The Impact of Single- and Double-Strand DNA Breaks in Human Spermatozoa on Assisted Reproduction




Abstract
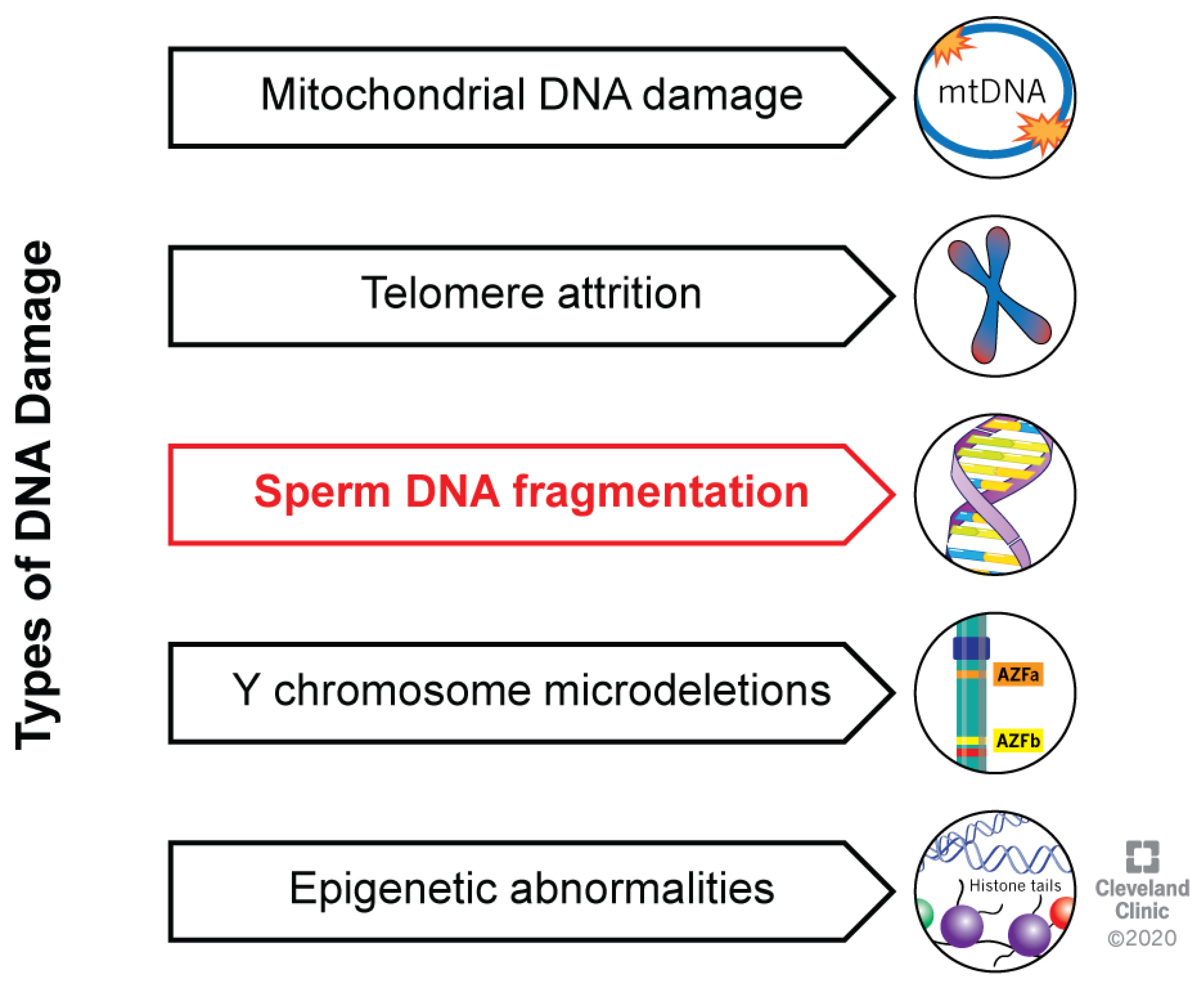
:1. Introduction
2. DNA Strand Breaks: Causes and Diagnosis
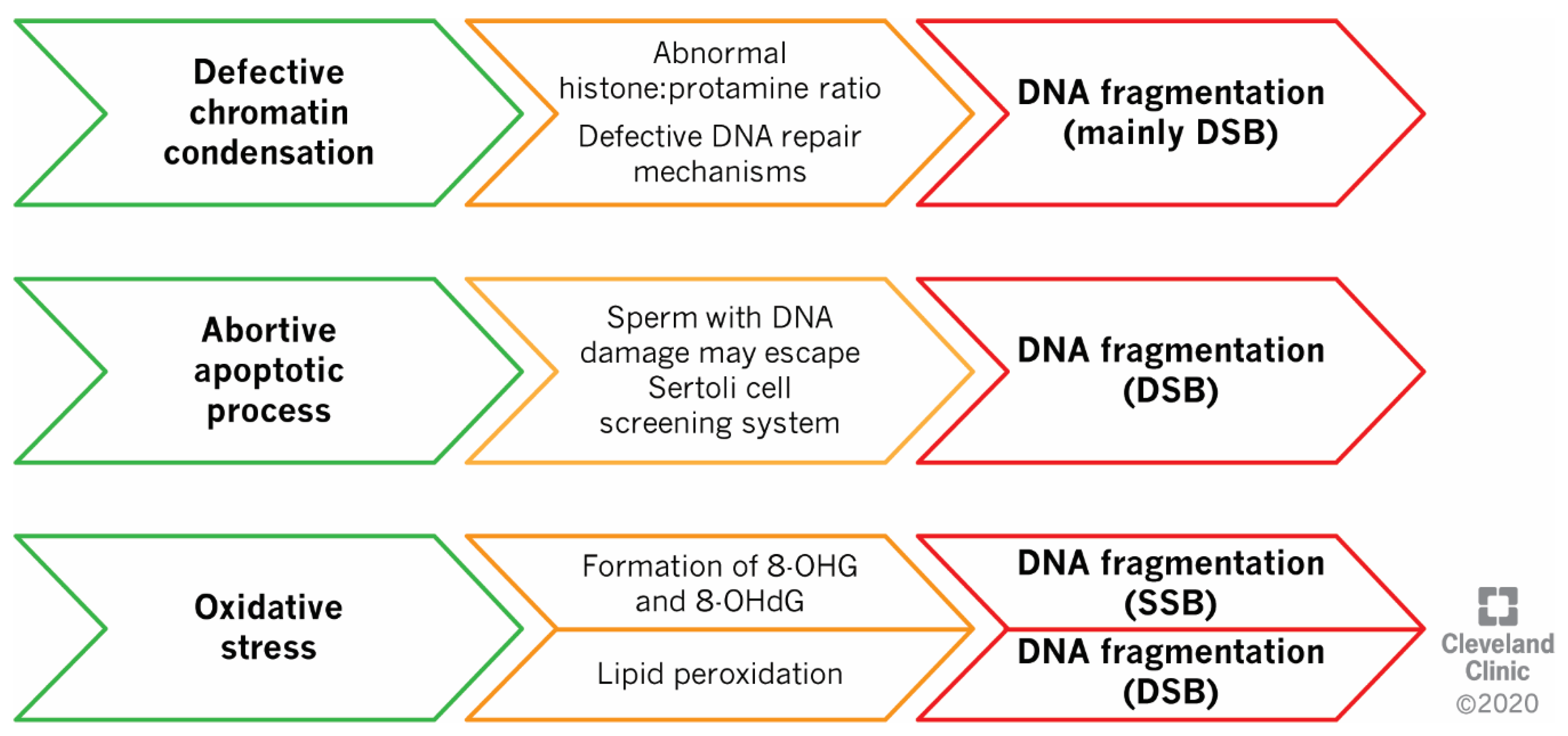
2.1. Defects in Sperm DNA Compaction
2.2. Abortive Apoptosis
2.3. Oxidative Stress
2.4. Clinical Tests for SDF Assessment
3. Association between DNA SSBs/DSBs and Reproductive Outcomes
3.1. Fertilization and Implantation Rates
3.2. Miscarriage Rate
3.3. Pregnancy Rate
3.4. Live Birth Rate
3.5. Brief Summary of the above Evidence
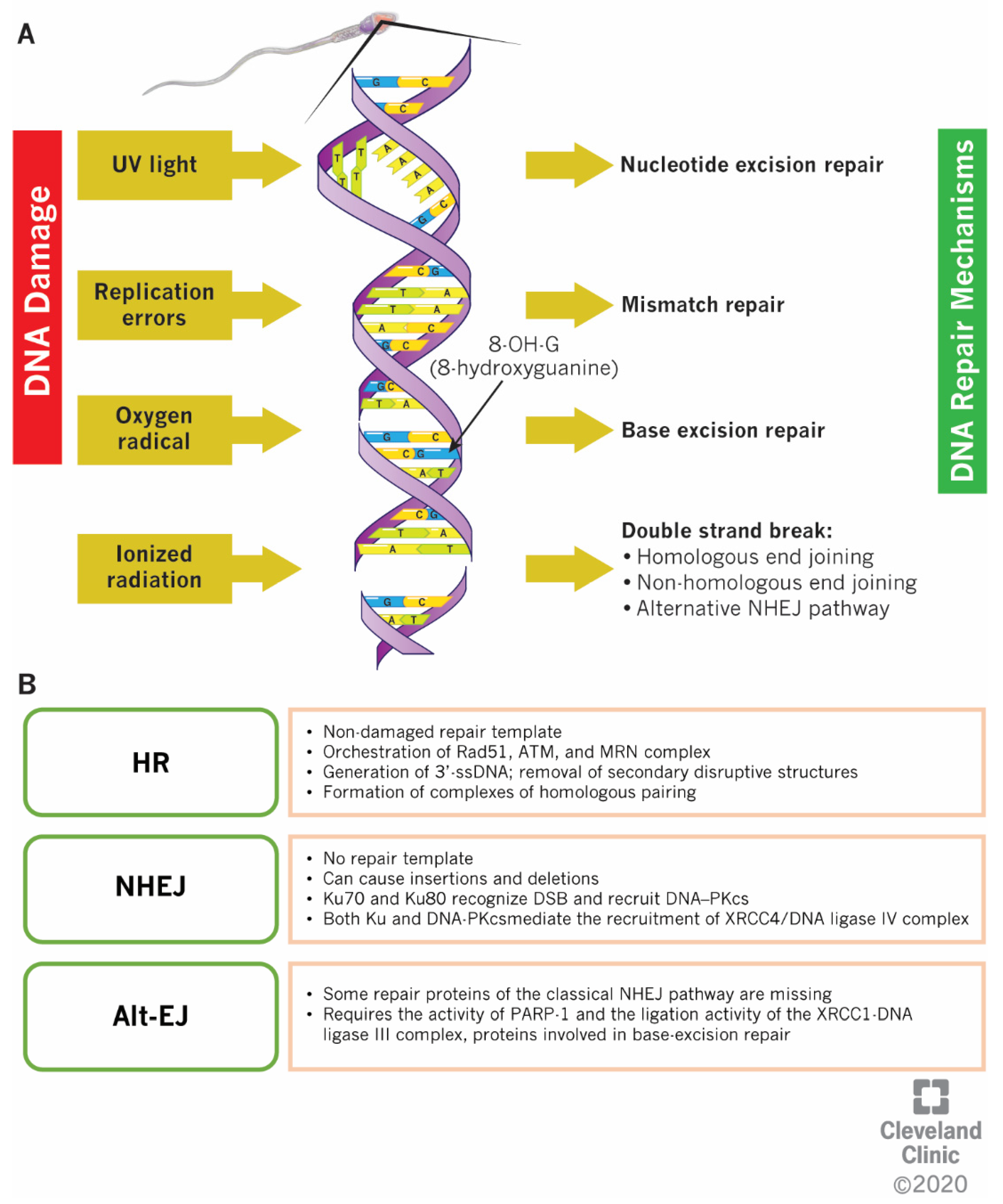
4. DNA Repair Mitigation Strategies: Differences between SSBs and DSBs
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ART | Assisted reproductive techniques |
| DGC | Density gradient centrifugation |
| DSBs | Double-strand DNA breaks |
| IVF | In vitro fertilization |
| SCD | Sperm chromatin dispersion |
| SCSA | Sperm chromatin structure assay |
| SSBs | Single-strand DNA breaks |
| SDF | Sperm DNA fragmentation |
| TUNEL | Terminal deoxynucleotidyl transferase nick end labelling |
References
- Ward, W.S. Function of sperm chromatin structural elements in fertilization and development. Mol. Hum. Reprod. 2009, 16, 30–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evgeni, E.; Charalabopoulos, K.; Asimakopoulos, B. Human Sperm DNA Fragmentation and its Correlation with Conventional Semen Parameters. J. Reprod. Infertil. 2014, 15, 2–14. [Google Scholar] [PubMed]
- Simon, L.; Murphy, K.; Shamsi, M.B.; Liu, L.; Emery, B.; I Aston, K.; Hotaling, J.; Carrell, D.T. Paternal influence of sperm DNA integrity on early embryonic development. Hum. Reprod. 2014, 29, 2402–2412. [Google Scholar] [CrossRef] [PubMed]
- Barroso, G.; Valdespin, C.; Vega, E.; Kershenovich, R.; Avila, R.; Avendaño, C.; Oehninger, S. Developmental sperm contributions: Fertilization and beyond. Fertil. Steril. 2009, 92, 835–848. [Google Scholar] [CrossRef]
- Lee, M.T.; Bonneau, A.R.; Giraldez, A.J. Zygotic genome activation during the maternal-to-zygotic transition. Annu. Rev. Cell Dev. Boil. 2014, 30, 581–613. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Selvam, M.K.P.; Baskaran, S.; Cho, C.-L. Sperm DNA damage and its impact on male reproductive health: A critical review for clinicians, reproductive professionals and researchers. Expert Rev. Mol. Diagn. 2019, 19, 443–457. [Google Scholar] [CrossRef]
- Garcia-Rodriguez, A.; Gosalvez, J.; Agarwal, A.; Roy, R.; Johnston, S.D. DNA Damage and Repair in Human Reproductive Cells. Int. J. Mol. Sci. 2018, 20, 31. [Google Scholar] [CrossRef] [Green Version]
- Sakkas, D.; Alvarez, J.G. Sperm DNA fragmentation: Mechanisms of origin, impact on reproductive outcome, and analysis. Fertil. Steril. 2010, 93, 1027–1036. [Google Scholar] [CrossRef]
- Aitken, R.; De Iuliis, G.; De Iuliis, G. On the possible origins of DNA damage in human spermatozoa. Mol. Hum. Reprod. 2009, 16, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.; Bronson, R.; Smith, T.B.; De Iuliis, G. The source and significance of DNA damage in human spermatozoa; a commentary on diagnostic strategies and straw man fallacies. Mol. Hum. Reprod. 2013, 19, 475–485. [Google Scholar] [CrossRef] [Green Version]
- Ward, W.S. DNA packaging and organization in mammalian spermatozoa: Comparison with somatic cells. Boil. Reprod. 1991, 44, 569–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrell, D.T.; Liu, L. Altered protamine 2 expression is uncommon in donors of known fertility, but common among men with poor fertilizing capacity, and may reflect other abnormalities of spermiogenesis. J. Androl. 2001, 22, 604–610. [Google Scholar] [PubMed]
- Simon, L.; Castillo, J.; Oliva, R.; E Lewis, S. Relationships between human sperm protamines, DNA damage and assisted reproduction outcomes. Reprod. Biomed. Online 2011, 23, 724–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Peiró, A.; Martínez-Heredia, J.; Oliver-Bonet, M.; Abad, C.; Amengual, M.J.; Navarro, J.; Jones, C.; Coward, K.; Gosalvez, J.; Benet, J. Protamine 1 to protamine 2 ratio correlates with dynamic aspects of DNA fragmentation in human sperm. Fertil. Steril. 2011, 95, 105–109. [Google Scholar] [CrossRef]
- Laberge, R.-M.; Boissonneault, G. On the Nature and Origin of DNA Strand Breaks in Elongating Spermatids1. Boil. Reprod. 2005, 73, 289–296. [Google Scholar] [CrossRef]
- Andrabi, S.M.H. Mammalian sperm chromatin structure and assessment of DNA fragmentation. J. Assist. Reprod. Genet. 2007, 24, 561–569. [Google Scholar] [CrossRef]
- Smith, A.; Haaf, T. DNA Nicks and Increased Sensitivity of DNA to Fluorescence In Situ End Labeling during Functional Spermiogenesis. Biotechniques 1998, 25, 496–502. [Google Scholar] [CrossRef]
- Kellner, U.; Rudolph, P.; Parwaresch, R. Human DNA-Topoisomerases - Diagnostic and Therapeutic Implications for Cancer. Onkologie 2000, 23, 424–430. [Google Scholar] [CrossRef]
- Hai, Y.; Hou, J.; Liu, Y.; Liu, Y.; Yang, H.; Li, Z.; He, Z. The roles and regulation of Sertoli cells in fate determinations of spermatogonial stem cells and spermatogenesis. Semin. Cell Dev. Boil. 2014, 29, 66–75. [Google Scholar] [CrossRef]
- Sakkas, D.; Seli, E.; Bizzaro, D.; Tarozzi, N.; Manicardi, G.C. Abnormal spermatozoa in the ejaculate: Abortive apoptosis and faulty nuclear remodelling during spermatogenesis. Reprod. Biomed. Online 2003, 7, 428–432. [Google Scholar] [CrossRef]
- Sakkas, D.; Mariethoz, E.; John, J.C.S. Abnormal Sperm Parameters in Humans Are Indicative of an Abortive Apoptotic Mechanism Linked to the Fas-Mediated Pathway. Exp. Cell Res. 1999, 251, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Marin, C.; Gosalvez, J.; Roy, R. Types, Causes, Detection and Repair of DNA Fragmentation in Animal and Human Sperm Cells. Int. J. Mol. Sci. 2012, 13, 14026–14052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakkas, D.; Seli, E.; Manicardi, G.C.; Nijs, M.; Ombelet, W.; Bizzaro, D. The presence of abnormal spermatozoa in the ejaculate: Did apoptosis fail? Hum. Fertil. 2004, 7, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Betteridge, D.J. What is oxidative stress? Metabolism 2000, 49, 3–8. [Google Scholar] [CrossRef]
- Badouard, C.; Ménézo, Y.; Panteix, G.; Ravanat, J.-L.; Douki, T.; Cadet, J.; Favier, A. Determination of new types of DNA lesions in human sperm. Zygote 2008, 16, 9–13. [Google Scholar] [CrossRef]
- Cortés-Gutiérrez, E.I.; Fernández, J.; Dávila-Rodríguez, M.I.; López-Fernández, C.; Gosalvez, J. Two-Tailed Comet Assay (2T-Comet): Simultaneous Detection of DNA Single and Double Strand Breaks. In Advanced Structural Safety Studies; Springer Science and Business Media LLC: Berlin, Germany, 2017; Volume 1560, pp. 285–293. [Google Scholar]
- Ribas-Maynou, J.; García-Peiró, A.; Encinas, A.F.; Abad, C.; Amengual, M.J.; Prada, E.; Navarro, J.; Benet, J. Comprehensive analysis of sperm DNA fragmentation by five different assays: TUNEL assay, SCSA, SCD test and alkaline and neutral Comet assay. Andrology 2013, 1, 715–722. [Google Scholar] [CrossRef]
- Valdiglesias, V.; Giunta, S.; Fenech, M.; Neri, M.; Bonassi, S. γH2AX as a marker of DNA double strand breaks and genomic instability in human population studies. Mutat. Res. Mutat. Res. 2013, 753, 24–40. [Google Scholar] [CrossRef]
- Gupta, S.; Sharma, R.; Agarwal, A. Inter-and Intra-Laboratory Standardization of TUNEL Assay for Assessment of Sperm DNA Fragmentation. Curr. Protoc. Toxicol. 2017, 74, 16.11.1–16.11.22. [Google Scholar] [CrossRef]
- Evenson, D.P.; Jost, L.; Marshall, D.; Zinaman, M.; Clegg, E.; Purvis, K.; De Angelis, P.; Claussen, O. Utility of the sperm chromatin structure assay as a diagnostic and prognostic tool in the human fertility clinic. Hum. Reprod. 1999, 14, 1039–1049. [Google Scholar] [CrossRef]
- Fernández, J.; Muriel, L.; Rivero, M.T.; Goyanes, V.; Vazquez, R.; Alvarez, J.G. The sperm chromatin dispersion test: A simple method for the determination of sperm DNA fragmentation. J. Androl. 2003, 24, 59–66. [Google Scholar]
- Ribas-Maynou, J.; Benet, J. Single and Double Strand Sperm DNA Damage: Different Reproductive Effects on Male Fertility. Genes 2019, 10, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enciso, M.; Sarasa, J.; Agarwal, A.; Fernández, J.; Gosalvez, J. A two-tailed Comet assay for assessing DNA damage in spermatozoa. Reprod. Biomed. Online 2009, 18, 609–616. [Google Scholar] [CrossRef]
- Ribas-Maynou, J.; García-Peiró, A.; Abad, C.; Amengual, M.J.; Navarro, J.; Benet, J. Alkaline and neutral Comet assay profiles of sperm DNA damage in clinical groups. Hum. Reprod. 2012, 27, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Singh, K.; Almasan, A. Histone H2AX Phosphorylation: A Marker for DNA Damage. Adv. Struct. Saf. Stud. 2012, 920, 613–626. [Google Scholar] [CrossRef]
- Santi, D.; Spaggiari, G.; Simoni, M. Sperm DNA Fragmentation Index as a Promising Predictive Tool for Male Infertility Diagnosis and Treatment Management–Meta-Analyses. Reprod. Biomed. Online 2018, 37, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Deng, C.; Li, T.; Xie, Y.; Guo, Y.; Yang, Q.-Y.; Liang, X.; Deng, C.-H.; Liu, G.-H. Sperm DNA fragmentation index influences assisted reproductive technology outcome: A systematic review and meta-analysis combined with a retrospective cohort study. Andrologia 2019, 51, e13263. [Google Scholar] [CrossRef]
- Simon, L.; Brunborg, G.; Stevenson, M.; Lutton, D.; McManus, J.; E Lewis, S. Clinical significance of sperm DNA damage in assisted reproduction outcome. Hum. Reprod. 2010, 25, 1594–1608. [Google Scholar] [CrossRef] [Green Version]
- Simon, L.; Lutton, D.; McManus, J.; E Lewis, S. Sperm DNA damage measured by the alkaline Comet assay as an independent predictor of male infertility and in vitro fertilization success. Fertil. Steril. 2011, 95, 652–657. [Google Scholar] [CrossRef]
- Casanovas, A.; Ribas-Maynou, J.; Lara-Cerrillo, S.; Jimenez-Macedo, A.R.; Hortal, O.; Benet, J.; Carrera, J.; García-Peiró, A. Double-stranded sperm DNA damage is a cause of delay in embryo development and can impair implantation rates. Fertil. Steril. 2019, 111, 699–707.e1. [Google Scholar] [CrossRef]
- Ribas-Maynou, J.; García-Peiró, A.; Encinas, A.F.; Amengual, M.J.; Prada, E.; Cortés, P.; Navarro, J.; Benet, J. Double Stranded Sperm DNA Breaks, Measured by Comet Assay, Are Associated with Unexplained Recurrent Miscarriage in Couples without a Female Factor. PLoS ONE 2012, 7, e44679. [Google Scholar] [CrossRef] [Green Version]
- Garolla, A.; Cosci, I.; Bertoldo, A.; Sartini, B.; Boudjema, E.; Foresta, C. DNA double strand breaks in human spermatozoa can be predictive for assisted reproductive outcome. Reprod. Biomed. Online 2015, 31, 100–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, L.; I Aston, K.; Emery, B.R.; Hotaling, J.; Carrell, D.T. Sperm DNA damage output parameters measured by the alkaline Comet assay and their importance. Andrologia 2016, 49, 49. [Google Scholar] [CrossRef] [PubMed]
- Garolla, A.; Ghezzi, M.; Cosci, I.; Sartini, B.; Bottacin, A.; Engl, B.; Di Nisio, A.; Foresta, C. FSH treatment in infertile males candidate to assisted reproduction improved sperm DNA fragmentation and pregnancy rate. Endocrine 2016, 56, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Simon, L.; Proutski, I.; Stevenson, M.; Jennings, D.; McManus, J.; Lutton, D.; E Lewis, S. Sperm DNA damage has a negative association with live-birth rates after IVF. Reprod. Biomed. Online 2013, 26, 68–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coban, O.; Serdarogullari, M.; Yarkiner, Z.; Serakinci, N. Investigating the level of DNA double-strand break in human spermatozoa and its relation to semen characteristics and IVF outcome using phospho-histone H2AX antibody as a biomarker. Andrologia 2019, 8, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Reichmann, J.; Nijmeijer, B.; Hossain, M.J.; Eguren, M.; Schneider, I.; Politi, A.Z.; Roberti, M.J.; Hufnagel, L.; Hiiragi, T.; Ellenberg, J. Dual-spindle formation in zygotes keeps parental genomes apart in early mammalian embryos. Science 2018, 361, 189–193. [Google Scholar] [CrossRef] [Green Version]
- Berg, J.V.D.; Manjón, A.G.; Kielbassa, K.; Feringa, F.M.; Freire, R.; Medema, R.H. A limited number of double-strand DNA breaks is sufficient to delay cell cycle progression. Nucleic Acids Res. 2018, 46, 10132–10144. [Google Scholar] [CrossRef]
- Belloc, S.; Benkhalifa, M.; Cohen-Bacrie, M.; Dalleac, A.; Amar, E.; Zini, A. Sperm deoxyribonucleic acid damage in normozoospermic men is related to age and sperm progressive motility. Fertil. Steril. 2014, 101, 1588–1593. [Google Scholar] [CrossRef]
- García-Ferreyra, J.; Villegas, L.; Obst, R.R.; Obst, P.Z.; Hilario, R.; Casafranca, G.; Dueñas-Chacón, J.; García-Ferreyra, L.V.J. Sperm DNA Fragmentation is Significantly Increased in Those Men with Morphologically Abnormal Spermatozoa. J. Fertil. Vitr. IVF-Worldwide Reprod. Med. Genet. Stem Cell Boil. 2014, 2, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Gunes, S.; Sertyel, S. Sperm DNA Damage and Oocyte Repair Capability. In A Clinician’s Guide to Sperm DNA and Chromatin Damage; Springer Science and Business Media LLC: Berlin, Germany, 2018; pp. 321–346. [Google Scholar]
- Ahmadi, A. Fertilizing ability of DNA-damaged spermatozoa. J. Exp. Zool. 1999, 284, 696–704. [Google Scholar] [CrossRef]
- Boiteux, S.; Radicella, J.P. Base excision repair of 8-hydroxyguanine protects DNA from endogenous oxidative stress. Biochimie 1999, 81, 59–67. [Google Scholar] [CrossRef]
- Güneş, S.; Al-Sadaan, M.; Agarwal, A.; Alsadaanasan, M. Spermatogenesis, DNA damage and DNA repair mechanisms in male infertility. Reprod. Biomed. Online 2015, 31, 309–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-S.; Krasieva, T.B.; Kurumizaka, H.; Chen, D.J.; Taylor, A.M.R.; Yokomori, K. Independent and sequential recruitment of NHEJ and HR factors to DNA damage sites in mammalian cells. J. Cell Boil. 2005, 170, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Brandsma, I.; Van Gent, D.C. Pathway choice in DNA double strand break repair: Observations of a balancing act. Genome Integr. 2012, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Steinfeld, J.B.; Liang, F.; Chen, X.; Maranon, D.G.; Ma, C.J.; Kwon, Y.; Rao, T.; Wang, W.; Sheng, C.; et al. BRCA1–BARD1 promotes RAD51-mediated homologous DNA pairing. Nature 2017, 550, 360–365. [Google Scholar] [CrossRef]
- Forget, A.L.; Kowalczykowski, S.C. Single-molecule imaging brings Rad51 nucleoprotein filaments into focus. Trends Cell Boil. 2010, 20, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Scully, R.; Chen, J.; Plug, A.; Xiao, Y.; Weaver, D.; Feunteun, J.; Ashley, T.; Livingston, D.M. Association of BRCA1 with Rad51 in Mitotic and Meiotic Cells. Cell 1997, 88, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Maréchal, A.; Zou, L. DNA Damage Sensing by the ATM and ATR Kinases. Cold Spring Harb. Perspect. Boil. 2013, 5, a012716. [Google Scholar] [CrossRef]
- Yoshida, K.; Miki, Y. Role of BRCA1 and BRCA2 as regulators of DNA repair, transcription, and cell cycle in response to DNA damage. Cancer Sci. 2004, 95, 866–871. [Google Scholar] [CrossRef]
- Lee, J.-H. ATM Activation by DNA Double-Strand Breaks Through the Mre11-Rad50-Nbs1 Complex. Science 2005, 308, 551–554. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Chaudhuri, A.R.; Lopes, M.; Costanzo, V. Rad51 protects nascent DNA from Mre11-dependent degradation and promotes continuous DNA synthesis. Nat. Struct. Mol. Boil. 2010, 17, 1305–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Liu, Y.; Wu, X.; Shell, S.M. Functions of human replication protein A (RPA): From DNA replication to DNA damage and stress responses. J. Cell. Physiol. 2006, 208, 267–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rulten, S.L.; Grundy, G.J. Non-homologous end joining: Common interaction sites and exchange of multiple factors in the DNA repair process. BioEssays 2017, 39, 1600209. [Google Scholar] [CrossRef] [PubMed]
- A Goodarzi, A.; Yu, Y.; Riballo, E.; Douglas, P.; A Walker, S.; Ye, R.; Härer, C.; Marchetti, C.; Morrice, N.; Jeggo, P.A.; et al. DNA-PK autophosphorylation facilitates Artemis endonuclease activity. EMBO J. 2006, 25, 3880–3889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riballo, E.; Woodbine, L.; Stiff, T.; Walker, S.A.; Goodarzi, A.A.; Jeggo, P.A. XLF-Cernunnos promotes DNA ligase IV-XRCC4 re-adenylation following ligation. Nucleic Acids Res. 2008, 37, 482–492. [Google Scholar] [CrossRef]
- Rothkamm, K.; Thompson, L.H.; Löbrich, M. Pathways of DNA Double-Strand Break Repair during the Mammalian Cell Cycle. Mol. Cell. Boil. 2003, 23, 5706–5715. [Google Scholar] [CrossRef] [Green Version]
- Myler, L.R.; Gallardo, I.F.; Soniat, M.M.; Deshpande, R.A.; Gonzalez, X.B.; Kim, Y.; Paull, T.T.; Finkelstein, I.J. Single-Molecule Imaging Reveals How Mre11-Rad50-Nbs1 Initiates DNA Break Repair. Mol. Cell 2017, 67, 891–898.e4. [Google Scholar] [CrossRef] [Green Version]
- Langerak, P.; Mejia-Ramirez, E.; Limbo, O.; Russell, P. Release of Ku and MRN from DNA Ends by Mre11 Nuclease Activity and Ctp1 Is Required for Homologous Recombination Repair of Double-Strand Breaks. PLoS Genet. 2011, 7, e1002271. [Google Scholar] [CrossRef] [Green Version]
- Teixeira-Silva, A.; Saada, A.A.; Hardy, J.; Iraqui, I.; Nocente, M.C.; Fréon, K.; Lambert, S. The end-joining factor Ku acts in the end-resection of double strand break-free arrested replication forks. Nat. Commun. 2017, 8, 1982. [Google Scholar] [CrossRef] [Green Version]
- Boboila, C.; Jankovic, M.; Yan, C.T.; Wang, J.H.; Wesemann, D.R.; Zhang, T.; Fazeli, A.; Feldman, L.; Nussenzweig, A.; Nussenzweig, M.; et al. Alternative end-joining catalyzes robust IgH locus deletions and translocations in the combined absence of ligase 4 and Ku70. Proc. Natl. Acad. Sci. USA 2010, 107, 3034–3039. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.T.; Boboila, C.; Souza, E.K.; Franco, S.; Hickernell, T.R.; Murphy, M.; Gumaste, S.; Geyer, M.B.; Zarrin, A.A.; Manis, J.P.; et al. IgH class switching and translocations use a robust non-classical end-joining pathway. Nature 2007, 449, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Nagaria, P.; Rassool, F.V. Alternative Non-Homologous End-Joining: Mechanisms and Targeting Strategies in Cancer. In Targeting the DNA Damage Response for Anti-Cancer Therapy. Cancer Drug Discovery and Development; Pollard, J., Curtin, N., Eds.; Humana Press: Cham, Switzerland, 2018; pp. 375–400. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Test | Principle | Result | Type of Damage Detected |
|---|---|---|---|
| TUNEL | Tagged nucleotides are linked to the DNA break | Sperm with fragmented DNA shows fluorescence | SSBs/DSBs |
| Comet assay | Fragmented DNA is separated electrophoretically | Shape of comet tail indirectly reflects the severity of DNA damage | Neutral Comet: DSBs Alkaline Comet: mostly SSBs |
| SCSA | The susceptibility of sperm DNA to denaturation is measured based on the metachromatic properties of acridine orange | Double-stranded DNA fluoresces green, denatured DNA fluoresces orange-red | SSBs/DSBs |
| SCD test/Halo Test | Chromatin dispersion is microscopically observed after denaturation | DNA integrity results in the generation of a characteristic halo of dispersed DNA loops, while no halo is reported in case of DNA damage | SSBs/DSBs |
| γH2AX | Antibodies are used to detect γH2AX, a marker of DSB | γH2AX levels correlate with increased levels of DSBs | DSBs |
| Reproductive Outcome | Study Description | Results | Author, Year |
|---|---|---|---|
| Fertilization Rate (FR) | 360 patients attending IVF (n = 230) and ICSI (n = 130); Alkaline Comet assay to evaluate SSBs in the native semen and after DGC | In IVF, FR decreased depending on the percentage of DGC-selected sperm showing SSBs; no difference in ICSI | Simon, 2010 [38] |
| 75 couples (IVF) and 28 fertile donors; Alkaline Comet assay to assess SSBs in the native semen and after DGC | In IVF, FR was negatively associated to the percentage of sperm with SSBs when both native and DGC sperm were used | Simon, 2011 [39] | |
| Semen sample used for ICSI was analyzed by two-tailed Comet assay | In ICSI, no difference in FR in case of high percentage of sperm with SSBs and DSBs | Casanovas, 2019 [40] | |
| Implantation Rate (IR) | 215 infertile men undergoing IVF/ICSI; samples were classified based on the percentage of SSBs in “low damage”, “intermediate damage” and “high damage” | In the native semen, IR decreased depending on the percentage of sperm with SSBs | Simon, 2014 [3] |
| Semen sample used for ICSI was analyzed by two-tailed Comet assay | In ICSI, reduced IR in case of high sperm percentage with DSBs | Casanovas, 2019 [40] | |
| Miscarriage Rate | 25 fertile men and 20 patients suffering for recurrent pregnancy loss SDF were analyzed by using two-tailed Comet assay, SCD test and pulsed-field gel electrophoresis (PFGE) | Higher percentage of sperm with SSBs and DSBs is reported in unexplained recurrent pregnancy loss patients than fertile donors | Ribas-Maynou, 2012 [41] |
| Pregnancy Rate (PR) | 360 patients attending IVF (n = 230) and ICSI (n = 130); Alkaline Comet to evaluate SSBs in the native semen and after DGC | In IVF, non-pregnant couples showed higher percentage of sperm with SSBs than pregnant couples in both native and DGC-selected sperm; cut-offs equal to 56% and 44% for percentage of sperm SSBs in native and DGC-selected semen, respectively, to predict a clinical pregnancy in ART | Simon, 2010 [38] |
| 75 couples (IVF) and 28 fertile donors; Alkaline Comet assay to assess SSBs in the native semen and after DGC | High percentage of sperm with SSBs (>52% for native semen; >46% for DGC-selected sperm) was associated with decreased pregnancy rate | Simon, 2011 [39] | |
| 25 fertile men and 20 patients suffering for recurrent pregnancy loss DF were analyzed by using two-tailed Comet assay, SCD test and pulsed-field gel electrophoresis (PFGE) | Alkaline Comet assay might better predict pregnancy than neutral Comet assay | Ribas-Maynou, 2012 [41] | |
| 215 infertile men undergoing IVF/ICSI Samples were classified based on the percentage of SSBs in “low damage”, “intermediate damage” and “high damage” | Reduced clinical PR in couples with high percentage of sperm having SSBs | Simon, 2014 [3] | |
| 100 infertile men undergoing ICSI and 61 fertile men DSBs were measured by γH2AX | Reduced percentage of sperm with DSBs in infertile patients who achieved a pregnancy compared to those infertile who failed | Garolla, 2015 [42] | |
| 47 fertile donors and 238 infertile couples; Alkaline Comet assay to evaluate the presence of SSBs | Alkaline Comet predicted clinical pregnancy with moderate sensitivity and specificity at a cut-off value of 52% | Simon, 2017 [43] | |
| 166 infertile male partners of couples undergoing ICSI 84 patients were receiving FSH treatment and 82 refused treatment (controls); DSBs were measured by γH2AX | Infertile patients undergoing FSH-treatment and ICSI showed reduced percentage of sperm with DSBs when the pregnancy was achieved | Garolla, 2017 [44] | |
| Live Birth Rate (LBR) | 339 couples attending IVF (n = 203) and ICSI (n = 136); Alkaline Comet assay to evaluate SSBs in native semen and after DGC | Following IVF, LBR was 33% and 13% in couples with <25% and >50% SSBs, respectively; no difference after ICSI | Simon, 2013 [45] |
| 60 ART cycles (52 autologous and 8 donors); DSBs assessed by detection of histone γH2AX | In IVF, live birth rate was associated with lower percentage of sperm with DSBs | Coban, 2019 [46] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agarwal, A.; Barbăroșie, C.; Ambar, R.; Finelli, R. The Impact of Single- and Double-Strand DNA Breaks in Human Spermatozoa on Assisted Reproduction. Int. J. Mol. Sci. 2020, 21, 3882. https://doi.org/10.3390/ijms21113882
Agarwal A, Barbăroșie C, Ambar R, Finelli R. The Impact of Single- and Double-Strand DNA Breaks in Human Spermatozoa on Assisted Reproduction. International Journal of Molecular Sciences. 2020; 21(11):3882. https://doi.org/10.3390/ijms21113882
Chicago/Turabian StyleAgarwal, Ashok, Cătălina Barbăroșie, Rafael Ambar, and Renata Finelli. 2020. "The Impact of Single- and Double-Strand DNA Breaks in Human Spermatozoa on Assisted Reproduction" International Journal of Molecular Sciences 21, no. 11: 3882. https://doi.org/10.3390/ijms21113882
APA StyleAgarwal, A., Barbăroșie, C., Ambar, R., & Finelli, R. (2020). The Impact of Single- and Double-Strand DNA Breaks in Human Spermatozoa on Assisted Reproduction. International Journal of Molecular Sciences, 21(11), 3882. https://doi.org/10.3390/ijms21113882