Fatty Acid Diets: Regulation of Gut Microbiota Composition and Obesity and Its Related Metabolic Dysbiosis


 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Medium-Chain Fatty Acids
3. Long-Chain Fatty Acids
4. Monounsaturated Fatty Acids
5. Polyunsaturated Fatty Acids
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hawkesworth, S.; Dangour, A.D.; Johnston, D.; Lock, K.; Poole, N.; Rushton, J.; Uauy, R.; Waae, J. Feeding the world healthily: The challenge of measuring the effects of agriculture on health. Phil. Trans. R. Soc. B. 2010, 365, 3083–3097. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Functional roles of fatty acids and their effects on human health. J. Parenter. Enteral. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef] [PubMed]
- FAO. Fats and fatty acids in human nutrition. In Report of an Expert Consultation; FAO Food and Nutrition Paper 91; FAO: Rome, Italy, 2010; ISBN 978-92-5-106733-8. [Google Scholar]
- Albahrani, A.A.; Greaves, R.F. Fat-soluble vitamins: Clinical indications and current challenges for chromatographic measurement. Clin. Biochem. Ver. 2016, 37, 27–47. [Google Scholar]
- Abedi, E.; Sahari, M.A. Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties. Food Sc. Nutr. 2014, 2, 443–463. [Google Scholar] [CrossRef] [PubMed]
- Choquet, H.; Meyre, D. Genetics of obesity: What have we learned? Curr. Genom. 2011, 12, 169–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwood, H.C.; Bloom, S.R.; Murphy, K.G. Peptides and their potential role in the treatment of diabetes and obesity. Rev. Diabet. Stud. 2011, 8, 355–368. [Google Scholar] [CrossRef] [Green Version]
- Camacho, S.; Ruppel, A. Is the calorie concept a real solution to the obesity epidemic? Global Health Action 2017, 10, 1289650. [Google Scholar] [CrossRef] [Green Version]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [Green Version]
- Lecocq, M.; Detry, B.; Guisset, A.; Pilette, C. FcαRI-mediated inhibition of IL-12 production and priming by IFN-γ of human monocytes and dendritic cells. J. Immunol. 2013, 190, 2362–2371. [Google Scholar] [CrossRef] [Green Version]
- Bourlioux, P.; Koletzko, B.; Guarner, F.; Braesco, V. The intestine and its microflora are partners for the protection of the host: Report on the danone symposium “The intelligent intestine”, held in Paris, June 14, 2002. Am. J. Clin. Nutr. 2003, 78, 675–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajoka, M.S.R.; Shi, J.; Mehwish, H.M.; Zhu, J.; Li, Q.; Shao, D.; Huang, Q.; Yang, H. Interaction between diet composition and gut microbiota and its impact on gastrointestinal tract health. Food Sci. Hum. Wellness 2017, 6, 121–130. [Google Scholar] [CrossRef]
- Cummings, J.H.; Macfarlane, G.T. The control and consequences of bacterial fermentation in the human colon. J. Appl. Bacteriol. 1991, 70, 443–459. [Google Scholar] [CrossRef] [PubMed]
- Ohira, H.; Tsutsui, W.; Fujioka, Y. Are short chain fatty acids in gut microbiota defensive players for inflammation and atherosclerosis? J. Atheroscler. Thromb. 2017, 24, 660–672. [Google Scholar] [CrossRef] [Green Version]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef] [Green Version]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef]
- Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; Leitch, C.M.; Scott, K.P.; Flint, H.; Louis, P. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014, 8, 1323. [Google Scholar] [CrossRef] [Green Version]
- Louis, P.; Duncan, S.H.; McCrae, S.I.; Millar, J.; Jackson, M.S.; Flint, H.J. Restricted distribution of the butyrate kinase pathway among butyrate-production bacteria from the human colon. J. Bacterol. 2004, 186, 2099–2106. [Google Scholar] [CrossRef] [Green Version]
- Roediger, W.E. Role of anaerobic bacteria in the metabolic welfare of the colonic mucosa in man. Gut 1980, 21, 793–798. [Google Scholar] [CrossRef] [Green Version]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [Green Version]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakkaer, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.M.; Yu, Y.N.; Wang, J.L.; Lin, Y.W.; Kong, X.; Yang, C.Q.; Yang, L.; Liu, Z.J.; Yuan, Y.Z.; Liu, F.; et al. Decreased dietary fiber intake and structural alteration of gut microbiota in patients with advanced colorectal adenoma. Am. J. Clin. Nutr. 2013, 97, 1044–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graf, D.; Cagno, R.D.; Fak, F.; Flint, H.J.; Nyman, M.; Saarela, M.; Watzl, B. Contribution of diet to the composition of the human gut microbiota. Microb. Ecol. Health Dis. 2015, 26, 26164. [Google Scholar] [CrossRef] [PubMed]
- Alou, M.T.; Lagier, J.C.; Roaoult, D. Diet influence on the gut microbiota and dysbiosis related to nutritional disorders. Human Microbiome Journal 2016, 1, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Bibbò, S.; Ianiro, G.; Giorgio, V.; Scaldaferri, F.; Masucci, L.; Gasbarrini, A.; Cammarota, G. The role of diet on gut microbiota composition. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4742–4749. [Google Scholar]
- Elmadfa, I.; Freisling, H. Fat intake, diet variety and health promotion. Forum Nutr. 2005, 57, 1–10. [Google Scholar]
- Honors, M.A.; Harnack, L.J.; Zhou, X.; Steffen, L.M. Trends in fatty acid intake of adults in the Minneapolis-St Paul, MN metropolitan area, 1980–1982 through 2007 – 2009. J. Am. Heart Assoc. 2014, 3, e001023. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, P.S.; Inada, A.C.; Marcelino, G.; Cardozo, C.M.L.; Freitas, K.C.; Guimarães, R.C.A.; Castro, A.P.; Nascimento, V.A.; Hiane, P.A. Fatty acids consumption: The role Metabolic aspects involved in obesity and its associated disorders. Nutrients 2017, 9, 1158. [Google Scholar] [CrossRef] [Green Version]
- Forouhi, N.G.; Krauss, R.M.; Taubes, G.; Willett, W. Dietary fat and cardiometabolic health: Evidence, controversies, and consensus for guidance. BMJ 2018, 361, k2139. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.H.; Kang, S.G.; Park, J.H.; Yanagisawa, M.; Kim, C.H. Short-chain fatty acids activate GPR41 and GPR43 on intestinal epithelial cells to promote inflammatory responses in mice. Gastroenterology 2013, 145, 396–406. [Google Scholar] [CrossRef]
- Lyons, C.L.; Kennedy, E.B.; Roche, H.M. Metabolic inflammation-differential modulation by dietary constituents. Nutrients 2016, 8, 247. [Google Scholar] [CrossRef] [PubMed]
- Goossens, G.H. The metabolic phenotype in obesity: Fat mass, body fat distribution, and adipose tissue function. Obes. Facts 2017, 10, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Tvrszicka, E.; Kremmyda, L.S.; Stankova, B.; Zak, A. Fatty acids as biocompounds: Their role in human metabolism, health and disease–a review. Part 1: Classification, dietary sources and biological functions. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc. Czech. Repub. 2011, 155, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Francois, C.A.; Cannor, S.L.; Wander, R.C.; Connor, W.E. Acute effects of dietary fatty acids on the fatty acids of human milk. Am. J. Clin. Nutr. 1998, 67, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Kok, S.; Ong-Abdullah, M.; Ee, C.G.; Namasivayam, P. Comparison of nutrient composition in kernel of tenera and clonal materials of oil palm (Elaeis guineensis Jacq.). Food Chem. 2011, 129, 1343–1347. [Google Scholar] [CrossRef]
- Orsavova, J.; Misurcova, L.; Ambrozova, J.V.; Vicha, R.; Mlcek, J. Fatty acids composition of vegetable oils and its contribution to dietary energy intake and dependence of cardiovascular mortality on dietary intake of fatty acids. Int. J. Mol. Sci. 2015, 16, 12871–12890. [Google Scholar] [CrossRef]
- Gardner, A.S.; Rahman, I.A.; Lai, C.T.; Hepworth, A.; Trengove, N.; Hartmann, P.E.; Geddes, D.T. Changes in fatty acid composition of human milk in response to cold-like symptoms in the lactating mother and infant. Nutrients 2017, 9, 1034. [Google Scholar] [CrossRef]
- Thomson, A.B.R.; Garg, M.K.M.L.; Clandinin, M.T. Intestinal aspects of absorption: In review. Can. J. Physiol. Pharmacol. 1989, 67, 179–191. [Google Scholar] [CrossRef]
- Decker, E.A. The role of stereospecific saturated fatty acid position on lipid nutrition. Nutr. Rev. 1996, 54, 108–110. [Google Scholar] [CrossRef]
- Briggs, M.A.; Petersen, K.S.; Kris-Etherton, P.M. Saturated fatty acids and cardiovascular disease: Replacements for saturated fat to reduce cardiovascular risk. Healthcare (Basel) 2017, 5, 29. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.O.; Peters, J.C.; Swift, L.L.; Yang, D.; Sharp, T.; Abumrad, N.; Greene, H.L. Changes in blood lipids during six days of overfeeding with medium or long chain triglycerides. J. Lipid Res. 1990, 31, 407–416. [Google Scholar] [PubMed]
- Turner, N.; Hariharan, K.; TidAng, J.; Frangioudakis, G.; Beale, S.M.; Wright, L.E.; Zeng, X.Y.; Leslie, S.J.; Li, J.Y.; Kraegen, E.W. Enhancement of muscle mitochondrial oxidative capacity and alterations in insulin action are lipid species dependent. Diabetes 2009, 58, 2547–2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, N.D.; Limketkai, B.N. The Use of Medium-Chain Triglycerides in Gastrointestinal Disorders. Pract. Gastroenterol. 2017, 41, 20–28. [Google Scholar]
- Wang, Y.; Liu, Z.; Han, Y.; Xu, J.; Haung, W.; Li, Z. Medium chain triglycerides enhances exercise endurance through the increased mitochondrial biogenesis and metabolism. PLoS ONE 2018, 13, e0191182. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Wan, Y.; Jacoby, J.; Jiang, Y.; Zhang, Y.; Yu, L. Effects of medium- and long-chain triacylglicerols on lipid metabolism and gut microbiota composition in C57BL/6L mice. J. Agric. Food Chem. 2017, 65, 6599–6607. [Google Scholar] [CrossRef]
- Djurasevic, S.; Bojic, S.; Nikolic, B.; Dimkic, I.; Todorovic, Z.; Djordjevic, J.; Mitic-Culafic, D. Beneficial effect of virgin coconut oil on alloxan-induced diabetes and microbiota composition in rats. Plant Foods Hum. Nutr. 2018, 73, 295–301. [Google Scholar] [CrossRef]
- Dias, M.M.; Siqueira, N.P.; Conceição, L.L.; Reis, S.A.; Valente, F.X.; Dias, M.M.S.; Rosa, C.O.B.; Paula, S.O.; Matta, S.L.P.; Oliveira, L.L.; et al. Consumption of virgin coconut oil in Wistar rats increases saturated fatty acids in the liver and adipose tissue, as well as adipose tissue inflammation. J. Funct. Foods 2018, 48, 472–480. [Google Scholar] [CrossRef]
- Patrone, V.; Minuti, A.; Lizier, M.; Miragoli, F.; Lucchini, F.; Trevisi, E.; Rossi, F.; Callegari, M.L. Differential effects of coconut versus soy oil on gut microbiota composition and predicted metabolic function in adult mice. BMC Genomics 2018, 19, 808. [Google Scholar] [CrossRef]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Li, S.; Gan, R.Y.; Zhou, T.; Xu, D.P.; Li, H.B. Impacts of gut bacteria on human health and diseases. Int. J. Mol. Sci. 2015, 16, 7493–7519. [Google Scholar] [CrossRef]
- Sanmiguel, C.; Gupta, A.; Mayer, E.A. Gut microbiome and obesity: A plausible explanation for obesity. Curr. Obes. Rep. 2015, 4, 250–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azad, A.K.; Sarker, M.; Li, T.; Yin, J. Probiotic species in the modulation of gut microbiota: An overview. BioMed Res. Int. 2018, 9478630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ossa, J.C.; Yáñez, D.; Valenzuela, R.; Gallardo, P.; Lucero, Y.; Farfán, M.J. Intestinal Inflammation in chilean infants fed with bovine formula vs. breast milk and its association with their gut microbiota. Front. Cell. Infect. Microbiol. 2018, 8, 190. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.Y.; Lee, Y.; Lu, H.; Chou, C.H.; Wang, C. Analysis of gut microbiota and the effect of lauric acid against necrotic enteritis in Clostridium perfringens and Eimeria side-by-side challenge model. PLoS ONE 2019, 14, e0205784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, J. Ecological role of lactobacilli in the gastrointestinal tract: Implications for fundamental and biomedical research. Appl. Environ. Microbiol. 2008, 74, 4985–4996. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–549. [Google Scholar] [CrossRef]
- Mendonça, M.A.; Araújo, W.M.C.; Borgo, L.A.; Alencar, E.R. Lipid profile of different infant formulas for infants. PLoS ONE 2017, 12, e0177812. [Google Scholar] [CrossRef] [Green Version]
- Mazzocchi, A.; D’Oria, V.; de Cosmi, V.; Bettocchi, S.; Milani, G.P.; Silano, M.; Agostoni, C. The role of lipids in human milk and infant formulae. Nutrients 2018, 10, 567. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes. 2018, 9, 304–325. [Google Scholar] [CrossRef] [Green Version]
- Sousa, F.P.; Silva, L.N.; Rezende, D.B.L.; Oliveira, L.C.A.; Pasa, V.M.D. Simultaneous deoxygenation, cracking and isomerization of palm kernel oil and palm olein over beta zeolite to produce biogasoline, green diesel and biojet-fuel. Fuel 2018, 223, 149–152. [Google Scholar] [CrossRef]
- Mohdaly, A.A.E.R.; Seliem, K.A.E.H.; El-Hassan, A.E.M.M.A. Effect of Refining Process on the Quality Characteristics of Soybean and Cotton seed Oils. Int. J. Curr. Microbiol. App. Sci. 2017, 6, 207–222. [Google Scholar] [CrossRef]
- Sette, S.; Le Donne, C.; Piccinelli, R.; Arcella, D.; Turrini, A.; Leclercq, C.; Arcella, D.; Bevilacqua, N.; Buonocore, P.; Capriotti, M.; et al. The third Italian national food consumption survey, INRAN-SCAI 2005-06–part 1: Nutrient intakes in Italy. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.M.; Neville, M.J.; Hoppa, M.B.; Frayn, K.N. De novo lipogenesis and stearoyl-CoA desaturase are coordinately regulated in the human adipocyte and protect against palmitate-induced cell injury. J. Biol. Chem. 2010, 285, 6044–6052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silbernagel, G.; Kovarova, M.; Cegan, A.; Machann, J.; Schick, F.; Lehmann, R.; Häring, H.U.; Stefan, N.; Schleicher, E.; Fritsche, A.; et al. High hepatic SCD1 activity is associated with low liver fat content in healthy subjects under a lipogenic diet. J. Clin. Endocrinol. Metab. 2012, 97, E2288–E2292. [Google Scholar] [CrossRef]
- De Wit, N.; Derrien, M.; Bosch-Vermeulen, H.; Oosterink, E.; Keshtkar, S.; Duval, C.; de Vogel-van den Bosch, J.; Kleerebezem, M.; Müller, M.; van der Meer, R. Saturated fat stimulates obesity and hepatic steatosis and affects gut microbiota composition by an enhanced overflow of dietary fat to the distal intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G589–G599. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.A.; Gu, W.; Lee, I.A.; Joh, E.H.; Kim, D.H. High fat diet-induced gut microbiota exacerbates inflammation and obesity in mice via TLR4 signaling pathway. PLoS ONE 2012, 7, e47713. [Google Scholar] [CrossRef]
- Huang, E.Y.; Leone, V.A.; Devkota, S.; Wang, Y.; Brady, M.J.; Chang, E.B. Composition of dietary fat source shape gut microbiota architecture and alters host inflammatory mediators in mouse adipose tissue. JPEN J. Parenter. Enteral Nutr. 2013, 37, 746–754. [Google Scholar] [CrossRef]
- Patterson, E.; O’Doherty, R.M.; Murphy, E.F.; Wall, R.; O’Sullivan, O.; Nilaweera, K.; Fitzgerald, G.F.; Cotter, P.D.; Ross, R.P.; Stanton, C. Impact of dietary fatty acids on metabolic activity and host intestinal microbiota composition in C57BL/6J mice. Br. J. Nutr. 2014, 111, 1905–1917. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Wolf, P.G.; Carbonero, F.; Zhong, W.; Reid, T.; Gaskins, R.; McIntosh, M.K. Intestinal and systemic inflammatory responses are positively associated with sulfidogenic bacteria abundance in high-fat-fed male C57BL/6J mice. J. Nutr. 2014, 144, 1181–1187. [Google Scholar] [CrossRef] [Green Version]
- Lam, Y.Y.; Ha, C.W.Y.; Hoffmann, J.M.A.; Oscarsson, J.; Dinudom, A.; Mather, T.J.; Cook, D.I.; Hunt, N.H.; Caterson, I.D.; Holmes, A.J.; et al. Effects of dietary fat profile on gut permeability and microbiota and their relationships with metabolic changes in mice. Obesity (Silver Spring) 2015, 23, 1429–1439. [Google Scholar] [CrossRef]
- Yamada, S.; Kamada, N.; Amiya, T.; Nakamoto, N.; Nakaoka, T.; Kimura, M.; Saito, Y.; Ejima, C.; Kanai, T.; Saito, H. Gut microbiota-mediated generation of saturated fatty acids elicits inflammation in the liver in murine high-fat diet-induced steatohepatitis. BMC Gastroenterol. 2017, 17, 136. [Google Scholar] [CrossRef] [PubMed]
- Prieto, I.; Hidalgo, M.; Segarra, A.B.; Martínez-Rodríguez, A.M.; Cobo, A.; Ramírez, M.; Abriouel, H.; Gálvez, A.; Martínez-Cañamero, M. Influence of a diet enriched with virgin olive oil or butter on mouse gut microbiota and its correlation to physiological and biochemical parameters related to metabolic syndrome. PLoS ONE 2018, 13, e0190368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, J.M.; Pan, C.; Cantor, R.M.; Tang, W.H.W.; Garcia-Garcia, J.C.; Kurtz, I.; Hazen, S.L.; Bergeron, N.; Krauss, R.M.; Lusis, A.J. Impact of individual traits, saturated fat, and protein source on the gut microbiome. mBio 2018, 9, e01604–e01618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenq, R.R.; Taur, Y.; Devlin, S.M.; Ponce, D.M.; Goldberg, J.D.; Ahr, K.F.; Littmann, E.R.; Ling, L.; Gobourne, A.C.; Miller, L.C.; et al. Intestinal Blautia is associated with reduction death from graft-versus-host disease. Biol. Blood Marrow Transplant. 2015, 21, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Chen, W.D.; Wang, Y.D. Gut microbiota: An integral moderator in health and disease. Front. Microbiol. 2018, 9, 151. [Google Scholar] [CrossRef]
- Garcia-Mantrana, I.; Selma-Royo, M.; Alcantara, C.; Collado, M.C. Shifts on gut microbiota associated to Mediterranean diet adherence and specific dietary intakes on general adult population. Front. Microbiol. 2018, 9, 890. [Google Scholar] [CrossRef]
- Li, Y.; Yang, H.; Xu, L.; Wang, Z.; Zhao, Y.; Chen, X. Effects of dietary fiber levels on cecal microbiota composition in geese. Asian-Australas J. Anim. Scie. 2018, 31, 1285–1290. [Google Scholar] [CrossRef]
- Jefferson, A.; Adolphus, K. The effects of intact cereal grain fibers, including wheat bran on the gut microbiota composition of healthy adults: A systematic review. Front. Nutr. 2019, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.C.; Yu, S.C.; Lo, Y.C.; Lin, I.H.; Tung, T.H.; Huang, S.Y. A high linoleic acid exacerbates metabolic responses and gut microbiota dysbiosis in obese rats with diabetes mellitus. Food Funct. 2019, 10, 786–798. [Google Scholar] [CrossRef]
- Wang, K.; Liao, M.; Zhou, N.; Bao, L.; Ma, K.; Zheng, Z.; Wang, Y.; Liu, C.; Wang, W.; Wang, J.; et al. Parabacteroides distasonis alleviates obesity and metabolic dysfunctions via production of succinate and secondary bile acids. Cell Reports 2019, 26, 222–235. [Google Scholar] [CrossRef] [Green Version]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, E.R.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Dng, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–485. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Backhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Koliada, A.; Syzenko, G.; Moseiko, V.; Budovska, L.; Puchkov, K.; Perederiy, V.P.; Gavalko, Y.; Dorofeyev, A.; Romanenko, M.; Tkach, S.; et al. Association between body mass index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population. BMC Microbiol. 2017, 17, 120. [Google Scholar] [CrossRef] [Green Version]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A common factor in human diseases. Biomed. Res. Int. 2017, 2017, 9351507. [Google Scholar] [CrossRef] [Green Version]
- De Souza, P.A.L.; Marcadenti, A.; Portal, V.L. Effects of olive oil phenolics compounds on Inflammation in the prevention and treatment of coronary artery disease. Nutrients 2017, 9, 1087. [Google Scholar] [CrossRef] [Green Version]
- Luque-Sierra, A.; Alvarez-Amor, L.; Kleemann, R.; Martín, F.; Varela, L.M. Extra-virgin olive oil with natural phenolic content exerts an anti-inflammatory effect in adipose tissue and attenuates the severity of atherosclerotic lesion in Ldlr–/–.Leiden mice. Mol. Nutr. Food Res. 2018, 62, 1800295. [Google Scholar] [CrossRef] [PubMed]
- Visioli, E.; Franco, M.; Toledo, E.; Luchsinger, J.; Willett, W.C.; Hu, F.B.; Martinez-Gonzalez, M.A. Olive oil and prevention of chronic diseases: Summary of an international conference. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Khoo, C.; Furtado, J.; Ikewaki, K.; Sacks, F.M. Dietary monounsaturated fat activates metabolic pathways for triglycerides-rich lipoproteins that involve apolipoproteins E and C-III. Am. J. Clin. Nur. 2008, 88, 272–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, A.; Perez-Martinez, P.; Delgado-Lista, J.; Marin, C.; Cortes, B.; Rodriguez-Cantalejo, F.; Gomez-Luna, M.J.; Cruz-Teno, C.; Perez-Jimenez, F.; Lopez-Miranda, J. Body mass interacts with fat quality to determine the postprandial lipoprotein response in healthy young adults. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 355–361. [Google Scholar] [CrossRef]
- Pu, S.; Khazanehei, H.; Jones, P.J.; Khafipour, E. Interactions between obesity status and dietary intake of monounsaturated and polyunsaturated oils on human gut microbiome profiles in the canola oil multicenter intervention trial (COMIT). Front. Microbiol. 2016, 7, 1612. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, M.; Prieto, I.; Abriouel, H.; Villarejo, A.B.; Ramírez-Sánchez, M.; Cobo, A.; Benomar, N.; Gálvez, A.; Martínez-Cañamero, M. Changes in gut microbiota linked to a reduction in systolic blood pressure in spontaneously hypertensive rats fed an extra virgin olive oil-enriched diet. Plant. Foods Hum. Nutr. 2018, 73, 1–6. [Google Scholar] [CrossRef]
- Bäckhed, F.; Manchester, J.K.; Semenkovich, C.F.; Gordon, J.I. Mechanisms underlying the resistance to diet-induced obesity in germ-freemice. Proc. Natl. Acad, Sci. USA. 2007, 16, 979–984. [Google Scholar] [CrossRef] [Green Version]
- De La Serre, C.B.; Ellis, C.L.; Lee, J.; Hartman, A.L.; Rtledge, J.C.; Raybould, H.E. Propensity to high-fat diet-induced obesity in rats is associated with changes in the gut microbiota and gut inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G440–G448. [Google Scholar] [CrossRef]
- Mujico, J.R.; Baccan, G.C.; Gheorghe, A.; Díaz, L.E.; Marcos, A. Changes in gut microbiota due to supplemented fatty acids in diet-induced obese mice. Br. J. Nutr. 2013, 110, 711–720. [Google Scholar] [CrossRef] [Green Version]
- Menni, C.; Zierer, J.; Pallister, T.; Jackson, M.A.; Long, T.; Mohney, R.P.; Steves, C.J.; Spector, T.D.; Valdes, A.M. Omega-3 fatty acids correlate with gut microbiome diversity and production of N-carbamylglutamate in middle aged and elderly women. Sci. Rep. 2017, 7, 11079. [Google Scholar] [CrossRef] [Green Version]
- Watson, H.; Mitra, S.; Croden, F.C.; Taylor, M.; Wood, H.M.; Perry, S.L.; Spencer, J.A.; Quirke, P.; Toogood, G.J.; Lawton, C.L.; et al. A randomised trial of the effect of omega-3 polyunsaturated fatty acid supplements on the human intestinal microbiota. Gut 2017, 67, 1974–1983. [Google Scholar] [CrossRef]
- Andrade-Oliveira, V.; Amano, M.Y.; Correa-Costa, M.; Castoldi, A.; Felizardo, R.J.F.; de Almeida, D.C.; Bassi, E.J.; Moraes-Vieira, P.M. Gut bacteria products prevent AKI induced by ischemia-reperfusion. J. Am. Soc. Nephrol. 2015, 26, 1877–1888. [Google Scholar] [CrossRef] [PubMed]
- Constantini, L.; Molinari, R.; Farinon, B.; Merendino, N. Impact of omega-3 fatty acids on the gut microbiota. Int. J. Mol. Sci. 2017, 18, 2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Rizzolo, D.A.; Kostov, B.; López-Siles, M.; Serra, A.; Colungo, C.; González-de-Paz, L.; Martinez-Medina, M.; Sisó-Almirall, A.; Gomis, R. Healthy dietary pattern and their corresponding gut microbiota profile are linked to a lower risk of type 2 diabetes, independent of the presence of obesity. Clin. Nutr. 2020, 39, 524–532. [Google Scholar] [CrossRef] [Green Version]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance–A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef] [PubMed]
- USDA Nutrient Database. Available online: http://ndb.nal.usda.gov/ndb/search (accessed on 22 February 2020).
- Bazinet, R.P.; Layé, S. Polyunsaturated fatty acids and their metabolites in brain function and disease. Nat. Rev. Neurosci. 2014, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- van Elst, K.; Bruining, H.; Birtoli, B.; Terreaux, C.; Buitelaar, J.K.; Kas, M.J. Food for thought: Dietary changes in essential fatty acid ratios and the increase in autism spectrum disorders. Neurosci. Biobehav. Rev. 2014, 45, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.W.; Chien, Y.S.; Chen, Y.J.; Ajuwon, K.M.; Mersmann, H.M.; Ding, S.T. Role of n-3 polyunsaturated fatty acids in ameliorating the obesity-induced metabolic syndrome in animal models and humans. Int. J. Mol. Sci. 2016, 17, 1689. [Google Scholar] [CrossRef] [PubMed]
- Lalia, A.Z.; Lanza, I.R. Insulin-sensitizing effects of omega-3 fatty acids: Lost in translation? Nutrients 2016, 8, 329. [Google Scholar] [CrossRef]
- Lepretti, M.; Martucciello, S.; Aceves, M.A.B.; Putti, R.; Lioneti, L. Omega-3 fatty acids and insulin resistance: Focus on the regulation of mitochondria and endoplasmic reticulum stress. Nutrients 2018, 10, 350. [Google Scholar] [CrossRef] [Green Version]
- Sokova-Wysoczanska, E.; Wysoczanski, T.; Wagner, J.; Czyz, K.; Bodkowski, R.; Lochynski, S.; Patkowska-Sokola, B. Polyunsaturated fatty acids and their potential therapeutic role in cardiovascular system disorders–A review. Nutrients 2018, 10, 1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, E.; Wall, R.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Health implications of high dietary omega-6 polyunsaturated fatty acids. J. Nutr. Metab. 2012, 2012, 539426. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Molcant, E.; DeCoffe, D.; Dai, C.; Gibson, D.L. Diets rich in n-6 PUFA induce intestinal microbial dysbiosis in aged mice. Br. J. Nutr. 2013, 110, 515–523. [Google Scholar] [CrossRef] [Green Version]
- Kris-Etherton, P.M.; Taylor, D.S.; Yu-Poth, S.; Huth, P.; Moriarty, K.; Fishell, V.; Hargrove, R.L.; Zhao, G.; Etherton, T.D. Polyunsaturated fatty acids in the food chain in the United States. Am. J. Clin. Nutr. 2000, 71, 179S–188S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druart, C.; Bindels, L.B.; Schmaltz, R.; Neyrinck, A.M.; Cani, P.D.; Walter, J.; Ramer-Tait, A.E.; Delzenne, N.M. Ability of the gut microbiota to produce PUFA-derived bacterial metabolites: Proof of concept in germ-free versus conventionalized mice. Mol. Nutr. Food Res. 2015, 59, 1603–1613. [Google Scholar] [CrossRef]
- Noriega, B.S.; Snchaz-Gonzalez, M.A.; Salyakina, D.; Coffman, J. Understanding the impact of omega-3 rich diet on the gut microbiota. Case Rep. Med. 2016, 2016, 3089303. [Google Scholar] [CrossRef] [Green Version]
- Hildebrandt, M.A.; Hoffmann, C.; Sherrill-Max, S.A.; Keilbaugh, S.A.; Hamady, M.; Chen, Y.Y.; Knight, R.; Ahima, R.S.; Bushman, F.; Wu, G.D. High-fat diet determines the composition of the murine gut microbiome independently of obesity. Gastroenterology 2009, 137, 1716–1724. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.N.; Zhu, J.; Pan, W.S.; Shen, S.R.; Shan, W.G.; Das, U.N. Effects of fish with a high content of n-3 polyunsaturated fatty acids on mouse gut microbiota. Arch. Med. Res. 2014, 45, 195–202. [Google Scholar] [CrossRef]
- Myles, I.A.; Pincus, N.B.; Fontecilla, N.M.; Datta, S.K. Effects of parental omega-3 fatty acid intake on offspring microbiome and immunity. PLoS ONE 2014, 9, e87181. [Google Scholar] [CrossRef]
- Pusceddu, M.M.; Aidy, S.E.; Crispie, F.; O’Sullivan, O.; Cotter, P.; Stanton, C.; Kelly, P.; Cryan, J.F.; Dinan, T.G. N-3 polyunsaturated fatty acids (PUFAs) reverse the impact of early-life stress on the gut microbiota. PLoS ONE 2015, 10, e0139721. [Google Scholar] [CrossRef]
- Robertson, R.C.; Oriach, C.S.; Murphy, K.; Moloney, G.M.; Cryan, J.F.; Dinan, T.G.; Ross, R.P.; Stanton, C. Omega-3 polyunsaturated fatty acids critically regulate behavior and gut microbiota development in adolescent and adulthood. Brain Behav. Immun. 2017, 59, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Robertson, R.C.; Kaliannan, K.; Strain, C.R.; Ross, R.P.; Stanton, C. Maternal omega-3 fatty acids regulate offspring obesity through persistent modulation of gut microbiota. Microbiome 2018, 6, 95. [Google Scholar] [CrossRef] [PubMed]
- Bravo, D.; Hoare, A.; Soto, C.; Valenzuela, M.A.; Quest, A.F.G. Helicobacter pylori in human health and disease: Mechanisms for local gastric and systemic effects. World, J. Gastroenterol. 2018, 24, 3071–3089. [Google Scholar] [CrossRef]
- Allin, K.H.; Tremaroli, V.; Caesar, R.; Jensen, B.A.H.; Damgaard, M.T.F.; Bahl, M.I.; Licht, T.R.; Hansen, T.H.; Nielsen, T.; Dantoft, T.M.; et al. Aberrant intestinal microbiota in individuals with prediabetes. Diabetologia 2018, 61, 810–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, R.X.; Goh, W.R.; Wu, R.N.; Yue, X.Q.; Luo, X.; Khine, W.W.T.; Wu, J.R.; Lee, Y.K. Revisit gut microbiota and its impact on health and disease. J. Food Drug Anal. 2019, 27, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Sandri, M.; Monego, S.D.; Conte, G.; Sgorlon, S.; Stefanon, B. Raw meat based diet influences faecal microbiome and end products of fermentation in healthy dogs. BMC Vet. Res. 2017, 13, 65. [Google Scholar] [CrossRef]
- Monda, V.; Villano, I.; Messina, A.; Valenzano, A.; Esposito, T.; Moscatelli, F.; Viggiano, A.; Cibelli, G.; Chieffi, S.; Monda, M.; et al. Exercise modifies the gut microbiota with positive health effects. Oxid. Med. Cell Longev. 2017, 2017, 3831972. [Google Scholar] [CrossRef]
- Hermann, E.; Young, W.; Rosendale, D.; Reichert-Grimm, V.; Riedel, C.U.; Conrad, R.; Egert, M. RNA-Based stable isotope probing suggests Allobaculum spp. as particularly active glucose assimilators in a complex murine microbiota cultured in vitro. BioMed Res. Int. 2017, 2017, 1829685. [Google Scholar] [CrossRef] [Green Version]
- Raza, G.S.; Putaala, H.; Hibberd, A.A.; Alhoniemi, E.; Tiihonen, K.; Mäkelä, K.A.; Herzig, K.H. Polydextrose changes the gut microbiome and attenuates fasting triglyceride and cholesterol levels in western diet fed mice. Sci. Rep. 2017, 7, 5294. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Zeng, M.Y.; Núñez, G. The interplay between host immune cells and gut microbiota in chronic inflammatory diseases. Exp. Mol. Med. 2017, 49, e339. [Google Scholar] [CrossRef] [Green Version]
- De La Cuesta-Zuluaga, J.; Corrales-Agudelo, V.; Velásques-Mejía, E.P.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Gut microbiota is associated with obesity and cardiometabolic disease in a population in the midst of westernization. Sci. Rep. 2018, 8, 11356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méndez-Salazar, E.O.; Ortiz-López, M.G.; Granados-Silvestre, M.A.; Palacios-González, B.; Menjivar, M. Altered gut microbiota and compositional changes in Firmicutes and Proteobacteria in Mexican undernourished and obese children. Front. Microbiol. 2018, 9, 2494. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]


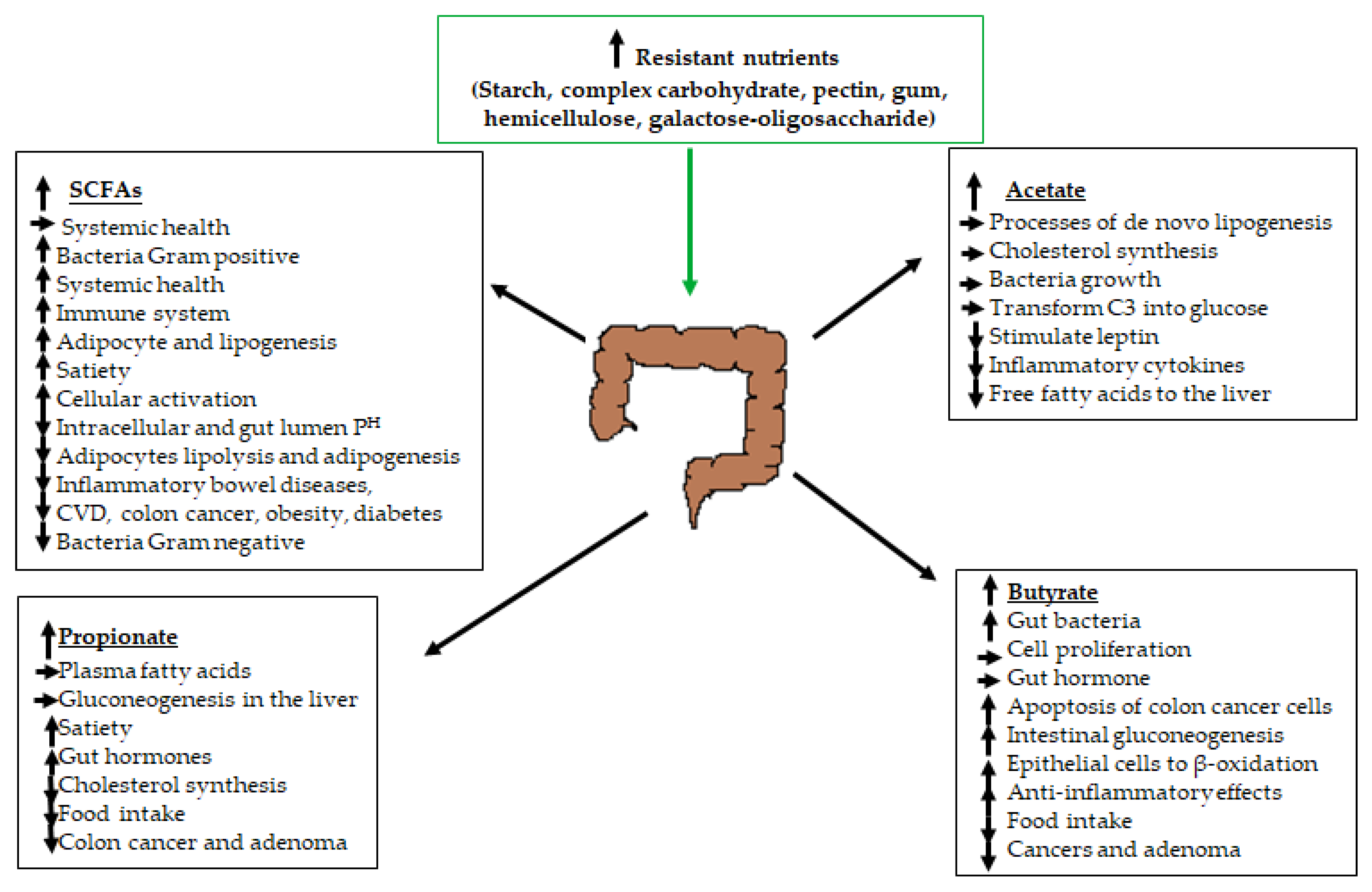{kind=link}
| Host | Diets | Main Outcomes | |
|---|---|---|---|
| Gut Microbiota | Related Diseases | ||
| Mice C57BL/6J (7 weeks old): healthy male [46] | HFD containing 20% (w/w) rapeseed oil with MCFAs (30%) for 6 weeks | Bacteroidetes↑ Allobaculum and Lachnospiraceae (Firmicutes)↓ Helicobacter spp. (Proteobacteria)↓ | IBD↓ Obesity↓ |
| Wistar rats (10 weeks old): male with induced diabetic [47] | Virgin coconut oil (caprylic, 6.57%; capric, 5.78%; and lauric, 48.51%) for 16 weeks | Bifidobacterium (Actinobacteria)↑ Allobaculum and Lactobacullum (Firmicutes)↑ | T2DM↔ |
| Wistar rats: female [48] | HFD (50 or 95%) of Virgin coconut oil (caprylic, 5.22%; capric, 5.41%; and lauric, 51.64%) for 10 weeks | Bacteroides and Prevotella (Bacteroidetes)↑ Bifidobacterium (Actinobacteria)↑ Lactobacullum and Enterocuccus (Firmicutes)↑ Clostridium histolyticum (Firmicutes)↓ | IBD↑ adipose tissue↑ NASH↑ |
| Mice C57BL/6N (3 weeks old): healthy female [49] | HFD containing coconut oil 25% and soy oil 0.25% for 8 weeks | Allobaculum, Staphylococcus, Clostridium, F16, YS2, Lactobacillus (Firmicutes)↑ Deltaproteobacteria (Proteobacteria)↑ Bacteroidetes↓ | Obesity↑ adipose tissue↑ plasma cholesterol↑. |
| Host | Diets | Main Outcomes | |
|---|---|---|---|
| Gut Microbiota | Metabolic | ||
| Mice C57BL/6J (3 weeks old): healthy male [66] | LFD palm oil (rich in palmitic acid) (10% kcal) for 3 weeks | Bacteroidetes↑ Bacilli and Clostridium cluster XI, XVII, and XVIII (Firmicutes)↓. | Adipose tissue↓ weight gain↓ NASH↓ insulin resistance↓ |
| HFD palm oil (rich in palmitic acid) (45% kcal) for 3 weeks | Bacilli and Clostridium clusters XI, XVII, and XVIII (Firmicutes)↑ Bacteroidetes↓. | Adipose tissue↑ weight gain↑ NASH↑ | |
| Mice (C57BL/6J) (8 weeks old): healthy male [67] | HFD (60 kcal % fat diet (HFD, D12492) for 8 weeks | Firmicutes↑ Enterobacteriaceae (Proteobacteria)↑ Rikenellaceae, Bacteroidaceae, and Provotellaceae (Bacteriodetes)↓ Ruminococcaceae and Clostridiales (Firmicutes)↓ Proteobacteria and Bifidobacterium (Actinobacteria)↓. | IBD↑ weight gain↑ |
| Mice C57BL/6J (7–10 weeks old): healthy male [68] | Milk fat (rich in palmitic, stearic, myristic, and oleic acids) for 4 weeks | Firmicutes↑ Proteobacteria↑ Actinobacteria↑ Bacteroidetes↓. | Adipose tissue↑ IBD↑ weight gain↑ |
| Mice RELMβ KO (13 weeks old): healthy female [68] | Safflower oil (rich in palmitic acid) for 4 weeks | Firmicutes↑ Tenericutes↑ Actinobacteria↑ Bacteroidetes↓ | Adipose tissue↑ IBD↑ weight gain↑ |
| Mice C57BL/6J (3 weeks old): healthy male [69] | HFD palm oil (rich in palmitic acid) with 45% energy for 16 weeks | Coprococcus, Erysipelotrichaceae, and Lachnospiraceae (Firmicutes)↑ Bacteroides, Bacteroidaceae (Bacteroidetes)↓ Deferribacteres↓ Actinobacteria↓ Proteobacteria↓. | Adipose tissue↑ weight gain↑ insulin resistance↑ |
| Mice (C57BL/6J) (3 weeks old): healthy male [70] | HFD (60% of energy from fat; 95% from lard; and 5% from soybean oil) for 6 weeks | Desulfovibrio and Bilophila wadsworthia (Proteobacteria)↑ Bifidobacterium spp. (Actinobacteria)↓. | Adipose tissue↑ IBD↑ |
| Mice C57BL/6J (6 weeks old): healthy female [71] | HFD saturated fatty acid with 34% energy for 8 weeks | Lactobacillus, erysipelotrichaceae, Lachnospiraceae, and Pseudoflavonifractor (Firmicutes)↑ Bilophila (Proteobacteria)↑ Allobaculum (Firmicutes)↓ Bamesiella (Bacteroidetes)↓ Mucispirillum (Deferribacteres)↓ Bacteroides (Bacteroidetes)↓ Bifidobacterium (Actinobacteria)↓. | Weight gain↑ adipose tissue↑ insulin resistance↑ IBD↑ gut permeability↑ |
| Mice SPF C57BL/6J (8 weeks old): healthy [72] | HFD with 72% fat/kcal for 9 weeks. | Clostridium (Firmicutes)↑ Bifidobacterium (Actinobacteria)↓ Enterococcus (Firmicutes)↓ Bacteroides (Bacteroidetes)↓. | NASH↑ |
| ICR Swiss mice (6 weeks old): healthy male [73] | Butter diet with 38% energy for 12 weeks | Alistipes indistinctus (Bacteroidetes)↑ Marvinbryantia, Lactobacillus spp. and Lactococcus (Firmicutes)↑ Anaerostipes butyaticus, Desulfovibrio desulfuricans and Escherichia fergunsoni (Proteobacteria)↑ Bacteroidetes↓ | Weight gain↑ hypertension↑ insulin resistance↑ total cholesterol↑ |
| Mice C57BL/6N (3 weeks old): healthy female [49] | HFD containing coconut oil 25% and soy oil 0.25% for 2–8 weeks. | Anaerotruncus, Syntrophomonas, Lutispora and Lactobacillus (Firmicutes)↑ Parabacteroidetes (Bacteroidestes)↑ Akkermansia (Verruncomicrobia)↑ Proteobacteria↑ Anaerostipes and Peptostreptococcaceae (Firmicutes)↓ Agrobacterium (Proteobacteria)↓ | Obesity↑ adipose tissue↑ plasma cholesterol↑ |
| Host | Diets | Main Outcomes | |
|---|---|---|---|
| Gut Microbiota | Metabolic | ||
| Mice C57BL/6J (germ free wild-type): healthy male [98] | Western diet with 41% energy from fat for 8 weeks | Bacteroidetes↑ Firmicutes↓ | Adipose tissue↓ obesity↓ |
| Rats: Sprague–Dawley healthy male [99] | LFD 10% (SFA 25.1%, MUFA 34.7%, and PUFA 40.2%) for 8 eight weeks | Bacteroidales (Bacteroidetes)↑ Clostridiales (Firmicutes)↓ Enterobacteriales (Proteobacteria)↓ | IBD↓ obesity↓ |
| Mice C57BL/6J (3 weeks old): healthy male [66] | HFD olive oil rich in oleic acid (45% kcal) for three weeks | Bacteroidetes↑ Bacilli and Clostridium cluster XI, XVII, and XVIII (Firmicutes)↓ | Adipose tissue↓ weight gain↓ NASH↓ insulin resistance↓ |
| ICR Swiss mice: 8-week-old healthy female [100] | HFD supplementation with an oleic acid (16% per day) for 19 weeks | Bacteroidetes↑ Bifidobacterium spp. (Actinobacteria)↑ Lactobacillus spp. (Firmicutes)↓ Clostridial cluster XIVa (Firmicutes)↓ Enterobacteriales (Proteobacteria)↓ | Obesity↓ IBD↓ |
| Mice C57BL/6J (3 weeks old): healthy male [69] | HFD olive oil (oleic acid) with 45% energy for 16 weeks | Allobculum, Erysipelotrichaceae (Firmicutes)↑ Bacteroides, Bacteroidaceae (Bacteroidetes)↓ Deferribacteres↓ Proteobacteria↓ Actinobacteria↓ | Weight gain↓ NASH↓ |
| Rats (4–5 weeks old): spontaneously hypertensive male [97] | EVOO diet: 20% of EVOO (oleic acid) with 75.5% energy for 12 weeks | Lachnospiraceae, Ruminococcaceae (Clostridia XIVa) and Lactobacillus (Firmicutes)↑ Bacteroidetes↓ Actinonobacteria↓ | Hypertension↓ |
| ICR Swiss mice (6 weeks old): healthy male [73] | EVOO with 38% energy for 12 weeks | Prevotellaceae, Marinillabiliaceae, Mucilaginibacter dageonensis, Bacteroides fragilis and Alistipes indictintus (Bacteroidetes)↑ Sutterellaceae and Marispirillum (Proteobacteria)↑ Christenellaceae, Erysipelotrichaceae and Clostridim cocleatum (Firmicutes)↑ Desulfovibrio (Firmicutes)↓ | Hypertension↓ weight gain↓ |
| Host | Diets | Main Outcomes | |
|---|---|---|---|
| Gut Microbiota | Metabolic | ||
| Men and women volunteers with risk of metabolic syndrome [96] | MUFA-rich oil (canola, 36%; canola/DHA, 39%; and canola oleic, 44% energy) for 4 weeks | Coprobacillus, Faecalibacterium Lactobacillus, Robinsoniella and Tepidimicrobium Fusibacter, Turicibacter (Firmicutes)↑ Flexithrix, Parabacteroides, and Prevotella (Bacteroidetes)↑ Enterobacteriaceaes (Proteobacteria)↑ Isobaculum (Firmicutes)↓ | BMI↓ |
| Men and women obese volunteers with prediabetes risk (≥65 years old) [105] | Lipids 40% (MUFA 19%) for 3 days | Prevotella (Bacteroidetes)↓ Faecalibacterium prausnitzzi, Lactic acid bacteria (Firmicutes)↑ Escherichia coli (Proteobacteria)↑ Firmicutes/Bacteroidetes ratio↑ | T2DM↓ |
| Men and women nonobese volunteers with prediabetes risk (≥65 years old) [105] | Lipids 41% (MUFA 19%) for 3 days | Firmicutes/Bacteroidetes↓ Prevotella (Bacteroidetes)↑ Faecalibacterium prausnitzzi, Lactic acid bacteria (Firmicutes)↑ Escherichia coli (Proteobacteria)↓ | T2DM↓ |
| Host | Diets | Main outcomes | |
|---|---|---|---|
| Gut microbiota | Metabolic | ||
| Mice wild-type (13 weeks old): healthy female [119] | Safflower oil (rich in linoleic acid) for 21 weeks | Clostridiaceae (Firmicutes)↑ Desulfovibrionaceae (Proteobacteria)↑ Bacteriodaceae, Prevotellaceae and Rickenellaceae (Bacteroidetes)↓ | Obesity↑ |
| Mice RELMβ KO (13 weeks old): healthy female [119] | Safflower oil (rich in linoleic acid) for 21 weeks | Clostridiaceae (Firmicutes)↑ Desulfovibrionaceae (Proteobacteria)↑ Bacteriodaceae, Prevotellaceae and Rickenellaceae (Bacteroidetes)↓ | Obesity↓ |
| Rats: Sprague–Dawley male [99] | LFD 10% (SFA 25%, MUFA 35%, and PUFA 40%) at eight weeks | Bacteroidales (Bacteroidetes)↑ Clostridiales (Firmicutes)↓ Enterobacteriales (Proteobacteria)↓ | IBD↓ obesity↓ |
| Mice C57BL/6J (3 weeks old): healthy male [66] | Safflower oil rich in linoleic acid (45% energy) for 8 weeks | Bacteroidetes↑ Clostridium cluster XI, XVII, and XVIII (Firmicutes)↑ Bacilli (Firmicutes)↓ | Adipose tissue↓ obesity↓ NASH↓ insulin resistance↓ |
| Mice C57Bl/6 (7–10 weeks old): healthy male [68] | HFD safflower oil (rich in linoleic acid) for 4 weeks | Firmicutes↑ Tenericutes↑ Actinobacteria↑ Deferibacteria↑ Proteobacteria↑ Bacteroidetes↓ | IBD↑ weight gain↑ |
| ICR Swiss mice: 8-week-old healthy female [100] | HFD supplementation with n–3 PUFAs (EPA + DHA) for 19 weeks | Bifidobacterium spp. (Actinobacteria)↑ Bacteroidetes↑. Lactobacillus spp. (Firmicutes)↑ Enterobacteriales (Proteobacteria)↑ Clostridial cluster XIVa (Firmicutes)↓ | IBD↓ Obesity↓ |
| Mice C57BL/6J (24 months old): healthy female [115] | 1. HFD of maize oil + rapeseed oil (rich in n–6 PUFAs) with 40% energy for 7 weeks | Firmicutes↑ Bacteroidetes↓ | Weight gain↑ IBD↑ |
| 2. LFD of maize oil plus fish oil supplemented (rich in n–3 PUFA (EPA + DHA) with 34% energy for 7 weeks | Bacteroidetes↑ Firmicutes↓ Proteobacteria↓ | Weight gain↓ IBD↓ | |
| Mice C57BL/6J (3 weeks old): healthy male [69] | 1. HFD safflower oil (linoleic acid n–6 PUFA) with 45% energy for 16 weeks | Allobaculum, Oscillibacter and Ruminococcaceae (Firmicutes)↑ Bacteroides and Parabacteroides (Bacteroidetes)↑ Bifidobacterium (Actinobacteria)↑ | Weight gain↑ Insulin resistance↑ |
| 2. HFD flaxseed/fish oil (α-linolenic acid n–3 PUFA) with 45% energy for 16 weeks | Allobaculum, Erysipalotrichaceae and Lachnospiraceae (Firmicutes)↑ Deferribacteres↑ Bifidobacteriaceae (Actinobacteria)↑ Bacteroides, Bacteroidaceae (Bacteroidetes)↓ Proteobacteria↓ | Weight gain↓ Insulin resistance↓ NASH↓ | |
| ICR mice (4 weeks old): healthy male and female (17–21 g) [120] | HFD fish oil (40% EPA and 27% DHA) n–3 PUFA for 2 weeks | Helicobacter, Pseudomonas sp., and Sphingomonadales (Proteobacteria)↓ Clostridiales (Firmicutes)↓. | Weight gain↔ |
| Mice BALB/c (3 weeks old): male and female pups from n–3 breeders [121] | HFD n–6/n–3 PUFAs (1/2) with 40% energy for 2 weeks | Blautia, Oscillibater, Clostridales, Robinsoniella, Lactococcus, and Eubacterium (Firmicutes)↑ Porphyromonadaceae (Bacteroidetes)↓ Lachnospiraceae and Rosebeuria, Euterococcus (Firmicutes)↓ | IBD↓ |
| Mice C57BL/6J (6 weeks old): healthy female [71] | HFD n–3 PUFA with 37% energy for 8 weeks | Lactobacillus, Allobaculum, Clostridium, and Turicibacter (Firmicutes)↑ Bifidobacterium (Actinobacteria)↑ Bamesella (Bacteroidetes)↓ Bilophila (Proteobacteria)↓ Akkemansia (Verrucomicrobia)↓ | Weight gain↓ adipose tissue↓ insulin resistance↓ |
| HFD n–6 PUFA with 31% energy for 8 weeks | Allobaculum, Erysipelotrichaceae, Lachnospiraceae and Oscillibacter (Firmicutes)↑ Mucispitillum (Deferribacteres)↑ Bilophila (Proteobacteria)↑ Lactobacillus and Acetivibrio (Firmicutes)↓ Bamesiella (Bacteroidetes)↓ Bifidobacterium (Actinobacteria)↓ | Weight gain↑ adipose tissue↑ insulin resistance↑ IBD↑ | |
| Rats (5 weeks old): early life stressed (weaned) female pups (250–300 g) with reduced Bacteroidetes/ Firmicutes ratio and inflamed gut [122] | HFD of n–3 PUFA (1 g EPA 80% + DHA 20%) for 17 weeks | Butyrivibrio, Jeotgalicoccus, and Peptococcus (Firmicutes)↑ Caldicoprobacter (Terrabacteria)↑ Bifidobacteria and Aerococcus (Actinobacteria)↑ Undibacterium (Proteobacteria)↓ | IBD↓ |
| Mice C57BL/6J (4–5 weeks old) and adulthood (11–13 weeks old): male offspring subsequently weaned onto the same diets as their mothers and stressed. Stressed adulthood [123] | HFD of n–3 PUFA-supplemented diet (1 g EPA + DHA/100 g diet) for 8 weeks | Bacteroidetes↑ Verrucomicrobia and bifidobacterium (Actionobacteria)↑ Firmicutes↓ Tenercutes and enterobacteria (Proteobacteria)↓ | IBD↓ |
| Mice C57BL/6 WT (4 weeks old): transgenic male and female lactated by mother lactated or foster mother [124] | Maternal n–3 PUFA for 4 weeks plus HFD 60% energy (SFA, 32%; MUFA, 36%; PUFA, 32%; n–6 PUFA, 30%; and n–3 PUFA, 2.1%) for six weeks | Helicobacter (Proteobacteria)↑ Bacteroides (Bacteroidetes)↑ Epsilonproteobacteria (Proteobacteria)↑ Lachnospiraceae and Ruminococcaceae (Firmicutes)↑ Akkermansia (Verrucomicrobia)↑ | Obesity↓ IBD↓ |
| Rats with diabetes mellitus (7 weeks old): male and female with type 2 diabetes mellitus [80] | 1. LFD n–6/n–3 (3/1) for 6 weeks 2. HFD with n–6/n–3 (9/1) for 6 weeks | Proteobacteria↑ Allobaculum (Firmicutes)↑ Actinobacteria↓ Firmicutes/Bacteroidetes↓ | Weight gain↓ IBD↓ insulin resistance↓ T2DM↓ |
| Host | Diets | Main Outcomes | |
|---|---|---|---|
| Gut Microbiota | Metabolic | ||
| Men and women (young): 98 healthy volunteers [135] | HFD n–3 PUFA | Bacteroidetes↑ Actinobacteria↑ Firmicutes↓ Proteobacteria↓ | Weight gain↓ |
| Men (45 years old): healthy and physically active [118] | Fish protein diet with vegetables that included over 600 mg of HFD n–3 PUFA for 2 weeks | Blautia, Coprococcus, Ruminococcus, Subdoligranulum, Eubacteria, Anaerosfipes, and Pseudobutyrivibrio (Firmicutes)↑ Roseburia and Faecalibacterium prausnitzii (Firmicites)↓ Akkermansia spp. (Verrucomicrobia)↓ Bacteroidetes↓ Actinobacteria↓ | IBD↓ T2DM↓ obesity↓ insulin resistance↓ |
| Men and women: volunteers with risk of metabolic syndrome [96] | HFD n–6 PUFA blended corn/safflower oil (25/75) with 42% energy and blended flax/safflower oil (6/4) with 42% energy for 4 weeks | Isobaculum (Firmicutes)↑ Parabacteroides and Prevotella, Bacteroidetes↓ Enterobacteriaceae↓ Turicibacter (Firmicutes)↓ | BMI↓ |
| Women twins (middle and elderly aged): 876 healthy [101] | HFD in n–6/n–3 PUFA (11/1) for 7 days | Lachnospiraceae (Firmicutes)↑ | BMI↓ obesity↓ |
| Men and women (≥50 years old): healthy [102] | Capsules and drink of n–3 PUFA (EPA + DHA) for 8 weeks | Bifidobacterium (Actinobacteria)↑ Oscillospira, Roseburia and Lachnospira (Firmicutes)↑ Coprococcus and Faecalibacterium (Firmicutes)↓ | BMI↓ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machate, D.J.; Figueiredo, P.S.; Marcelino, G.; Guimarães, R.d.C.A.; Hiane, P.A.; Bogo, D.; Pinheiro, V.A.Z.; Oliveira, L.C.S.d.; Pott, A. Fatty Acid Diets: Regulation of Gut Microbiota Composition and Obesity and Its Related Metabolic Dysbiosis. Int. J. Mol. Sci. 2020, 21, 4093. https://doi.org/10.3390/ijms21114093
Machate DJ, Figueiredo PS, Marcelino G, Guimarães RdCA, Hiane PA, Bogo D, Pinheiro VAZ, Oliveira LCSd, Pott A. Fatty Acid Diets: Regulation of Gut Microbiota Composition and Obesity and Its Related Metabolic Dysbiosis. International Journal of Molecular Sciences. 2020; 21(11):4093. https://doi.org/10.3390/ijms21114093
Chicago/Turabian StyleMachate, David Johane, Priscila Silva Figueiredo, Gabriela Marcelino, Rita de Cássia Avellaneda Guimarães, Priscila Aiko Hiane, Danielle Bogo, Verônica Assalin Zorgetto Pinheiro, Lincoln Carlos Silva de Oliveira, and Arnildo Pott. 2020. "Fatty Acid Diets: Regulation of Gut Microbiota Composition and Obesity and Its Related Metabolic Dysbiosis" International Journal of Molecular Sciences 21, no. 11: 4093. https://doi.org/10.3390/ijms21114093
APA StyleMachate, D. J., Figueiredo, P. S., Marcelino, G., Guimarães, R. d. C. A., Hiane, P. A., Bogo, D., Pinheiro, V. A. Z., Oliveira, L. C. S. d., & Pott, A. (2020). Fatty Acid Diets: Regulation of Gut Microbiota Composition and Obesity and Its Related Metabolic Dysbiosis. International Journal of Molecular Sciences, 21(11), 4093. https://doi.org/10.3390/ijms21114093