Pre-Growth Culture Conditions Affect Type 1 Fimbriae-Dependent Adhesion of Salmonella


Abstract
:1. Introduction
2. Results
2.1. Adhesion of Salmonella to IPEC-J2 Cells Depended on Pre-Infection Bacterial Growth Conditions
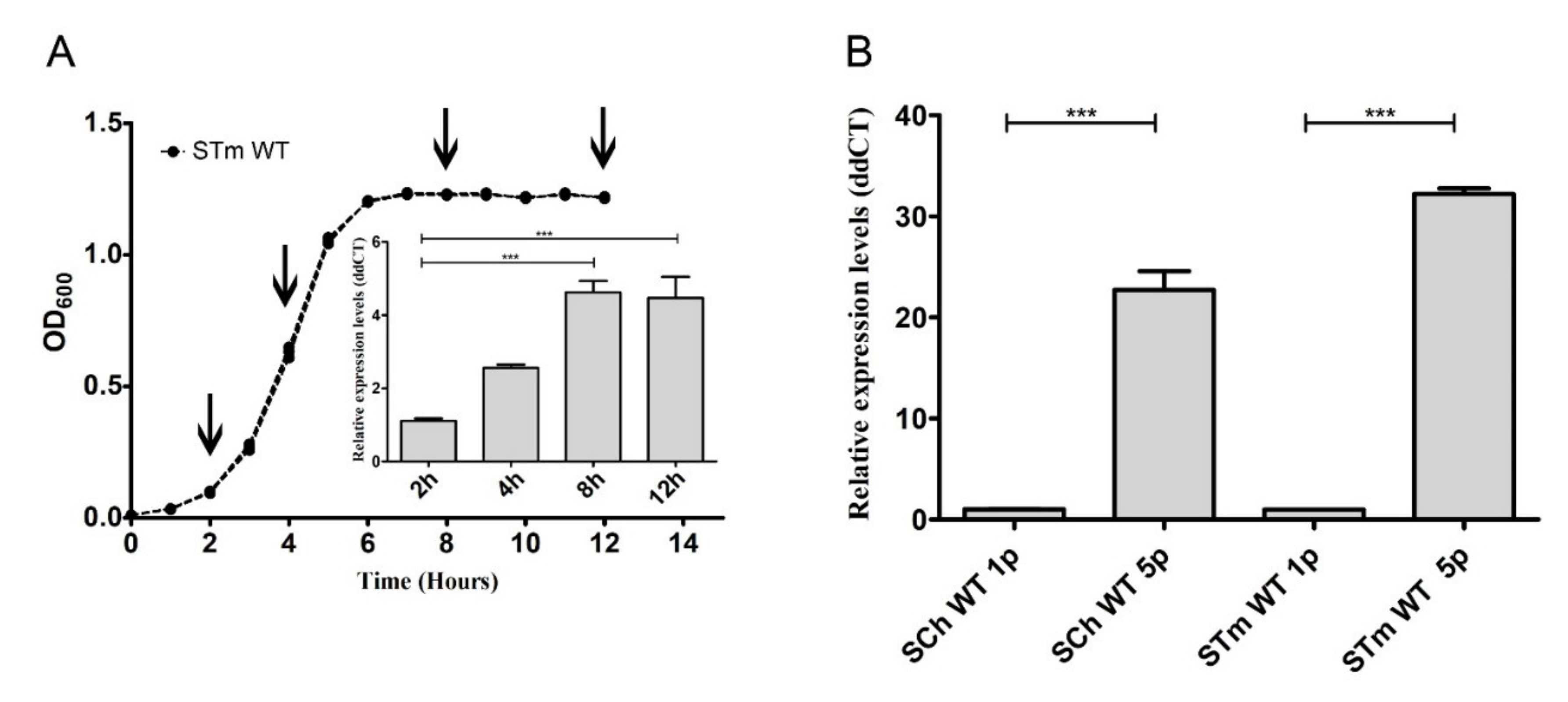
2.2. Salmonella FimH Expression Was Dependent on Growth Conditions and Correlated with Adhesion Level
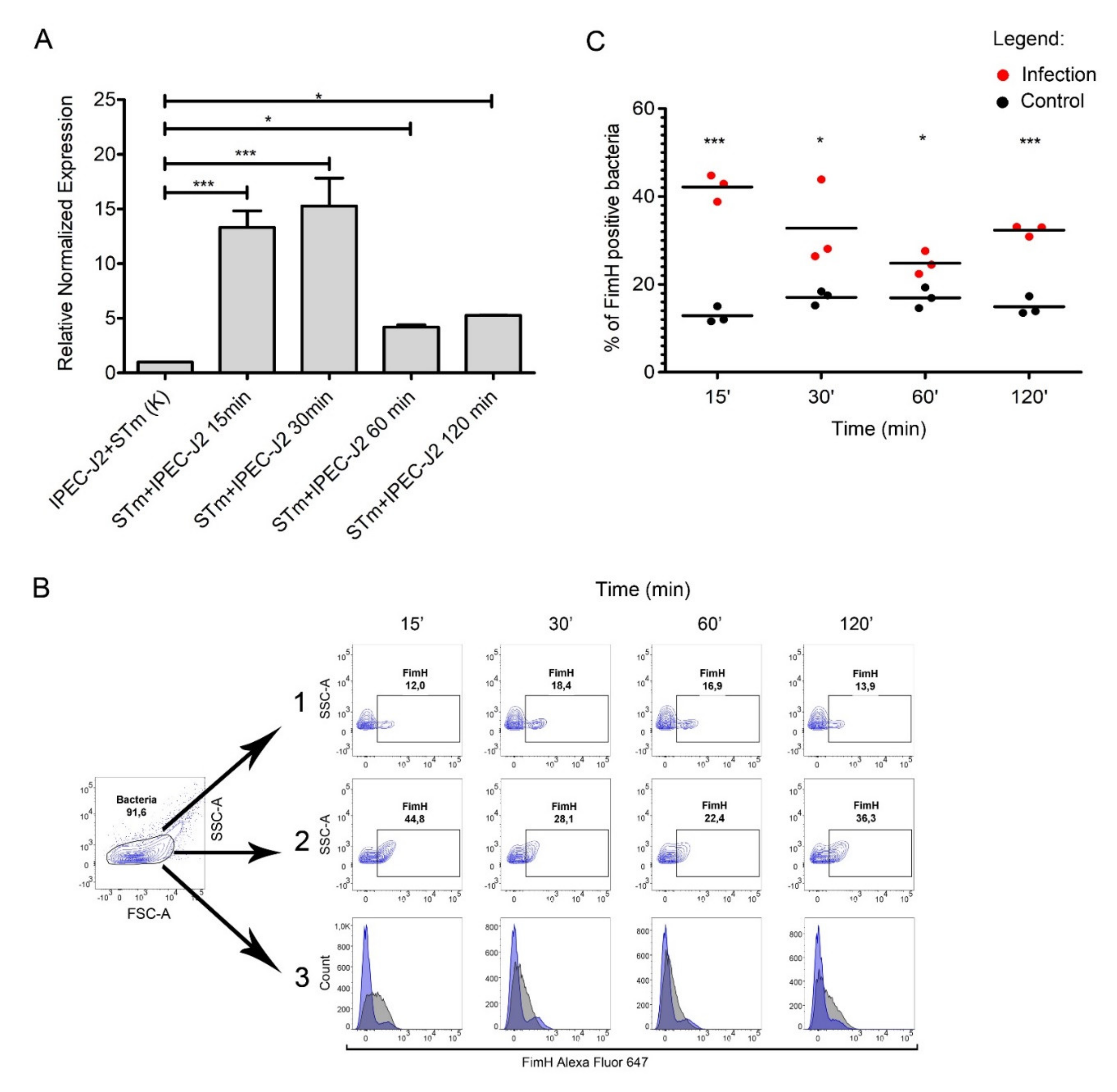
2.3. Salmonella fimH Gene Expression Increased during Contact with IPEC-J2 Cells
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Cells and Cell Culture
4.3. Construction of Strains and Plasmids
4.4. Determination of the Growth Curves
4.5. Adhesion/Infection Assays
4.6. Infection Experiments
4.7. Quantitative PCR (qPCR)
4.8. Flow Cytometry
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| T1F | Type 1 fimbriae |
| T3SS | Type three secretion system |
| SPI-1 | Salmonella pathogenicity island 1 |
| MOI | Multiplicity of infection |
References
- Uzzau, S.; Brown, D.J.; Wallis, T.; Rubino, S.; Leori, G.; Bernard, S.; Casadesus, J.; Platt, D.J.; Olsen, J.E. Host adapted serotypes of Salmonella enterica. Epidemiol. Infect. 2000, 125, 229–255. [Google Scholar] [CrossRef] [PubMed]
- Tsolis, R.M.; Xavier, M.N.; Santos, R.L.; Baumler, A.J. How to become a top model: Impact of animal experimentation on human Salmonella disease research. Infect. Immun. 2011, 79, 1806–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, S.; Slauch, J.M.; Aldridge, P.D.; Rao, C.V. Role of cross talk in regulating the dynamic expression of the flagellar Salmonella pathogenicity island 1 and type 1 fimbrial genes. J. Bacteriol. 2010, 192, 5767–5777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, S.; Pearl, J.A.; Rao, C.V. Role of FimW, FimY, and FimZ in regulating the expression of type i fimbriae in Salmonella enterica serovar Typhimurium. J. Bacteriol. 2009, 191, 3003–3010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolenda, R.; Ugorski, M.; Grzymajlo, K. Everything You Always Wanted to Know About Salmonella Type 1 Fimbriae, but Were Afraid to Ask. Front. Microbiol. 2019, 10, 1017. [Google Scholar] [CrossRef]
- Hahn, E.; Wild, P.; Hermanns, U.; Sebbel, P.; Glockshuber, R.; Haner, M.; Taschner, N.; Burkhard, P.; Aebi, U.; Muller, S.A. Exploring the 3D molecular architecture of Escherichia coli type 1 pili. J. Mol. Biol. 2002, 323, 845–857. [Google Scholar] [CrossRef]
- Grzymajlo, K.; Ugorski, M.; Suchanski, J.; Kedzierska, A.E.; Kolenda, R.; Jarzab, A.; Biernatowska, A.; Schierack, P. The Novel Type 1 Fimbriae FimH Receptor Calreticulin Plays a Role in Salmonella Host Specificity. Front. Cell. Infect. Microbiol. 2017, 7, 326. [Google Scholar] [CrossRef] [PubMed]
- Grzymajlo, K.; Ugorski, M.; Kolenda, R.; Kedzierska, A.; Kuzminska-Bajor, M.; Wieliczko, A. FimH adhesin from host unrestricted Salmonella Enteritidis binds to different glycoprotein ligands expressed by enterocytes from sheep, pig and cattle than FimH adhesins from host restricted Salmonella Abortus-ovis, Salmonella Choleraesuis and Salmonella Dublin. Vet. Microbiol. 2013, 166, 550–557. [Google Scholar] [CrossRef]
- Guo, A.; Cao, S.; Tu, L.; Chen, P.; Zhang, C.; Jia, A.; Yang, W.; Liu, Z.; Chen, H.; Schifferli, D.M. FimH alleles direct preferential binding of Salmonella to distinct mammalian cells or to avian cells. Microbiology 2009, 155, 1623–1633. [Google Scholar] [CrossRef] [Green Version]
- Kisiela, D.I.; Chattopadhyay, S.; Libby, S.J.; Karlinsey, J.E.; Fang, F.C.; Tchesnokova, V.; Kramer, J.J.; Beskhlebnaya, V.; Samadpour, M.; Grzymajlo, K.; et al. Evolution of Salmonella enterica virulence via point mutations in the fimbrial adhesin. PLoS Pathog. 2012, 8, e1002733. [Google Scholar] [CrossRef] [Green Version]
- Purcell, B.K.; Pruckler, J.; Clegg, S. Nucleotide sequences of the genes encoding type 1 fimbrial subunits of Klebsiella pneumoniae and Salmonella typhimurium. J. Bacteriol. 1987, 169, 5831–5834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Old, D.C.; Duguid, J.P. Selective outgrowth of fimbriate bacteria in static liquid medium. J. Bacteriol. 1970, 103, 447–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duguid, J.P.; Anderson, E.S.; Campbell, I. Fimbriae and adhesive properties in Salmonellae. J. Pathol. Bacteriol. 1966, 92, 107–138. [Google Scholar] [CrossRef] [PubMed]
- Silverman, M.; Zieg, J.; Hilmen, M.; Simon, M. Phase variation in Salmonella: Genetic analysis of a recombinational switch. Proc. Natl. Acad. Sci. USA 1979, 76, 391–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosnahan, A.J.; Brown, D.R. Porcine IPEC-J2 intestinal epithelial cells in microbiological investigations. Vet. Microbiol. 2012, 156, 229–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeiner, S.A.; Dwyer, B.E.; Clegg, S. FimA, FimF, and FimH are necessary for assembly of type 1 fimbriae on Salmonella enterica serovar Typhimurium. Infect. Immun. 2012, 80, 3289–3296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weening, E.H.; Barker, J.D.; Laarakker, M.C.; Humphries, A.D.; Tsolis, R.M.; Baumler, A.J. The Salmonella enterica serotype Typhimurium lpf, bcf, stb, stc, std, and sth fimbrial operons are required for intestinal persistence in mice. Infect. Immun. 2005, 73, 3358–3366. [Google Scholar] [CrossRef] [Green Version]
- Dibb-Fuller, M.P.; Allen-Vercoe, E.; Thorns, C.J.; Woodward, M.J. Fimbriae- and flagella-mediated association with and invasion of cultured epithelial cells by Salmonella enteritidis. Microbiology 1999, 145, 1023–1031. [Google Scholar] [CrossRef] [Green Version]
- Collazo, C.M.; Galan, J.E. Requirement for exported proteins in secretion through the invasion-associated type III system of Salmonella typhimurium. Infect. Immun. 1996, 64, 3524–3531. [Google Scholar] [CrossRef] [Green Version]
- Kaniga, K.; Tucker, S.; Trollinger, D.; Galan, J.E. Homologs of the Shigella IpaB and IpaC invasins are required for Salmonella typhimurium entry into cultured epithelial cells. J. Bacteriol. 1995, 177, 3965–3971. [Google Scholar] [CrossRef] [Green Version]
- Kuzminska-Bajor, M.; Grzymajlo, K.; Ugorski, M. Type 1 fimbriae are important factors limiting the dissemination and colonization of mice by Salmonella Enteritidis and contribute to the induction of intestinal inflammation during Salmonella invasion. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Kuzminska-Bajor, M.; Kuczkowski, M.; Grzymajlo, K.; Wojciech, L.; Sabat, M.; Kisiela, D.; Wieliczko, A.; Ugorski, M. Decreased colonization of chicks by Salmonella enterica serovar Gallinarum expressing mannose-sensitive FimH adhesin from Salmonella enterica serovar Enteritidis. Vet. Microbiol. 2012, 158, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Hancox, L.S.; Yeh, K.S.; Clegg, S. Construction and characterization of type 1 non-fimbriate and non-adhesive mutants of Salmonella typhimurium. FEMS Immunol. Med Microbiol. 1997, 19, 289–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolenda, R.; Burdukiewicz, M.; Schiebel, J.; Rodiger, S.; Sauer, L.; Szabo, I.; Orlowska, A.; Weinreich, J.; Nitschke, J.; Bohm, A.; et al. Adhesion of Salmonella to Pancreatic Secretory Granule Membrane Major Glycoprotein GP2 of Human and Porcine Origin Depends on FimH Sequence Variation. Front. Microbiol. 2018, 9, 1905. [Google Scholar] [CrossRef] [PubMed]
- Rajashekara, G.; Munir, S.; Alexeyev, M.F.; Halvorson, D.A.; Wells, C.L.; Nagaraja, K.V. Pathogenic role of SEF14, SEF17, and SEF21 fimbriae in Salmonella enterica serovar enteritidis infection of chickens. Appl. Environ. Microbiol. 2000, 66, 1759–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumler, A.J.; Tsolis, R.M.; Heffron, F. Contribution of fimbrial operons to attachment to and invasion of epithelial cell lines by Salmonella typhimurium. Infect. Immun. 1996, 64, 1862–1865. [Google Scholar] [CrossRef] [Green Version]
- Patterson, S.K.; Borewicz, K.; Johnson, T.; Xu, W.; Isaacson, R.E. Characterization and differential gene expression between two phenotypic phase variants in Salmonella enterica serovar Typhimurium. PLoS ONE 2012, 7, e43592. [Google Scholar] [CrossRef]
- Isaacson, R.E.; Argyilan, C.; Kwan, L.; Patterson, S.; Yoshinaga, K. Phase variable switching of in vivo and environmental phenotypes of Salmonella typhimurium. Adv. Exp. Med. Biol. 1999, 473, 281–289. [Google Scholar] [CrossRef]
- Schierack, P.; Nordhoff, M.; Pollmann, M.; Weyrauch, K.D.; Amasheh, S.; Lodemann, U.; Jores, J.; Tachu, B.; Kleta, S.; Blikslager, A.; et al. Characterization of a porcine intestinal epithelial cell line for in vitro studies of microbial pathogenesis in swine. Histochem. Cell Biol. 2006, 125, 293–305. [Google Scholar] [CrossRef]
- Yue, M.; Han, X.; De Masi, L.; Zhu, C.; Ma, X.; Zhang, J.; Wu, R.; Schmieder, R.; Kaushik, R.S.; Fraser, G.P.; et al. Allelic variation contributes to bacterial host specificity. Nat. Commun. 2015, 6, 8754. [Google Scholar] [CrossRef] [Green Version]
- Wagner, C.; Hensel, M. Adhesive mechanisms of Salmonella enterica. Adv. Exp. Med. Biol. 2011, 715, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, J.A.; Knodler, L.A.; Sturdevant, D.E.; Virtaneva, K.; Carmody, A.B.; Fischer, E.R.; Porcella, S.F.; Steele-Mortimer, O. Induction of Salmonella pathogenicity island 1 under different growth conditions can affect Salmonella-host cell interactions in vitro. Microbiology 2010, 156, 1120–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClelland, M.; Sanderson, K.E.; Spieth, J.; Clifton, S.W.; Latreille, P.; Courtney, L.; Porwollik, S.; Ali, J.; Dante, M.; Du, F.; et al. Complete genome sequence of Salmonella enterica serovar Typhimurium LT2. Nature 2001, 413, 852–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphries, A.D.; Raffatellu, M.; Winter, S.; Weening, E.H.; Kingsley, R.A.; Droleskey, R.; Zhang, S.P.; Figueiredo, J.; Khare, S.; Nunes, J.; et al. The use of flow cytometry to detect expression of subunits encoded by 11 Salmonella enterica serotype Typhimurium fimbrial operons. Mol. Microbiol. 2003, 48, 1357–1376. [Google Scholar] [CrossRef] [PubMed]
- Nuccio, S.P.; Baumler, A.J. Evolution of the chaperone/usher assembly pathway: Fimbrial classification goes Greek. Microbiol. Mol. Biol. Rev. 2007, 71, 551–575. [Google Scholar] [CrossRef] [Green Version]
- Hurtado-Escobar, G.A.; Grepinet, O.; Raymond, P.; Abed, N.; Velge, P.; Virlogeux-Payant, I. H-NS is the major repressor of Salmonella Typhimurium Pef fimbriae expression. Virulence 2019, 10, 849–867. [Google Scholar] [CrossRef]
- Sterzenbach, T.; Nguyen, K.T.; Nuccio, S.P.; Winter, M.G.; Vakulskas, C.A.; Clegg, S.; Romeo, T.; Baumler, A.J. A novel CsrA titration mechanism regulates fimbrial gene expression in Salmonella typhimurium. Embo J. 2013, 32, 2872–2883. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, A.; Lam, L.; Rajendram, M.; Tamburini, F.; Honeycutt, J.; Pham, T.; Van Treuren, W.; Pruss, K.; Stabler, S.R.; Lugo, K.; et al. A Gut Commensal-Produced Metabolite Mediates Colonization Resistance to Salmonella Infection. Cell Host Microbe 2018, 24, 296–307. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Garcia, E.; de Lorenzo, V. Engineering multiple genomic deletions in Gram-negative bacteria: Analysis of the multi-resistant antibiotic profile of Pseudomonas putida KT2440. Environ. Microbiol. 2011, 13, 2702–2716. [Google Scholar] [CrossRef]
- Owen, S.V.; Wenner, N.; Canals, R.; Makumi, A.; Hammarlof, D.L.; Gordon, M.A.; Aertsen, A.; Feasey, N.A.; Hinton, J.C. Characterization of the Prophage Repertoire of African Salmonella Typhimurium ST313 Reveals High Levels of Spontaneous Induction of Novel Phage BTP1. Front. Microbiol. 2017, 8, 235. [Google Scholar] [CrossRef] [Green Version]
- Kisiela, D.; Sapeta, A.; Kuczkowski, M.; Stefaniak, T.; Wieliczko, A.; Ugorski, M. Characterization of FimH adhesins expressed by Salmonella enterica serovar Gallinarum biovars Gallinarum and Pullorum: Reconstitution of mannose-binding properties by single amino acid substitution. Infect. Immun. 2005, 73, 6187–6190. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Strain Tag | Characteristic | Reference |
|---|---|---|---|
| S. Choleraesuis ΔfimH | SCΔfimH | S. Choleraesuis 6150 with fimH gene knockout | [7] |
| S. Typhimurium | WT | SL1344 | Monack lab strain collection [38] |
| S. Typhimurium SL1344_∆fimH | ∆fimH | S. Typhimurium SL 1344 with fimH gene knockout | This study |
| S. Abortusovis | 520 | Wild type, fimbriate | [8] |
| S. Dublin | 508 | Wild type, fimbriate | [8] |
| S. Enteritidis | 327 | Wild type, fimbriate | [8] |
| E. coli DH5α λpir | Monack lab strain collection | ||
| E. coli S17-1 λpir | Monack lab strain collection |
| Plasmid | Characteristic | Reference |
|---|---|---|
| pEMG | Suicide plasmid; KmR | [39] |
| pSW-2 | Plasmid for m-toluate-inducible expression of the I-SceI enzyme; GmR | [39] |
| pEMG | ∆fimH | This study |
| FIMH_F | TACAGCGGCAAAGTGGAAGT |
| FIMH_R | GCCCCCGCCTGACTAAATAA |
| 16S RNA_F | CAGAAGAAGCACCGGCTAAC |
| 16S RNA_R | GCGCTTTACGCCCAGTAATT |
| CG_F | CGGAATTCGCCACGTTTAACGCCAACCGCAACCG |
| CG_R | CGGATCCCGTACCCCCAAAGGCGGGCAC |
| ∆FIMH_B | CATTATGCCTCCCTCTATTTTTCCTTTTATGACGCCGGACG |
| ∆FIMH_D | GGAAAAATAGAGGGAGGCATAATGATCCTTCGGCGCG |
| rpoD_F | ACATGGGTATTCAGGTAATGGAAG |
| rpoD_R | CGGTGCTGGTGGTATTTTCA |
| F_seq | GGCGATTACGATAGCCAGCGC |
| R_seq | CAGCGGGCTGAACAAAACACAAC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klasa, B.; Kędzierska, A.E.; Grzymajło, K. Pre-Growth Culture Conditions Affect Type 1 Fimbriae-Dependent Adhesion of Salmonella. Int. J. Mol. Sci. 2020, 21, 4206. https://doi.org/10.3390/ijms21124206
Klasa B, Kędzierska AE, Grzymajło K. Pre-Growth Culture Conditions Affect Type 1 Fimbriae-Dependent Adhesion of Salmonella. International Journal of Molecular Sciences. 2020; 21(12):4206. https://doi.org/10.3390/ijms21124206
Chicago/Turabian StyleKlasa, Beata, Anna Ewa Kędzierska, and Krzysztof Grzymajło. 2020. "Pre-Growth Culture Conditions Affect Type 1 Fimbriae-Dependent Adhesion of Salmonella" International Journal of Molecular Sciences 21, no. 12: 4206. https://doi.org/10.3390/ijms21124206
APA StyleKlasa, B., Kędzierska, A. E., & Grzymajło, K. (2020). Pre-Growth Culture Conditions Affect Type 1 Fimbriae-Dependent Adhesion of Salmonella. International Journal of Molecular Sciences, 21(12), 4206. https://doi.org/10.3390/ijms21124206