Human Sperm Capacitation Involves the Regulation of the Tyr-Phosphorylation Level of the Anion Exchanger 1 (AE1)


 and
and 
Abstract
:1. Introduction
2. Results
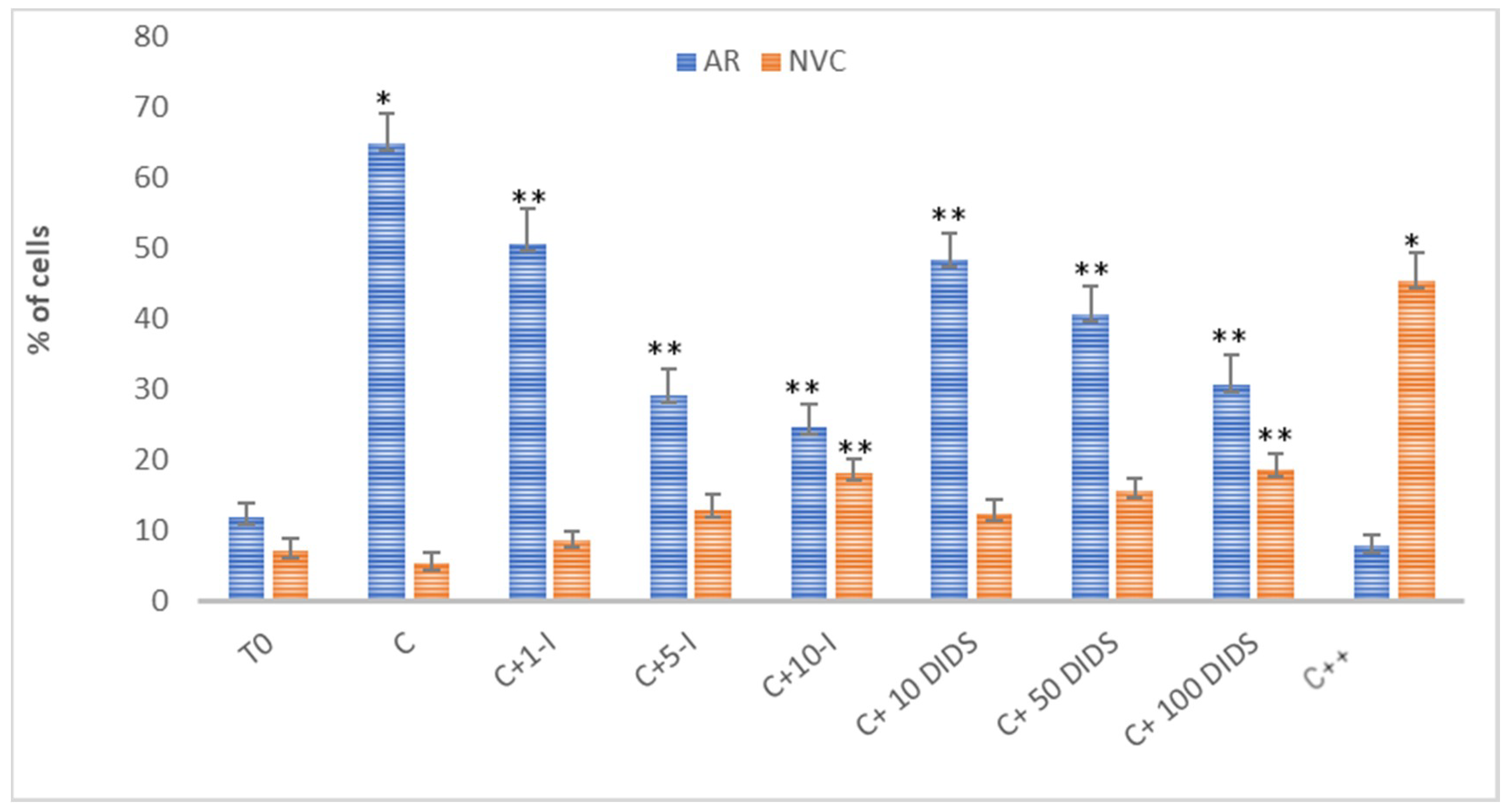
2.1. Effect of the Inhibition of CFTR and SLC4 Family Channels on Sperm AR and Viability
2.1.1. Effect of I-172 and DIDS on Acrosome Reaction (AR) and Sperm Viability
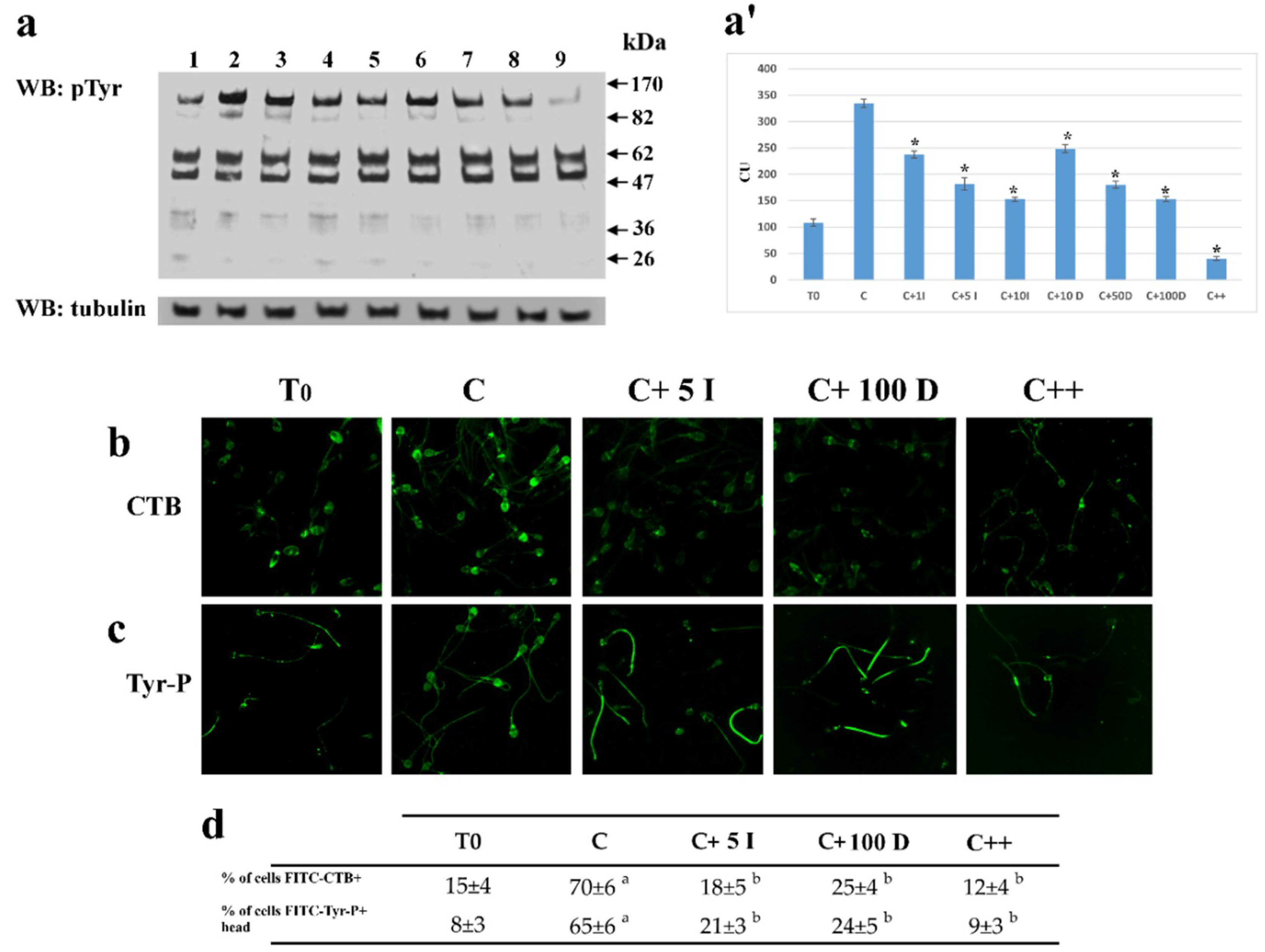
2.1.2. Effect of I-172 and DIDS on Tyr-P Level and Location as well as Membrane Rearrangement
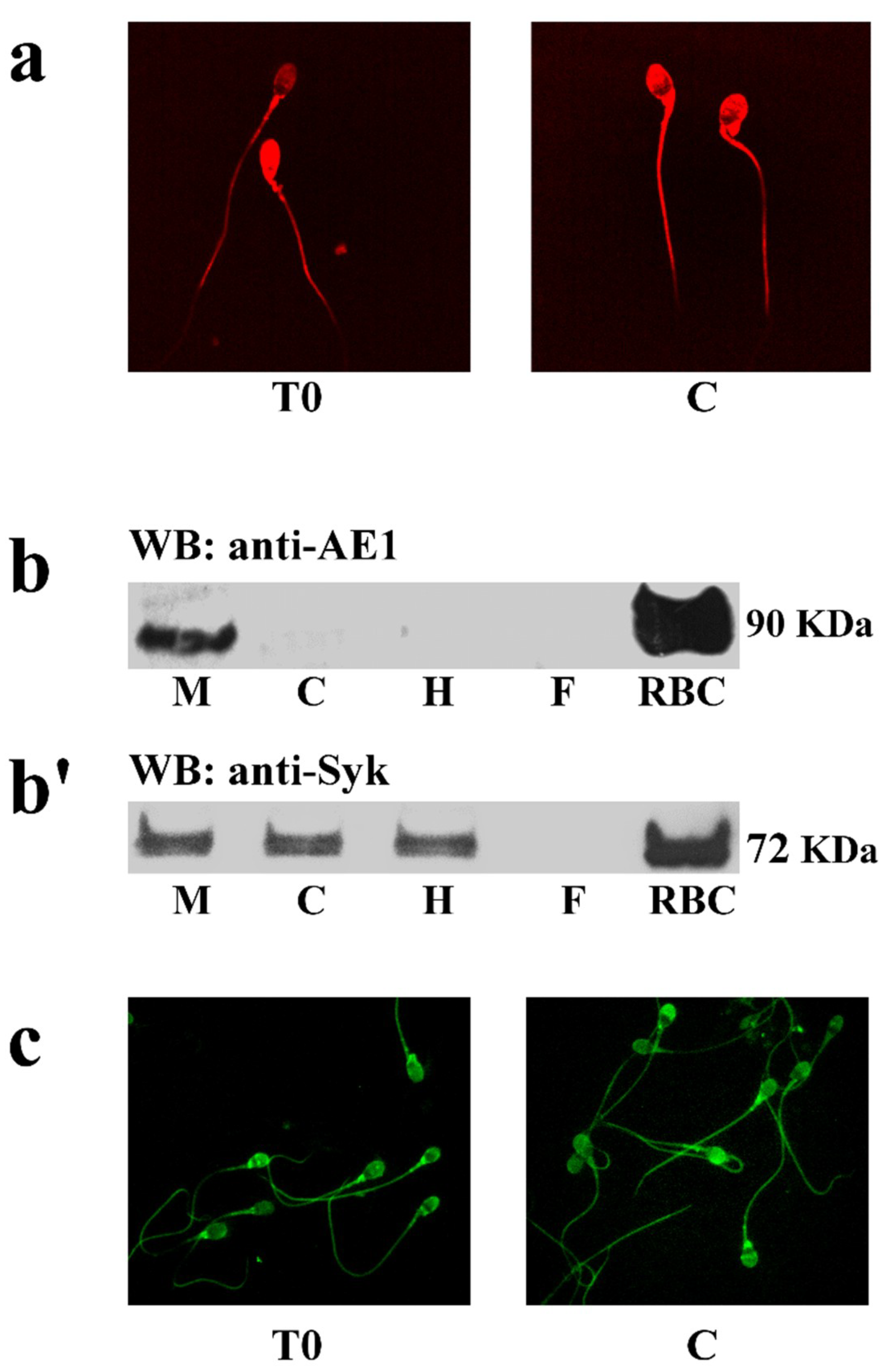
2.2. Identification and Localization of Sperm SLC4A1 (spAE1) and Protein Kinase Syk
2.3. Relationship between spAE1 Tyr-P and Capacitation
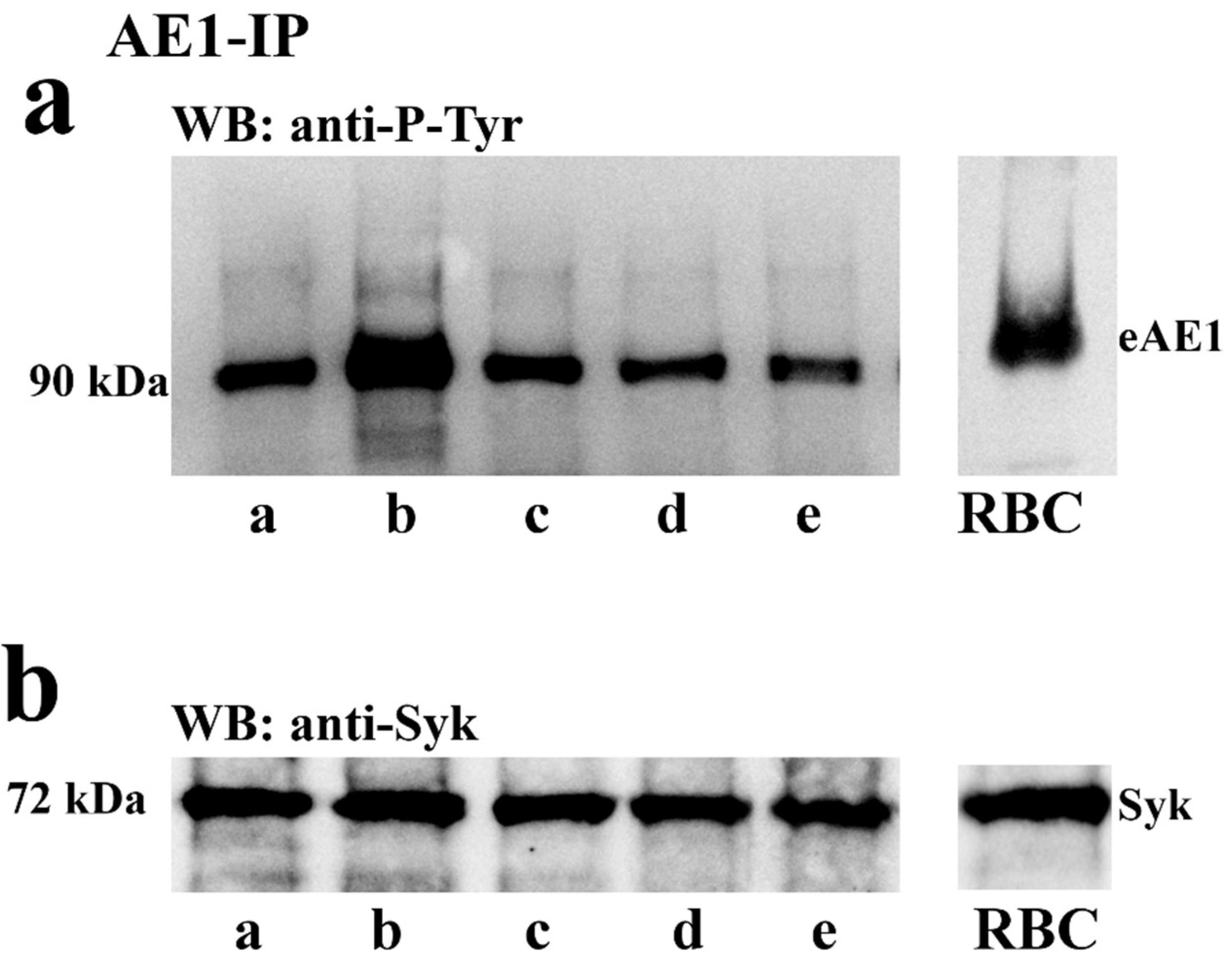
2.3.1. Effect of Capacitation and I-172 or DIDS on the Tyr-P Level of spAE1 and spAE1’s Interaction with Syk
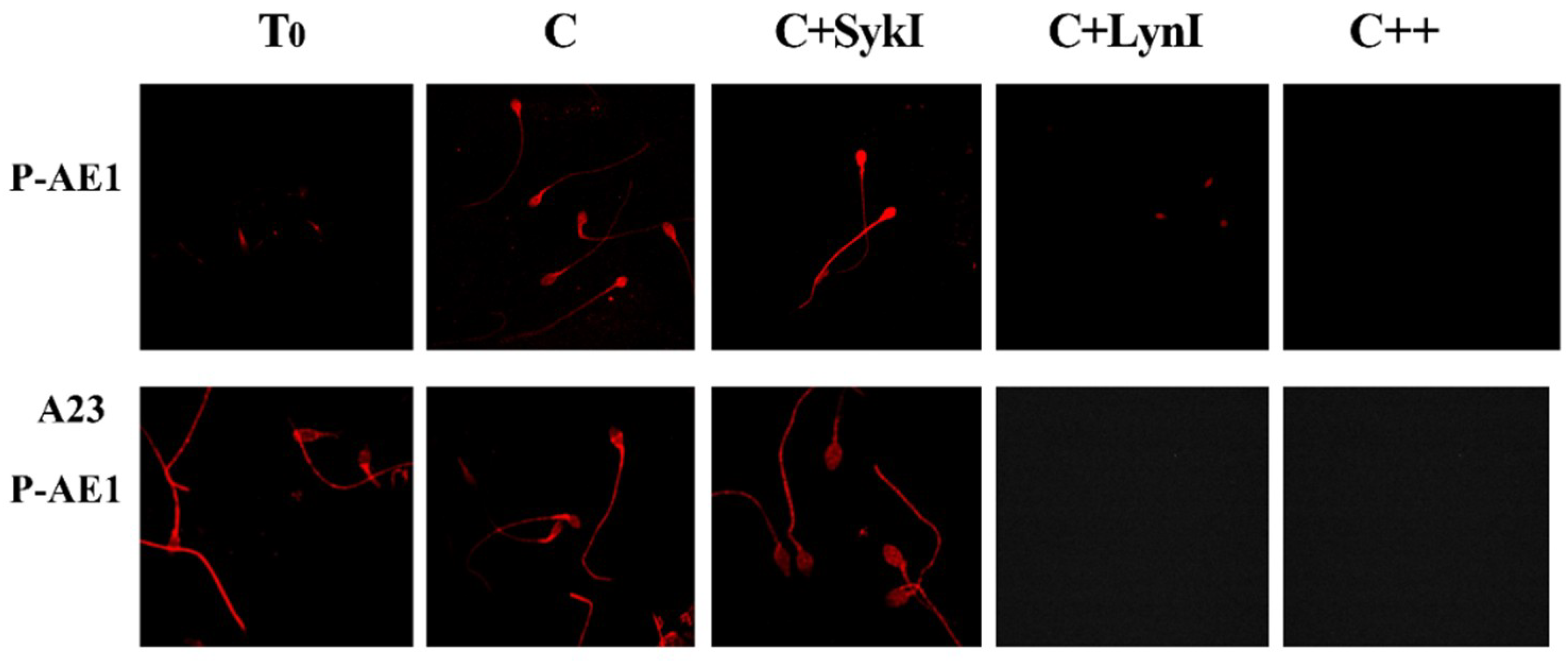
2.3.2. Effect of SykI and LynI on Capacitation (Tyr-P, CTB, P-AE1) and AR
2.3.3. Evaluation of Sperm Motility
3. Discussion
4. Materials and Methods
4.1. Semen Collection and Analysis
4.2. Chemicals
4.3. Sample Preparation
4.4. Computer Assisted Sperm Analysis (CASA)
4.5. Anti-Tyr-P and Anti-P-AE1 and Anti-Syk Immunofluorescence Evaluations
4.6. Evaluation of A23187-Induced Acrosome Reaction and Sperm Viability
4.7. Evaluation of Membrane Rearrangement
4.8. sp AE1 and Syk Localization and Interaction
4.8.1. Anti-AE1 (sp/eAE1-IP) and Anti-Syk Immunoprecipitations
4.8.2. spAE1 Localization in Sub-Cellular Fractions
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ARC | Acrosome Reacted Cells |
| NVC | Not viable Cells |
| CTB | Cholera Toxin beta subunit |
| SLC4A1 | Solute channel 4 A1 |
References
- Demarco, I.A.; Espinosa, F.; Edwards, J.; Sosnik, J.; de la Vega-Beltrán, J.L.; Hockensmith, J.W.; Kopf, G.S.; Darszon, A.; Visconti, P.E. Involvement of a Na+/HCO Cotransporter in Mouse Sperm Capacitation. J. Biol. Chem. 2003, 278, 7001–7009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buffone, M.G.; Wertheimer, E.V.; Visconti, P.E.; Krapf, D. Central role of soluble adenylyl cyclase and cAMP in sperm physiology. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2014, 1842, 2610–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Clark, E.N.; Florman, H.M. Sperm Membrane Potential: Hyperpolarization during Capacitation Regulates Zona Pellucida-Dependent Acrosomal Secretion. Dev. Biol. 1995, 171, 554–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alper, S.L. Molecular physiology and genetics of Na+-independent SLC4 anion exchangers. J. Exp. Biol. 2009, 212, 1672–1683. [Google Scholar] [CrossRef] [Green Version]
- Sindić, A.; Chang, M.-H.; Mount, D.B.; Romero, M.F. Renal physiology of SLC26 anion exchangers. Curr. Opin. Nephrol. Hypertens. 2007, 16, 484–490. [Google Scholar] [CrossRef]
- Ishiguro, H.; Steward, M.C.; Naruse, S.; Ko, S.B.H.; Goto, H.; Case, R.M.; Kondo, T.; Yamamoto, A. CFTR functions as a bicarbonate channel in pancreatic duct cells. J. Gen. Physiol. 2009, 133, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Bernardino, R.L.; Carrageta, D.F.; Sousa, M.; Alves, M.G.; Oliveira, P.F. pH and male fertility: Making sense on pH homeodynamics throughout the male reproductive tract. Cell. Mol. Life Sci. CMLS 2019, 76, 3783–3800. [Google Scholar] [CrossRef]
- Parkkila, S.; Rajaniemi, H.; Kellokumpu, S. Polarized Expression of a Band 3-Related Protein in Mammalian Sperm Cells. Biol. Reprod. 1993, 49, 326–331. [Google Scholar] [CrossRef] [Green Version]
- El Khouri, E.; Touré, A. Functional interaction of the cystic fibrosis transmembrane conductance regulator with members of the SLC26 family of anion transporters (SLC26A8 and SLC26A9): Physiological and pathophysiological relevance. Int. J. Biochem. Cell Biol. 2014, 52, 58–67. [Google Scholar] [CrossRef]
- Puga Molina, L.C.; Pinto, N.A.; Torres, N.I.; González-Cota, A.L.; Luque, G.M.; Balestrini, P.A.; Romarowski, A.; Krapf, D.; Santi, C.M.; Treviño, C.L.; et al. CFTR/ENaC-dependent regulation of membrane potential during human sperm capacitation is initiated by bicarbonate uptake through NBC. J. Biol. Chem. 2018, 293, 9924–9936. [Google Scholar] [CrossRef] [Green Version]
- Touré, A. Importance of SLC26 Transmembrane Anion Exchangers in Sperm Post-testicular Maturation and Fertilization Potential. Front. Cell Dev. Biol. 2019, 7, 230. [Google Scholar] [CrossRef] [PubMed]
- Sorum, B.; Czégé, D.; Csanády, L. Timing of CFTR Pore Opening and Structure of Its Transition State. Cell 2015, 163, 724–733. [Google Scholar] [CrossRef] [Green Version]
- Puga Molina, L.C.; Pinto, N.A.; Torres Rodríguez, P.; Romarowski, A.; Vicens Sanchez, A.; Visconti, P.E.; Darszon, A.; Treviño, C.L.; Buffone, M.G. Essential Role of CFTR in PKA-Dependent Phosphorylation, Alkalinization, and Hyperpolarization During Human Sperm Capacitation. J. Cell. Physiol. 2017, 232, 1404–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardino, R.; Jesus, T.; Martins, A.; Sousa, M.; Barros, A.; Cavaco, J.; Socorro, S.; Alves, M.; Oliveira, P. Molecular Basis of Bicarbonate Membrane Transport in the Male Reproductive Tract. Curr. Med. Chem. 2013, 20, 4037–4049. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Low, P.S. Mapping of glycolytic enzyme-binding sites on human erythrocyte band 3. Biochem. J. 2006, 400, 143–151. [Google Scholar] [CrossRef]
- Chu, H.; Breite, A.; Ciraolo, P.; Franco, R.S.; Low, P.S. Characterization of the deoxyhemoglobin binding site on human erythrocyte band 3: Implications for O2 regulation of erythrocyte properties. Blood 2008, 111, 932–938. [Google Scholar] [CrossRef] [Green Version]
- Brunati, A.M.; Bordin, L.; Clari, G.; James, P.; Quadroni, M.; Baritono, E.; Pinna, L.A.; Donella-Deana, A. Sequential phosphorylation of protein band 3 by Syk and Lyn tyrosine kinases in intact human erythrocytes: Identification of primary and secondary phosphorylation sites. Blood 2000, 96, 1550–1557. [Google Scholar] [CrossRef]
- Brunati, A.M.; Bordin, L.; Clari, G.; Moret, V. The Lyn-catalyzed Tyr phosphorylation of the transmembrane band-3 protein of human erythrocytes. Eur. J. Biochem. 1996, 240, 394–399. [Google Scholar] [CrossRef]
- Leclerc, P.; de Lamirande, E.; Gagnon, C. Regulation of protein-tyrosine phosphorylation and human sperm capacitation by reactive oxygen derivatives. Free Radic. Biol. Med. 1997, 22, 643–656. [Google Scholar] [CrossRef]
- Donà, G.; Fiore, C.; Tibaldi, E.; Frezzato, F.; Andrisani, A.; Ambrosini, G.; Fiorentin, D.; Armanini, D.; Bordin, L.; Clari, G. Endogenous reactive oxygen species content and modulation of tyrosine phosphorylation during sperm capacitation: ROS, Tyr-phosphorylation and capacitation in human sperms. Int. J. Androl. 2011, 34, 411–419. [Google Scholar] [CrossRef]
- Varano, G.; Lombardi, A.; Cantini, G.; Forti, G.; Baldi, E.; Luconi, M. Src activation triggers capacitation and acrosome reaction but not motility in human spermatozoa. Hum. Reprod. 2008, 23, 2652–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrisani, A.; Donà, G.; Ambrosini, G.; Bonanni, G.; Bragadin, M.; Cosmi, E.; Clari, G.; Armanini, D.; Bordin, L. Effect of various commercial buffers on sperm viability and capacitation. Syst. Biol. Reprod. Med. 2014, 60, 239–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrisani, A.; Donà, G.; Tibaldi, E.; Brunati, A.; Sabbadin, C.; Armanini, D.; Alvisi, G.; Gizzo, S.; Ambrosini, G.; Ragazzi, E.; et al. Astaxanthin Improves Human Sperm Capacitation by Inducing Lyn Displacement and Activation. Mar. Drugs 2015, 13, 5533–5551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-Y.; Jia, Y.-P.; Duan, L.-Y.; Li, K.-M. Participation of the inositol 1,4,5-trisphosphate-gated calcium channel in the zona pellucida- and progesterone-induced acrosome reaction and calcium influx in human spermatozoa. Asian J. Androl. 2020, 22, 192. [Google Scholar] [CrossRef]
- Romero, M.F.; Chen, A.-P.; Parker, M.D.; Boron, W.F. The SLC4 family of bicarbonate transporters. Mol. Aspects Med. 2013, 34, 159–182. [Google Scholar] [CrossRef] [Green Version]
- Romero, M.F.; Fulton, C.M.; Boron, W.F. The SLC4 family of HCO3− transporters. Pflügers Arch. Eur. J. Physiol. 2004, 447, 495–509. [Google Scholar] [CrossRef]
- Shin, J.-H.; Son, E.J.; Lee, H.S.; Kim, S.J.; Kim, K.; Choi, J.Y.; Lee, M.G.; Yoon, J.-H. Molecular and functional expression of anion exchangers in cultured normal human nasal epithelial cells. Acta Physiol. 2007, 191, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Mukaibo, T.; Gao, X.; Kurtz, I.; Melvin, J.E. Slc4a11 disruption causes duct cell loss and impairs NaCl reabsorption in female mouse submandibular glands. Physiol. Rep. 2019, 7, e14232. [Google Scholar] [CrossRef] [Green Version]
- Donà, G.; Fiore, C.; Andrisani, A.; Ambrosini, G.; Brunati, A.; Ragazzi, E.; Armanini, D.; Bordin, L.; Clari, G. Evaluation of correct endogenous reactive oxygen species content for human sperm capacitation and involvement of the NADPH oxidase system. Hum. Reprod. 2011, 26, 3264–3273. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.-W.; Li, Y.; Zou, L.-L.; Guan, Y.-T.; Peng, S.; Zheng, L.-X.; Deng, S.-M.; Zhu, L.-Y.; Wang, L.-W.; Chen, L.-X. Chloride channels are involved in sperm motility and are downregulated in spermatozoa from patients with asthenozoospermia. Asian J. Androl. 2017, 19, 418. [Google Scholar] [CrossRef]
- Donà, G.; Kožuh, I.; Brunati, A.; Andrisani, A.; Ambrosini, G.; Bonanni, G.; Ragazzi, E.; Armanini, D.; Clari, G.; Bordin, L. Effect of Astaxanthin on Human Sperm Capacitation. Mar. Drugs 2013, 11, 1909–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nixon, B.; Mitchell, L.A.; Anderson, A.L.; Mclaughlin, E.A.; O’bryan, M.K.; Aitken, R.J. Proteomic and functional analysis of human sperm detergent resistant membranes. J. Cell. Physiol. 2011, 226, 2651–2665. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Majzoub, A.; Esteves, S.C.; Ko, E.; Ramasamy, R.; Zini, A. Clinical utility of sperm DNA fragmentation testing: Practice recommendations based on clinical scenarios. Transl. Androl. Urol. 2016, 5, 935–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farinha, C.M.; Swiatecka-Urban, A.; Brautigan, D.L.; Jordan, P. Regulatory Crosstalk by Protein Kinases on CFTR Trafficking and Activity. Front. Chem. 2016, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Baggio, B.; Bordin, L.; Gambaro, G.; Piccoli, A.; Marzaro, G.; Clari, G. Evidence of a link between erythrocyte band 3 phosphorylation and anion transport in patients with “idiopathic” calcium oxalate nephrolithiasis. Miner. Electrolyte Metab. 1993, 19, 17–20. [Google Scholar]
- Baggio, B.; Priante, G.; Brunati, A.M.; Clari, G.; Bordin, L. Specific modulatory effect of arachidonic acid on human red blood cell oxalate transport: Clinical implications in calcium oxalate nephrolithiasis. J. Am. Soc. Nephrol. JASN 1999, 10 (Suppl. S14), S381–S384. [Google Scholar]
- Baggio, B.; Bordin, L.; Clari, G.; Gambaro, G.; Moret, V. Functional correlation between the Ser/Thr-phosphorylation of band-3 and band-3-mediated transmembrane anion transport in human erythrocytes. Biochim. Biophys. Acta 1993, 1148, 157–160. [Google Scholar] [CrossRef]
- Ferru, E.; Giger, K.; Pantaleo, A.; Campanella, E.; Grey, J.; Ritchie, K.; Vono, R.; Turrini, F.; Low, P.S. Regulation of membrane-cytoskeletal interactions by tyrosine phosphorylation of erythrocyte band 3. Blood 2011, 117, 5998–6006. [Google Scholar] [CrossRef] [Green Version]
- Breitbart, H.; Finkelstein, M. Regulation of Sperm Capacitation and the Acrosome Reaction by PIP2 and Actin Modulation. Asian J. Androl. 2015, 17, 597. [Google Scholar] [CrossRef]
- Okamura, N.; Tajima, Y.; Ishikawa, H.; Yoshii, S.; Koiso, K.; Sugita, Y. Lowered levels of bicarbonate in seminal plasma cause the poor sperm motility in human infertile patients. Fertil. Steril. 1986, 45, 265–272. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization: Geneva, Switzerland, 2010; ISBN 978-92-4-154778-9. [Google Scholar]
- Zonta, F.; Pagano, M.A.; Trentin, L.; Tibaldi, E.; Frezzato, F.; Trimarco, V.; Facco, M.; Zagotto, G.; Pavan, V.; Ribaudo, G.; et al. Lyn sustains oncogenic signaling in chronic lymphocytic leukemia by strengthening SET-mediated inhibition of PP2A. Blood 2015, 125, 3747–3755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Fujikura, K.; Higashiyama, T.; Takata, K. DNA Staining for Fluorescence and Laser Confocal Microscopy. J. Histochem. Cytochem. 1997, 45, 49–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, R.; Fukuda, M.; Takeuchi, H.; Itoh, S.; Takada, M. Flow Cytometry to Evaluate Acrosome-Reacted Sperm. Arch. Androl. 1990, 25, 243–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, C.; Bagatolli, L.A.; Volovyk, Z.N.; Thompson, N.L.; Levi, M.; Jacobson, K.; Gratton, E. Lipid Rafts Reconstituted in Model Membranes. Biophys. J. 2001, 80, 1417–1428. [Google Scholar] [CrossRef] [Green Version]
- Amaral, A.; Castillo, J.; Estanyol, J.M.; Ballescà, J.L.; Ramalho-Santos, J.; Oliva, R. Human Sperm Tail Proteome Suggests New Endogenous Metabolic Pathways. Mol. Cell. Proteom. 2013, 12, 330–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tyr-P (%) | CTB (%) | P-AE1 (%) | A23-P-AE1 (%) | ARC (%) | NVC (%) | |
|---|---|---|---|---|---|---|
| Head | Head | Head | Head | |||
| T0 | 18.2 ± 2.1 | 15.4 ± 1.7 | 9.2 ± 1.2 | 10.8 ± 1.2 | 11.3 ± 2.3 | 7.1 ± 2.3 |
| C | 66.3 ± 2.9 * | 67.0 ± 3.6 * | 63.1 ± 2.5 * | 68.9 ± 7.5 * | 69.7 ± 5.2 * | 9.1 ± 1.2 * |
| C+SykI | 58.7 ± 2.3 b | 55.5 ± 3.5 b | 85.5 ± 12.7 b | 89.7 ± 7.7 b | 52.9 ± 3.2 a | 12.3 ± 1.1 b |
| C+LynI | 21.3 ± 1.7 a | 23.3 ± 4.2 a | 16.3 ± 3.5 a | 15.3 ± 4.6 a | 20.4 ± 4.1 a | 60.6 ± 1.3 b |
| C++ | 10.2 ± 1.3 a | 12.8 ± 3.5 a | 9.9 ± 2.2 a | 8.5 ± 4.2 a | 14.6 ± 3.3 a | 65.4 ± 3.2 b |
| Motility (%) | VSL (µm/s) | VAP (µm/s) | ALH (µm) | |
|---|---|---|---|---|
| T0 | 69 ± 3 | 58.4 ± 8.9 | 54.0 ± 6.7 | 3.1 ± 0.5 |
| C | 76 ± 7 | 77.8 ± 13.9 a | 69.6 ± 11.0 a | 4.9 ± 0.9 a |
| C+Syk- | 74 ± 8 | 76.9 ± 10.4 a | 67.3 ± 9.0 a | 4.8 ± 0.8 a |
| C+LynI | 74 ± 12 | 76.3 ± 9.4 a | 68.3 ± 9.5 a | 4.7 ± 0.5 a |
| C++ | 72 ± 6 | 75.9 ± 12.7 a | 67.6± 9.8 a | 4.9 ± 0.7 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donà, G.; Tibaldi, E.; Andrisani, A.; Ambrosini, G.; Sabbadin, C.; Pagano, M.A.; Brunati, A.M.; Armanini, D.; Ragazzi, E.; Bordin, L. Human Sperm Capacitation Involves the Regulation of the Tyr-Phosphorylation Level of the Anion Exchanger 1 (AE1). Int. J. Mol. Sci. 2020, 21, 4063. https://doi.org/10.3390/ijms21114063
Donà G, Tibaldi E, Andrisani A, Ambrosini G, Sabbadin C, Pagano MA, Brunati AM, Armanini D, Ragazzi E, Bordin L. Human Sperm Capacitation Involves the Regulation of the Tyr-Phosphorylation Level of the Anion Exchanger 1 (AE1). International Journal of Molecular Sciences. 2020; 21(11):4063. https://doi.org/10.3390/ijms21114063
Chicago/Turabian StyleDonà, Gabriella, Elena Tibaldi, Alessandra Andrisani, Guido Ambrosini, Chiara Sabbadin, Mario Angelo Pagano, Anna Maria Brunati, Decio Armanini, Eugenio Ragazzi, and Luciana Bordin. 2020. "Human Sperm Capacitation Involves the Regulation of the Tyr-Phosphorylation Level of the Anion Exchanger 1 (AE1)" International Journal of Molecular Sciences 21, no. 11: 4063. https://doi.org/10.3390/ijms21114063
APA StyleDonà, G., Tibaldi, E., Andrisani, A., Ambrosini, G., Sabbadin, C., Pagano, M. A., Brunati, A. M., Armanini, D., Ragazzi, E., & Bordin, L. (2020). Human Sperm Capacitation Involves the Regulation of the Tyr-Phosphorylation Level of the Anion Exchanger 1 (AE1). International Journal of Molecular Sciences, 21(11), 4063. https://doi.org/10.3390/ijms21114063