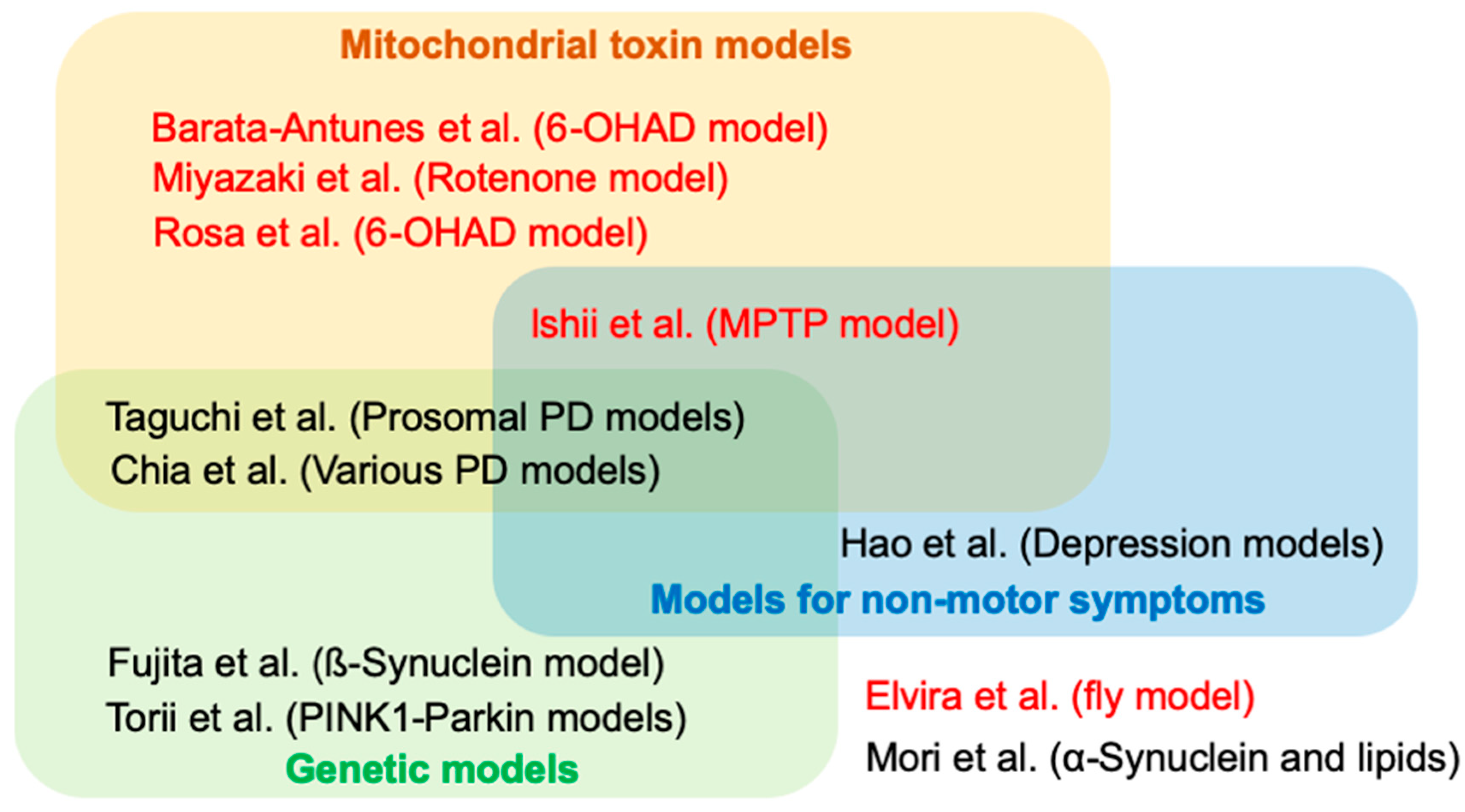
Editorial for the Special Issue “Animal Models of Parkinson’s Disease and Related Disorders”


{kind=link}
Funding
Conflicts of Interest
Abbreviations
| 6-OHDA | 6-Hydroxydopamine |
| DLB | Dementia with Lewy bodies |
| HIF-1 | hypoxia-inducible factor-1 |
| IPAS | Inhibitory PAS domain protein |
| LB | Lewy body |
| MPP+ | 1-Methyl-4-phenylpyridinium |
| MPTP | 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| PD | Parkinson’s disease |
| PERK | Protein kinase RNA-like endoplasmic reticulum kinase |
| ROS | Reactive oxygen species |
References
- Langston, J.W.; Ballard, P.; Tetrud, J.W.; Irwin, I. Chronic Parkinsonism in Humans Due to a Product of Meperidine-Analog Synthesis. Science 1983, 219, 979–980. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, Y.; Saitoh, T.; Sone, N. Inhibition of Mitochondrial NADH-Ubiquinone Oxidoreductase Activity by 1-methyl-4-phenylpyridinium Ion. Biochem. Biophys. Res. Commun. 1987, 143, 294–299. [Google Scholar] [CrossRef]
- Kitayama, S.; Mitsuhata, C.; Davis, S.; Wang, J.B.; Sato, T.; Morita, K.; Uhl, G.R.; Dohi, T. MPP+ Toxicity and Plasma Membrane Dopamine Transporter: Study Using Cell Lines Expressing the Wild-Type and Mutant Rat Dopamine Transporters. Biochim. Biophys. Acta 1998, 1404, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, K.; Tada, Y.; Muroi, Y.; Unno, T.; Ishii, T. Selective Loss of Dopaminergic Neurons in the Substantia Nigra Pars Compacta after Systemic Administration of MPTP Facilitates Extinction Learning. Life Sci. 2015, 137, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Kinoshita, K.I.; Muroi, Y. Serotonin 5-HT4 receptor Agonists Improve Facilitation of Contextual Fear Extinction in an MPTP-Induced Mouse Model of Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 5340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, I.; Di Censo, D.; Ranieri, B.; Di Giovanni, G.; Scarnati, E.; Alecci, M.; Galante, A.; Florio, T.M. Comparison between Tail Suspension Swing Test and Standard Rotation Test in Revealing Early Motor Behavioral Changes and Neurodegeneration in 6-OHDA Hemiparkinsonian rats. Int. J. Mol. Sci. 2020, 21, 2874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barata-Antunes, S.; Teixeira, F.G.; Mendes-Pinheiro, B.; Domingues, A.V.; Vilaca-Faria, H.; Marote, A.; Silva, D.; Sousa, R.A.; Salgado, A.J. Impact of Aging on the 6-OHDA-Induced Rat Model of Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 3459. [Google Scholar] [CrossRef] [PubMed]
- Betarbet, R.; Sherer, T.B.; MacKenzie, G.; Garcia-Osuna, M.; Panov, A.V.; Greenamyre, J.T. Chronic Systemic Pesticide Exposure Reproduces Features of Parkinson’s Disease. Nat. Neurosci. 2000, 3, 1301–1306. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, I.; Isooka, N.; Imafuku, F.; Sun, J.; Kikuoka, R.; Furukawa, C.; Asanuma, M. Chronic Systemic Exposure to Low-Dose Rotenone Induced Central and Peripheral Neuropathology and Motor Deficits in Mice: Reproducible Animal Model of Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 3254. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, T.; Ikuno, M.; Yamakado, H.; Takahashi, R. Animal Model for Prodromal Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Y.; Ge, H.; Sun, M.; Gao, Y. Selecting an Appropriate Animal Model of Depression. Int. J. Mol. Sci. 2019, 20, 4827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, A.; Imai, Y.; Hattori, N. Lipids: Key Players That Modulate alpha-Synuclein Toxicity and Neurodegeneration in Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 3301. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Ho, G.; Takamatsu, Y.; Wada, R.; Ikeda, K.; Hashimoto, M. Possible Role of Amyloidogenic Evolvability in Dementia with Lewy Bodies: Insights from Transgenic Mice Expressing P123H beta-Synuclein. Int. J. Mol. Sci. 2020, 21, 2849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitada, T.; Asakawa, S.; Hattori, N.; Matsumine, H.; Yamamura, Y.; Minoshima, S.; Yokochi, M.; Mizuno, Y.; Shimizu, N. Mutations in the Parkin Gene Cause Autosomal Recessive Juvenile Parkinsonism. Nature 1998, 392, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Valente, E.M.; Abou-Sleiman, P.M.; Caputo, V.; Muqit, M.M.; Harvey, K.; Gispert, S.; Ali, Z.; Del Turco, D.; Bentivoglio, A.R.; Healy, D.G.; et al. Hereditary early-onset Parkinson’s disease caused by mutations in PINK1. Science 2004, 304, 1158–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, Y. PINK1-Parkin signaling in Parkinson’s disease: Lessons from Drosophila. Neurosci. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Torii, S.; Kasai, S.; Yoshida, T.; Yasumoto, K.I.; Shimizu, S. Mitochondrial E3 Ubiquitin Ligase Parkin: Relationships with Other Causal Proteins in Familial Parkinson’s Disease and Its Substrate-Involved Mouse Experimental Models. Int. J. Mol. Sci. 2020, 21, 1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elvira, R.; Cha, S.J.; Noh, G.M.; Kim, K.; Han, J. PERK-Mediated eIF2alpha Phosphorylation Contributes to The Protection of Dopaminergic Neurons from Chronic Heat Stress in Drosophila. Int. J. Mol. Sci. 2020, 21, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chia, S.J.; Tan, E.K.; Chao, Y.X. Historical Perspective: Models of Parkinson’ Disease. Int. J. Mol. Sci. 2020, 21, 2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imai, Y. Editorial for the Special Issue “Animal Models of Parkinson’s Disease and Related Disorders”. Int. J. Mol. Sci. 2020, 21, 4250. https://doi.org/10.3390/ijms21124250
Imai Y. Editorial for the Special Issue “Animal Models of Parkinson’s Disease and Related Disorders”. International Journal of Molecular Sciences. 2020; 21(12):4250. https://doi.org/10.3390/ijms21124250
Chicago/Turabian StyleImai, Yuzuru. 2020. "Editorial for the Special Issue “Animal Models of Parkinson’s Disease and Related Disorders”" International Journal of Molecular Sciences 21, no. 12: 4250. https://doi.org/10.3390/ijms21124250
APA StyleImai, Y. (2020). Editorial for the Special Issue “Animal Models of Parkinson’s Disease and Related Disorders”. International Journal of Molecular Sciences, 21(12), 4250. https://doi.org/10.3390/ijms21124250