Expression of HTRA Genes and Its Association with Microsatellite Instability and Survival of Patients with Colorectal Cancer

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
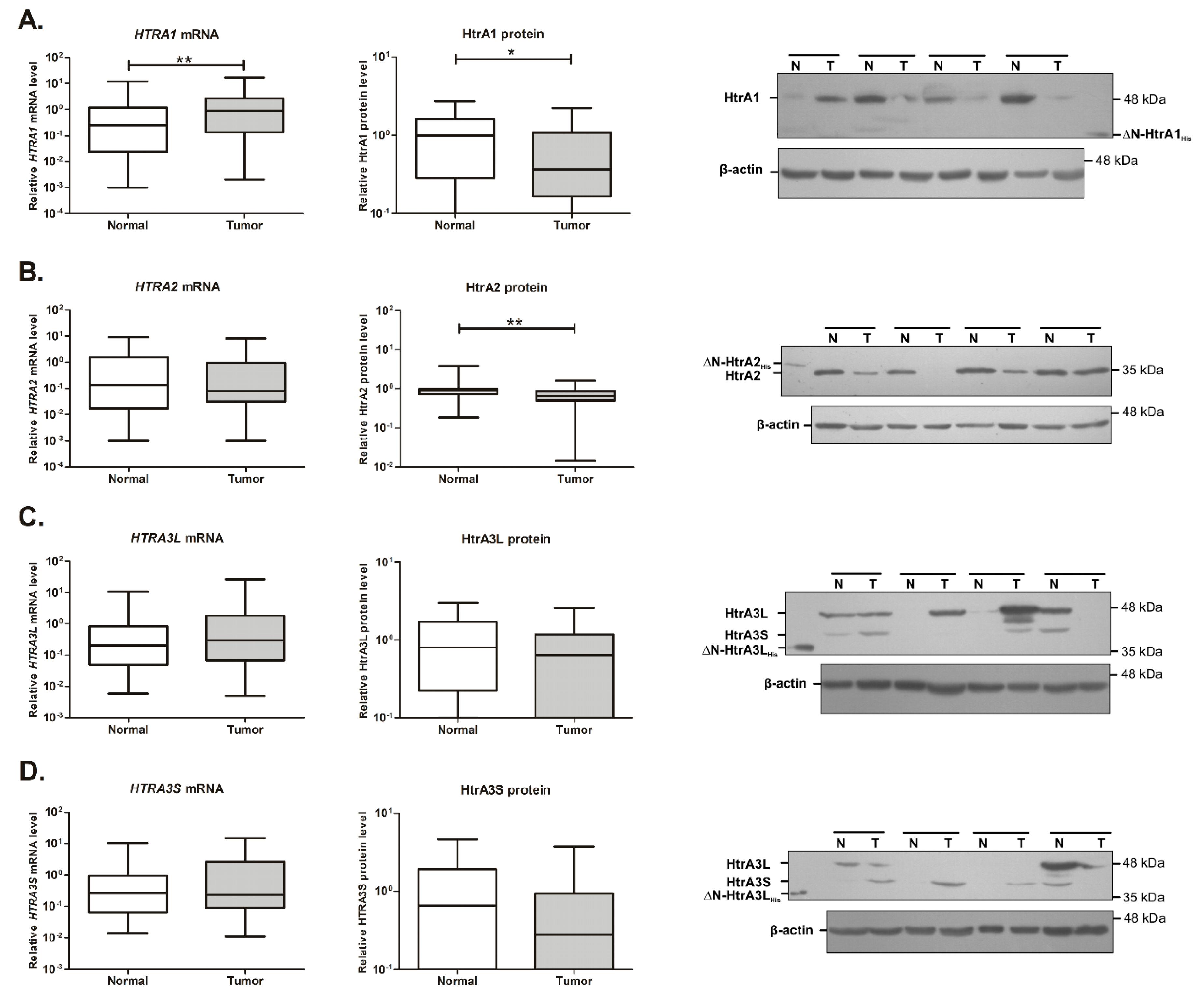
2.1. Expression of the HTRA1 and HTRA2 Genes Is Changed in Colorectal Tumors
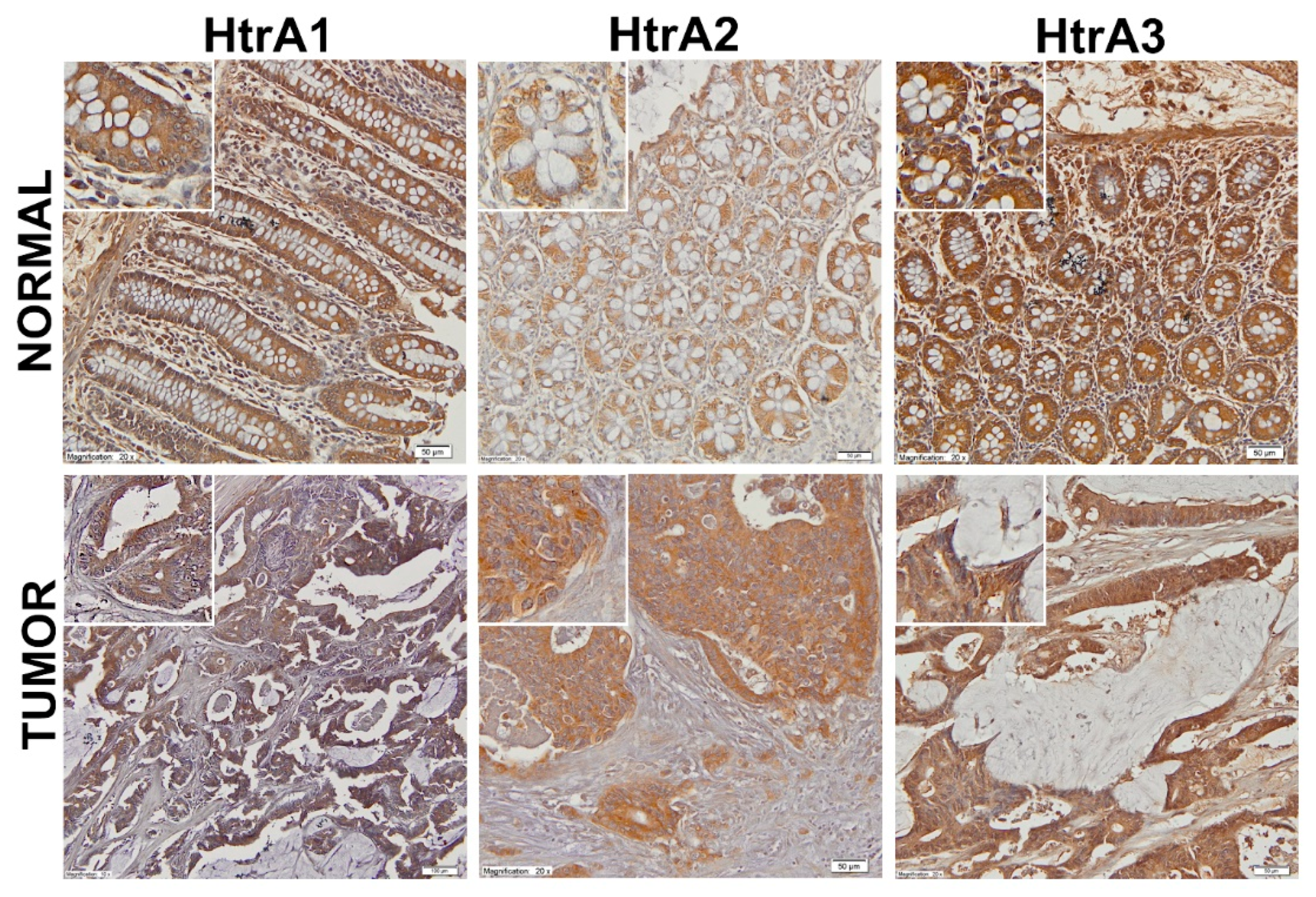
2.2. Location of the HtrA Proteins in CRC Tissue and Colorectal Mucosa
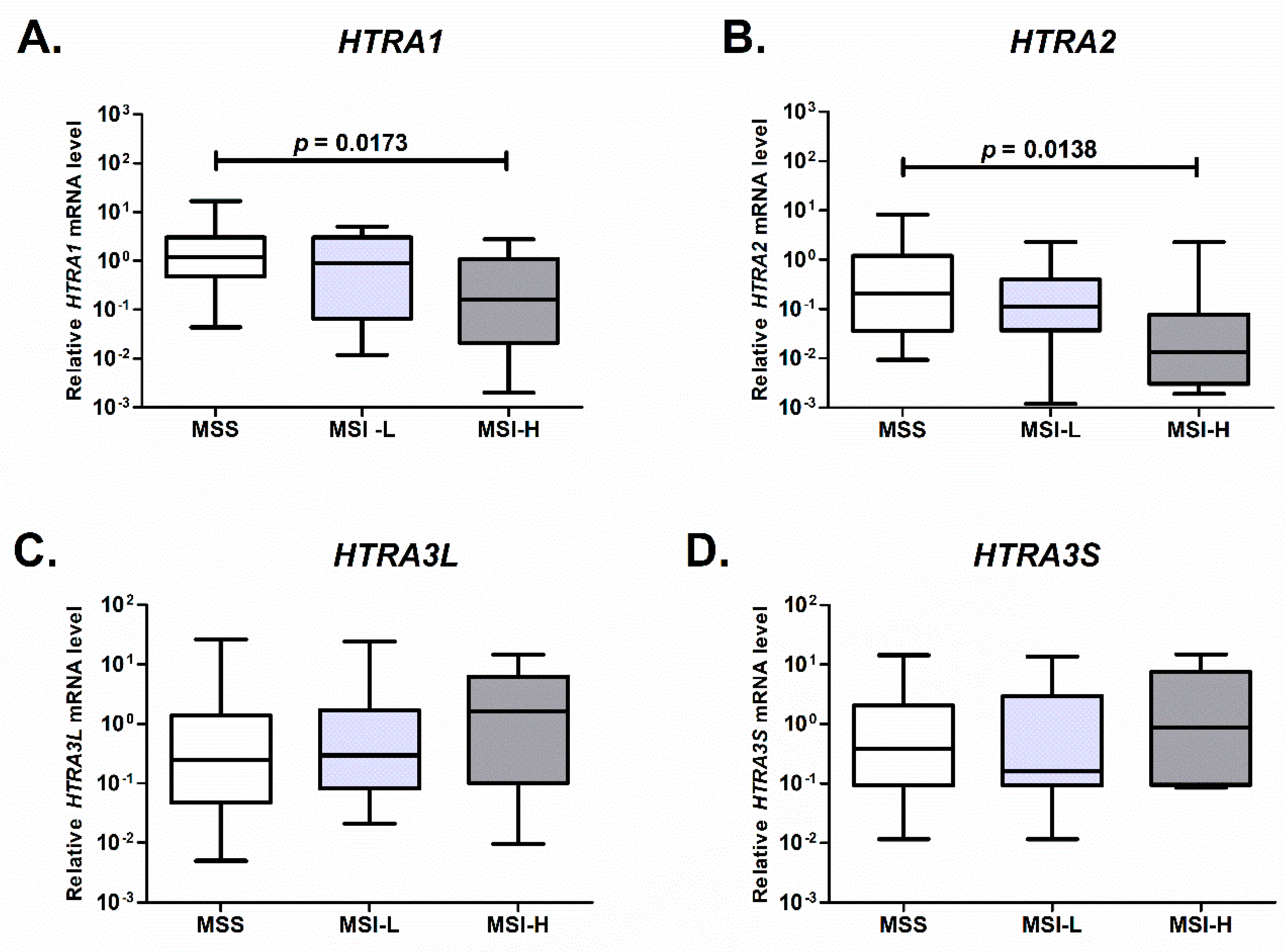
2.3. Expression of HTRA1 and HTRA2 Is Associated with Microsatellite Status of the Colorectal Tumor
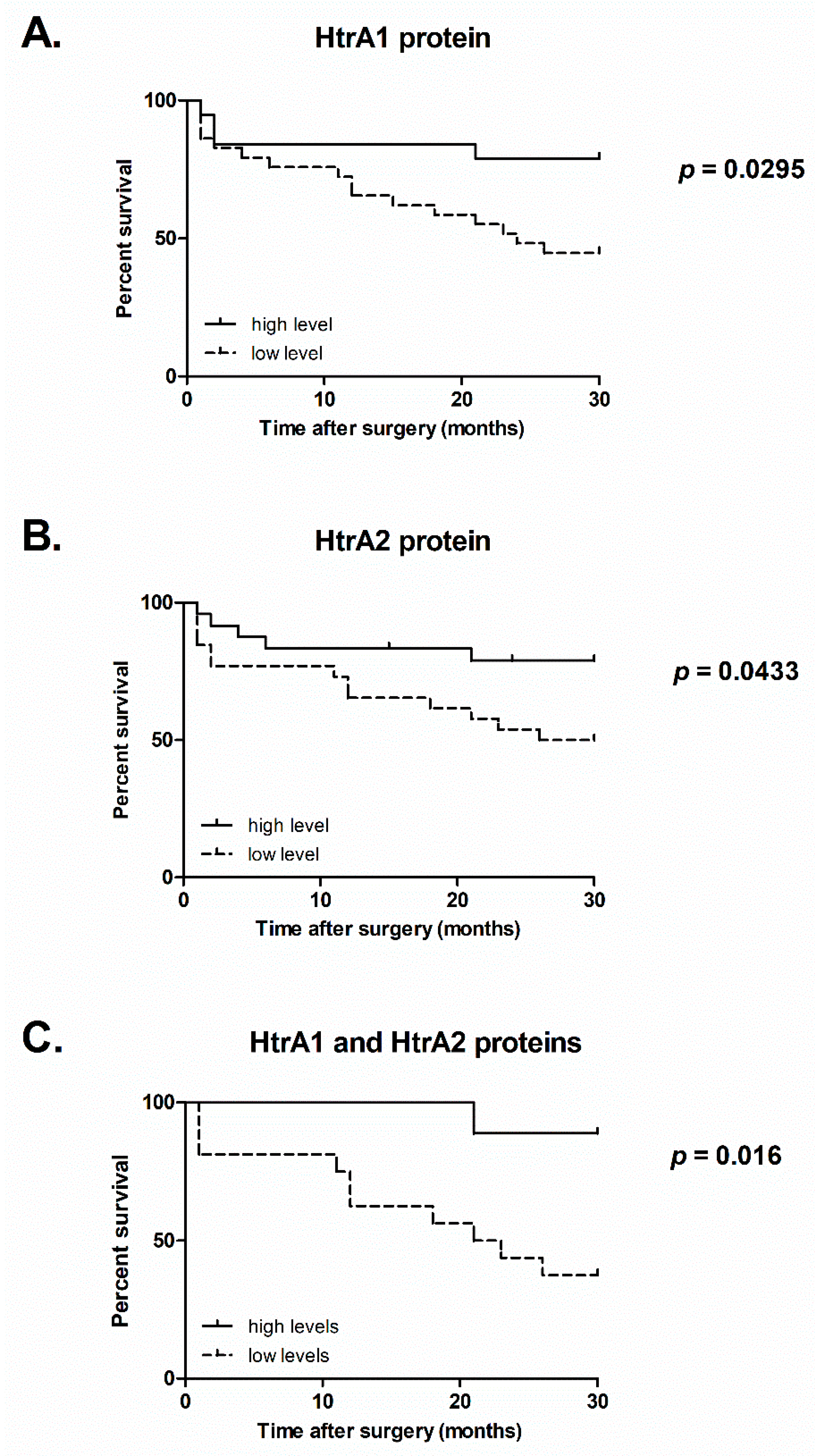
2.4. Survival of CRC Patients Correlates with the Levels of HtrA1 and HtrA2 Proteins
3. Discussion
4. Materials and Methods
4.1. Patients and Specimens
4.2. RNA Isolation and cDNA Synthesis
4.3. Real-Time PCR
4.4. Preparation of Tissue Extracts
4.5. Protein Assay and Electrophoresis
4.6. Western Blotting
4.7. Immunohistochemical Staining
4.8. Microsatellite Instability Typing by High-Resolution-Melting PCR
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global patterns and trends in colorectal cancer incidence and mortality. Gut 2017, 66, 683–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuipers, E.J.; Grady, W.M.; Lieberman, D.; Seufferlein, T.; Sung, J.J.; Boelens, P.G.; van de Velde, C.J.H.; Watanabe, T. Colorectal cancer. Nat. Rev. Dis. Primer 2015, 1, 15065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenner, H.; Kloor, M.; Pox, C.P. Colorectal cancer. Lancet Lond. Engl. 2014, 383, 1490–1502. [Google Scholar] [CrossRef]
- Mundade, R.; Imperiale, T.F.; Prabhu, L.; Loehrer, P.J.; Lu, T. Genetic pathways, prevention, and treatment of sporadic colorectal cancer. Oncoscience 2014, 1, 400–406. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-J.; Wang, A.-Q.; Chen, Q.-Q. DNA methylation assay for colorectal carcinoma. Cancer Biol. Med. 2017, 14, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Pancione, M.; Remo, A.; Colantuoni, V. Genetic and epigenetic events generate multiple pathways in colorectal cancer progression. Pathol. Res. Int. 2012, 2012, 509348. [Google Scholar] [CrossRef] [Green Version]
- Zoratto, F.; Rossi, L.; Verrico, M.; Papa, A.; Basso, E.; Zullo, A.; Tomao, L.; Romiti, A.; Lo Russo, G.; Tomao, S. Focus on genetic and epigenetic events of colorectal cancer pathogenesis: Implications for molecular diagnosis. Tumor Biol. 2014, 35, 6195–6206. [Google Scholar] [CrossRef]
- Zurawa-Janicka, D.; Wenta, T.; Jarzab, M.; Skorko-Glonek, J.; Glaza, P.; Gieldon, A.; Ciarkowski, J.; Lipinska, B. Structural insights into the activation mechanisms of human HtrA serine proteases. Arch. Biochem. Biophys. 2017, 621, 6–23. [Google Scholar] [CrossRef]
- Altobelli, E.; Marzioni, D.; Lattanzi, A.; Angeletti, P.M. HtrA1: Its future potential as a novel biomarker for cancer. Oncol. Rep. 2015, 34, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Skorko-Glonek, J.; Zurawa-Janicka, D.; Koper, T.; Jarzab, M.; Figaj, D.; Glaza, P.; Lipinska, B. HtrA protease family as therapeutic targets. Curr. Pharm. Des. 2013, 19, 977–1009. [Google Scholar] [CrossRef]
- Tiaden, A.N.; Richards, P.J. The emerging roles of HTRA1 in musculoskeletal disease. Am. J. Pathol. 2013, 182, 1482–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, J.; Campioni, M.; Shridhar, V.; Baldi, A. HtrA serine proteases as potential therapeutic targets in cancer. Curr. Cancer Drug Targets 2009, 9, 451–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altobelli, E.; Angeletti, P.M.; Morroni, M.; Profeta, V.F. HtrA1 as a promising tissue marker in cancer: A meta-analysis. BMC Cancer 2018, 18, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldi, A.; Mottolese, M.; Vincenzi, B.; Campioni, M.; Mellone, P.; Di Marino, M.; di Crescenzo, V.G.; Visca, P.; Menegozzo, S.; Spugnini, E.P.; et al. The serine protease HtrA1 is a novel prognostic factor for human mesothelioma. Pharmacogenomics 2008, 9, 1069–1077. [Google Scholar] [CrossRef]
- Catalano, V.; Mellone, P.; d’Avino, A.; Shridhar, V.; Staccioli, M.P.; Graziano, F.; Giordani, P.; Rossi, D.; Baldelli, A.M.; Alessandroni, P.; et al. HtrA1, a potential predictor of response to cisplatin-based combination chemotherapy in gastric cancer. Histopathology 2011, 58, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Chien, J.; Aletti, G.; Baldi, A.; Catalano, V.; Muretto, P.; Keeney, G.L.; Kalli, K.R.; Staub, J.; Ehrmann, M.; Cliby, W.A.; et al. Serine protease HtrA1 modulates chemotherapy-induced cytotoxicity. J. Clin. Investig. 2006, 116, 1994–2004. [Google Scholar] [CrossRef] [Green Version]
- Franco, R.; Collina, F.; Di Bonito, M.; Botti, G.; Montanaro, D.; Di Maio, L.; Vincenzi, B.; Landi, G.; D’Aiuto, M.; Caraglia, M.; et al. HtrA1 loss is related to aggressive behavior parameters in sentinel node positive breast cancer. Histol. Histopathol. 2015, 30, 707–714. [Google Scholar] [CrossRef]
- Lehner, A.; Magdolen, V.; Schuster, T.; Kotzsch, M.; Kiechle, M.; Meindl, A.; Sweep, F.C.G.J.; Span, P.N.; Gross, E. Downregulation of serine protease HTRA1 is associated with poor survival in breast cancer. PLoS ONE 2013, 8, e60359. [Google Scholar] [CrossRef] [Green Version]
- Baldi, A.; De Luca, A.; Morini, M.; Battista, T.; Felsani, A.; Baldi, F.; Catricalà, C.; Amantea, A.; Noonan, D.M.; Albini, A.; et al. The HtrA1 serine protease is down-regulated during human melanoma progression and represses growth of metastatic melanoma cells. Oncogene 2002, 21, 6684–6688. [Google Scholar] [CrossRef]
- Chien, J.; Staub, J.; Hu, S.-I.; Erickson-Johnson, M.R.; Couch, F.J.; Smith, D.I.; Crowl, R.M.; Kaufmann, S.H.; Shridhar, V. A candidate tumor suppressor HtrA1 is downregulated in ovarian cancer. Oncogene 2004, 23, 1636–1644. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Khurana, A.; Maguire, J.L.; Chien, J.; Shridhar, V. HtrA1 sensitizes ovarian cancer cells to cisplatin-induced cytotoxicity by targeting XIAP for degradation. Int. J. Cancer 2012, 130, 1029–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Ota, T.; Liu, P.; Su, C.; Chien, J.; Shridhar, V. Downregulation of HtrA1 promotes resistance to anoikis and peritoneal dissemination of ovarian cancer cells. Cancer Res. 2010, 70, 3109–3118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullany, S.A.; Moslemi-Kebria, M.; Rattan, R.; Khurana, A.; Clayton, A.; Ota, T.; Mariani, A.; Podratz, K.C.; Chien, J.; Shridhar, V. Expression and functional significance of HtrA1 loss in endometrial cancer. Clin. Cancer Res. 2011, 17, 427–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.; Wang, F.; Wang, L.; Fan, Q. Elevated serine protease HtrA1 inhibits cell proliferation, reduces invasion, and induces apoptosis in esophageal squamous cell carcinoma by blocking the nuclear factor-κB signaling pathway. Tumor Biol. 2013, 34, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Chien, J.; Ota, T.; Aletti, G.; Shridhar, R.; Boccellino, M.; Quagliuolo, L.; Baldi, A.; Shridhar, V. Serine protease HtrA1 associates with microtubules and inhibits cell migration. Mol. Cell. Biol. 2009, 29, 4177–4187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.-L.; Zhao, L.-J. Mechanistic basis and clinical relevance of the role of transforming growth factor-β in cancer. Cancer Biol. Med. 2015, 12, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Lv, L.; Liu, Y.; Chen, B.; Li, M.; Ni, T.; Yang, D.; Zhu, H.; Xue, Q.; Ni, R. The expression levels and prognostic value of high temperature required A2 (HtrA2) in NSCLC. Pathol. Res. Pract. 2014, 210, 939–943. [Google Scholar] [CrossRef]
- Miyamoto, M.; Takano, M.; Iwaya, K.; Shinomiya, N.; Goto, T.; Kato, M.; Suzuki, A.; Aoyama, T.; Hirata, J.; Nagaoka, I.; et al. High-temperature-required protein A2 as a predictive marker for response to chemotherapy and prognosis in patients with high-grade serous ovarian cancers. Br. J. Cancer 2015, 112, 739–744. [Google Scholar] [CrossRef]
- Zhang, F.; Yu, T.; Yi, C.; Sun, X.-F. Radiation-inducible HtrA2 gene enhances radiosensitivity of uveal melanoma OCM-1 cells in vitro and in vivo. Clin. Experiment. Ophthalmol. 2014, 42, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Dynon, K.; Heng, S.; Puryer, M.; Li, Y.; Walton, K.; Endo, Y.; Nie, G. HtrA3 as an early marker for preeclampsia: Specific monoclonal antibodies and sensitive high-throughput assays for serum screening. PLoS ONE 2012, 7, e45956. [Google Scholar] [CrossRef]
- Li, Y.; Puryer, M.; Lin, E.; Hale, K.; Salamonsen, L.A.; Manuelpillai, U.; Tong, S.; Chan, W.; Wallace, E.M.; Nie, G. Placental HtrA3 is regulated by oxygen tension and serum levels are altered during early pregnancy in women destined to develop preeclampsia. J. Clin. Endocrinol. Metab. 2011, 96, 403–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, G.-Y.; Li, Y.; Minoura, H.; Batten, L.; Ooi, G.T.; Findlay, J.K.; Salamonsen, L.A. A novel serine protease of the mammalian HtrA family is up-regulated in mouse uterus coinciding with placentation. Mol. Hum. Reprod. 2003, 9, 279–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, H.; Makino, S.-I.; Endo, Y.; Nie, G. Inhibition of HTRA3 stimulates trophoblast invasion during human placental development. Placenta 2010, 31, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Beleford, D.; Rattan, R.; Chien, J.; Shridhar, V. High temperature requirement A3 (HtrA3) promotes etoposide- and cisplatin-induced cytotoxicity in lung cancer cell lines. J. Biol. Chem. 2010, 285, 12011–12027. [Google Scholar] [CrossRef] [Green Version]
- Wenta, T.; Rychlowski, M.; Jurewicz, E.; Jarzab, M.; Zurawa-Janicka, D.; Filipek, A.; Lipinska, B. The HtrA3 protease promotes drug-induced death of lung cancer cells by cleavage of the X-linked inhibitor of apoptosis protein (XIAP). FEBS J. 2019, 286, 4579–4596. [Google Scholar] [CrossRef]
- Beleford, D.; Liu, Z.; Rattan, R.; Quagliuolo, L.; Boccellino, M.; Baldi, A.; Maguire, J.; Staub, J.; Molina, J.; Shridhar, V. Methylation induced gene silencing of HtrA3 in smoking-related lung cancer. Clin. Cancer Res. 2010, 16, 398–409. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Zhang, J.; Zhang, X.; Feng, M.; Qu, J. High temperature requirement A3 (HTRA3) expression predicts postoperative recurrence and survival in patients with non-small-cell lung cancer. Oncotarget 2016, 7, 40725–40734. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.; Li, Y.; Fuller, P.J.; Harrison, C.; Rao, J.; Stephens, A.N.; Nie, G. HtrA3 is Downregulated in Cancer Cell Lines and Significantly Reduced in Primary Serous and Granulosa Cell Ovarian Tumors. J. Cancer 2013, 4, 152–164. [Google Scholar] [CrossRef] [Green Version]
- Zurawa-Janicka, D.; Kobiela, J.; Galczynska, N.; Stefaniak, T.; Lipinska, B.; Lachinski, A.; Skorko-Glonek, J.; Narkiewicz, J.; Proczko-Markuszewska, M.; Sledzinski, Z. Changes in expression of human serine protease HtrA1, HtrA2 and HtrA3 genes in benign and malignant thyroid tumors. Oncol. Rep. 2012, 28, 1838–1844. [Google Scholar] [CrossRef] [Green Version]
- Tocharus, J.; Tsuchiya, A.; Kajikawa, M.; Ueta, Y.; Oka, C.; Kawaichi, M. Developmentally regulated expression of mouse HtrA3 and its role as an inhibitor of TGF-beta signaling. Dev. Growth Differ. 2004, 46, 257–274. [Google Scholar] [CrossRef]
- Narkiewicz, J.; Lapinska-Szumczyk, S.; Zurawa-Janicka, D.; Skorko-Glonek, J.; Emerich, J.; Lipinska, B. Expression of human HtrA1, HtrA2, HtrA3 and TGF-beta1 genes in primary endometrial cancer. Oncol. Rep. 2009, 21, 1529–1537. [Google Scholar] [PubMed]
- Narkiewicz, J.; Klasa-Mazurkiewicz, D.; Zurawa-Janicka, D.; Skorko-Glonek, J.; Emerich, J.; Lipinska, B. Changes in mRNA and protein levels of human HtrA1, HtrA2 and HtrA3 in ovarian cancer. Clin. Biochem. 2008, 41, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Altobelli, E.; Latella, G.; Morroni, M.; Licini, C.; Tossetta, G.; Mazzucchelli, R.; Profeta, V.F.; Coletti, G.; Leocata, P.; Castellucci, M.; et al. Low HtrA1 expression in patients with long-standing ulcerative colitis and colorectal cancer. Oncol. Rep. 2017, 38, 418–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forse, C.L.; Rahimi, M.; Diamandis, E.P.; Assarzadegan, N.; Dawson, H.; Grin, A.; Kennedy, E.; O’Connor, B.; Messenger, D.E.; Riddell, R.H.; et al. HtrA3 stromal expression is correlated with tumor budding in stage II colorectal cancer. Exp. Mol. Pathol. 2017, 103, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Boland, C.R.; Thibodeau, S.N.; Hamilton, S.R.; Sidransky, D.; Eshleman, J.R.; Burt, R.W.; Meltzer, S.J.; Rodriguez-Bigas, M.A.; Fodde, R.; Ranzani, G.N.; et al. A National Cancer Institute Workshop on Microsatellite Instability for cancer detection and familial predisposition: Development of international criteria for the determination of microsatellite instability in colorectal cancer. Cancer Res. 1998, 58, 5248–5257. [Google Scholar] [PubMed]
- Xiong, Z.; Fu, Z.; Shi, J.; Jiang, X.; Wan, H. HtrA1 Down-regulation Induces Cisplatin Resistance in Colon Cancer by Increasing XIAP and Activating PI3K/Akt Pathway. Ann. Clin. Lab. Sci. 2017, 47, 264–270. [Google Scholar]
- Zurawa-Janicka, D.; Kobiela, J.; Stefaniak, T.; Wozniak, A.; Narkiewicz, J.; Wozniak, M.; Limon, J.; Lipinska, B. Changes in expression of serine proteases HtrA1 and HtrA2 during estrogen-induced oxidative stress and nephrocarcinogenesis in male Syrian hamster. Acta Biochim. Pol. 2008, 55, 9–19. [Google Scholar] [CrossRef]
- Liu, C.; Xing, F.; He, Y.; Zong, S.; Luo, C.; Li, C.; Duan, T.; Wang, K.; Zhou, Q. Elevated HTRA1 and HTRA4 in severe preeclampsia and their roles in trophoblast functions. Mol. Med. Rep. 2018, 18, 2937–2944. [Google Scholar] [CrossRef] [Green Version]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef]
- Sarro, S.M.; Unruh, T.L.; Zuccolo, J.; Sanyal, R.; Luider, J.M.; Auer-Grzesiak, I.A.; Mansoor, A.; Deans, J.P. Quantification of CD20 mRNA and protein levels in chronic lymphocytic leukemia suggests a post-transcriptional defect. Leuk. Res. 2010, 34, 1670–1673. [Google Scholar] [CrossRef]
- Stark, A.M.; Pfannenschmidt, S.; Tscheslog, H.; Maass, N.; Rösel, F.; Mehdorn, H.M.; Held-Feindt, J. Reduced mRNA and protein expression of BCL-2 versus decreased mRNA and increased protein expression of BAX in breast cancer brain metastases: A real-time PCR and immunohistochemical evaluation. Neurol. Res. 2006, 28, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Taquet, N.; Dumont, S.; Vonesch, J.-L.; Hentsch, D.; Reimund, J.-M.; Muller, C.D. Differential between protein and mRNA expression of CCR7 and SSTR5 receptors in Crohn’s disease patients. Mediat. Inflamm. 2009, 2009, 285812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flusberg, D.A.; Sorger, P.K. Surviving apoptosis: Life-death signaling in single cells. Trends Cell Biol. 2015, 25, 446–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulda, S. Evasion of apoptosis as a cellular stress response in cancer. Int. J. Cell Biol. 2010, 2010, 370835. [Google Scholar] [CrossRef]
- Han, R.; Huang, G.; Wang, Y.; Xu, Y.; Hu, Y.; Jiang, W.; Wang, T.; Xiao, T.; Zheng, D. Increased gene expression noise in human cancers is correlated with low p53 and immune activities as well as late stage cancer. Oncotarget 2016, 7, 72011–72020. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, N.; Irle, I.; Ripkens, K.; Lux, V.; Nelles, J.; Johannes, C.; Parry, L.; Greenow, K.; Amir, S.; Campioni, M.; et al. Epigenetic silencing of serine protease HTRA1 drives polyploidy. BMC Cancer 2016, 16, 399. [Google Scholar] [CrossRef] [Green Version]
- Lampropoulos, P.; Zizi-Sermpetzoglou, A.; Rizos, S.; Kostakis, A.; Nikiteas, N.; Papavassiliou, A.G. TGF-beta signalling in colon carcinogenesis. Cancer Lett. 2012, 314, 1–7. [Google Scholar] [CrossRef]
- Langenskiöld, M.; Holmdahl, L.; Falk, P.; Angenete, E.; Ivarsson, M.-L. Increased TGF-beta 1 protein expression in patients with advanced colorectal cancer. J. Surg. Oncol. 2008, 97, 409–415. [Google Scholar] [CrossRef]
- Miura, K.; Fujibuchi, W.; Ishida, K.; Naitoh, T.; Ogawa, H.; Ando, T.; Yazaki, N.; Watanabe, K.; Haneda, S.; Shibata, C.; et al. Inhibitor of apoptosis protein family as diagnostic markers and therapeutic targets of colorectal cancer. Surg. Today 2011, 41, 175–182. [Google Scholar] [CrossRef]
- Liu, Z.; Li, H.; Derouet, M.; Berezkin, A.; Sasazuki, T.; Shirasawa, S.; Rosen, K. Oncogenic Ras inhibits anoikis of intestinal epithelial cells by preventing the release of a mitochondrial pro-apoptotic protein Omi/HtrA2 into the cytoplasm. J. Biol. Chem. 2006, 281, 14738–14747. [Google Scholar] [CrossRef] [Green Version]
- Yoo, B.H.; Wang, Y.; Erdogan, M.; Sasazuki, T.; Shirasawa, S.; Corcos, L.; Sabapathy, K.; Rosen, K.V. Oncogenic ras-induced down-regulation of pro-apoptotic protease caspase-2 is required for malignant transformation of intestinal epithelial cells. J. Biol. Chem. 2011, 286, 38894–38903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, K. Colorectal cancer development and advances in screening. Clin. Interv. Aging 2016, 11, 967–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boland, C.R.; Goel, A. Microsatellite instability in colorectal cancer. Gastroenterology 2010, 138, 2073–2087. [Google Scholar] [CrossRef] [PubMed]
- Karagiannis, G.S.; Treacy, A.; Messenger, D.; Grin, A.; Kirsch, R.; Riddell, R.H.; Diamandis, E.P. Expression patterns of bone morphogenetic protein antagonists in colorectal cancer desmoplastic invasion fronts. Mol. Oncol. 2014, 8, 1240–1252. [Google Scholar] [CrossRef]
- AJCC Cancer Staging Manual, 8th ed.; Amin, M.B.; Edge, S.; Greene, F.; Byrd, D.R.; Brookland, R.K.; Washington, M.K.; Gershenwald, J.E.; Compton, C.C.; Hess, K.R.; Sullivan, D.C.; et al. (Eds.) American Joint Committee on Cancer: Chicago, IL, USA, 2017; ISBN 978-3-319-40617-6. [Google Scholar]
- Green, M.R.; Sambrook, J.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012; ISBN 978-1-936113-41-5. [Google Scholar]
- Sørby, L.A.; Andersen, S.N.; Bukholm, I.R.K.; Jacobsen, M.B. Evaluation of suitable reference genes for normalization of real-time reverse transcription PCR analysis in colon cancer. J. Exp. Clin. Cancer Res. 2010, 29, 144. [Google Scholar] [CrossRef] [Green Version]
- Wierzbicki, P.M.; Klacz, J.; Rybarczyk, A.; Slebioda, T.; Stanislawowski, M.; Wronska, A.; Kowalczyk, A.; Matuszewski, M.; Kmiec, Z. Identification of a suitable qPCR reference gene in metastatic clear cell renal cell carcinoma. Tumor Biol. 2014, 35, 12473–12487. [Google Scholar] [CrossRef] [Green Version]
- Van Eijsden, R.G.E.; Stassen, C.; Daenen, L.; Van Mulders, S.E.; Bapat, P.M.; Siewers, V.; Goossens, K.V.Y.; Nielsen, J.; Delvaux, F.R.; Van Hummelen, P.; et al. A universal fixation method based on quaternary ammonium salts (RNAlater) for omics-technologies: Saccharomyces cerevisiae as a case study. Biotechnol. Lett. 2013, 35, 891–900. [Google Scholar] [CrossRef]
- Glaza, P.; Osipiuk, J.; Wenta, T.; Zurawa-Janicka, D.; Jarzab, M.; Lesner, A.; Banecki, B.; Skorko-Glonek, J.; Joachimiak, A.; Lipinska, B. Structural and Functional Analysis of Human HtrA3 Protease and Its Subdomains. PLoS ONE 2015, 10, e0131142. [Google Scholar] [CrossRef]
- Stanislawowski, M.; Wierzbicki, P.M.; Golab, A.; Adrych, K.; Kartanowicz, D.; Wypych, J.; Godlewski, J.; Smoczynski, M.; Kmiec, Z. Decreased Toll-like receptor-5 (TLR-5) expression in the mucosa of ulcerative colitis patients. J. Physiol. Pharmacol. 2009, 60 (Suppl. 4), 71–75. [Google Scholar]
- Janavicius, R.; Matiukaite, D.; Jakubauskas, A.; Griskevicius, L. Microsatellite instability detection by high-resolution melting analysis. Clin. Chem. 2010, 56, 1750–1757. [Google Scholar] [CrossRef] [Green Version]
- Odenthal, M.; Barta, N.; Lohfink, D.; Drebber, U.; Schulze, F.; Dienes, H.P.; Baldus, S.E. Analysis of microsatellite instability in colorectal carcinoma by microfluidic-based chip electrophoresis. J. Clin. Pathol. 2009, 62, 850–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients Demographics | No. of Cases - n (%) |
|---|---|
| Total | 65 (100) |
| Gender | |
| Male | 41 (63) |
| Female | 24 (37) |
| Age | |
| Years ± SD | 67 ± 10.5 |
| Tumor stage * | |
| I + II | 30 (46) |
| III + IV | 35 (54) |
| Anatomical location | |
| Right-sided | 29 (45) |
| Left-sided | 36 (55) |
| Gene | Oligonucleotide Sequence 5′→ 3′ | Annealing Temp. (°C) | PCR Product Size (bp) | RefSeq | Ref. |
|---|---|---|---|---|---|
| HTRA1 | Forward: CGGAAGATGGACTGATCGTGAC Reverse: GGTGATGGCTTTTCCTTTGGCC | 62 | 506 | NM_002775.4 | [42] |
| HTRA2 | Forward: CCCTATCTCGAACGGCTCAGG Reverse: CCATGCTGAACATCGGGAAAGC | 63 | 639 | NM_013247.4 | [42] |
| HTRA3L | Forward: AGATCAAAGACTGGAAGAAGCG Reverse: ATGATGTCACCATCTTGGATGC | 60 | 181 | NM_053044.1 | This work |
| HTRA3S | Forward: ATCCATCCCAAGAAAAAGCTCC Reverse: CTCAATGAACTGCCAGTGAGG | 60 | 130 | NM_001297559.1 | This work |
| PPIA | Forward: CTTGGGCCGCGTCTCCTTTGAG Reverse: GCTTGCCATCCAACCACTCAGTC | 59 | 329 | NM_001300981.1 | [68] |
| IPO8 | Forward: TTGGAAGAAACCGCGCTTGAGG Reverse: ACCAGGCTGCATCTCGACTCTG | 59 | 118 | NM_001190995 | [68] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zurawa-Janicka, D.; Kobiela, J.; Slebioda, T.; Peksa, R.; Stanislawowski, M.; Wierzbicki, P.M.; Wenta, T.; Lipinska, B.; Kmiec, Z.; Biernat, W.; et al. Expression of HTRA Genes and Its Association with Microsatellite Instability and Survival of Patients with Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 3947. https://doi.org/10.3390/ijms21113947
Zurawa-Janicka D, Kobiela J, Slebioda T, Peksa R, Stanislawowski M, Wierzbicki PM, Wenta T, Lipinska B, Kmiec Z, Biernat W, et al. Expression of HTRA Genes and Its Association with Microsatellite Instability and Survival of Patients with Colorectal Cancer. International Journal of Molecular Sciences. 2020; 21(11):3947. https://doi.org/10.3390/ijms21113947
Chicago/Turabian StyleZurawa-Janicka, Dorota, Jarek Kobiela, Tomasz Slebioda, Rafal Peksa, Marcin Stanislawowski, Piotr Mieczyslaw Wierzbicki, Tomasz Wenta, Barbara Lipinska, Zbigniew Kmiec, Wojciech Biernat, and et al. 2020. "Expression of HTRA Genes and Its Association with Microsatellite Instability and Survival of Patients with Colorectal Cancer" International Journal of Molecular Sciences 21, no. 11: 3947. https://doi.org/10.3390/ijms21113947
APA StyleZurawa-Janicka, D., Kobiela, J., Slebioda, T., Peksa, R., Stanislawowski, M., Wierzbicki, P. M., Wenta, T., Lipinska, B., Kmiec, Z., Biernat, W., Lachinski, A. J., & Sledzinski, Z. (2020). Expression of HTRA Genes and Its Association with Microsatellite Instability and Survival of Patients with Colorectal Cancer. International Journal of Molecular Sciences, 21(11), 3947. https://doi.org/10.3390/ijms21113947