mapping of Pm57, a Powdery Mildew Resistance Gene Derived from Aegilops searsii. International Journal of Molecular Sciences. 2020; 21(1):322. https://doi.org/10.3390/ijms21010322
Chicago/Turabian StyleDong, Zhenjie, Xiubin Tian, Chao Ma, Qing Xia, Beilin Wang, Qifan Chen, Sunish K. Sehgal, Bernd Friebe, Huanhuan Li, and Wenxuan Liu. 2020. "Physical Mapping of Pm57, a Powdery Mildew Resistance Gene Derived from Aegilops searsii" International Journal of Molecular Sciences 21, no. 1: 322. https://doi.org/10.3390/ijms21010322
APA StyleDong, Z., Tian, X., Ma, C., Xia, Q., Wang, B., Chen, Q., Sehgal, S. K., Friebe, B., Li, H., & Liu, W. (2020). Physical Mapping of Pm57, a Powdery Mildew Resistance Gene Derived from Aegilops searsii. International Journal of Molecular Sciences, 21(1), 322. https://doi.org/10.3390/ijms21010322