Artificial Sporulation Induction (ASI) by kinA Overexpression Affects the Proteomes and Properties of Bacillus subtilis Spores

 ,
,  and
and
Abstract
:1. Introduction
2. Results
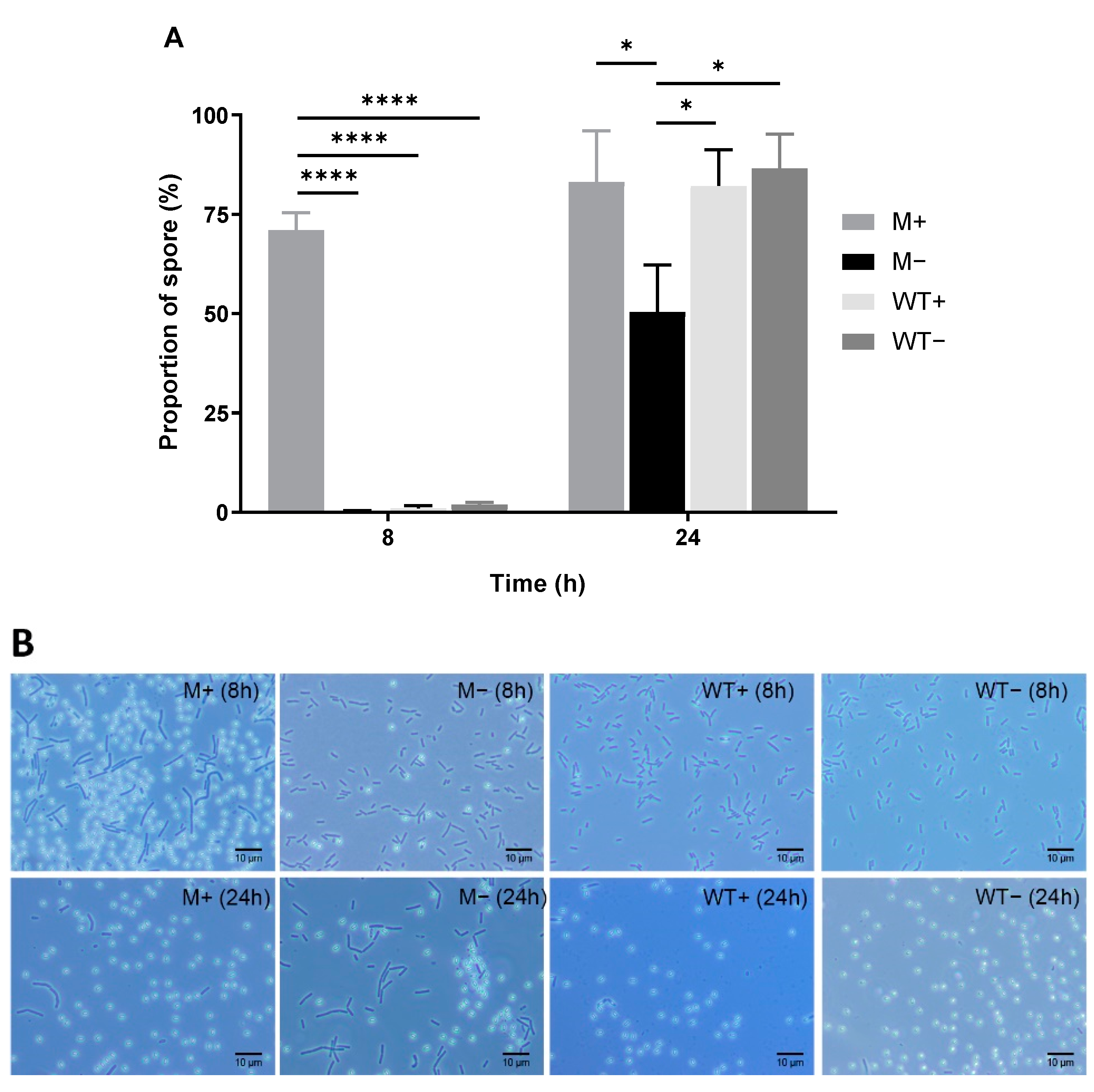
2.1. Overexpression of kinA Minimizes Heterogeneity in Sporulation Initiation
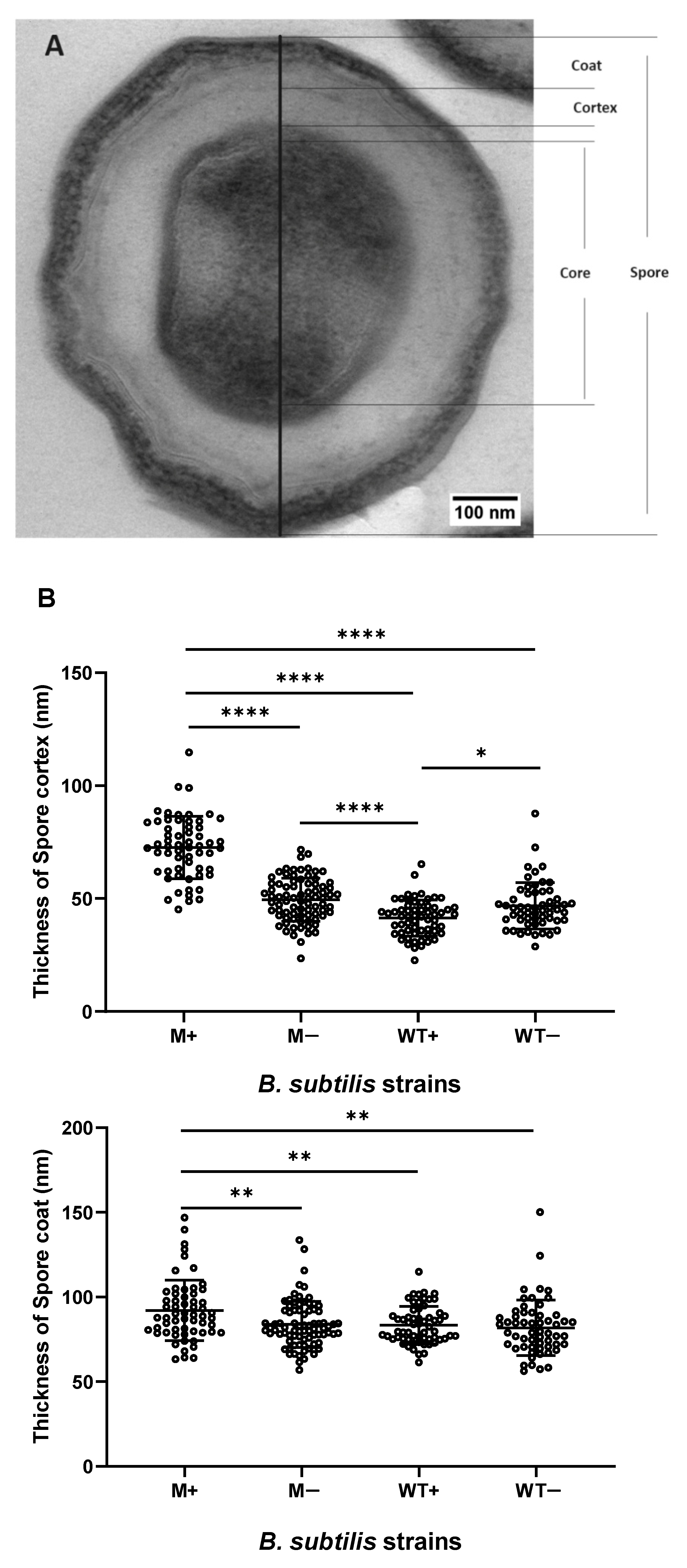
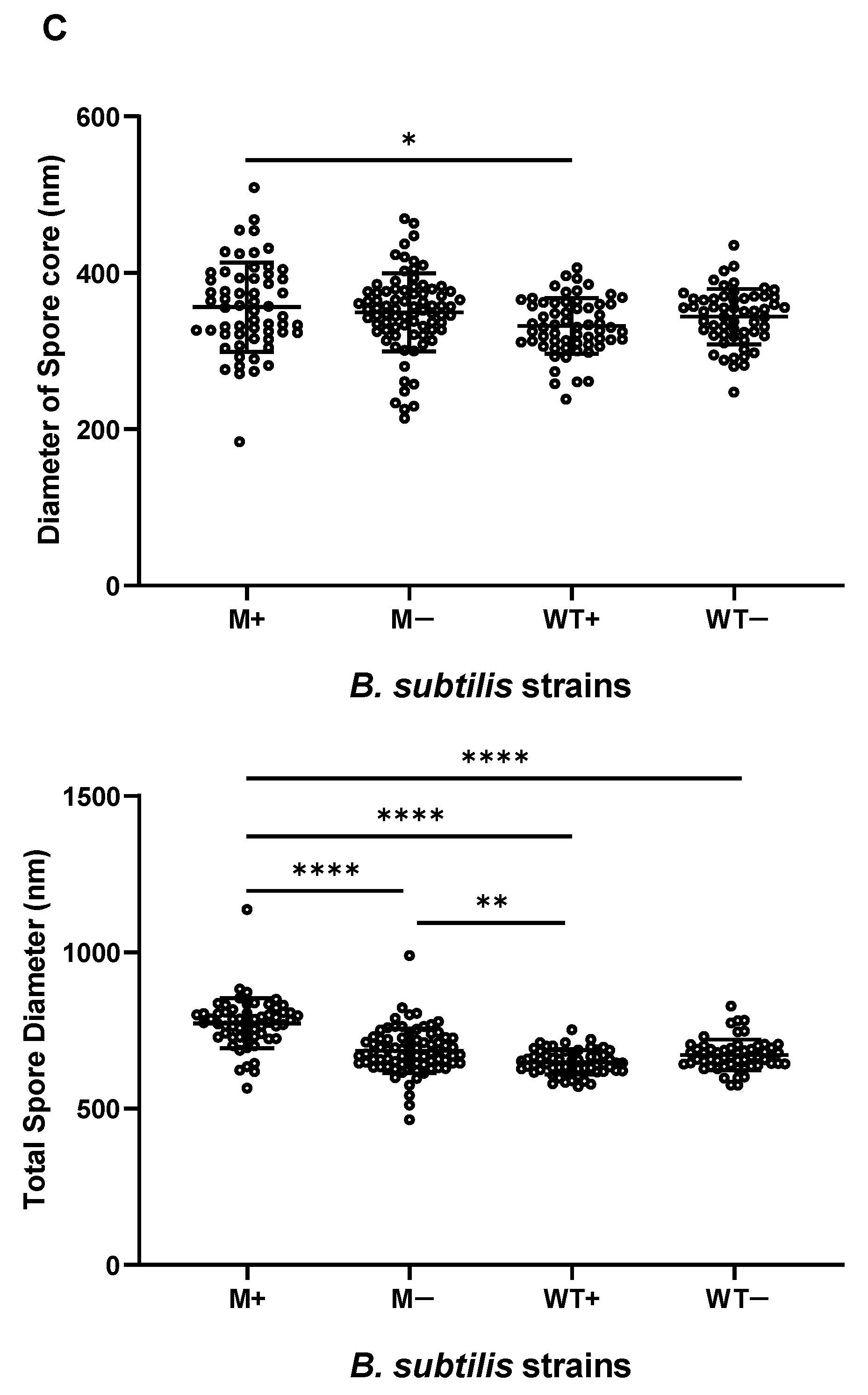
2.2. Synchronized Sporulation Yields Spores with a Thicker Cortex, Coat Layers and a Larger Spore Diameter
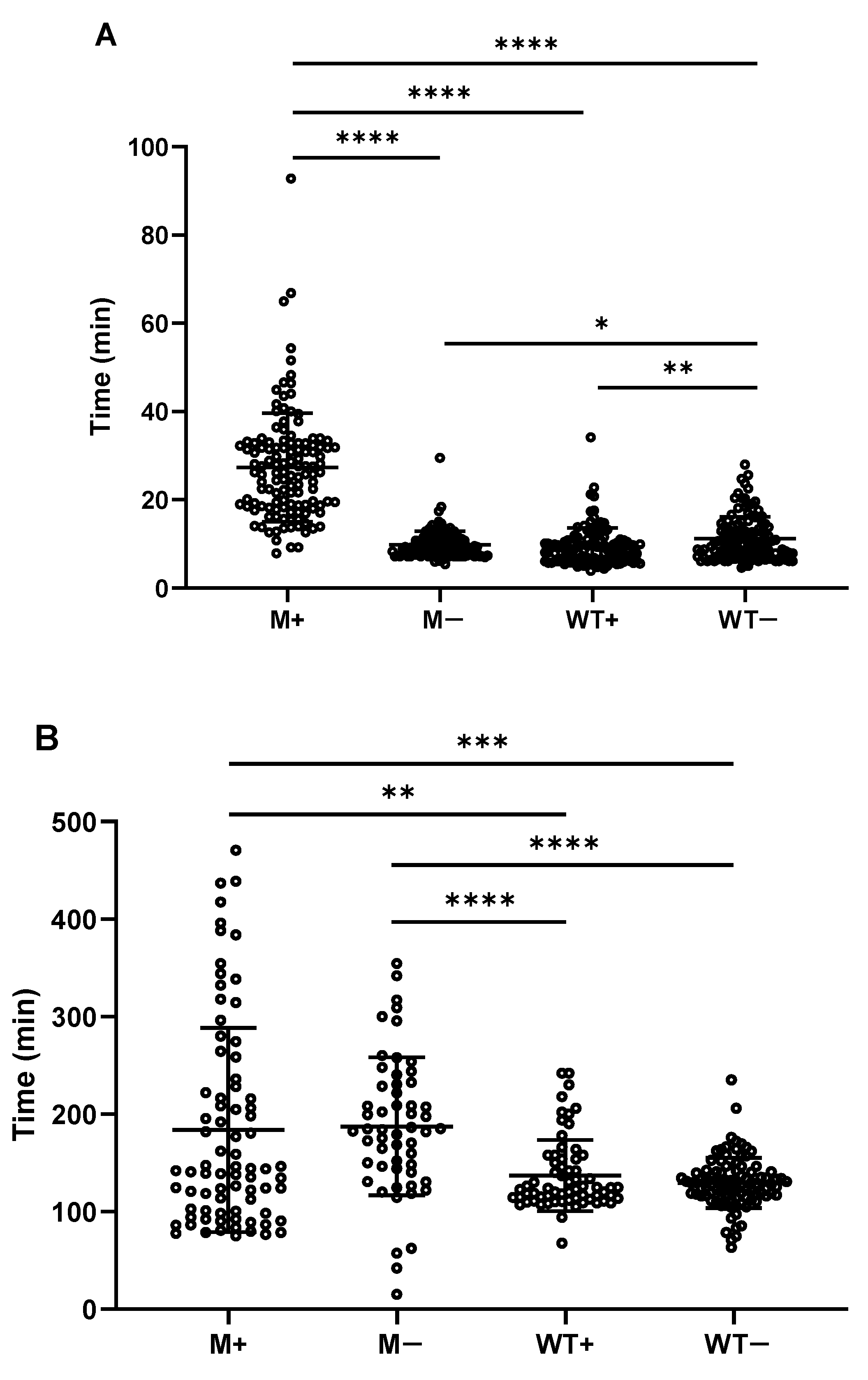
2.3. Spores upon Synchronously Initiating Sporulation Acquire Increased Wet Heat Resistance and Are Delayed in Start of Germination and Burst
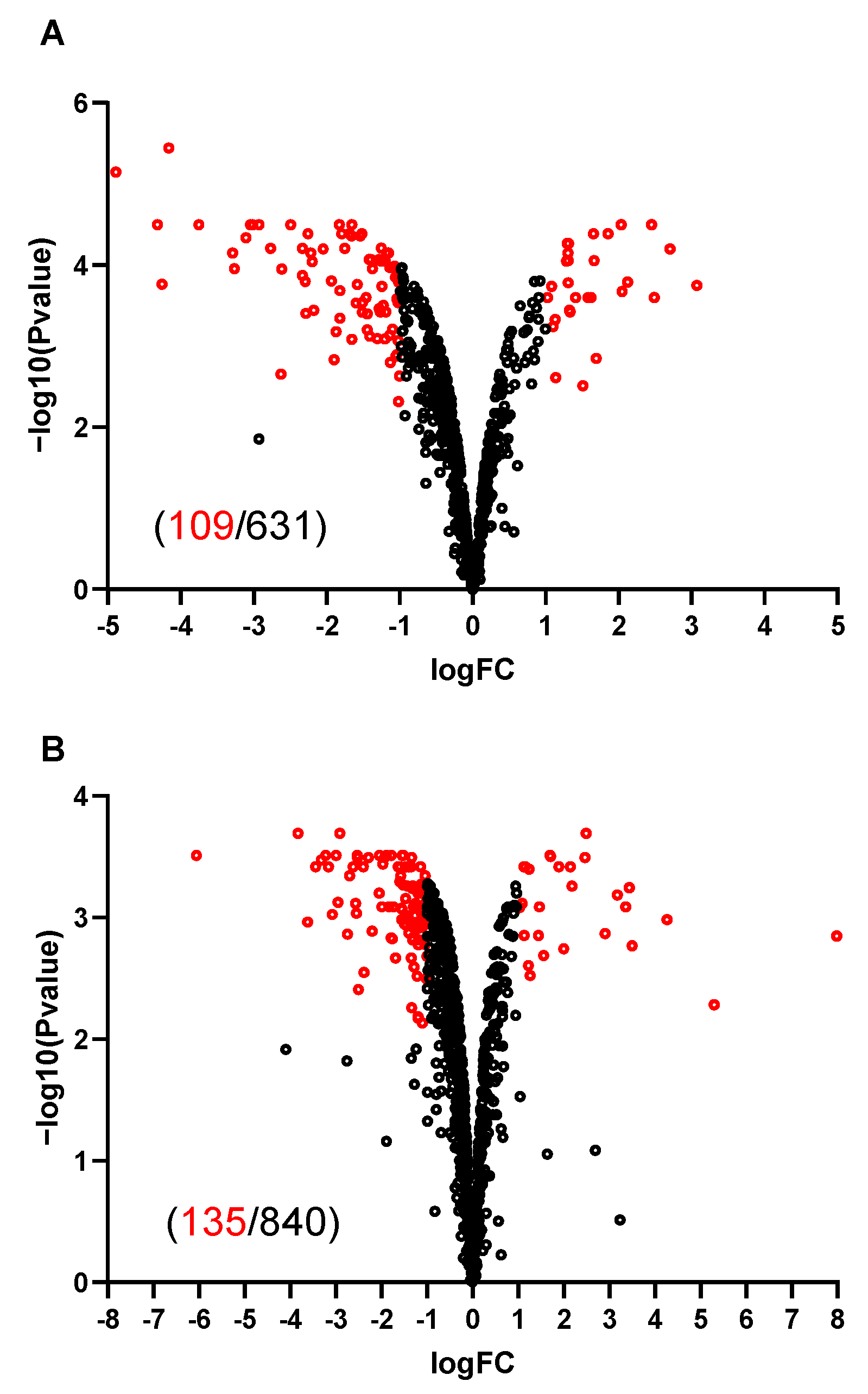
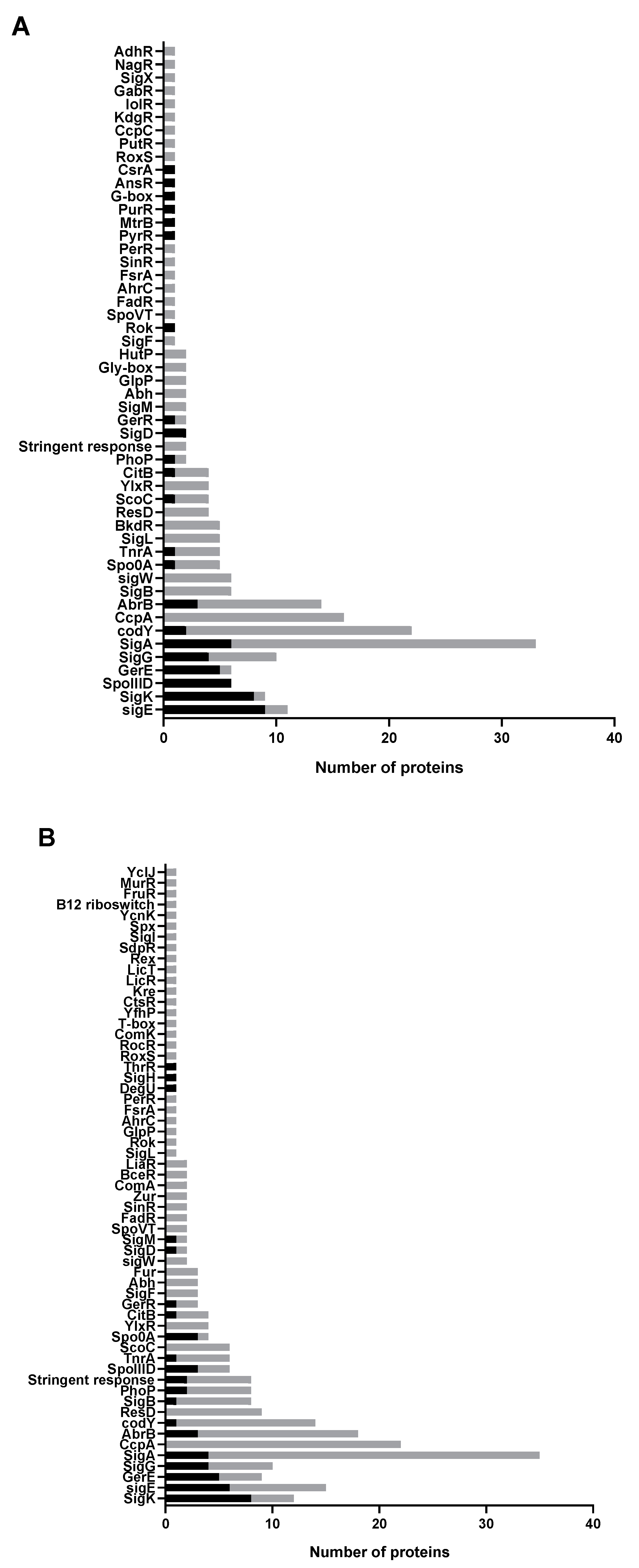
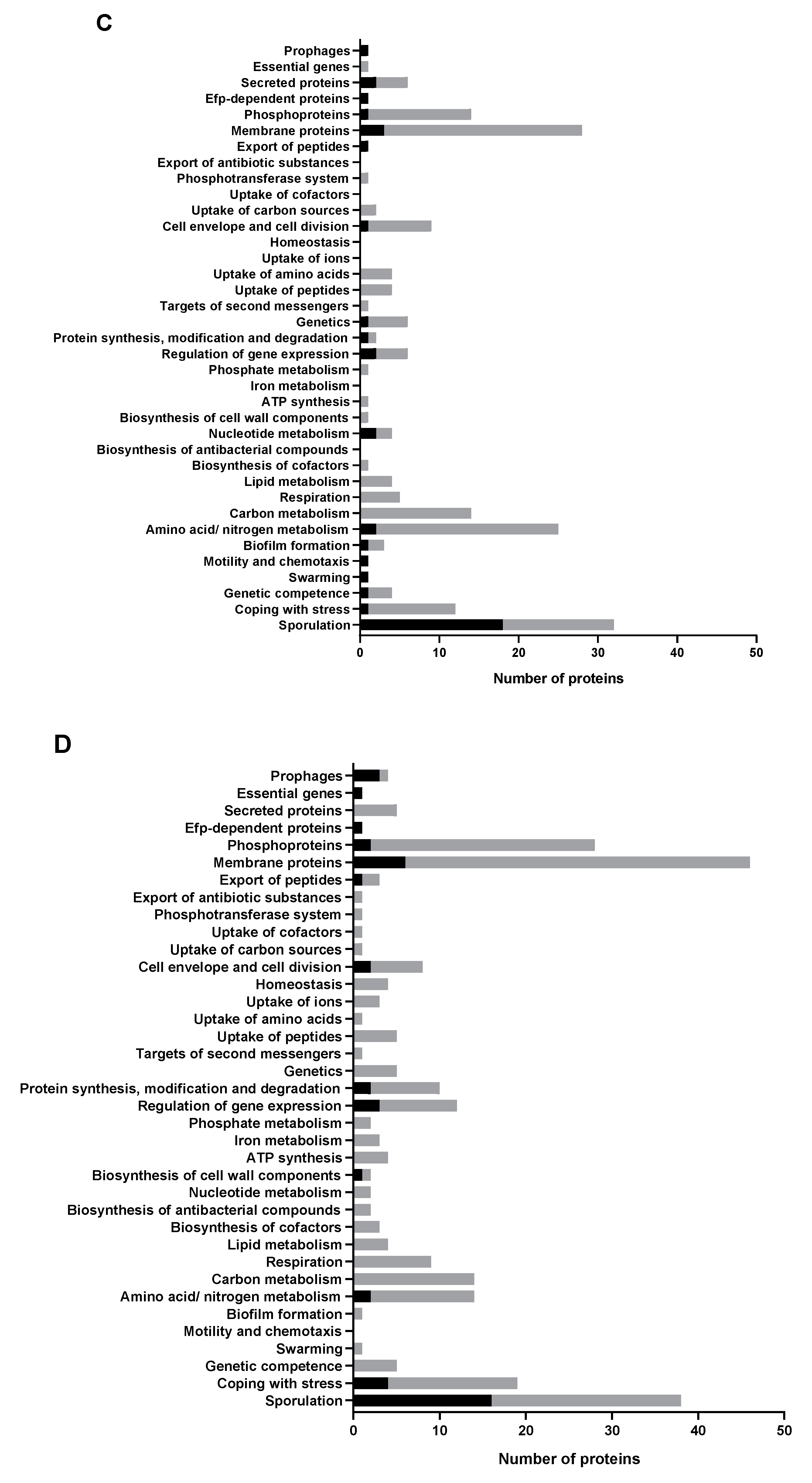
2.4. Most Upregulated Proteins and a Subgroup of Downregulated Proteins in M+ Spores Are Related to Sporulation
2.5. Abolishing KinA Synthesis Has Profound Influence on the Proteome of Vegetative Cells and Spores
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and Sporulation
4.2. Sporulation Efficiency and Sample Harvesting
4.3. Electron Microscopy
4.4. Live Imaging of Spores
4.5. Heat Resistance of Spores
4.6. Sample Preparation for LCMS Analysis
4.7. LCMS Analysis
4.8. Data Processing and Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Phillips, Z.E.V.; Strauch, M.A. Bacillus subtilis sporulation and stationary phase gene expression. Cell. Mol. Life Sci. 2002, 59, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.; Dworkin, J. Recent progress in Bacillus subtilis sporulation. FEMS Microbiol. Rev. 2012, 36, 131–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatt, J.K.; Youngman, P. Spo0A Mutants of Bacillus subtilis with Sigma Factor-Specific Defects in Transcription Activation. J. Bacteriol. 1998, 180, 3584–3591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.-J.; Piggot, P.J.; Tatti, K.M.; Moran, C.P., Jr. Transcription of the Bacillus subtilis spoIIA locus. Gene 1991, 101, 113–116. [Google Scholar] [CrossRef]
- Baldus, J.M.; Green, B.D.; Youngman, P.; Moran, C.P. Phosphorylation of Bacillus subtilis transcription factor Spo0A stimulates transcription from the spoIIG promoter by enhancing binding to weak 0A boxes. J. Bacteriol. 1994, 176, 296–306. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Piggot, P.J. Development of a two-part transcription probe to determine the completeness of temporal and spatial compartmentalization of gene expression during bacterial development. Proc. Natl. Acad. Sci. USA 2001, 98, 12538–12543. [Google Scholar] [CrossRef] [Green Version]
- Riley, E.P.; Trinquier, A.; Reilly, M.L.; Durchon, M.; Perera, V.R.; Pogliano, K.; Lopez-Garrido, J. Spatiotemporally regulated proteolysis to dissect the role of vegetative proteins during Bacillus subtilis sporulation: Cell-specific requirement of σH and σA. Mol. Microbiol. 2018, 108, 45–62. [Google Scholar] [CrossRef] [Green Version]
- Burbulys, D.; Trach, K.A.; Hoch, J.A. Initiation of sporulation in B. subtilis is controlled by a multicomponent phosphorelay. Cell 1991, 64, 545–552. [Google Scholar]
- Fujita, M.; Losick, R. Evidence that entry into sporulation in Bacillus subtilis is governed by a gradual increase in the level and activity of the master regulator Spo0A. Genes Dev. 2005, 19, 2236–2244. [Google Scholar] [CrossRef] [Green Version]
- de Jong, I.G.; Veening, J.-W.; Kuipers, O.P. Heterochronic phosphorelay gene expression as a source of heterogeneity in Bacillus subtilis spore formation. J. Bacteriol. 2010, 192, 2053–2067. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; He, Z.; Gao, F.; Yan, J.; Huang, X. Sensor kinase KinB and its pathway-associated key factors sense the signal of nutrition starvation in sporulation of Bacillus subtilis. MicrobiologyOpen 2018, 7, e00566. [Google Scholar] [CrossRef] [Green Version]
- Russell, J.R.; Cabeen, M.T.; Wiggins, P.A.; Paulsson, J.; Losick, R. Noise in a phosphorelay drives stochastic entry into sporulation in Bacillus subtilis. EMBO J. 2017, 36, 2856–2869. [Google Scholar] [CrossRef]
- Narula, J.; Kuchina, A.; Dong-yeon, D.L.; Fujita, M.; Süel, G.M.; Igoshin, O.A. Chromosomal arrangement of phosphorelay genes couples sporulation and DNA replication. Cell 2015, 162, 328–337. [Google Scholar] [CrossRef] [Green Version]
- Narula, J.; Kuchina, A.; Zhang, F.; Fujita, M.; Süel, G.M.; Igoshin, O.A. Slowdown of growth controls cellular differentiation. Mol. Syst. Biol. 2016, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- González-Pastor, J.E. Cannibalism: A social behavior in sporulating Bacillus subtilis. FEMS Microbiol. Rev. 2011, 35, 415–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Mathys, A. Superdormant spores as a hurdle for gentle germination-inactivation based spore control strategies. Front. Microbiol. 2019, 9, 3163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narula, J.; Devi, S.N.; Fujita, M.; Igoshin, O.A. Ultrasensitivity of the Bacillus subtilis sporulation decision. Proc. Natl. Acad. Sci. USA 2012, 109, E3513–E3522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiehler, B.; Haggett, L.; Fujita, M. The PAS domains of the major sporulation kinase in Bacillus subtilis play a role in tetramer formation that is essential for the autokinase activity. MicrobiologyOpen 2017, 6, e00481. [Google Scholar] [CrossRef] [Green Version]
- Chirakkal, H.; O’Rourke, M.; Atrih, A.; Foster, S.J.; Moir, A. Analysis of spore cortex lytic enzymes and related proteins in Bacillus subtilis endospore germination. Microbiology 2002, 148, 2383–2392. [Google Scholar] [CrossRef] [Green Version]
- El Mammeri, N.; Hierrezuelo, J.; Tolchard, J.; Cámara-Almirón, J.; Caro-Astorga, J.; Álvarez-Mena, A.; Dutour, A.; Berbon, M.; Shenoy, J.; Morvan, E. Molecular architecture of bacterial amyloids in Bacillus biofilms. FASEB J. 2019, 33, 12146–12163. [Google Scholar] [CrossRef] [Green Version]
- Shafikhani, S.H.; Núñez, E.; Leighton, T. ScoC mediates catabolite repression of sporulation in Bacillus subtilis. Curr. Microbiol. 2003, 47, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ray, W.K.; Helm, R.F.; Melville, S.B.; Popham, D.L. Levels of germination proteins in Bacillus subtilis dormant, superdormant, and germinating spores. PLoS ONE 2014, 9, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; Scotland, M.; Setlow, P. Levels of germination proteins in dormant and superdormant spores of Bacillus subtilis. J. Bacteriol. 2012, 194, 2221–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, I.S.; Ramamurthi, K.S. Spore formation in Bacillus subtilis. Environ. Microbiol. Rep. 2014, 6, 212–225. [Google Scholar] [CrossRef] [Green Version]
- Swarge, B.; Roseboom, W.; de Koster, C.; Brul, S.; de Koning, L. Towards the minimal proteome of life: Proteome profiles of Bacillus subtilis vegetative cells and spores. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Eswaramoorthy, P.; Duan, D.; Dinh, J.; Dravis, A.; Devi, S.N.; Fujita, M. The threshold level of the sensor histidine kinase KinA governs entry into sporulation in Bacillus subtilis. J. Bacteriol. 2010, 192, 3870–3882. [Google Scholar] [CrossRef] [Green Version]
- Kort, R.; O’brien, A.C.; Van Stokkum, I.H.; Oomes, S.J.; Crielaard, W.; Hellingwerf, K.J.; Brul, S. Assessment of heat resistance of bacterial spores from food product isolates by fluorescence monitoring of dipicolinic acid release. Appl. Environ. Microbiol. 2005, 71, 3556–3564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eswaramoorthy, P.; Guo, T.; Fujita, M. In vivo domain-based functional analysis of the major sporulation sensor kinase, KinA, in Bacillus subtilis. J. Bacteriol. 2009, 191, 5358–5368. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Korza, G.; Maciejewski, M.; Setlow, P. Analysis of metabolism in dormant spores of Bacillus species by 31P nuclear magnetic resonance analysis of low-molecular-weight compounds. J. Bacteriol. 2015, 197, 992–1001. [Google Scholar] [CrossRef] [Green Version]
- Pandey, R.; Ter Beek, A.; Vischer, N.O.; Smelt, J.P.; Brul, S.; Manders, E.M. Live cell imaging of germination and outgrowth of individual Bacillus subtilis spores; the effect of heat stress quantitatively analyzed with SporeTracker. PLoS ONE 2013, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Omardien, S.; Drijfhout, J.W.; Zaat, S.A.; Brul, S. Cationic amphipathic antimicrobial peptides perturb the inner membrane of germinated spores thus inhibiting their outgrowth. Front. Microbiol. 2018, 9, 2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omardien, S.; Ter Beek, A.; Vischer, N.; Montijn, R.; Schuren, F.; Brul, S. Evaluating novel synthetic compounds active against Bacillus subtilis and Bacillus cereus spores using Live imaging with SporeTrackerX. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Abhyankar, W.R.; Kamphorst, K.; Swarge, B.N.; van Veen, H.; van der Wel, N.N.; Brul, S.; de Koster, C.G.; de Koning, L.J. The influence of sporulation conditions on the spore coat protein composition of Bacillus subtilis spores. Front. Microbiol. 2016, 7, 1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abhyankar, W.; Stelder, S.; de Koning, L.; de Koster, C.; Brul, S. ‘Omics’ for microbial food stability: Proteomics for the development of predictive models for bacterial spore stress survival and outgrowth. Int. J. Food Microbiol. 2017, 240, 11–18. [Google Scholar] [CrossRef]
- Swarge, B.N.; Roseboom, W.; Zheng, L.; Abhyankar, W.R.; Brul, S.; de Koster, C.G.; de Koning, L.J. “ One-Pot” Sample Processing Method for Proteome-Wide Analysis of Microbial Cells and Spores. Proteom.-Clin. Appl. 2018, 12, 1700169. [Google Scholar] [CrossRef] [Green Version]
- Scopes, R.K. Measurement of protein by spectrophotometry at 205 nm. Anal. Biochem. 1974, 59, 277–282. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.I.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Zhu, B.; Stülke, J. Subti Wiki in 2018: From genes and proteins to functional network annotation of the model organism Bacillus subtilis. Nucleic Acids Res. 2018, 46, D743–D748. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Log2(M+/M−) | Log2(M+/WT+) | Function | Product |
|---|---|---|---|---|
| Upregulation | ||||
| CotQ | 1.66 | 1.70 | protection of the spore | spore coat protein |
| CotU | 2.49 | 2.00 | resistance of the spore | outer spore coat protein |
| GerE | 2.70 | 2.90 | regulation of SigK-dependent gene expression | transcriptional regulator (LuxR-FixJ family) |
| YheC | 3.07 | 3.17 | Unknown | ATP-binding spore coat protein |
| YhxC | 2.03 | 1.24 | Unknown | unknown |
| YjqC | 1.66 | 2.49 | protection of the spore | spore coat protein |
| YpqA | 1.31 | 2.14 | Unknown | unknown |
| YraF | 1.41 | 1.09 | Unknown | unknown |
| YtcC | 2.05 | 2.46 | lipopolysaccharide biosynthesis | sporulation protein |
| YxeD | 1.29 | 1.47 | Unknown | unknown |
| CotC | 1.51 | N.A. | resistance of the spore | spore coat protein (outer) |
| CotJA | 1.31 | N.A. | polypeptide composition of the spore coat | unknown |
| CotJB | 1.30 | N.A. | polypeptide composition of the spore coat | unknown |
| CotJC | 1.32 | N.A. | polypeptide composition of the spore coat, may protect against oxidative stress | putative manganese catalase |
| GerT | 1.86 | N.A. | germination | spore coat protein |
| YdhD | 1.31 | N.A. | Unknown | spore coat glycosylase |
| YqfT | 1.03 | N.A. | Unknown | unknown |
| YraG | 1.33 | N.A. | Unknown | unknown |
| KinA | 4.11 a | 5.29 | initiation of sporulation | two-component sensor kinase |
| MurG | N.A. | 1.13 | peptidoglycan precursor biosynthesis | UDP-N-acetylglucosamine-N-acetylmuramyl-(pentapeptide)pyrophosphoryl-undecaprenol N-acetylglucosamine transferase |
| SpsB | N.A. | 1.71 | spore coat polysaccharide synthesis | unknown |
| YabG | N.A. | 1.03 | modification of spore coat proteins | protease |
| YojB | N.A. | 1.26 | Unknown | unknown |
| YraD | N.A. | 1.15 | Unknown | unknown |
| Downregulation | ||||
| OppA | −4.16 | −2.53 | initiation of sporulation, competence development | oligopeptide ABC transporter (binding protein) |
| OppD | −2.93 | −1.42 | initiation of sporulation, competence development | oligopeptide ABC transporter (ATP-binding protein) |
| OppF | −3.04 | −1.84 | initiation of sporulation, competence development | oligopeptide ABC transporter (ATP-binding protein) |
| PdaA | −1.01 | −1.12 | spore cortex peptidoglycan synthesis | N-acetylmuramic acid deacetylase |
| SspA | −1.18 | −1.59 | protection of spore DNA | small acid-soluble spore protein (major alpha-type SASP) |
| YckD | −1.14 | −1.43 | Unknown | unknown |
| YugP | −1.02 | −1.20 | Unknown | unknown |
| CgeA | −3.27 | N.A. | maturation of the outermost layer of the spore | spore crust protein |
| GlnH | −2.49 | N.A. | glutamine uptake | glutamine ABC transporter (binding protein) |
| ParA | −1.01 | N.A. | forespore chromosome partitioning/negative regulation of Sporulation initiation | negative regulator of Sporulation initiation |
| PbpF | −1.10 | N.A. | bifunctional glucosyltransferase/transpeptidase | penicillin-binding protein 2C |
| YbfJ | −1.14 | N.A. | Unknown | unknown |
| YhfN | −1.02 | N.A. | Unknown | unknown |
| YuaG | −2.22 | N.A. | involved in the control of membrane fluidity | membrane-associated scaffold protein |
| BdbD | N.A. | −1.40 | oxidative folding of proteins | thiol-disulfide oxidoreductase |
| CotG | N.A. | −1.10 | resistance of the spore | spore coat protein |
| CotW | N.A. | −1.34 | resistance of the spore | spore crust protein (insoluble fraction) |
| CotX | N.A. | −1.31 | spore crust assembly | spore crust protein (insoluble fraction) |
| DacB | N.A. | −1.06 | carboxypeptidase | penicillin-binding protein 5*, D-alanyl-D-alanine carboxypeptidase |
| GerBC | N.A. | −1.28 | germination | nutrient receptor |
| OppC | N.A. | −1.52 | initiation of sporulation, competence development | oligopeptide ABC transporter (permease) |
| PhoP | N.A. | −1.55 | regulation of phosphate metabolism (phoA, phoB, phoD, resABCDE, tagA-tagB, tagDEF, [tuaA-H]) | two-component response regulator (OmpR family) |
| PhoR | N.A. | −1.93 | regulation of phosphate metabolism | two-component sensor kinase |
| spoVD | N.A. | −1.30 | spore morphogenesis | penicillin-binding protein (spore cortex) |
| SspE | N.A. | −1.09 | Unknown | small acid-soluble spore protein (major gamma-type SASP) |
| SspG | N.A. | −1.01 | protection of spore DNA | small acid-soluble spore protein (minor) |
| YbbC | N.A. | −1.32 | Unknown | unknown |
| YdcC | N.A. | −1.30 | Unknown | unknown |
| YjaZ | N.A. | −1.32 | Unknown | unknown |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, Z.; R. Abhyankar, W.; N. Swarge, B.; van der Wel, N.; Kramer, G.; Brul, S.; J. de Koning, L. Artificial Sporulation Induction (ASI) by kinA Overexpression Affects the Proteomes and Properties of Bacillus subtilis Spores. Int. J. Mol. Sci. 2020, 21, 4315. https://doi.org/10.3390/ijms21124315
Tu Z, R. Abhyankar W, N. Swarge B, van der Wel N, Kramer G, Brul S, J. de Koning L. Artificial Sporulation Induction (ASI) by kinA Overexpression Affects the Proteomes and Properties of Bacillus subtilis Spores. International Journal of Molecular Sciences. 2020; 21(12):4315. https://doi.org/10.3390/ijms21124315
Chicago/Turabian StyleTu, Zhiwei, Wishwas R. Abhyankar, Bhagyashree N. Swarge, Nicole van der Wel, Gertjan Kramer, Stanley Brul, and Leo J. de Koning. 2020. "Artificial Sporulation Induction (ASI) by kinA Overexpression Affects the Proteomes and Properties of Bacillus subtilis Spores" International Journal of Molecular Sciences 21, no. 12: 4315. https://doi.org/10.3390/ijms21124315
APA StyleTu, Z., R. Abhyankar, W., N. Swarge, B., van der Wel, N., Kramer, G., Brul, S., & J. de Koning, L. (2020). Artificial Sporulation Induction (ASI) by kinA Overexpression Affects the Proteomes and Properties of Bacillus subtilis Spores. International Journal of Molecular Sciences, 21(12), 4315. https://doi.org/10.3390/ijms21124315