N-Acetylcysteine Reduced Ischemia and Reperfusion Damage Associated with Steatohepatitis in Mice

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
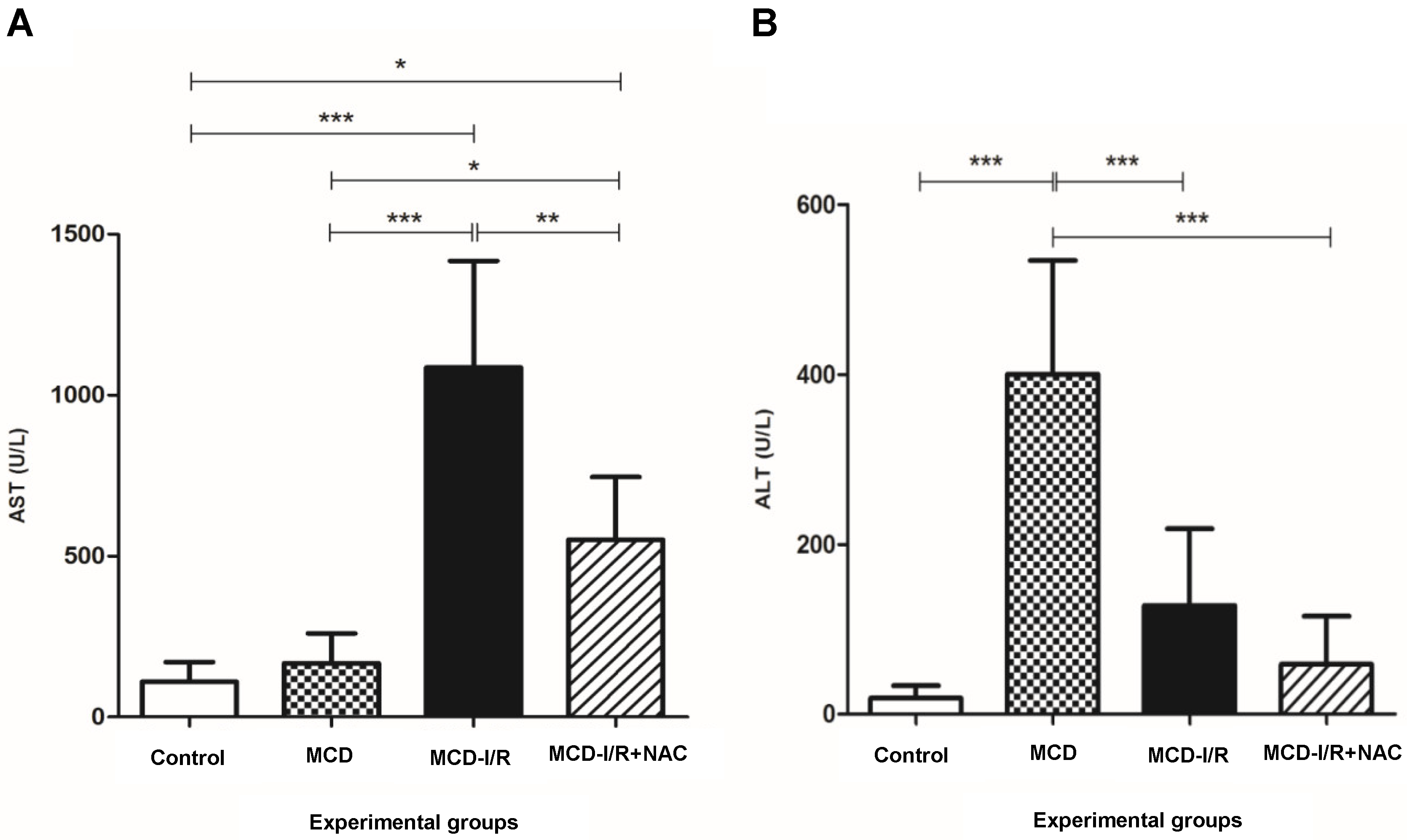
2.1. Hepatic Enzymes
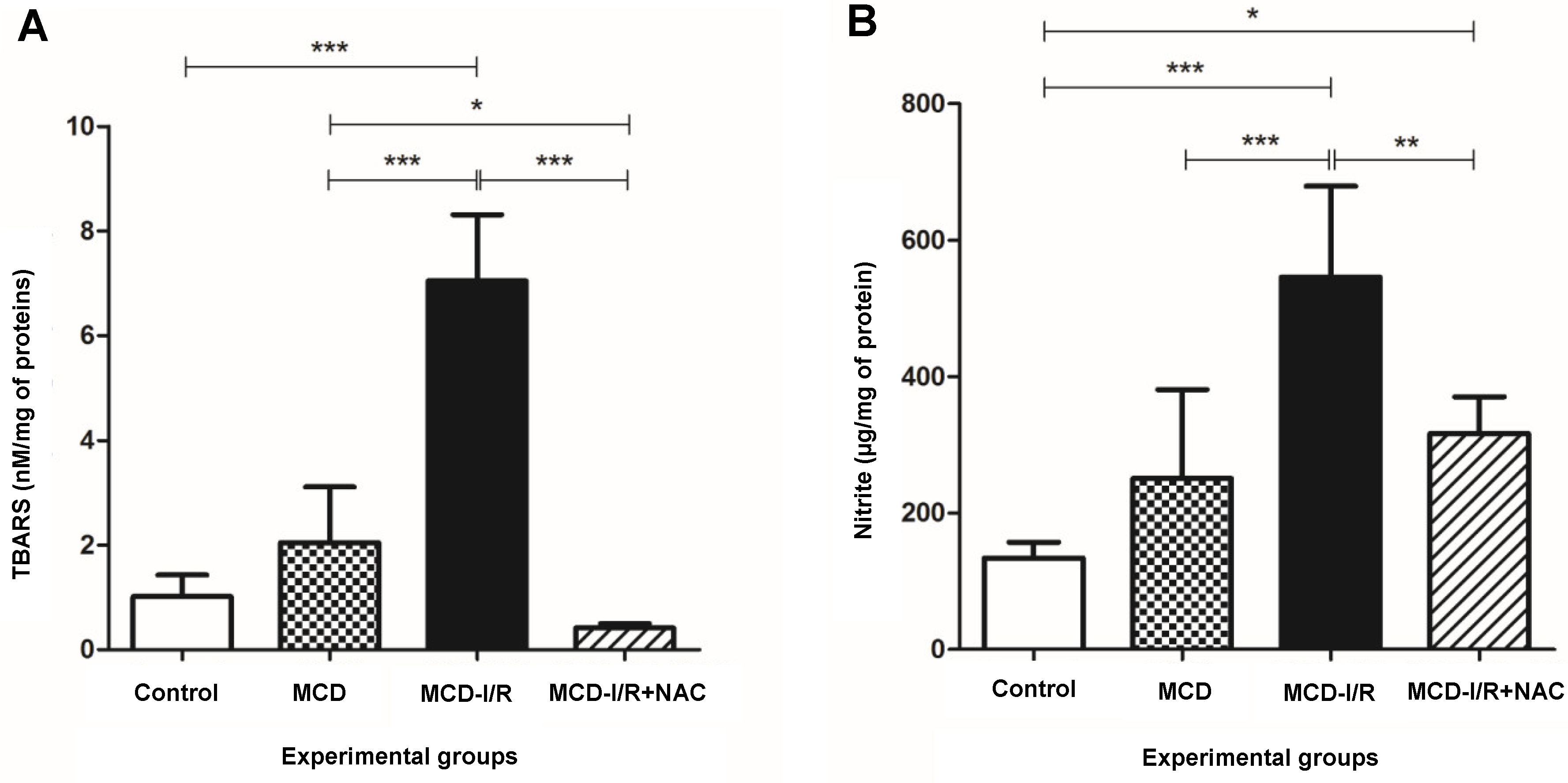
2.2. Oxidative Stress
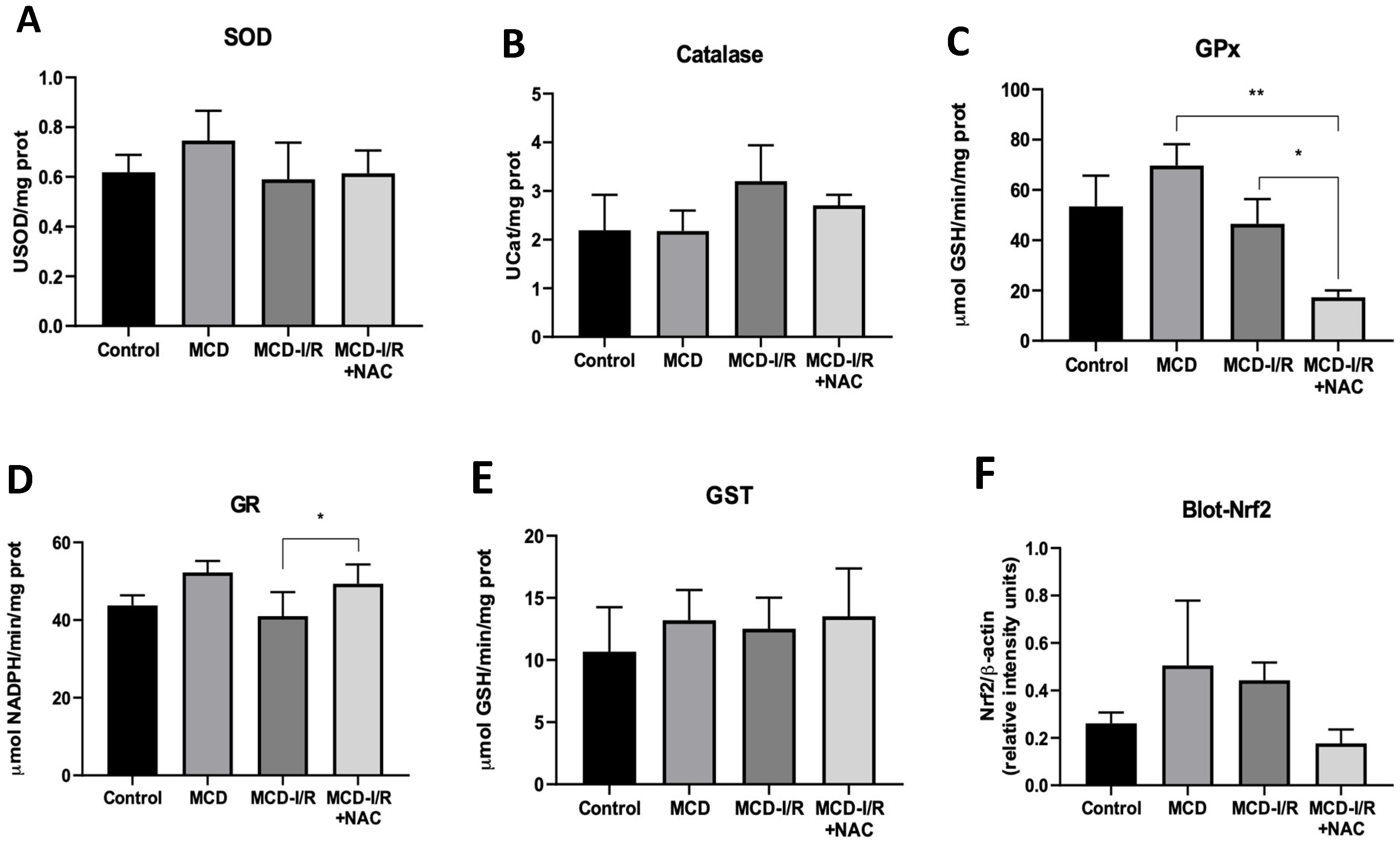
2.3. Antioxidant Enzyme Activities and Transcription Factor Nrf2
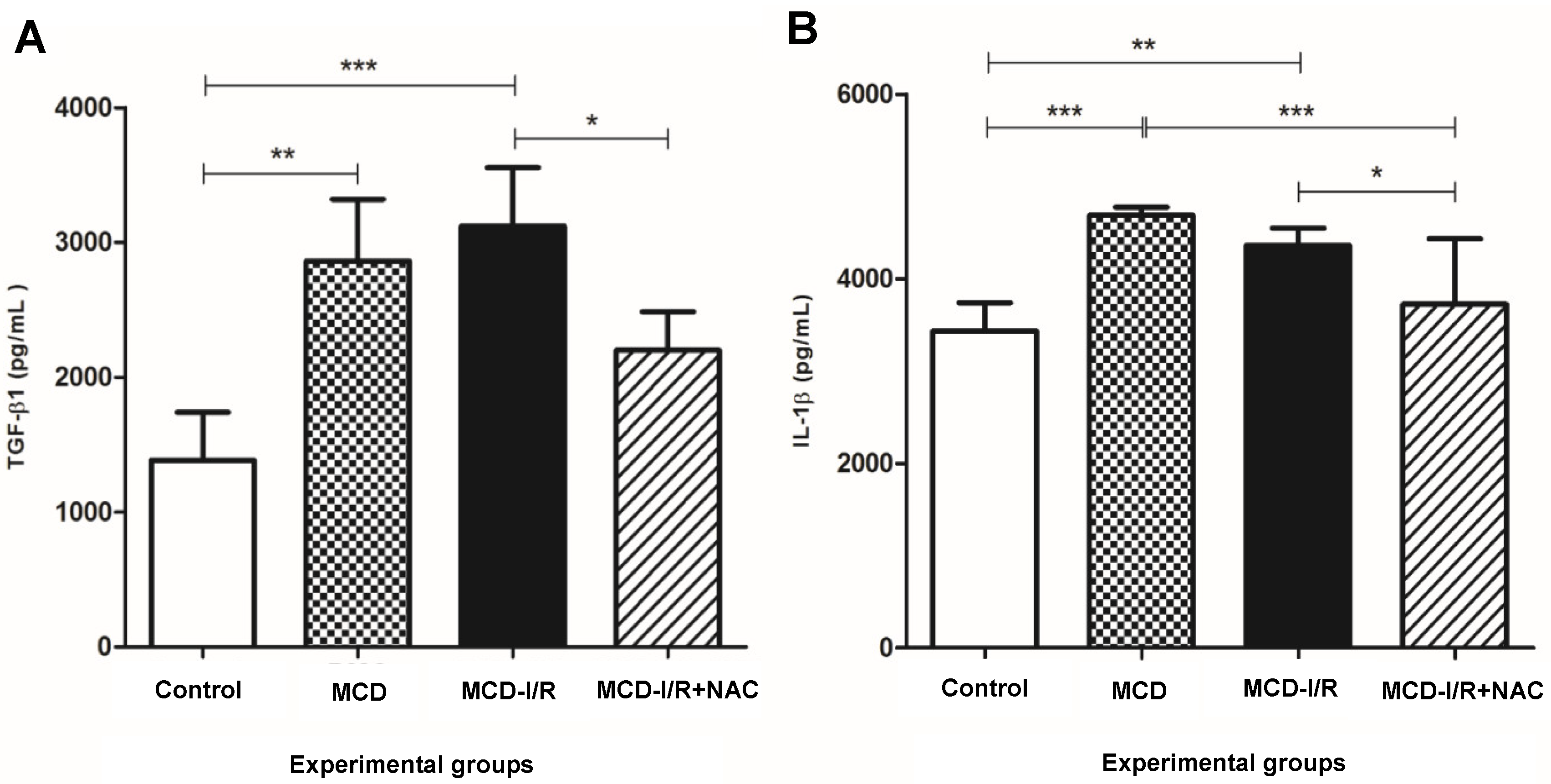
2.4. Hepatic Cytokines
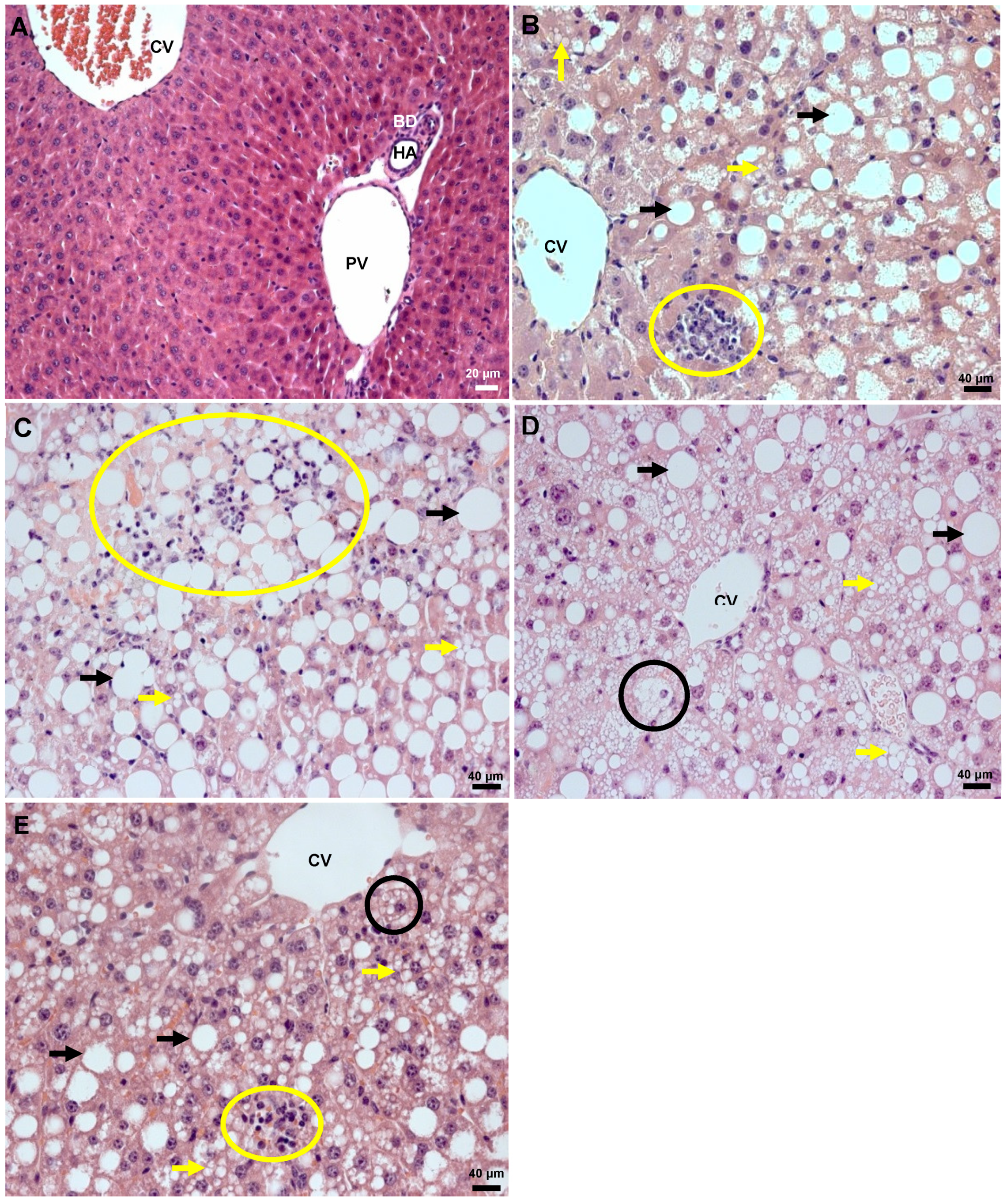
2.5. Histology
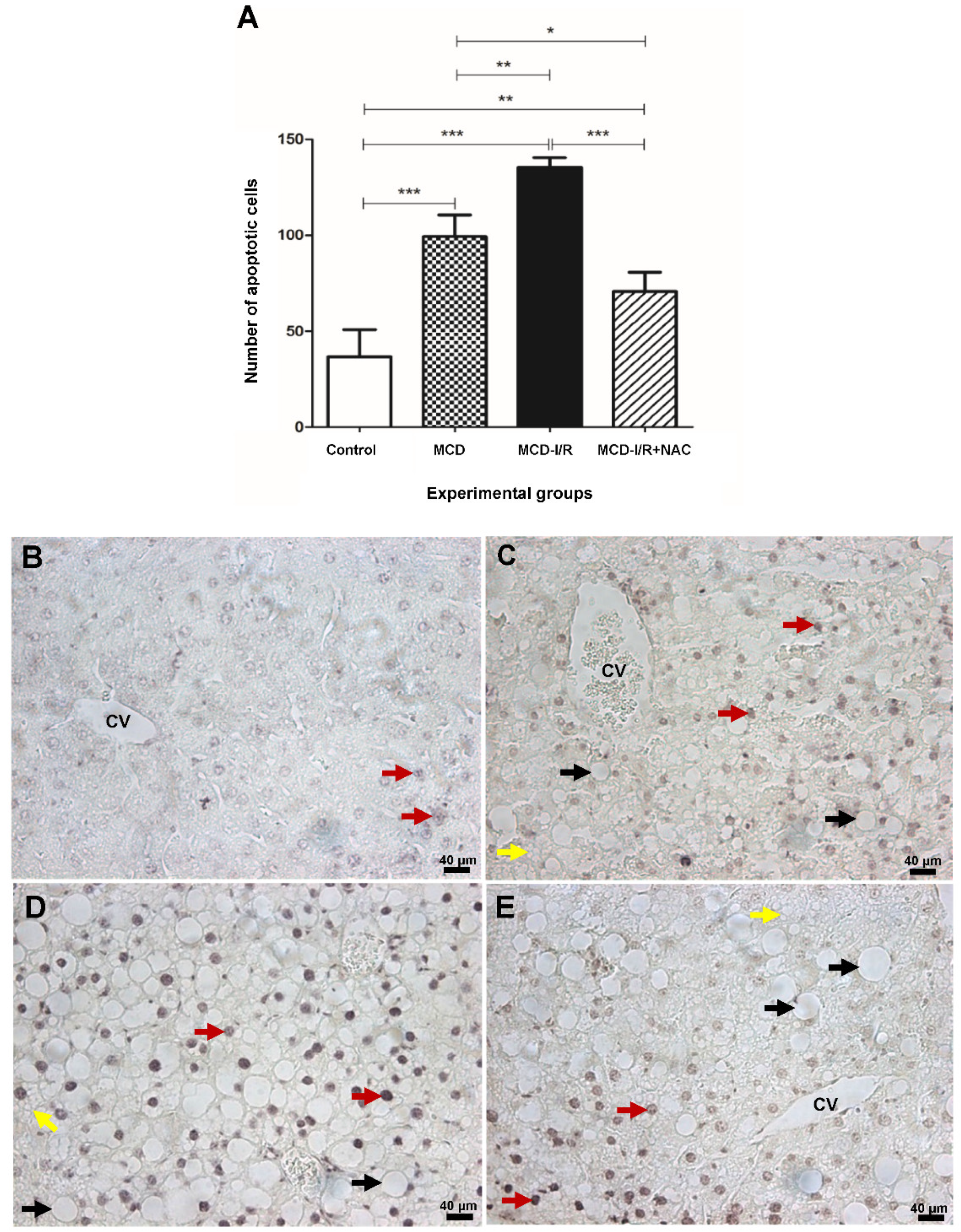
2.6. Apoptosis
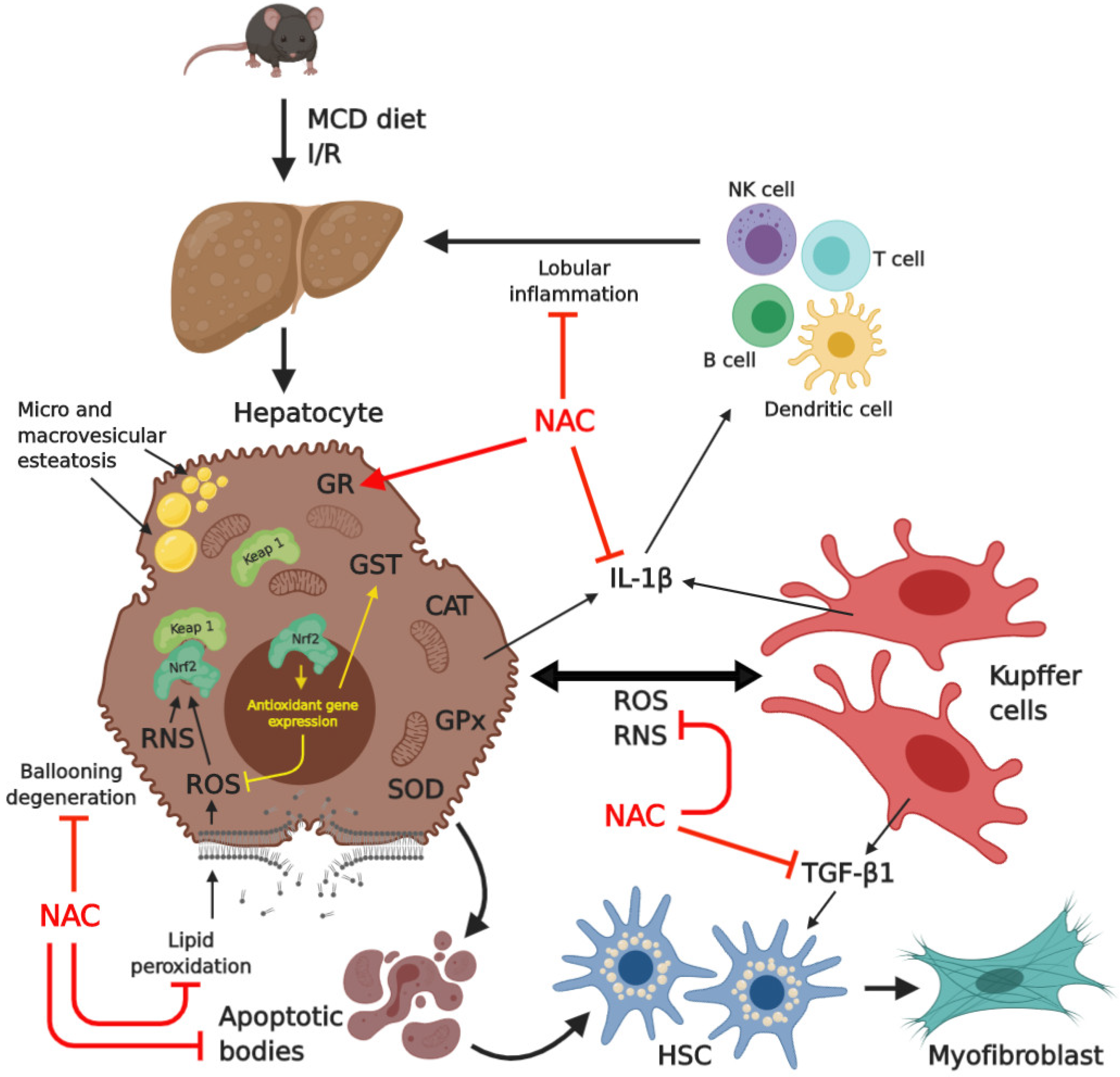
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Surgical Procedures
4.3. Hepatic Enzymes
4.4. Oxidative Stress
4.4.1. TBARS
4.4.2. Nitrite
4.4.3. Sample Preparation for Measurement of Antioxidant Enzymes
Determination of SOD Activity
Determination of CAT Activity
Determination of GPx Activity
Determination of GR Activity
Determination of GST Activity
Western Blotting
4.5. Hepatic Cytokines
4.6. Histology
4.7. Apoptosis
4.8. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALT | Alanine aminotransferase |
| ANOVA | Analysis of variance |
| ARE | Antioxidant response element |
| AST | Aspartate aminotransferase |
| BSA | Bovine serum albumin |
| CAT | Catalase |
| CEUA | Committee on ethics in animal use |
| DAB | Diaminobenzidine |
| ELISA | Enzyme-linked immunosorbent assay |
| FBS | Fetal bovine serum |
| FMUSP | Faculdade de Medicina da Universidade de São Paulo |
| GHS | Reduced glutathione |
| GPx | Glutathione peroxidase |
| GR | Glutathione reductase |
| GST | Glutathione S-transferase |
| HE | Hematoxylin and eosin |
| HFD | High fat diet |
| HSC | Hepatic stellate cells |
| ICHC-FMUSP | Instituto Central do Hospital das Clínicas-FMUSP |
| IFN-γ | Interferon gamma |
| IL-1β | Interleukin 1 beta |
| I/R | Ischemia/reperfusion |
| Keap1 | Kelch-like ECH-associated protein 1 |
| MCD | methionine- and choline-deficient |
| MCD-I/R+NAC | methionine- and choline-deficient with ischemia/reperfusion plus N-acetylcysteine |
| MCD-I/R | methionine- and choline-deficient with ischemia/reperfusion |
| NAC | N-acetylcysteine |
| MDA | Malondialdehyde |
| NAFLD | Nonalcoholic fatty liver disease |
| NASH | Nonalcoholic steatohepatitis |
| NF-κB | Nuclear factor κ-light-chain enhancer activated B cells |
| NK | Natural killer |
| Nrf2 | Nuclear factor erythroid-2-related factor 2 |
| PBS | Phosphate-buffered saline |
| RNS | Reactive nitrogen substances |
| ROS | Reactive oxygen substances |
| SOD | Superoxide dismutase |
| TBA | Thiobarbituric acid |
| TBARS | Thiobarbituric acid reactive substances |
| TCA | Trichloroacetic acid |
| TGF-β1 | Transforming growth factor beta 1 |
| TNF-α | Tumor necrosis factor alpha |
| TUNEL | Terminal deoxynucleotidyl transferase dUTP nick end labeling |
| USP | Universidade de São Paulo |
References
- Nassir, F.; Rector, R.S.; Hammoud, G.M.; Ibdah, J.A. Pathogenesis and Prevention of Hepatic Steatosis. Gastroenterol. Hepatol. 2015, 11, 167–175. [Google Scholar]
- Pham, T.; Dick, T.B.; Charlton, M.R. Nonalcoholic Fatty Liver Disease and Liver Transplantation. Clin. Liver Dis. 2016, 20, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Vernon, G.; Baranova, A.; Younossi, Z.M. Systematic review: The epidemiology and natural history of non-alcoholic fatty liver disease and non-alcoholic steato-hepatitis in adults. Aliment. Pharm. Ther. 2011, 34, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Sherif, Z.A.; Saeed, A.; Ghavimi, S.; Nouraie, S.M.; Laiyemo, A.O.; Brim, H.; Ash-ktorab, H. Global epidemiology of nonalcoholic fatty liver disease and perspectives on US minority populations. Dig. Dis. Sci. 2016, 61, 1214–1225. [Google Scholar] [CrossRef] [Green Version]
- Younossi, Z.M.; Koenig, A.B.; Abdellatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of non-alcoholic fatty liver disease a-analytic assessment of prevalence, incidence and outcomes. Hepatology 2015, 64, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Diehl, A.M.; Brunt, E.M.; Cusi, K.; Charlton, M.; Sanyal, A.J. The diagnosis and management of non-alcoholic fatty liver disease: Practice guideline by the American Association for the Study of Liver Diseases, American College of Gastroenterology, and the American Gastroenterological Association. Hepatology 2012, 55, 2005–2023. [Google Scholar] [CrossRef]
- Lefere, S.; Van Steenkiste, C.; Verhelst, X.; Van Vlierberghe, H.; Devisscher, L.; Geerts, A. Hypoxia-regulated mechanisms in the pathogenesis of obesity and non-alcoholic fatty liver disease. Cell. Mol. Life Sci. 2016, 73, 3419–3431. [Google Scholar] [CrossRef]
- Ramalho, F.S.; Fernandez-Monteiro, I.; Rosello-Catafau, J.; Peralta, C. Hepatic mi-crocirculatory failure. Acta Cir. Bras. 2006, 21, 48–53. [Google Scholar] [CrossRef]
- DuBray, B.J.; Conzen, K.D.; Upadhya, G.A.; Gunter, K.L.; Jia, J.; Knolhoff, B.L.; Monahakumar, T.; Chapman, W.C.; Anderson, C.D. BH3-only proteins contribute to steatotic liver ischemia and reperfusion injury. J. Surg. Res. 2015, 194, 653–658. [Google Scholar] [CrossRef] [Green Version]
- Tashiro, H.; Kuroda, S.; Mikuriya, Y.; Ohdan, H. Ischemia–reperfusion injury in patients with fatty liver and the clinical impact of steatotic liver on hepatic surgery. Surg. Today 2013, 44, 1611–1625. [Google Scholar] [CrossRef] [PubMed]
- McCormack, L.; Petrowsky, H.; Jochum, W.; Furrer, K.; Clavien, P.-A. Hepatic Steatosis Is a Risk Factor for Postoperative Complications After Major Hepatectomy. Ann. Surg. 2007, 245, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.K.; Marsh, J.W.; Varley, P.R.; Mock, B.K.; Chopra, K.B.; Geller, D.A.; Tsung, A. Underlying steatohepatitis, but not simple hepatic steatosis, increases morbidity after liver resection: A case-control study. Hepatology 2012, 56, 2221–2230. [Google Scholar] [CrossRef] [PubMed]
- Trevisani, F.; Colantoni, A.; Caraceni, P.; Van Thiel, D.H. The use of donor fatty liver for liver transplantation: A challenge or a quagmire? J. Hepatol. 1996, 24, 114–121. [Google Scholar] [CrossRef]
- Marsman, W.A.; Wiesner, R.H.; Rodriguez, I.; Batts, K.P.; Porayko, M.K.; Hay, J.E.; Gores, G.J.; Krom, R.A. Use of fatty donor liver is associated with diminished early patient and graft survival. Transplantation 1996, 62, 1246–1251. [Google Scholar] [CrossRef]
- Lee, L.-Y.; Harberg, C.; Matkowskyj, K.A.; Cook, S.; Roenneburg, D.; Werner, S.; Johnson, D.A.; Johnson, J.A.; Foley, D.P. Cell-specific over-activation of Nrf2-mediated gene expression in myeloid cells decreases hepatic ischemia reperfusion injury. Liver Transpl. 2016, 22, 1115–1128. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Castro, M.B.; Cornide-Petronio, M.E.; Gracia-Sancho, J.; Peralta, C. Inflammasome-mediated inflammation in liver ischemia-reperfusion injury. Cells 2019, 8, 1131. [Google Scholar] [CrossRef] [Green Version]
- Cohen-Naftaly, M.; Friedman, S.L. Currentstatus of novel antifibrotic therapies in patients with chronic liver disease. Ther. Adv. Gastroenterol. 2011, 4, 391–417. [Google Scholar] [CrossRef]
- Jiang, Y.; Tang, J.J.; Wu, B.Q.; Yuan, B.; Qu, Z. The protective effects of different-time-ischemic preconditioning on the reperfusion injury in fatty livers in rats. PLoS ONE 2013, 8, e58086. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-T.; Deng, J.-S.; Huang, W.-C.; Shieh, P.-C.; Chung, M.-I.; Huang, G.-J. Salvianolic acid C against acetaminophen-induced acute liver injury by attenuating inflammation, oxidative stress, and apoptosis through inhibition of the Keap1/Nrf2/HO-1 signaling. Oxidative Med. Cell Longev. 2019, 2019, 9056845. [Google Scholar] [CrossRef]
- Vargas-Mendoza, N.; Morales-González, A.; Madrigal-Santillán, E.O.; Madrigal-Bujaidar, E.; Álvarez-González, I.; García-Melo, L.F.; Anguiano-Robledo, L.; Fregoso-Aguilar, T.; Morales-Gonzalez, J.A. Antioxidant and adaptative response mediated by Nrf2 during physical exercise. Antioxidants 2019, 8, 196. [Google Scholar] [CrossRef] [Green Version]
- Winbladh, A.; Björnsson, B.; Trulsson, L.; Bojmar, L.; Sundqvist, T.; Gullstrand, P.; Sandström, P. N-acetylcysteine improves glycogenesis after segmental liver ischemia and reperfusion injury in pigs. Scand. J. Gastroenterol. 2012, 47, 225–236. [Google Scholar] [CrossRef]
- Seguro, A.C.; De Figueiredo, L.F.P.; Shimizu, M.H.M. N-acetylcysteine (NAC) Protects Against Acute Kidney Injury (AKI) Following Prolonged Pneumoperitoneum in the Rat. J. Surg. Res. 2012, 175, 312–315. [Google Scholar] [CrossRef]
- Saad, K.; Saad, P.F.; Filho, L.D.; De Brito, J.M.; Koike, M.; Zanoni, F.L.; Dolhnikoff, M.; Montero, E.F.D.S. Pulmonary impact of N-acetylcysteine in a controlled hemorrhagic shock model in rats. J. Surg. Res. 2013, 182, 108–115. [Google Scholar] [CrossRef]
- Saad, P.F.; Saad, K.; Filho, L.D.D.O.; Ferreira, S.G.; Koike, M.; Montero, E.F.D.S. Effect of N-acetylcysteine on pulmonary cell death in a controlled hemorrhagic shock model in rats. Acta Cir. Bras. 2012, 27, 561–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portella, A.O.V.; Montero, E.F.S.; Figueiredo, L.F.P.; Bueno, A.S.; Thurow, A.A.; Rodrigues, F.G. Effects of n-acetylcysteine in hepatic ischemia-reperfusion injury during hemorrhagic shock. Transplant. Proc. 2004, 36, 846–848. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Silva, S.M.; Simões, M.D.J.; Montero, E.F. Effect of n-acetylcysteine in liver ischemia-reperfusion injury after 30% hepatectomy in mice. Acta. Cir. Bras. 2012, 27, 346–349. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.M.; Carbonel, A.A.F.; Taha, M.O.; Simões, M.J.; Montero, E.F.S. Proliferative activity in ischemia/reperfusion injury in hepatectomized mice: Effect of N-acetylcysteine. Transpl. Proc. 2012, 44, 2321–2325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galhardo, M.A.; Quireze Júnior, C.; Navarro, P.G.R.; Morello, R.J.; Simões, M.J.; Montero, E.F.S. Liver and lung late alterations following hepatic reperfusion associated to ischemic preconditioning or N-acetylcysteine. Microsurgery 2007, 27, 295–299. [Google Scholar] [CrossRef]
- Samuhasaneeto, S.; Thong-Ngam, D.; Kulaputaba, O.; Patumraj, S.; Klaikeaw, N. Effects of N-acetylcysteine on oxidative stress in rats with non-alcoholic steatohepatitis. J. Med. Assoc. Thai. 2007, 90, 788–797. [Google Scholar]
- Baumgardner, J.N.; Shankar, K.; Hennings, L.; Albano, E.; Badger, T.M.; Ronis, M.J. N-acetylcysteine attenuates progression of liver pathology in a rat model of non-alcoholic steatohepatitis. J. Nutr. 2008, 138, 1872–1879. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.W.; Adams, L.A. Non-alcoholic fatty liver disease. Crit. Rev. Clin. Lab. Sci. 2011, 48, 97–113. [Google Scholar] [CrossRef]
- Zhou, Y.; Wei, F.; Fan, Y. High serum uric acid and risk of nonalcoholic fatty liver disease: A systematic review and meta-analysis. Clin. Biochem. 2016, 49, 636–642. [Google Scholar] [CrossRef]
- Amancher, D.E.; Schomaker, S.J.; Aubrecht, J. Development of blood biomarkers for drug-induced liver injury: An evaluation of their potential for risk assessment and diagnostics. Mol. Diagn. Ther. 2013, 17, 343–354. [Google Scholar] [CrossRef]
- Wang, C.; Chen, K.; Xia, Y.; Dai, W.; Wang, F.; Shen, M.; Cheng, P.; Wang, J.; Lu, J.; Zhang, Y.; et al. N-acetylcysteine attenuates ischemia-reperfusion-induced apoptosis and autophagy in mouse liver via regulation of the ROS/JNK/Bcl-2 pathway. PLoS ONE 2014, 9, e108855. [Google Scholar] [CrossRef]
- Ali, M.H.; Messiha, B.A.; Abdel-Latif, H.A. Protective effect of ursodeoxycholic acid, resveratrol, and N-acetylcysteine on nonalcoholic fatty liver disease in rats. Pharm. Biol. 2015, 54, 1198–1208. [Google Scholar] [CrossRef]
- Nasiri, M.; Karimi, M.H.; Azarpira, N.; Saadat, I. Gene Expression profile of Toll-like receptor/adaptor/interferon regulatory factor/cytokine axis during liver regeneration after partial ischemia-reperfusion injury. Exp. Clin. Transplant. 2020, 18, 215–223. [Google Scholar] [CrossRef]
- Del Campo, J.A.; Gallego, P.; Grande, L. Role of inflammatory response in liver diseases: Therapeutic strategies. World J. Hepatol. 2018, 10, 1–7. [Google Scholar] [CrossRef]
- Sun, Y.; Pu, L.Y.; Wang, X.H.; Zhang, F.; Rao, J.H. N-acetylcysteine attenuates re-active-oxygen-species-mediated endoplasmic reticulum stress during liver ischemia-reperfusion injury. World J. Gastroenterol. 2014, 20, 15289–15298. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.C.; Hsieh, S.C.; Chiu, J.H.; Wu, Y.L. Protective Effects of N-acetylcysteine and a Prostaglandin E1 Analog, Alprostadil, Against Hepatic Ischemia: Reperfusion Injury in Rats. J. Tradit. Complement. Med. 2014, 4, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Fusai, G.; Glantzouniss, G.K.; Hafez, T.; Yang, W.; Quaglia, A.; Sheth, H.; Kanoria, S.; Parkes, R.; Seifalian, A.; Davidson, B.R. N-acetycysteine ameliorates the late phase of liver ischaemia/reperfusion injury in rabbit with hepatic steatosis. Clin. Sci. 2005, 109, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Nakano, H.; Nagasaki, H.; Yoshida, K.; Kigawa, G.; Fujiwara, Y.; Kitamura, N.; Takeuchi, S.; Sasaki, J.; Shimura, H.; Yamaguchi, M.; et al. N-acetylcysteine and anti-ICAM-1 monoclonal antibody reduce ischemia-reperfusion injury of the steatotic rat liver. Transplant. Proc. 1998, 30, 3763. [Google Scholar] [CrossRef]
- Rushworth, G.F.; Megson, I.L. Existing and potential therapeutic uses for N-acetylcysteine: The need for conversion to intracellular glutathione for antioxidant benefits. Pharmacol. Ther. 2014, 141, 150–159. [Google Scholar] [CrossRef]
- Demir, S.; Inal-Erden, M. Pentoxifylline and N-acetylcysteine in hepatic ischemia/reperfusion injury. Clin. Chim. Acta 1998, 275, 127–135. [Google Scholar] [CrossRef]
- Georgious-Siafis, S.K.; Samiotaki, M.K.; Demopoulos, V.J.; Panayotou, G.; Tsiftsoglou, A.S. Formation of novel N-acetylcysteine-hemi adducts abrogates hemi-induced cytotoxicity ans suppresses the Nrf2-driven stress response in human pro-erythroid K562 cells. Eur. J. Pharmacol. 2020, 25, 173077. [Google Scholar] [CrossRef]
- Romanque, P.; Cornejo, P.; Valdés, S.; Videla, L.A. Thyroid hormone administration induces rat liver Nrf2 activation: Suppression by N-acetylcysteine pretreatment. Thyroid 2011, 21, 655–662. [Google Scholar] [CrossRef]
- Li, J.; Sapper, T.N.; Mah, E.; Rudraiah, S.; Schill, K.E.; Chitchumroonchokchai, C.; Moller, M.V.; McDonald, J.D.; Rohrer, P.R.; Manautou, J.E.; et al. Green tea extract provides extensive Nrf2-independent protection against lipid accumulation and NFB proinflammatory responses during nonalcoholic steatohepatitis in mice fed a high-fat diet. Mol. Nutr. Food Res. 2016, 60, 858–870. [Google Scholar] [CrossRef]
- Chambel, S.S.; Santos-Gonçalves, A.; Duarte, T.L. The dual role of Nrf2 in nonalcoholic fatty liver disease: Regulation of antioxidant defenses and hepatic lipid metabolism. Biomed. Res. Int. 2015, 2015, 597134. [Google Scholar] [CrossRef] [Green Version]
- Charron, M.J.; Williams, L.; Seki, Y.; Du, X.Q.; Chaurasia, B.; Saghatelian, A.; Summers, S.A.; Katz, E.B.; Vuguin, P.M.; Reznik, S.E. Antioxidant Effects of N-Acetylcysteine Prevent Programmed Metabolic Disease in Mice. Diabetes. 2020, db191129. [Google Scholar] [CrossRef]
- Fan, H.; Le, J.W.; Zhu, J.H. Protective Effect of N-Acetylcysteine Pretreatment on Acute Kidney Injury in Septic Rats. J. Surg. Res. 2020, 254, 125–134. [Google Scholar] [CrossRef]
- Kretzmann, N.A.; Filippi-Chiela, E.; Matte, U.; Marroni, N.P.; Marroni, C.A. N-acetylcysteine improves antitumoural response of Interferon alpha by NF-kB downregulation in liver cancer cells. Comp. Hepatol. 2012, 11, 4. [Google Scholar] [CrossRef] [Green Version]
- Alexandropoulos, D.; Bazigos, G.V.; Doulamis, I.P.; Tzani, A.; Konstantopoulos, P.; Tragotsalou, N.; Kondi-Pafiti, A.; Kotsis, T.; Arkadopoulos, N.; Smyrniotis, V.; et al. Protective effects of N -acetylcystein and atorvastatin against renal and hepatic injury in a rat model of intestinal ischemia-reperfusion. Biomed. Pharmacother. 2017, 89, 673–680. [Google Scholar] [CrossRef] [PubMed]
- El-Lakkany, N.M.; Din, S.H.S.; Sabra, A.-N.A.-L.; Hammam, O.A.; Ebeid, F.A.-L. Co-administration of metformin and N-acetylcysteine with dietary control improves the biochemical and histological manifestations in rats with non-alcoholic fatty liver. Res. Pharm. Sci. 2016, 11, 374–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lappas, C.M.; Day, Y.-J.; Marshall, M.A.; Engelhard, V.H.; Linden, J. Adenosine A2A receptor activation reduces hepatic ischemia reperfusion injury by inhibiting CD1d-dependent NKT cell activation. J. Cell Boil. 2006, 175, 2639–2648. [Google Scholar] [CrossRef]
- Olthof, P.B.; Van Golen, R.F.; Meijer, B.; Van Beek, A.A.; Bennink, R.J.; Verheij, J.; Van Guik, T.M.; Heger, M. Warm ischemia time.dependent variation in liver damage, inflammation, and function in hepatic ischemia/reperfusion injury. Biochim. Biophys. Acta 2016, 1863, 375–385. [Google Scholar] [CrossRef]
- Ellet, J.D.; Evans, Z.P.; Atkinson, C.; Schmidt, M.G.; Schnellmann, R.G.; Chavin, K. Toll-like receptor 4 is a key mediator of murine steatotic liver warm ischemia/reperfusion injury. Liver Transplant. 2009, 15, 1101–1109. [Google Scholar] [CrossRef] [Green Version]
- Weigand, K.; Brost, S.; Steinebrunner, N.; Büchler, M.; Schemmer, P.; Müller, M. Ischemia/Reperfusion Injury in Liver Surgery and Transplantation: Pathophysiology. HPB Surg. 2012, 2012, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.J.; Toledo-Pereyra, L.H. Toll-like receptor signaling in liver ischemia and reperfusion. J. Invest. Surg. 2012, 25, 271–277. [Google Scholar] [CrossRef]
- Wu, M.-Y.; Yiang, G.-T.; Liao, W.-T.; Tsai, A.P.-Y.; Cheng, Y.-L.; Cheng, P.-W.; Li, C.-Y.; Yiang, G.-T. Current Mechanistic Concepts in Ischemia and Reperfusion Injury. Cell. Physiol. Biochem. 2018, 46, 1650–1667. [Google Scholar] [CrossRef]
- Shi, T.; Yang, X.; Zhou, H.; Xi, J.; Sun, J.; Ke, Y.; Zhang, J.; Shao, Y.; Jiang, X.; Pan, X.; et al. Activated carbon N-acetylcysteine microcapsule protects against nonalcoholic fatty liver disease in young rats via activating telomerase and inhibiting apoptosis. PLoS ONE 2018, 13, e0189856. [Google Scholar] [CrossRef] [Green Version]
- Ukeda, H.; Maeda, S.; Ishii, T.; Sawamura, M. Spectrophotometric assay for superoxide dismutase based on tetrazolium salt 3’-1-(phenylamino)-carbonyl-3, 4-tetrazolium]-bis(4-methoxy-6-nitro)benzenesulfonic acid hydrate reduction by xanthine-xanthine oxidase. Anal. Biochem. 1997, 251, 206–209. [Google Scholar] [CrossRef]
- Flohé, L.; Günzler, W.A. Assay of glutathione peroxidase. Methods Enzymol. 1984, 105, 114–121. [Google Scholar]
- Carlberg, I.; Mannervik, B. Glutathione reductase. Methods Enzymol. 1985, 113, 484–490. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Fleischener, G.; Gatmaitan, Z.; Arias, I.M.; Jakoby, W.B. The identity of Glutthione-S-transferase B with ligandin, a major binding protein of liver. Proc. Natl. Acad Sci. USA 1974, 71, 3879–3882. [Google Scholar] [CrossRef] [Green Version]
- Rong, Y.; Baudry, M. Seizure activity results in a rapid induction of nuclear factor-kappa B in adult but not juvenile rat limbic structures. J. Neurochem. 1996, 67, 662–668. [Google Scholar] [CrossRef]
- Munhoz, C.D.; Lepsch, L.B.; Kawamoto, E.; Malta, M.B.; Lima, L.D.S.; Avellar, M.C.W.; Sapolsky, R.M.; Scavone, C. Chronic Unpredictable Stress Exacerbates Lipopolysaccharide-Induced Activation of Nuclear Factor-κB in the Frontal Cortex and Hippocampus via Glucocorticoid Secretion. J. Neurosci. 2006, 26, 3813–3820. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Brunt, E.M.; Van, N.M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrel, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Nonalcoholic Steatohepatitis Clinical Research Network. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Histologic Feature of NAFLD | Category | % Responses in the Groups (N = 4–6, per Group) | |||
|---|---|---|---|---|---|
| Control | MCD | MCD-I/R | MCD-I/R+NAC | ||
| Steatosis Grade | <5% | 25% | |||
| 5–33% | 50% | ||||
| >33–66% | 25% | 20% | 25% | ||
| >66% | 80% | 100% | 75% | ||
| Location | Periportal (zone 1) | 100% | 75% | ||
| Panacinar | 100% | 100% | 25% | ||
| Microvesicular steatosis | Present | 100% | 100% | 100% | 100% |
| Macrovesicular steatosis | Present | 0% | 100% | 100% | 100% |
| Lobular inflammation | No foci | 25% | |||
| <2 foci | 25% | ||||
| 2–4 foci | 50% | 100% | |||
| >4 foci | 100% | 100% | |||
| Ballooning degeneration | None | 100% | 80% | 66% | 75% |
| Few | 0% | 20% | 34% | 25% | |
| NASH | Present | 0% | 100% | 100% | 100% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaves Cayuela, N.; Kiyomi Koike, M.; Jacysyn, J.d.F.; Rasslan, R.; Azevedo Cerqueira, A.R.; Pereira Costa, S.K.; Picanço Diniz-Júnior, J.A.; Massazo Utiyama, E.; Frasson de Souza Montero, E. N-Acetylcysteine Reduced Ischemia and Reperfusion Damage Associated with Steatohepatitis in Mice. Int. J. Mol. Sci. 2020, 21, 4106. https://doi.org/10.3390/ijms21114106
Chaves Cayuela N, Kiyomi Koike M, Jacysyn JdF, Rasslan R, Azevedo Cerqueira AR, Pereira Costa SK, Picanço Diniz-Júnior JA, Massazo Utiyama E, Frasson de Souza Montero E. N-Acetylcysteine Reduced Ischemia and Reperfusion Damage Associated with Steatohepatitis in Mice. International Journal of Molecular Sciences. 2020; 21(11):4106. https://doi.org/10.3390/ijms21114106
Chicago/Turabian StyleChaves Cayuela, Natalie, Marcia Kiyomi Koike, Jacqueline de Fátima Jacysyn, Roberto Rasslan, Anderson Romério Azevedo Cerqueira, Soraia Katia Pereira Costa, José Antônio Picanço Diniz-Júnior, Edivaldo Massazo Utiyama, and Edna Frasson de Souza Montero. 2020. "N-Acetylcysteine Reduced Ischemia and Reperfusion Damage Associated with Steatohepatitis in Mice" International Journal of Molecular Sciences 21, no. 11: 4106. https://doi.org/10.3390/ijms21114106
APA StyleChaves Cayuela, N., Kiyomi Koike, M., Jacysyn, J. d. F., Rasslan, R., Azevedo Cerqueira, A. R., Pereira Costa, S. K., Picanço Diniz-Júnior, J. A., Massazo Utiyama, E., & Frasson de Souza Montero, E. (2020). N-Acetylcysteine Reduced Ischemia and Reperfusion Damage Associated with Steatohepatitis in Mice. International Journal of Molecular Sciences, 21(11), 4106. https://doi.org/10.3390/ijms21114106