Ablation of the miRNA Cluster 24 Has Profound Effects on Extracellular Matrix Protein Abundance in Cartilage

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Mirc24 Cluster Is Expressed in Articular Cartilage and Downregulated in Experimental OA
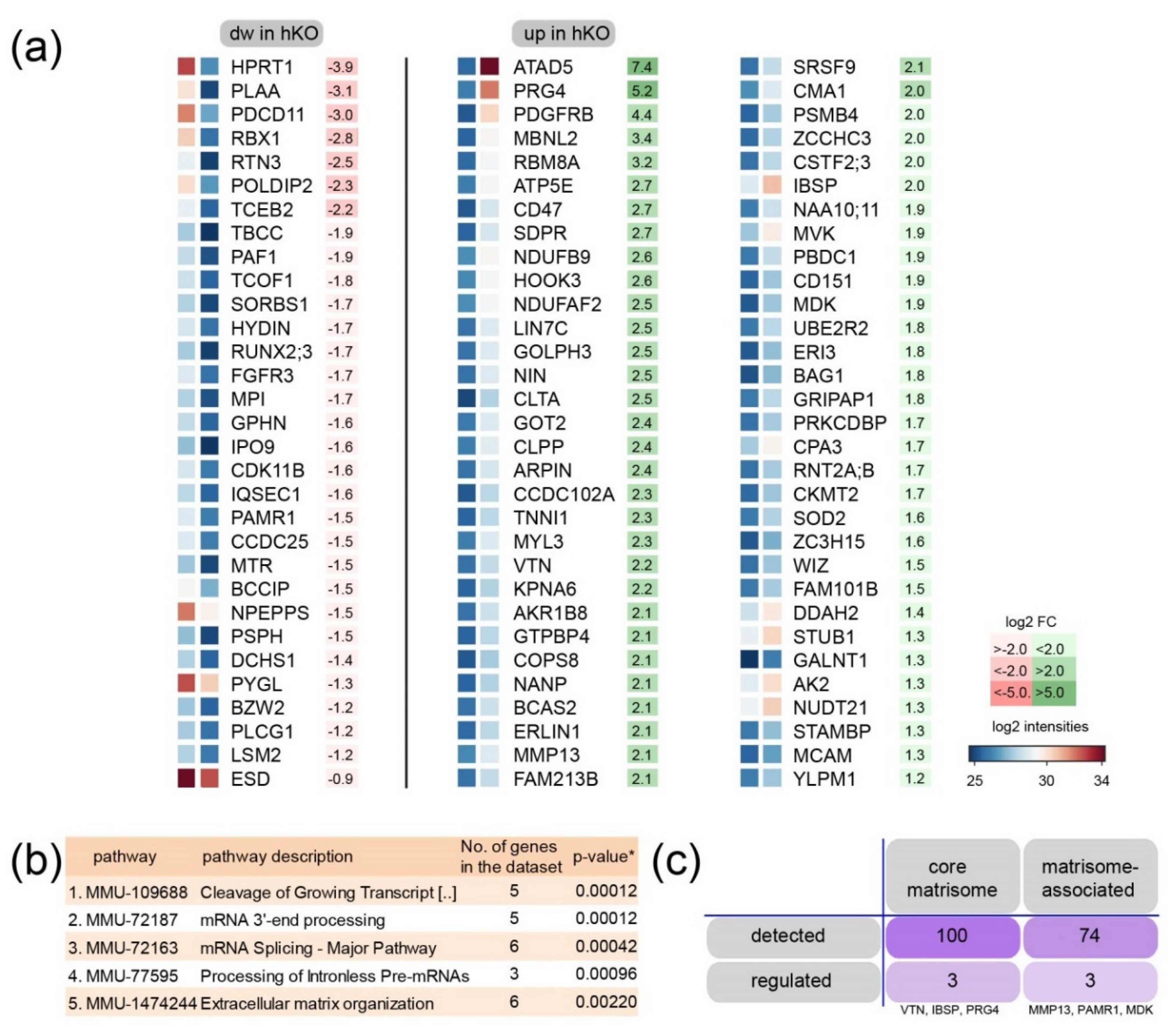
2.2. ECM Entities Are Enriched in Proteome Analysis of Mirc24 Cluster-Deficient Epiphyseal Cartilage Extracts
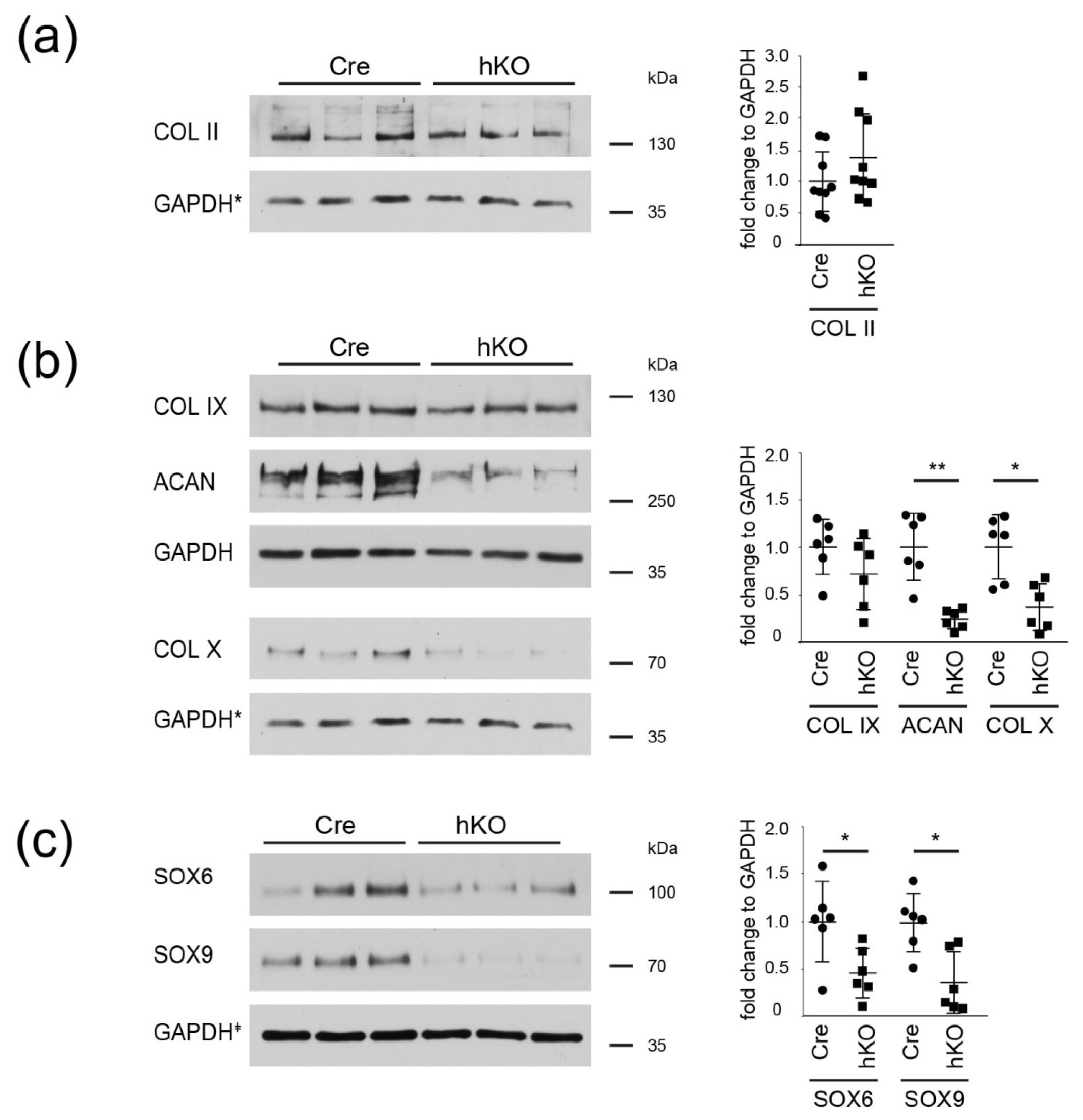
2.3. ECM Composition and SOX 6/9 Expression Is Altered in Mirc24-Cluster Deficient Epiphyseal Cartilage
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. In Situ Detection of LacZ Reporter Gene Activity
4.3. miRNA Expression Analysis in the GEO Dataset GSE93008
4.4. Protein Extraction
4.5. Proteome Analysis
4.6. RNA Isolation and Semiquantitative PCR
4.7. Fluorescent In Situ Hybridization Analysis
4.8. Immunoblot Analysis
4.9. Gelatin Zymography
4.10. In Situ Zymography
4.11. Immunofluorescence Analysis
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACAN | Aggrecan |
| ADAM | A disintegrin and metalloproteinase |
| ADAMTS | ADAM with thrombospondin motifs |
| COL | Collagen |
| DMM | Destabilization of the medial meniscus |
| DTT | Dithiothreitol |
| ECM | Extracellular matrix |
| EDTA | Ethylenediaminetetraacetic acid |
| EGTA | Ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid |
| FC | Fold change |
| FCS | Fetal calf serum |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GuHCl | Guanidinium hydrochloride |
| hKO | Hemizygous knock-out |
| IBSP | Bone sialoprotein 2 |
| MDK | Midkine |
| MgCl2 | Magnesium chloride |
| miR, miRNA | MicroRNA |
| MMP | Matrix metallopeptidase |
| MS | Mass spectrometry |
| OA | Osteoarthritis |
| SDS | Sodium dodecyl sulfate |
| SOX | SRY-related HMG box-containing |
| TBS | Tris-buffered saline |
| VTN | Vitronectin |
References
- Endisha, H.; Rockel, J.; Jurisica, I.; Kapoor, M. The complex landscape of microRNAs in articular cartilage: Biology, pathology, and therapeutic targets. JCI insight 2018, 3, e121630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kung, L.H.; Zaki, S.; Ravi, V.; Rowley, L.; Smith, M.M.; Bell, K.M.; Bateman, J.F.; Little, C.B. Utility of circulating serum miRNAs as biomarkers of early cartilage degeneration in animal models of post-traumatic osteoarthritis and inflammatory arthritis. Osteoarthr. Cartil. 2017, 25, 426–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyaki, S.; Sato, T.; Inoue, A.; Otsuki, S.; Ito, Y.; Yokoyama, S.; Kato, Y.; Takemoto, F.; Nakasa, T.; Yamashita, S.; et al. MicroRNA-140 plays dual roles in both cartilage development and homeostasis. Genes Dev. 2010, 24, 1173–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aszodi, A.; Bateman, J.F.; Gustafsson, E.; Boot-Handford, R.; Fassler, R. Mammalian skeletogenesis and extracellular matrix: What can we learn from knockout mice? Cell Struct. Funct. 2000, 25, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etich, J.; Holzer, T.; Pitzler, L.; Bluhm, B.; Brachvogel, B. MiR-26a modulates extracellular matrix homeostasis in cartilage. Matrix. Biol. 2015, 43, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Bluhm, B.; Ehlen, H.W.A.; Holzer, T.; Georgieva, V.S.; Heilig, J.; Pitzler, L.; Etich, J.; Bortecen, T.; Frie, C.; Probst, K.; et al. miR-322 stabilizes MEK1 expression to inhibit RAF/MEK/ERK pathway activation in cartilage. Development 2017, 144, 3562–3577. [Google Scholar] [CrossRef] [Green Version]
- Geng, H.; Song, Q.; Cheng, Y.; Li, H.; Yang, R.; Liu, S.; Hao, L. MicroRNA 322 Aggravates Dexamethasone-Induced Muscle Atrophy by Targeting IGF1R and INSR. Int. J. Mol. Sci. 2020, 21, 1111. [Google Scholar] [CrossRef] [Green Version]
- Marchand, A.; Atassi, F.; Mougenot, N.; Clergue, M.; Codoni, V.; Berthuin, J.; Proust, C.; Tregouet, D.A.; Hulot, J.S.; Lompre, A.M. miR-322 regulates insulin signaling pathway and protects against metabolic syndrome-induced cardiac dysfunction in mice. Biochim. Biophys. Acta 2016, 1862, 611–621. [Google Scholar] [CrossRef]
- Zhao, M.J.; Xie, J.; Shu, W.J.; Wang, H.Y.; Bi, J.; Jiang, W.; Du, H.N. MiR-15b and miR-322 inhibit SETD3 expression to repress muscle cell differentiation. Cell Death Dis. 2019, 10, 183. [Google Scholar] [CrossRef] [Green Version]
- Hou, Q.; Huang, Y.; Luo, Y.; Wang, B.; Liu, Y.; Deng, R.; Zhang, S.; Liu, F.; Chen, D. MiR-351 negatively regulates osteoblast differentiation of MSCs induced by (+)-cholesten-3-one through targeting VDR. Am. J. Transl. Res. 2017, 9, 4963–4973. [Google Scholar]
- He, X.; Sun, Y.; Lei, N.; Fan, X.; Zhang, C.; Wang, Y.; Zheng, K.; Zhang, D.; Pan, W. MicroRNA-351 promotes schistosomiasis-induced hepatic fibrosis by targeting the vitamin D receptor. Proc. Natl. Acad. Sci. USA 2018, 115, 180–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Xu, J.; Xu, L.; Zhang, J.; Chan, K.; Pan, X.; Li, G. MiR-503 Promotes Bone Formation in Distraction Osteogenesis through Suppressing Smurf1 Expression. Sci. Rep. 2017, 7, 409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kung LH, W.; Ravi, V.; Rowley, L.; Angelucci, C.; Fosang, A.J.; Bell, K.M.; Little, C.B.; Bateman, J.F. Cartilage MicroRNA Dysregulation During the Onset and Progression of Mouse Osteoarthritis Is Independent of Aggrecanolysis and Overlaps With Candidates From End-Stage Human Disease. Arthritis Rheumatol. 2018, 70, 383–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brachvogel, B.; Zaucke, F.; Dave, K.; Norris, E.L.; Stermann, J.; Dayakli, M.; Koch, M.; Gorman, J.J.; Bateman, J.F.; Wilson, R. Comparative proteomic analysis of normal and collagen IX null mouse cartilage reveals altered extracellular matrix composition and novel components of the collagen IX interactome. J. Biol. Chem. 2013, 288, 13481–13492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic. Acids. Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Fabregat, A.; Sidiropoulos, K.; Garapati, P.; Gillespie, M.; Hausmann, K.; Haw, R.; Jassal, B.; Jupe, S.; Korninger, F.; McKay, S.; et al. The Reactome pathway Knowledgebase. Nucleic. Acids. Res. 2016, 44, D481–D487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etich, J.; Koch, M.; Wagener, R.; Zaucke, F.; Fabri, M.; Brachvogel, B. Gene Expression Profiling of the Extracellular Matrix Signature in Macrophages of Different Activation Status: Relevance for Skin Wound Healing. Int. J. Mol. Sci. 2019, 20, 5086. [Google Scholar] [CrossRef] [Green Version]
- Bergmeier, V.; Etich, J.; Pitzler, L.; Frie, C.; Koch, M.; Fischer, M.; Rappl, G.; Abken, H.; Tomasek, J.J.; Brachvogel, B. Identification of a myofibroblast-specific expression signature in skin wounds. Matrix. Biol. 2018, 65, 59–74. [Google Scholar] [CrossRef]
- Steele, B.L.; Alvarez-Veronesi, M.C.; Schmidt, T.A. Molecular weight characterization of PRG4 proteins using multi-angle laser light scattering (MALLS). Osteoarthr. Cartil. 2013, 21, 498–504. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4. [Google Scholar] [CrossRef]
- Stickens, D.; Behonick, D.J.; Ortega, N.; Heyer, B.; Hartenstein, B.; Yu, Y.; Fosang, A.J.; Schorpp-Kistner, M.; Angel, P.; Werb, Z. Altered endochondral bone development in matrix metalloproteinase 13-deficient mice. Development 2004, 131, 5883–5895. [Google Scholar] [CrossRef] [Green Version]
- Knauper, V.; Cowell, S.; Smith, B.; Lopez-Otin, C.; O’Shea, M.; Morris, H.; Zardi, L.; Murphy, G. The role of the C-terminal domain of human collagenase-3 (MMP-13) in the activation of procollagenase-3, substrate specificity, and tissue inhibitor of metalloproteinase interaction. J. Biol. Chem. 1997, 272, 7608–7616. [Google Scholar] [CrossRef] [Green Version]
- Swaney, D.L.; Wenger, C.D.; Coon, J.J. Value of using multiple proteases for large-scale mass spectrometry-based proteomics. J. Proteome Res. 2010, 9, 1323–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akiyama, H.; Chaboissier, M.C.; Martin, J.F.; Schedl, A.; de Crombrugghe, B. The transcription factor Sox9 has essential roles in successive steps of the chondrocyte differentiation pathway and is required for expression of Sox5 and Sox6. Genes Dev. 2002, 16, 2813–2828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Lefebvre, V. L-Sox5 and Sox6 drive expression of the aggrecan gene in cartilage by securing binding of Sox9 to a far-upstream enhancer. Mol. Cell Biol. 2008, 28, 4999–5013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dy, P.; Wang, W.; Bhattaram, P.; Wang, Q.; Wang, L.; Ballock, R.T.; Lefebvre, V. Sox9 directs hypertrophic maturation and blocks osteoblast differentiation of growth plate chondrocytes. Dev. Cell 2012, 22, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Park, C.Y.; Jeker, L.T.; Carver-Moore, K.; Oh, A.; Liu, H.J.; Cameron, R.; Richards, H.; Li, Z.; Adler, D.; Yoshinaga, Y.; et al. A resource for the conditional ablation of microRNAs in the mouse. Cell Rep. 2012, 1, 385–391. [Google Scholar] [CrossRef] [Green Version]
- Ovchinnikov, D.A.; Deng, J.M.; Ogunrinu, G.; Behringer, R.R. Col2a1-directed expression of Cre recombinase in differentiating chondrocytes in transgenic mice. Genesis 2000, 26, 145–146. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell Proteomics 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [Green Version]
- Nolte, H.; MacVicar, T.D.; Tellkamp, F.; Krüger, M. Instant Clue: A Software Suite for Interactive Data Visualization and Analysis. Sci. Rep. 2018, 8, 12648. [Google Scholar] [CrossRef] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic. Acids. Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]
- Budde, B.; Blumbach, K.; Ylostalo, J.; Zaucke, F.; Ehlen, H.W.; Wagener, R.; Ala-Kokko, L.; Paulsson, M.; Bruckner, P.; Grässel, S. Altered integration of matrilin-3 into cartilage extracellular matrix in the absence of collagen IX. Mol. Cell Biol. 2005, 25, 10465–10478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girkontaite, I.; Frischholz, S.; Lammi, P.; Wagner, K.; Swoboda, B.; Aigner, T.; Von der Mark, K. Immunolocalization of type X collagen in normal fetal and adult osteoarthritic cartilage with monoclonal antibodies. Matrix. Biol. 1996, 15, 231–238. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Degasperi, A.; Birtwistle, M.R.; Volinsky, N.; Rauch, J.; Kolch, W.; Kholodenko, B.N. Evaluating strategies to normalise biological replicates of Western blot data. PLoS ONE 2014, 9, e87293. [Google Scholar] [CrossRef] [Green Version]
- Heilig, J.; Dietmar, H.F.; Brachvogel, B.; Paulsson, M.; Zaucke, F.; Niehoff, A. Collagen IX deficiency leads to premature vascularization and ossification of murine femoral heads through an imbalance of pro- and antiangiogenic factors. Osteoarthr. Cartil. 2020. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Georgieva, V.S.; Etich, J.; Bluhm, B.; Zhu, M.; Frie, C.; Wilson, R.; Zaucke, F.; Bateman, J.; Brachvogel, B. Ablation of the miRNA Cluster 24 Has Profound Effects on Extracellular Matrix Protein Abundance in Cartilage. Int. J. Mol. Sci. 2020, 21, 4112. https://doi.org/10.3390/ijms21114112
Georgieva VS, Etich J, Bluhm B, Zhu M, Frie C, Wilson R, Zaucke F, Bateman J, Brachvogel B. Ablation of the miRNA Cluster 24 Has Profound Effects on Extracellular Matrix Protein Abundance in Cartilage. International Journal of Molecular Sciences. 2020; 21(11):4112. https://doi.org/10.3390/ijms21114112
Chicago/Turabian StyleGeorgieva, Veronika S., Julia Etich, Björn Bluhm, Mengjie Zhu, Christian Frie, Richard Wilson, Frank Zaucke, John Bateman, and Bent Brachvogel. 2020. "Ablation of the miRNA Cluster 24 Has Profound Effects on Extracellular Matrix Protein Abundance in Cartilage" International Journal of Molecular Sciences 21, no. 11: 4112. https://doi.org/10.3390/ijms21114112
APA StyleGeorgieva, V. S., Etich, J., Bluhm, B., Zhu, M., Frie, C., Wilson, R., Zaucke, F., Bateman, J., & Brachvogel, B. (2020). Ablation of the miRNA Cluster 24 Has Profound Effects on Extracellular Matrix Protein Abundance in Cartilage. International Journal of Molecular Sciences, 21(11), 4112. https://doi.org/10.3390/ijms21114112