Antiviral Activity of Compound L3 against Dengue and Zika Viruses In Vitro and In Vivo

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
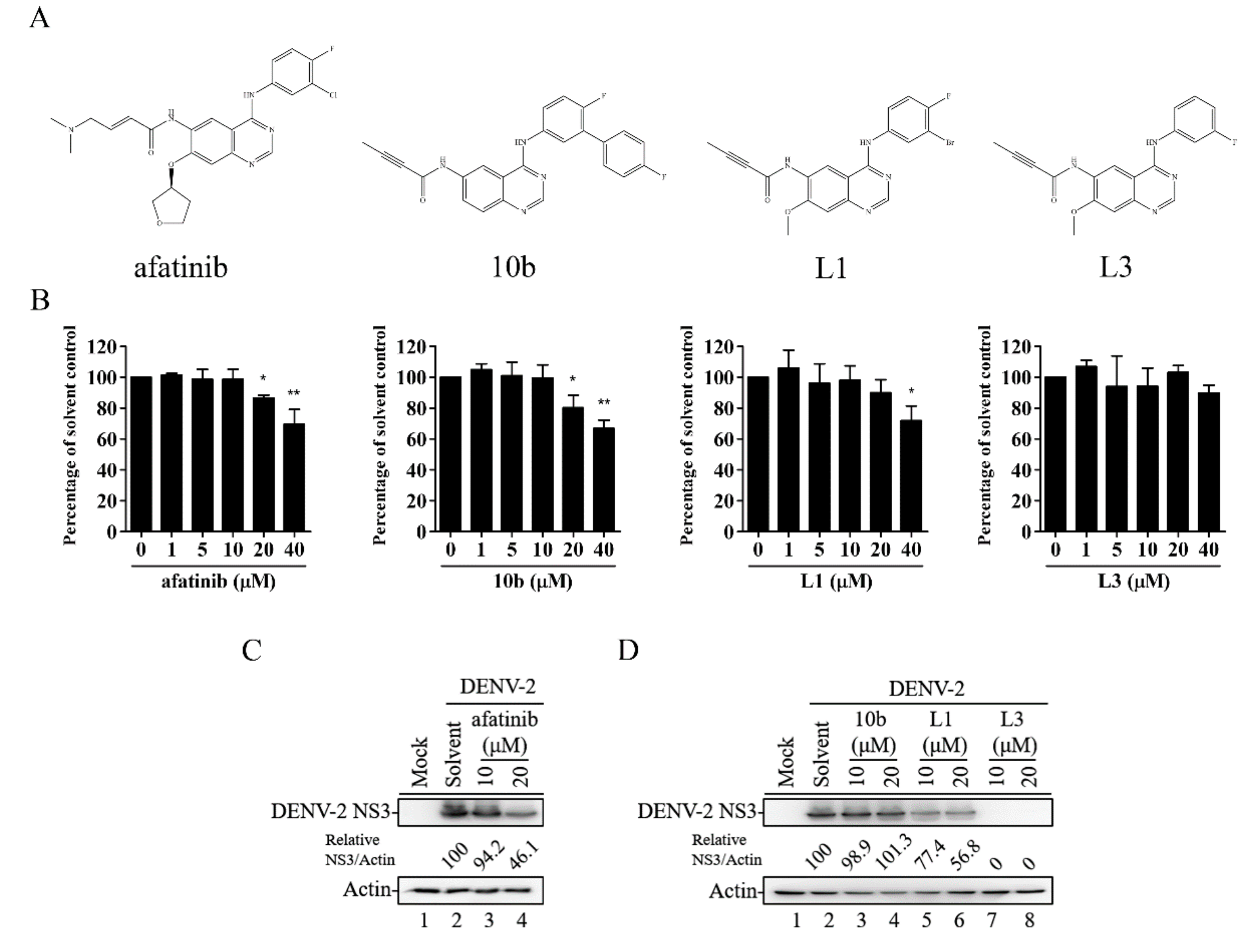
2.1. Series of Afatinib-Derivative TKI Compounds Exhibiting Antiviral Activities without Cytotoxic Effects
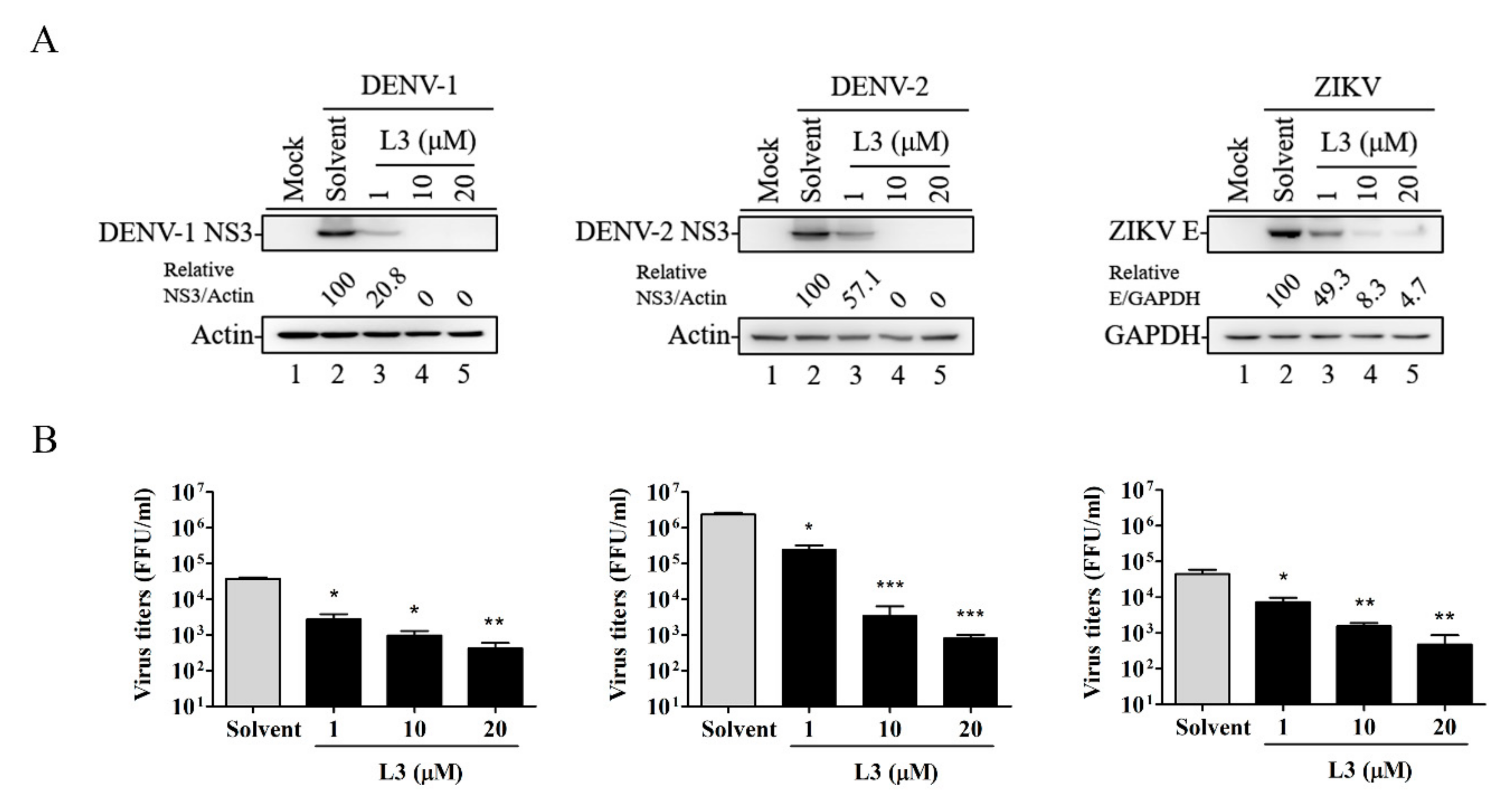
2.2. Compound L3 Exhibits Antiviral Activity against DENV-1, DENV-2, and ZIKV
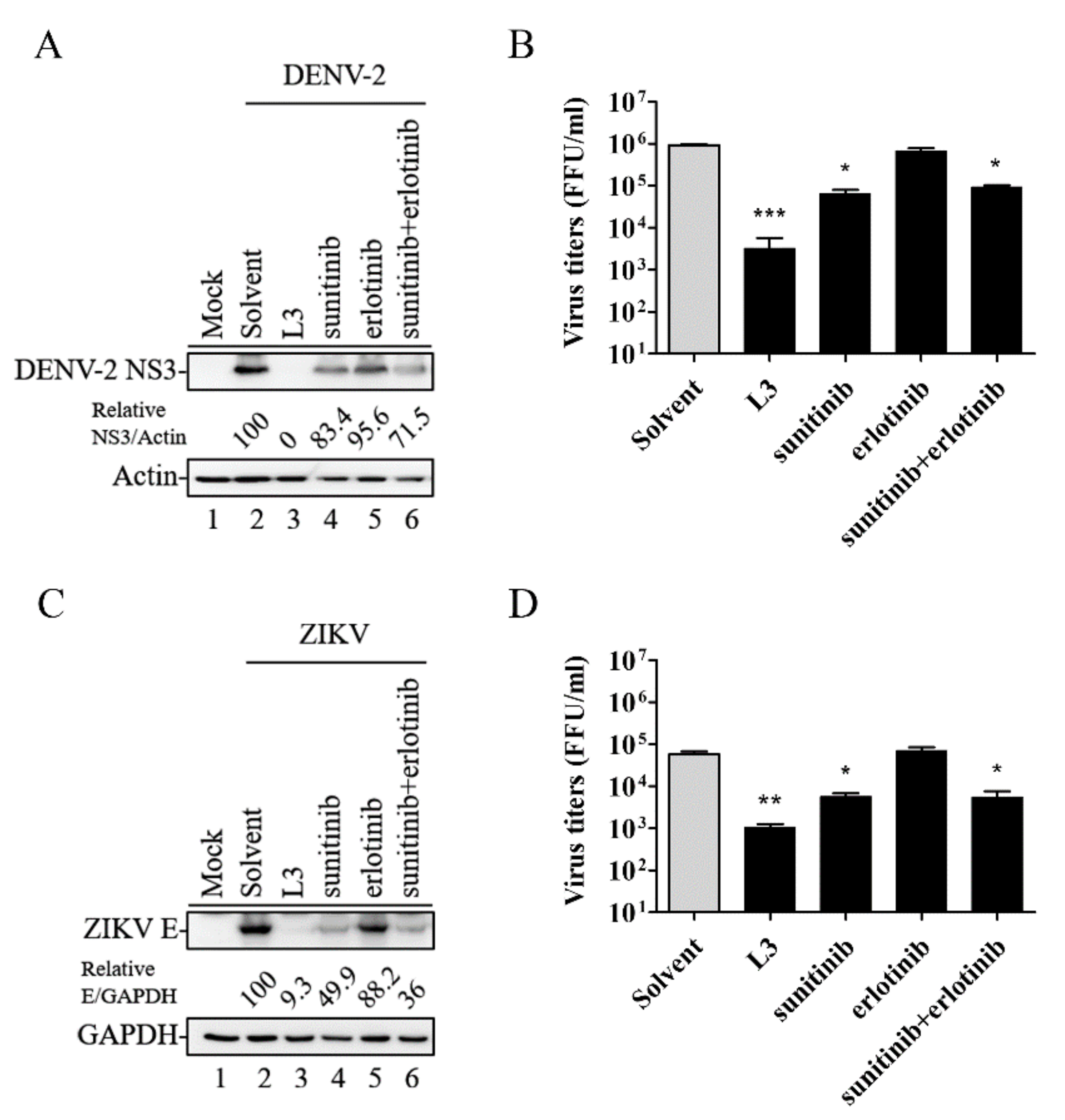
2.3. Compound L3 Shows Therapeutic Efficacy against DENV-2 and ZIKV Compared to Other TKI Inhibitors
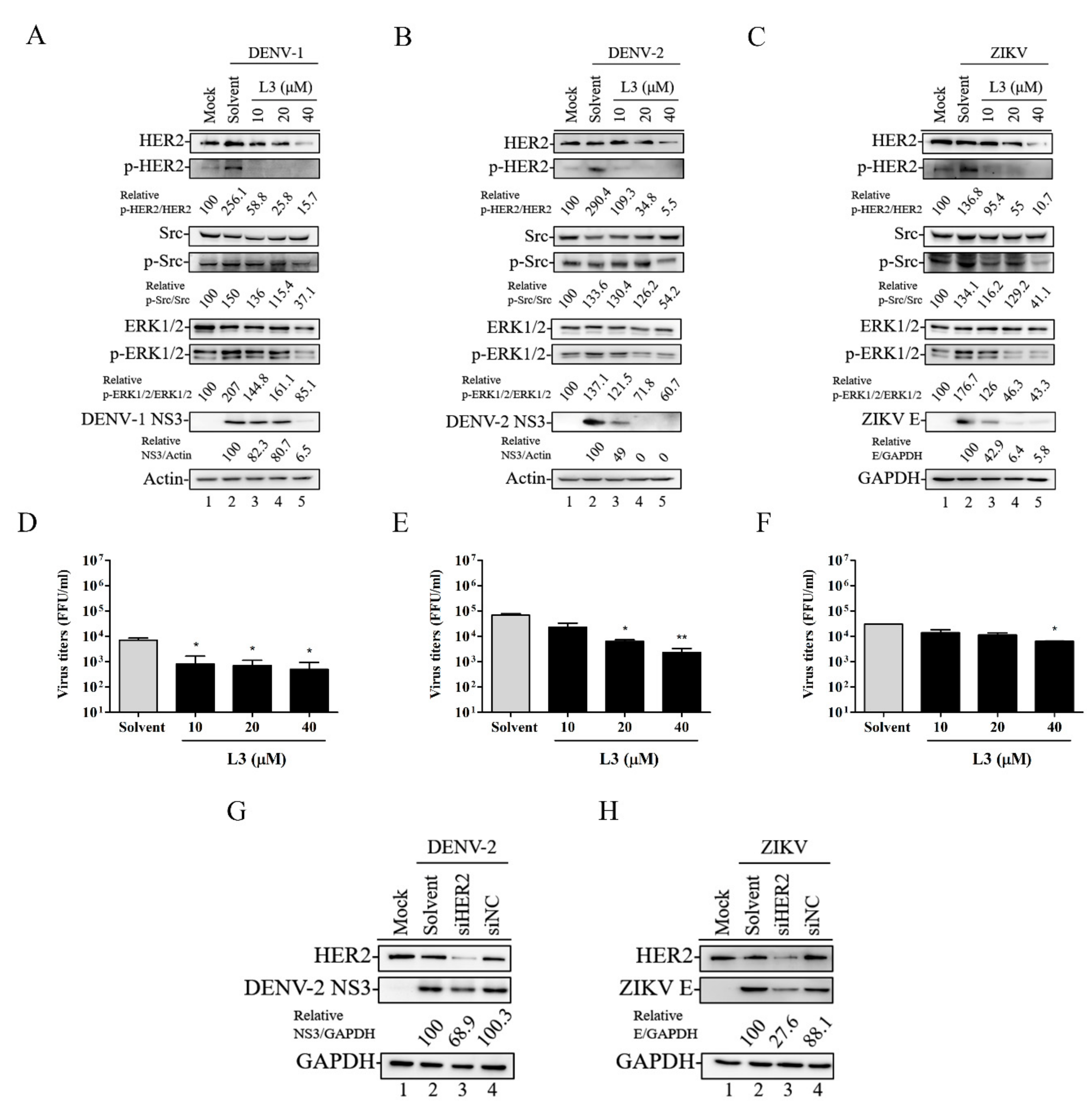
2.4. Compound L3 Inhibits DENV and ZIKV Replication through the HER2 Signaling Pathway
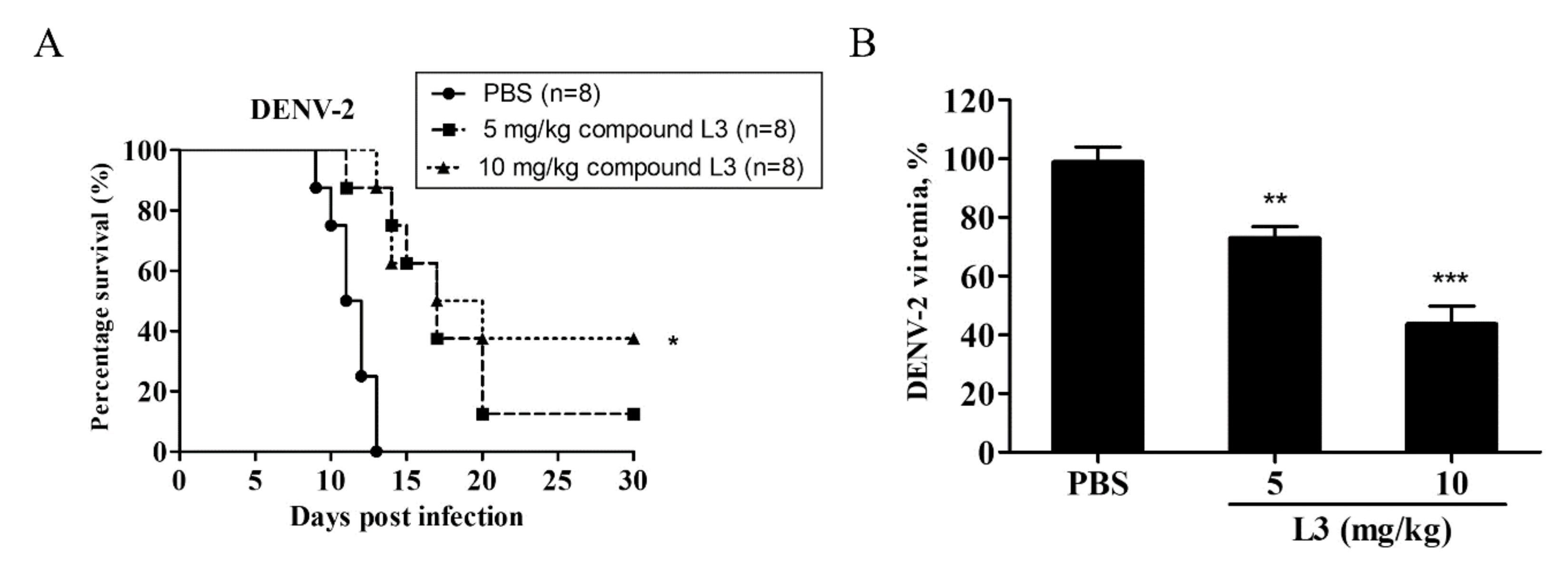
2.5. Compound L3 Alleviates DENV-2 Infection in AG129 Mice
3. Discussion
4. Materials and Methods
4.1. Synthesis of Compounds 10b, L1, and L3
4.2. Cell Lines and Viruses
4.3. Drug Cytotoxicity Assay
4.4. Western Blotting
4.5. HER2 Silencing
4.6. Mouse Model
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Neufeldt, C.J.; Cortese, M.; Acosta, E.G.; Bartenschlager, R. Rewiring cellular networks by members of the flaviviridae family. Nat. Rev. Microbiol. 2018, 16, 125–142. [Google Scholar] [CrossRef]
- Chong, H.Y.; Leow, C.Y.; Abdul Majeed, A.B.; Leow, C.H. Flavivirus infection-a review of immunopathogenesis, immunological response, and immunodiagnosis. Virus Res. 2019, 274, 197770. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Franco, L.; Gushi, L.T.; Luiz, W.B.; Amorim, J.H. Seeking flavivirus cross-protective immunity. Front. Immunol. 2019, 10, 2260. [Google Scholar] [CrossRef]
- Quicke, K.M.; Bowen, J.R.; Johnson, E.L.; McDonald, C.E.; Ma, H.; O’Neal, J.T.; Rajakumar, A.; Wrammert, J.; Rimawi, B.H.; Pulendran, B.; et al. Zika virus infects human placental macrophages. Cell Host Microbe 2016, 20, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, S.A.; Jamieson, D.J.; Honein, M.A.; Petersen, L.R. Zika virus and birth defects--reviewing the evidence for causality. N. Engl. J. Med. 2016, 374, 1981–1987. [Google Scholar] [CrossRef] [PubMed]
- Manet, C.; Roth, C.; Tawfik, A.; Cantaert, T.; Sakuntabhai, A.; Montagutelli, X. Host genetic control of mosquito-borne flavivirus infections. Mamm. Genome Off. J. Int. Mamm. Genome Soc. 2018, 29, 384–407. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Garcia, M.D.; Mazzon, M.; Jacobs, M.; Amara, A. Pathogenesis of flavivirus infections: Using and abusing the host cell. Cell Host Microbe 2009, 5, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Stein, D.A.; Shum, D.; Fischer, M.A.; Radu, C.; Bhinder, B.; Djaballah, H.; Nelson, J.A.; Fruh, K.; Hirsch, A.J. Inhibition of dengue virus replication by a class of small-molecule compounds that antagonize dopamine receptor d4 and downstream mitogen-activated protein kinase signaling. J. Virol. 2014, 88, 5533–5542. [Google Scholar] [CrossRef] [Green Version]
- Meertens, L.; Carnec, X.; Lecoin, M.P.; Ramdasi, R.; Guivel-Benhassine, F.; Lew, E.; Lemke, G.; Schwartz, O.; Amara, A. The tim and tam families of phosphatidylserine receptors mediate dengue virus entry. Cell Host Microbe 2012, 12, 544–557. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Agrawal, T.; Khan, N.A.; Nakayama, Y.; Medigeshi, G.R. Identification and characterization of the role of c-terminal src kinase in dengue virus replication. Sci. Rep. 2016, 6, 30490. [Google Scholar] [CrossRef]
- De Wispelaere, M.; LaCroix, A.J.; Yang, P.L. The small molecules azd0530 and dasatinib inhibit dengue virus rna replication via fyn kinase. J. Virol. 2013, 87, 7367–7381. [Google Scholar] [CrossRef] [Green Version]
- Zakaria, M.K.; Carletti, T.; Marcello, A. Cellular targets for the treatment of flavivirus infections. Front. Cell. Infect. Microbiol. 2018, 8, 398. [Google Scholar] [CrossRef]
- Bekerman, E.; Neveu, G.; Shulla, A.; Brannan, J.; Pu, S.Y.; Wang, S.; Xiao, F.; Barouch-Bentov, R.; Bakken, R.R.; Mateo, R.; et al. Anticancer kinase inhibitors impair intracellular viral trafficking and exert broad-spectrum antiviral effects. J. Clin. Investig. 2017, 127, 1338–1352. [Google Scholar] [CrossRef]
- Chu, J.J.; Yang, P.L. C-src protein kinase inhibitors block assembly and maturation of dengue virus. Proc. Natl. Acad. Sci. USA 2007, 104, 3520–3525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdallah, S.M.; Hirsh, V. Irreversible tyrosine kinase inhibition of epidermal growth factor receptor with afatinib in egfr activating mutation-positive advanced non-small-cell lung cancer. Curr. Oncol. 2018, 25, S9–S17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsh, V. Next-generation covalent irreversible kinase inhibitors in nsclc: Focus on afatinib. BioDrugs Clin. Immunothe. Biopharm. Gene Ther. 2015, 29, 167–183. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Gao, X.; Maragh, S.; Telford, W.G.; Tona, A. Cell lines as candidate reference materials for quality control of erbb2 amplification and expression assays in breast cancer. Clin. Chem. 2009, 55, 1307–1315. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Hackshaw, A.; Feng, Q.; Fu, X.; Zhang, Y.; Mao, C.; Tang, J. Comparison of gefitinib, erlotinib and afatinib in non-small cell lung cancer: A meta-analysis. Int. J. Cancer 2017, 140, 2805–2819. [Google Scholar] [CrossRef]
- Samanta, J.; Sharma, V. Dengue and its effects on liver. World J. Clin. Cases 2015, 3, 125–131. [Google Scholar] [CrossRef]
- Thepparit, C.; Khakpoor, A.; Khongwichit, S.; Wikan, N.; Fongsaran, C.; Chingsuwanrote, P.; Panraksa, P.; Smith, D.R. Dengue 2 infection of hepg2 liver cells results in endoplasmic reticulum stress and induction of multiple pathways of cell death. BMC Res. Notes 2013, 6, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.L.; Liu, C.C.; Lei, H.Y.; Yeh, T.M.; Lin, Y.S.; Chen, R.M.; Liu, H.S. Infection of five human liver cell lines by dengue-2 virus. J. Med. Virol. 2000, 60, 425–431. [Google Scholar] [CrossRef]
- Pu, S.Y.; Xiao, F.; Schor, S.; Bekerman, E.; Zanini, F.; Barouch-Bentov, R.; Nagamine, C.M.; Einav, S. Feasibility and biological rationale of repurposing sunitinib and erlotinib for dengue treatment. Antivir. Res. 2018, 155, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, B. Targeted therapeutic options and future perspectives for her2-positive breast cancer. Signal. Trans. Target. Ther. 2019, 4, 34. [Google Scholar] [CrossRef] [Green Version]
- Gingras, I.; Gebhart, G.; de Azambuja, E.; Piccart-Gebhart, M. Her2-positive breast cancer is lost in translation: Time for patient-centered research. Nat. Rev. Clin.l Oncol. 2017, 14, 669–681. [Google Scholar] [CrossRef]
- Vu, T.; Claret, F.X. Trastuzumab: Updated mechanisms of action and resistance in breast cancer. Front. Oncol. 2012, 2, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.J.; Higgs, S.; Horne, K.M.; Vanlandingham, D.L. Flavivirus-mosquito interactions. Viruses 2014, 6, 4703–4730. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Estofolete, C.F.; Terzian, A.C.B.; Colombo, T.E.; de Freitas Guimaraes, G.; Ferraz, H.C.J.; da Silva, R.A.; Greque, G.V.; Nogueira, M.L. Co-infection between zika and different dengue serotypes during denv outbreak in brazil. J. Infect. Public Health 2019, 12, 178–181. [Google Scholar] [CrossRef]
- Laredo-Tiscareno, S.V.; Garza-Hernandez, J.A.; Salazar, M.I.; De Luna-Santillana, E.J.; Tangudu, C.S.; Cetina-Trejo, R.C.; Doria-Cobos, G.L.; Carmona-Aguirre, S.D.; Garcia-Rejon, J.E.; Machain-Williams, C.; et al. Surveillance for flaviviruses near the mexico-u.S. Border: Co-circulation of dengue virus serotypes 1, 2, and 3 and west nile virus in tamaulipas, northern mexico, 2014–2016. Am. J. Trop. Med. Hyg. 2018, 99, 1308–1317. [Google Scholar] [CrossRef] [Green Version]
- Behnam, M.A.; Nitsche, C.; Boldescu, V.; Klein, C.D. The medicinal chemistry of dengue virus. J. Med. Chem. 2016, 59, 5622–5649. [Google Scholar] [CrossRef] [Green Version]
- Ben-Shachar, R.; Schmidler, S.; Koelle, K. Drivers of inter-individual variation in dengue viral load dynamics. PLoS Comput.Biol. 2016, 12, e1005194. [Google Scholar] [CrossRef]
- Stein, D.A.; Perry, S.T.; Buck, M.D.; Oehmen, C.S.; Fischer, M.A.; Poore, E.; Smith, J.L.; Lancaster, A.M.; Hirsch, A.J.; Slifka, M.K.; et al. Inhibition of dengue virus infections in cell cultures and in ag129 mice by a small interfering rna targeting a highly conserved sequence. J. Virol. 2011, 85, 10154–10166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreekanth, G.P.; Chuncharunee, A.; Sirimontaporn, A.; Panaampon, J.; Srisawat, C.; Morchang, A.; Malakar, S.; Thuwajit, P.; Kooptiwut, S.; Suttitheptumrong, A.; et al. Role of erk1/2 signaling in dengue virus-induced liver injury. Virus Res. 2014, 188, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.Y.; Pan, C.H.; Hsieh, C.H.; Tsai, J.P.; Chen, M.Y.; Liu, H.H.; Liu, S.J.; Chong, P.; Leng, C.H.; Chen, H.W. Lipidated dengue-2 envelope protein domain iii independently stimulates long-lasting neutralizing antibodies and reduces the risk of antibody-dependent enhancement. PLoS Negle. Trop. Dis. 2013, 7, e2432. [Google Scholar] [CrossRef] [PubMed]
- Simanjuntak, Y.; Liang, J.J.; Chen, S.Y.; Li, J.K.; Lee, Y.L.; Wu, H.C.; Lin, Y.L. Ebselen alleviates testicular pathology in mice with zika virus infection and prevents its sexual transmission. PLoS Pathog. 2018, 14, e1006854. [Google Scholar] [CrossRef]
- Hsieh, M.S.; Chen, M.Y.; Hsieh, C.H.; Pan, C.H.; Yu, G.Y.; Chen, H.W. Detection and quantification of dengue virus using a novel biosensor system based on dengue ns3 protease activity. PLoS ONE 2017, 12, e0188170. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | CC50 a (μM) | IC50 b (μM) | SI c (CC50/IC50) |
|---|---|---|---|
| DENV-1 | 61.4 | 2.0 | 30.7 |
| DENV-2 | 61.4 | 2.3 | 26.7 |
| ZIKV | 61.4 | 1.8 | 34.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuang, F.-K.; Liao, C.-L.; Hu, M.-K.; Chiu, Y.-L.; Lee, A.-R.; Huang, S.-M.; Chiu, Y.-L.; Tsai, P.-L.; Su, B.-C.; Chang, T.-H.; et al. Antiviral Activity of Compound L3 against Dengue and Zika Viruses In Vitro and In Vivo. Int. J. Mol. Sci. 2020, 21, 4050. https://doi.org/10.3390/ijms21114050
Chuang F-K, Liao C-L, Hu M-K, Chiu Y-L, Lee A-R, Huang S-M, Chiu Y-L, Tsai P-L, Su B-C, Chang T-H, et al. Antiviral Activity of Compound L3 against Dengue and Zika Viruses In Vitro and In Vivo. International Journal of Molecular Sciences. 2020; 21(11):4050. https://doi.org/10.3390/ijms21114050
Chicago/Turabian StyleChuang, Fu-Kai, Ching-Len Liao, Ming-Kuan Hu, Yi-Lin Chiu, An-Rong Lee, Shih-Ming Huang, Yu-Lung Chiu, Pei-Ling Tsai, Bo-Cyuan Su, Tsung-Hsien Chang, and et al. 2020. "Antiviral Activity of Compound L3 against Dengue and Zika Viruses In Vitro and In Vivo" International Journal of Molecular Sciences 21, no. 11: 4050. https://doi.org/10.3390/ijms21114050
APA StyleChuang, F. -K., Liao, C. -L., Hu, M. -K., Chiu, Y. -L., Lee, A. -R., Huang, S. -M., Chiu, Y. -L., Tsai, P. -L., Su, B. -C., Chang, T. -H., Lin, C. -C., Shih, C. -C., & Yen, L. -C. (2020). Antiviral Activity of Compound L3 against Dengue and Zika Viruses In Vitro and In Vivo. International Journal of Molecular Sciences, 21(11), 4050. https://doi.org/10.3390/ijms21114050