Purified Tea (Camellia sinensis (L.) Kuntze) Flower Saponins Induce the p53-Dependent Intrinsic Apoptosis of Cisplatin-Resistant Ovarian Cancer Cells





Abstract
:1. Introduction
2. Results
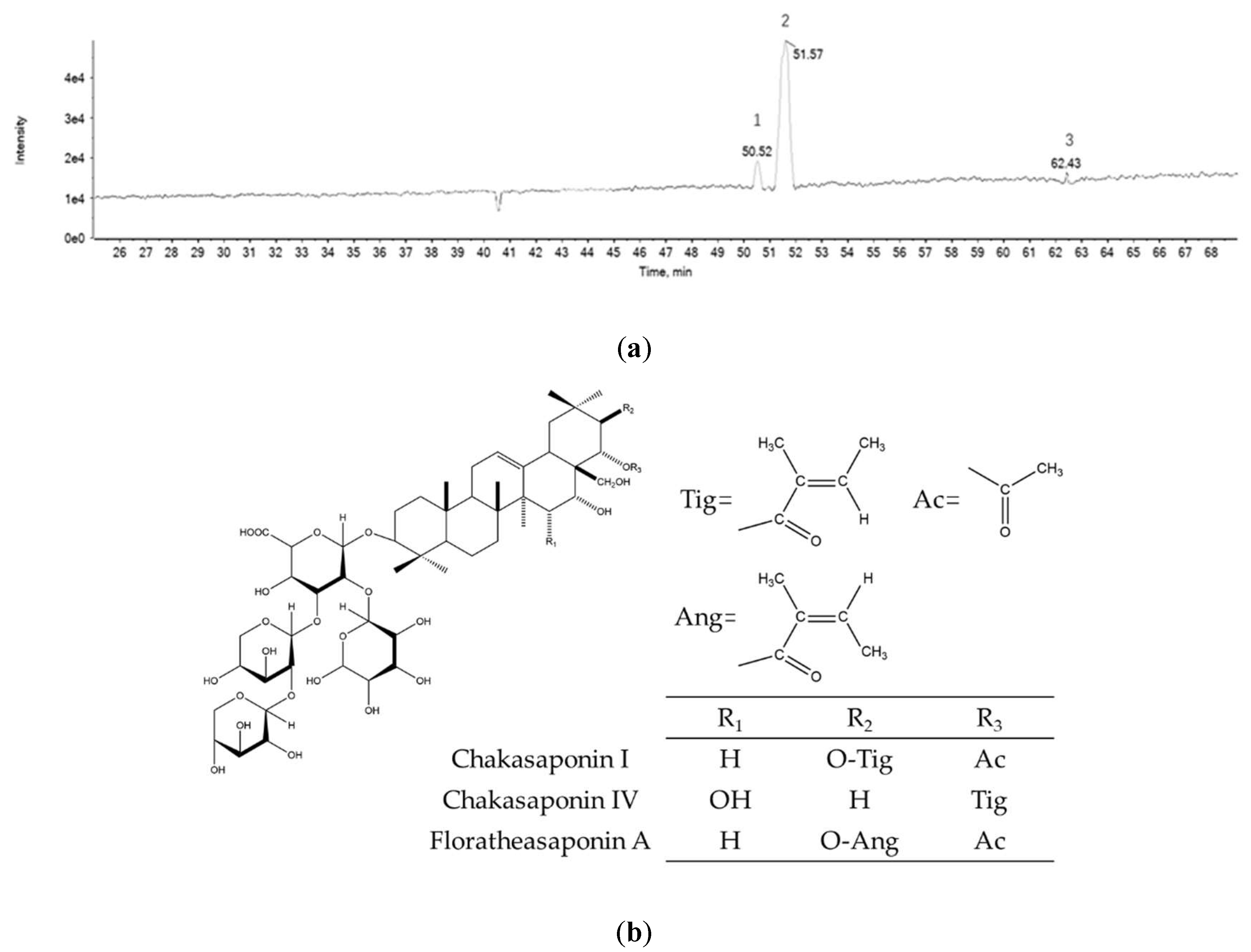
2.1. Chemical Composition of PTFSs
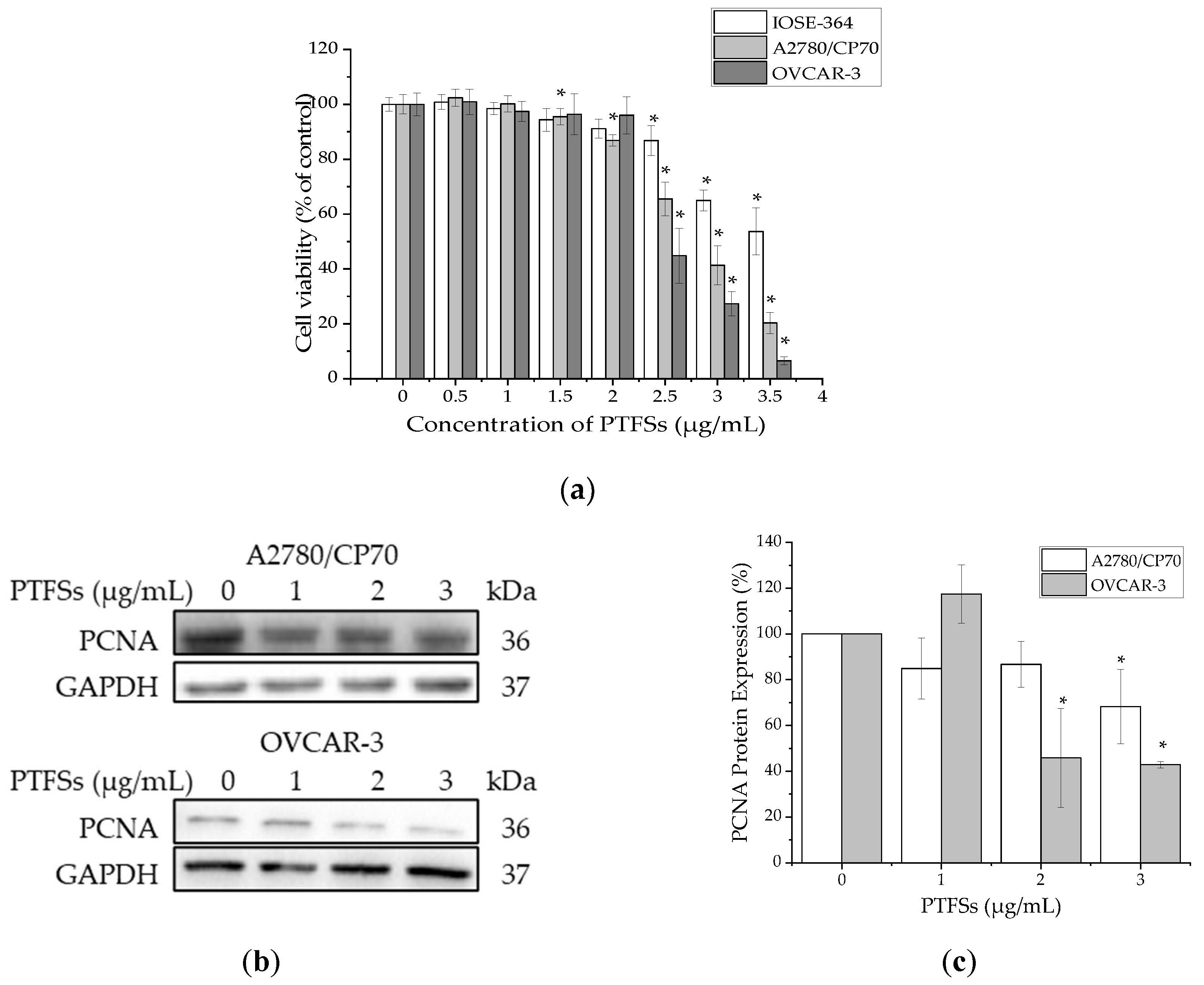
2.2. PTFSs Inhibit Cell Viability and Cell Proliferation in A2780/CP70s and OVCAR-3s
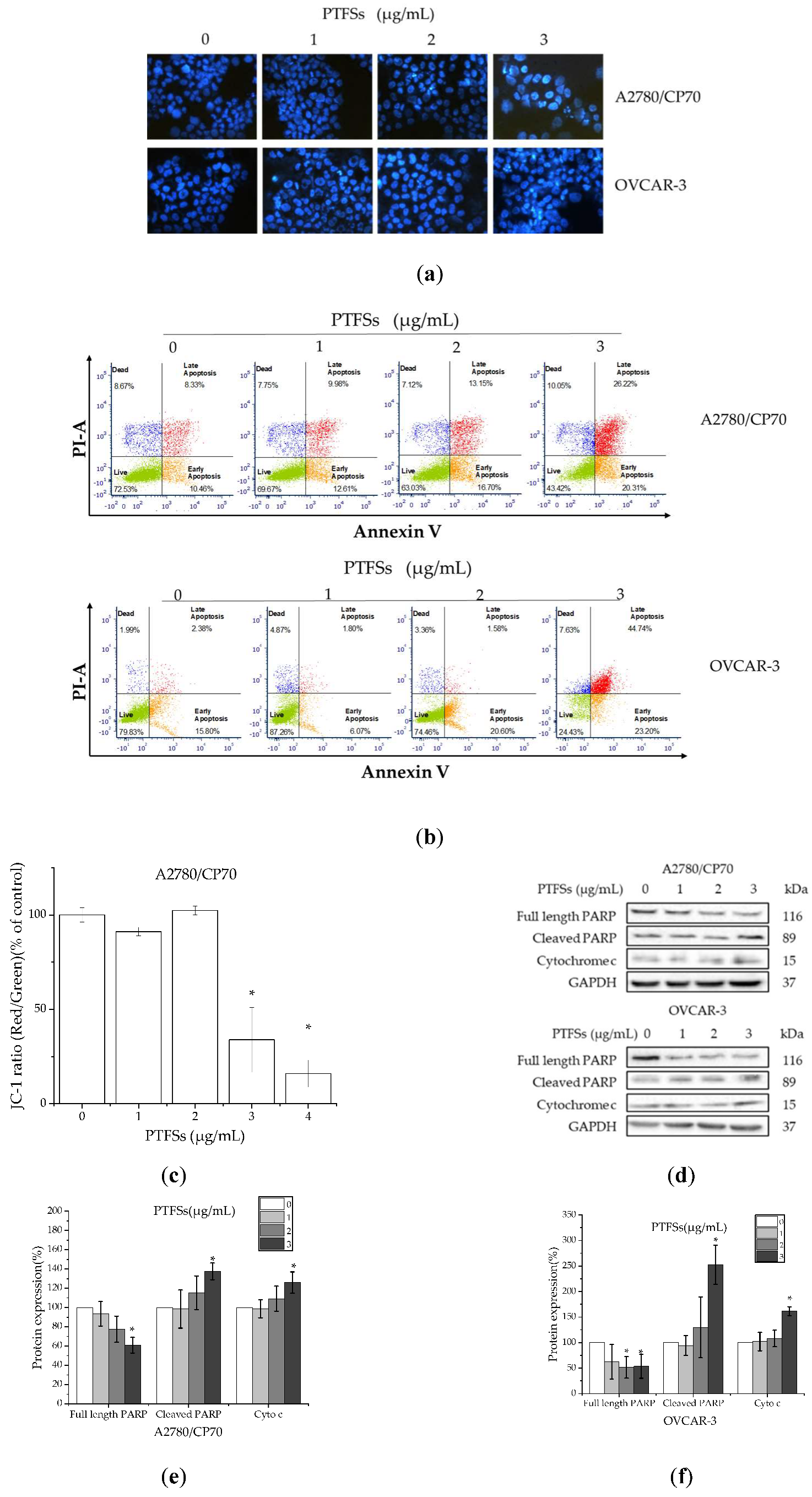
2.3. PTFSs Induce Apoptosis in A2780/CP70s and OVCAR-3s
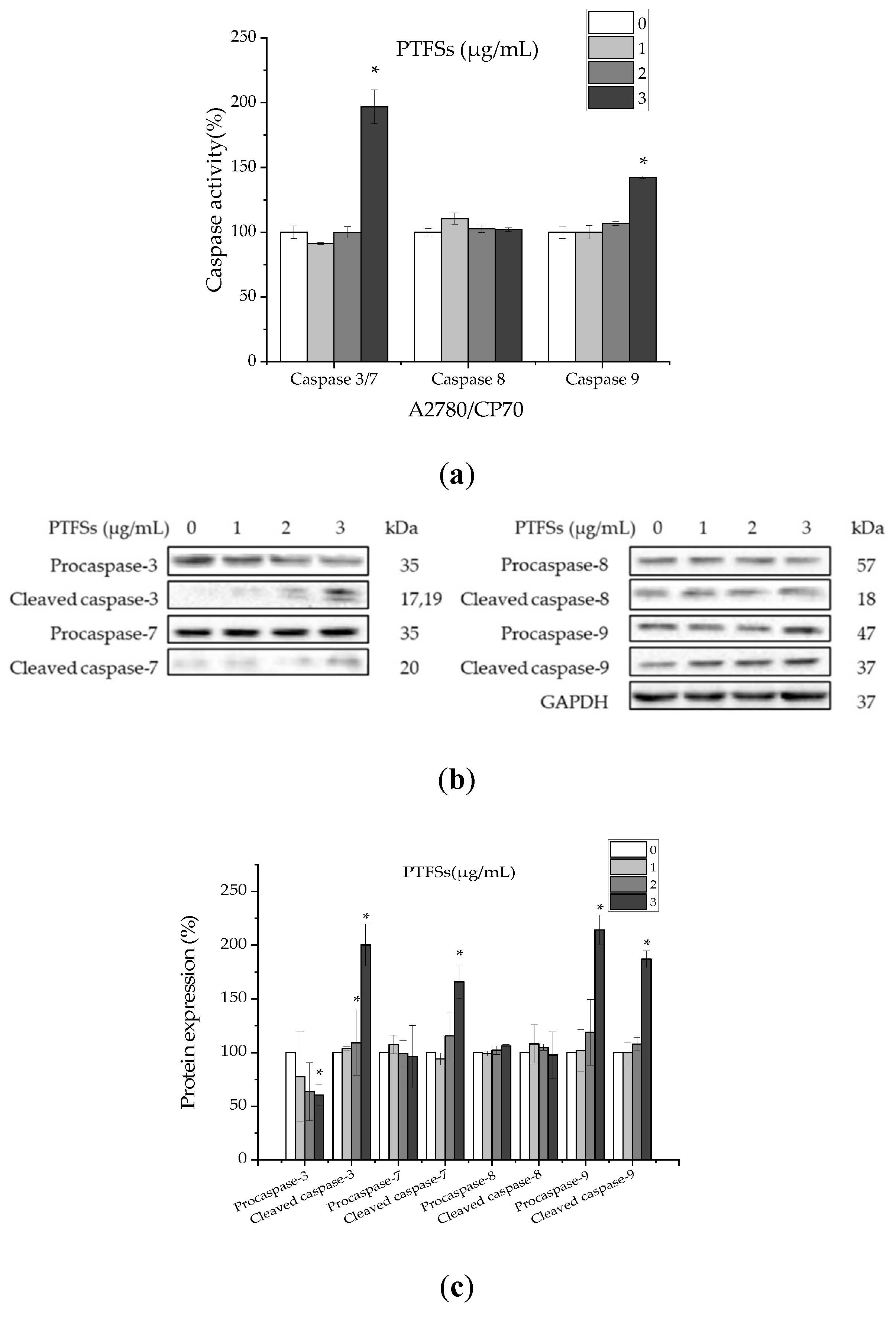
2.4. PTFSs Induce Apoptosis via Caspase-3/7 and -9 Activation in A2780/CP70s
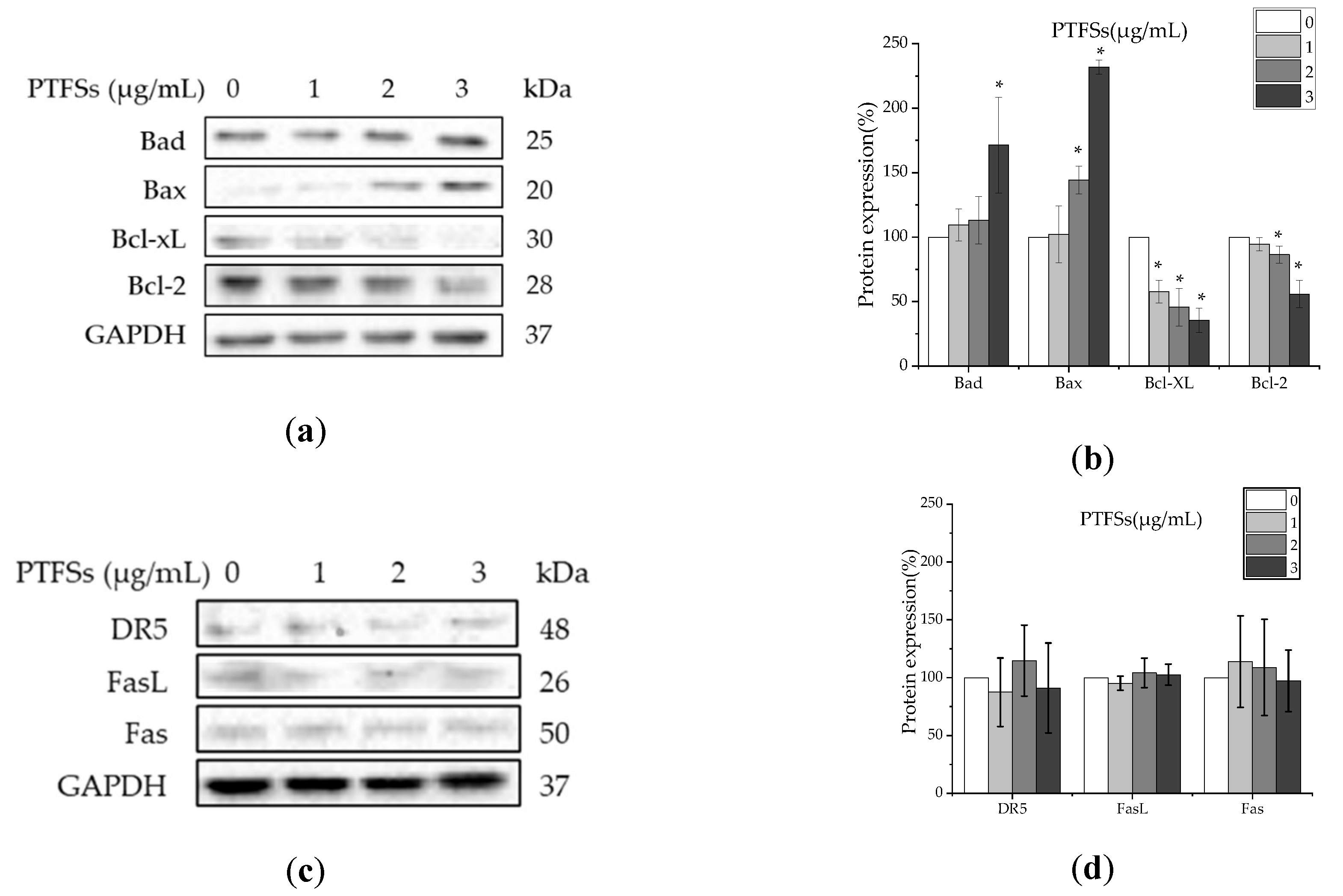
2.5. PTFSs Induce Apoptosis through the Intrinsic Rather than Extrinsic Apoptotic Pathway in A2780/CP70s
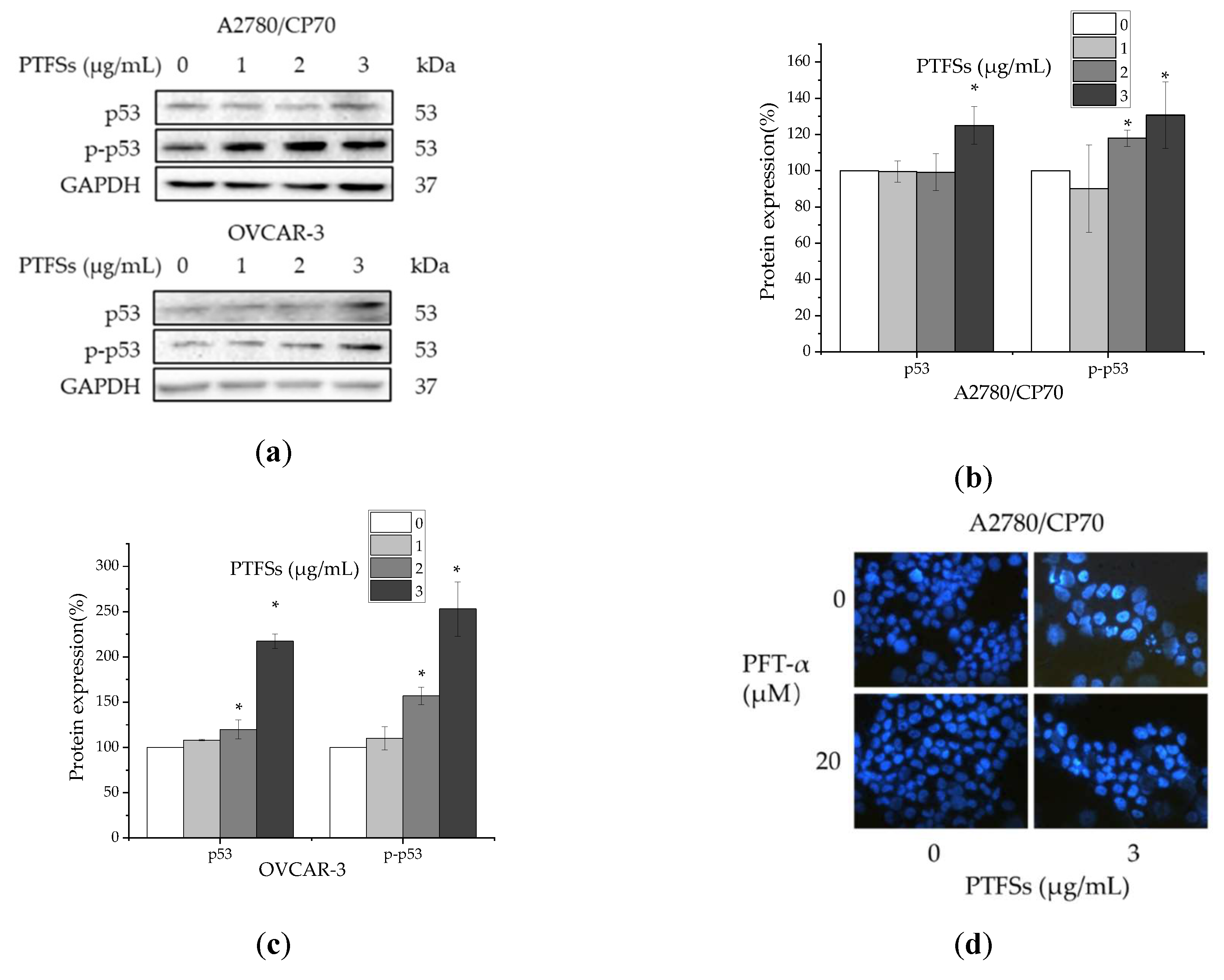
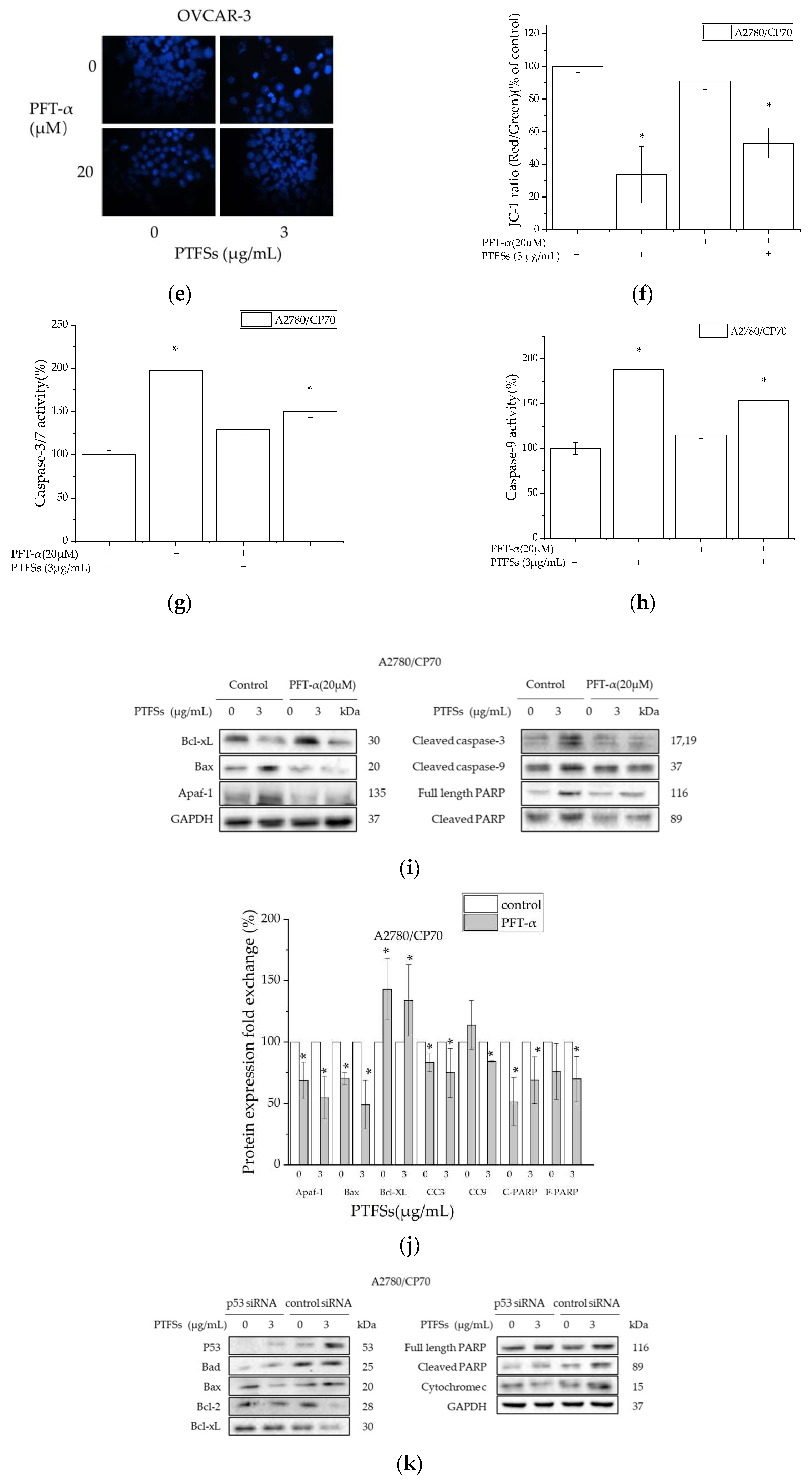
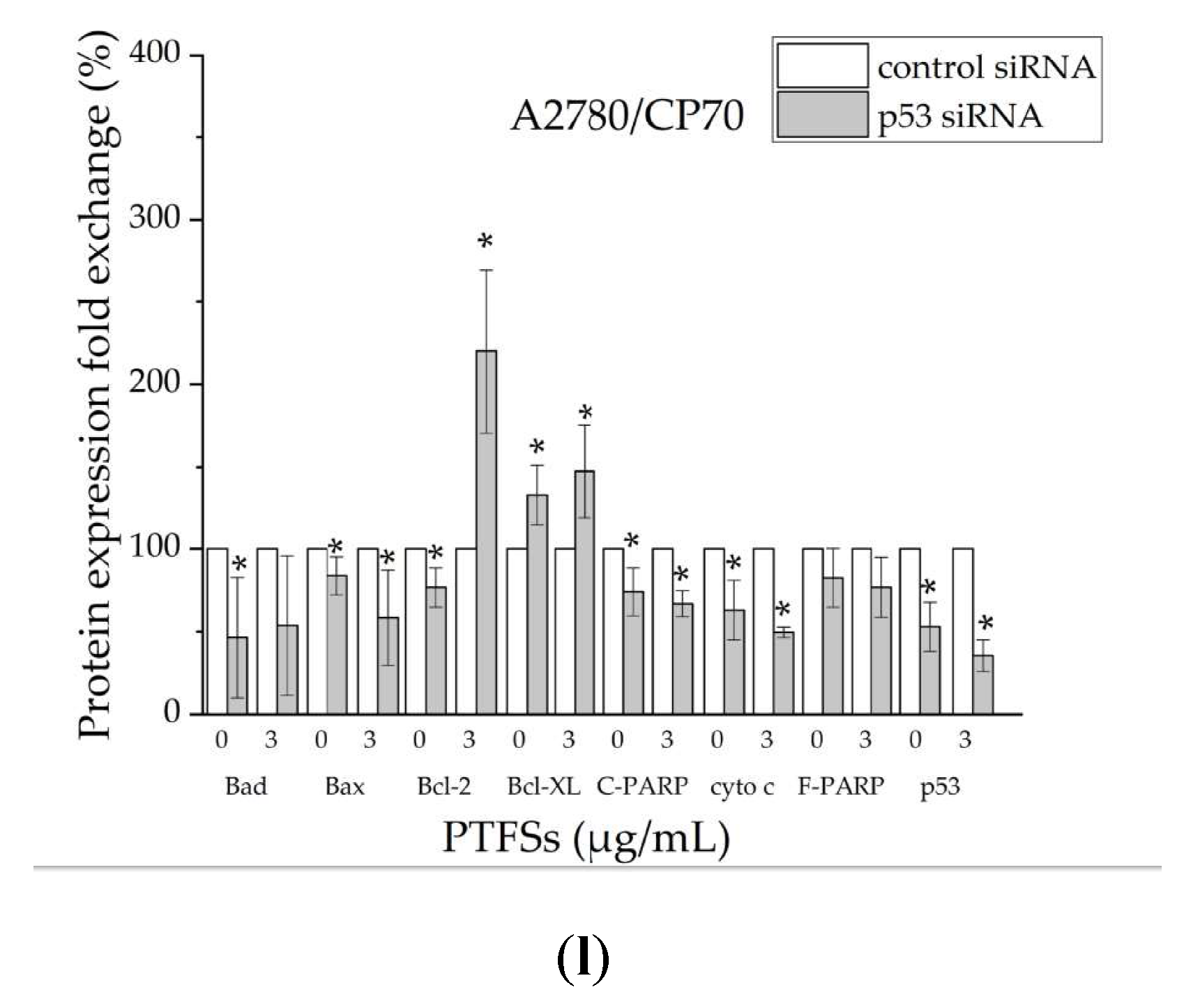
2.6. PTFS-Induced Intrinsic Apoptosis Is p53-Dependent in Ovarian Cancer Cells
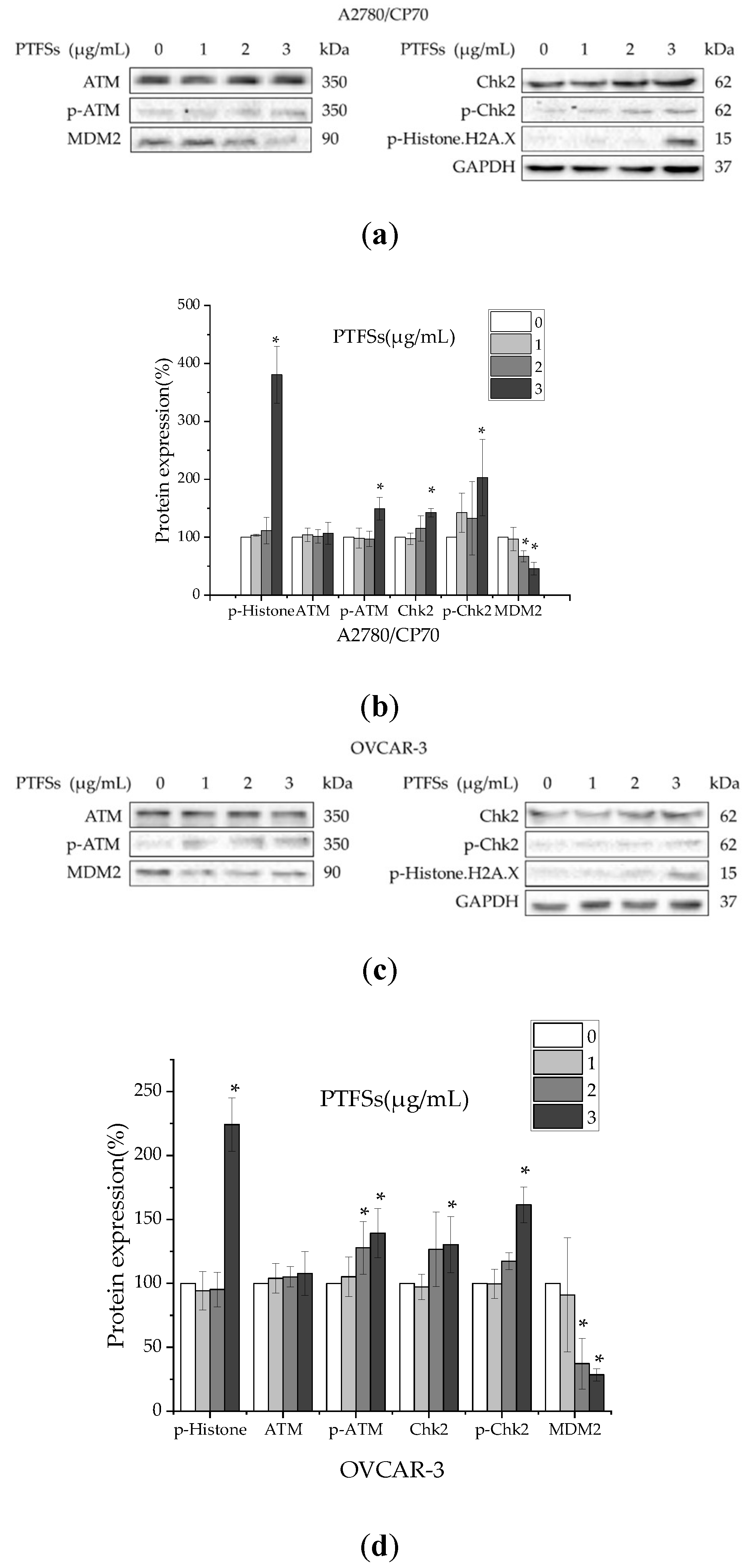
2.7. PTFSs Induce DNA Damage in Ovarian Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Preparation of Purified Tea Flower Saponins (PTFSs)
4.2. Cell Culture
4.3. Cell Viability
4.4. Hoechst 33342 Staining
4.5. Evaluating Mitochondrial Membrane Potential
4.6. Apoptosis Analysis by Flow Cytometry
4.7. Cellular Caspase Activity Assay
4.8. Small Interfering RNA (siRNA) Transfecting
4.9. Western Blotting
4.10. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BSA | Bovine serum albumin |
| BV | Bed volume |
| CC3 | Cleaved caspase 3 |
| CC9 | Cleaved caspase 9 |
| C-PARP | Cleaved PARP |
| F-PARP | Full length PARP |
| cyto c | Cytochrome c |
| DMSO | Dimethyl sulfoxide |
| FBS | Fetal bovine serum |
| PBS | Phosphate-buffered saline |
| PFT-α | Pifithrin-α |
| PI | Propidium iodide |
| PTFSs | Purified tea (Camellia sinensis) flower saponins |
| SEM | Standard error of mean |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Kevin, B.; Nicolas, G.; Eric, A. Chemoresistance and targeted therapies in ovarian and endometrial cancers. Oncotarget 2017, 8, 4008–4042. [Google Scholar]
- Agarwal, R.; Kaye, S.B. Ovarian cancer: Strategies for overcoming resistance to chemotherapy. Nat. Rev. Cancer 2003, 3, 502–516. [Google Scholar] [CrossRef] [PubMed]
- Norouzi-Barough, L.; Sarookhani, M.R.; Sharifi, M.; Moghbelinejad, S.; Jangjoo, S.; Salehi, R. Molecular mechanisms of drug resistance in ovarian cancer. J. Cell. Physiol. 2018, 233, 4546–4562. [Google Scholar] [CrossRef] [PubMed]
- Cooley, M.; Fang, P.; Fang, F.; Nephew, K.P.; Chien, J. Molecular determinants of chemotherapy resistance in ovarian cancer. Pharmacogenomics 2015, 16, 1763–1767. [Google Scholar] [CrossRef]
- Moxley, K.M.; McMeekin, D.S. Endometrial carcinoma: A review of chemotherapy, drug resistance, and the search for new agents. Oncologist 2010, 15, 1026–1033. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Y.; Zhang, P.-Y. Recent perspectives of epithelial ovarian carcinoma (review). Oncol. Lett. 2016, 12, 3055–3058. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.S.; Wang, X.; Lu, G.; Picinich, S.C. Cancer prevention by tea: Animal studies, molecular mechanisms and human relevance. Nat. Rev. Cancer 2009, 9, 429–439. [Google Scholar] [CrossRef] [Green Version]
- Lu, G.; Liao, J.; Yang, G.; Reuhl, K.R.; Yang, C.S.J.C.R. Inhibition of adenoma progression to adenocarcinoma in a 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone-induced lung tumorigenesis model in a/j mice by tea polyphenols and caffeine. Cancer Res. 2007, 66, 11494–11501. [Google Scholar] [CrossRef] [Green Version]
- Landau, J.M.; Wang, Z.Y.; Yang, G.Y.; Ding, W.; Yang, C.S. Inhibition of spontaneous formation of lung tumors and rhabdomyosarcomas in a/j mice by black and green tea. Carcinogenesis 1998, 19, 501–507. [Google Scholar] [CrossRef] [Green Version]
- Sazuka, M.; Murakami, S.; Isemura, M.; Satoh, K.; Nukiwa, T. Inhibitory effects of green tea infusion on in vitro invasion and in vivo metastasis of mouse lung carcinoma cells. Cancer Lett. 1995, 98, 27–31. [Google Scholar] [CrossRef]
- Li, N.; Chen, X.; Liao, J.; Yang, G.; Wang, S.; Josephson, Y.; Han, C.; Chen, J.; Huang, M.T.; Yang, C.S. Inhibition of 7,12-dimethylbenz[a]anthracene (dmba)-induced oral carcinogenesis in hamsters by tea and curcumin. Carcinogenesis 2002, 23, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Wang, L.D.; Lee, M.J.; Ho, C.T.; Huang, M.T.; Conney, A.H.; Yang, C.S. Inhibition of n-nitrosomethylbenzylamine-induced esophageal tumorigenesis in rats by green and black tea. Carcinogenesis 1995, 16, 2143–2148. [Google Scholar] [CrossRef]
- Yamane, T.; Takahashi, T.; Kuwata, K.; Oya, K.; Inagake, M.; Kitao, Y.; Suganuma, M.; Fujiki, H. Inhibition of n-methyl-n’-nitro-n-nitrosoguanidine-induced carcinogenesis by (-)-epigallocatechin gallate in the rat glandular stomach. Cancer Res. 1995, 55, 2081–2084. [Google Scholar] [PubMed]
- Ping, D.Q.; Sangkil, N.; Smith, D.M. Tea Polyphenol Esters and Analogs Thereof for Cancer Prevention and Treatment. U.S. Patent 6,713,506, 30 March 2004. [Google Scholar]
- Zong, J.; Wang, D.; Jiao, W.; Zhang, L.; Bao, G.; Ho, C.-T.; Hou, R.; Wan, X. Oleiferasaponin c6 from the seeds of camellia oleifera abel.: A novel compound inhibits proliferation through inducing cell-cycle arrest and apoptosis on human cancer cell lines in vitro. RSC Adv. 2016, 6, 91386–91393. [Google Scholar] [CrossRef]
- Zhao, W.H.; Li, N.; Zhang, X.R.; Wang, W.L.; Li, J.Y.; Si, Y.Y. Cancer chemopreventive theasaponin derivatives from the total tea seed saponin of camellia sinensis. J. Funct. Foods 2015, 12, 192–198. [Google Scholar] [CrossRef]
- Zong, J.; Wang, R.; Bao, G.; Ling, T.; Zhang, L.; Zhang, X.; Hou, R. Novel triterpenoid saponins from residual seed cake of camellia oleifera abel. Show anti-proliferative activity against tumor cells. Fitoterapia 2015, 104, 7–13. [Google Scholar] [CrossRef]
- Lee, A.H.; Su, D.; Pasalich, M.; Binns, C.W. Tea consumption reduces ovarian cancer risk. Cancer Epidemiol. 2013, 37, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Way, T.-D.; Lin, H.-Y.; Hua, K.-T.; Lee, J.-C.; Li, W.-H.; Lee, M.-R.; Shuang, C.-H.; Lin, J.-K. Beneficial effects of different tea flowers against human breast cancer mcf-7 cells. Food Chem. 2009, 114, 1231–1236. [Google Scholar] [CrossRef]
- Xu, R.; Ye, H.; Sun, Y.; Tu, Y.; Zeng, X. Preparation, preliminary characterization, antioxidant, hepatoprotective and antitumor activities of polysaccharides from the flower of tea plant (camellia sinensis). Food Chem. Toxicol. 2012, 50, 2473–2480. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Jin, Y.; Xu, Y.; Wu, Y.; Xu, J.; Tu, Y. Safety evaluation of tea (camellia sinensis (l.) o. Kuntze) flower extract: Assessment of mutagenicity, and acute and subchronic toxicity in rats. J. Ethnopharmacol. 2011, 133, 583–590. [Google Scholar] [CrossRef]
- Shen, X.; Shi, L.; Pan, H.; Li, B.; Wu, Y.; Tu, Y. Identification of triterpenoid saponins in flowers of four camellia sinensis cultivars from zhejiang province: Differences between cultivars, developmental stages, and tissues. Ind. Crop. Prod. 2017, 95, 140–147. [Google Scholar] [CrossRef]
- Matsuda, H.; Nakamura, S.; Morikawa, T.; Muraoka, O.; Yoshikawa, M. New biofunctional effects of the flower buds of camellia sinensis and its bioactive acylated oleanane-type triterpene oligoglycosides. J. Nat. Med. 2016, 70, 689–701. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, H.; Hamao, M.; Nakamura, S.; Kon’i, H.; Murata, M.; Yoshikawa, M. Medicinal flowers. Xxxiii. Anti-hyperlipidemic and anti-hyperglycemic effects of chakasaponins i-iii and structure of chakasaponin iv from flower buds of chinese tea plant (camellia sinensis). Chem. Pharm. Bull. (Tokyo) 2012, 60, 674–680. [Google Scholar] [CrossRef] [Green Version]
- Kitagawa, N.; Morikawa, T.; Motai, C.; Ninomiya, K.; Okugawa, S.; Nishida, A.; Yoshikawa, M.; Muraoka, O. The antiproliferative effect of chakasaponins i and ii, floratheasaponin a, and epigallocatechin 3-o-gallate isolated from camellia sinensis on human digestive tract carcinoma cell lines. Int. J. Mol. Sci. 2016, 17, 1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, M.; Sugimoto, S.; Kato, Y.; Nakamura, S.; Wang, T.; Yamashita, C.; Matsuda, H. Acylated oleanane-type triterpene saponins with acceleration of gastrointestinal transit and inhibitory effect on pancreatic lipase from flower buds of chinese tea plant (camellia sinensis). Chem. Biodivers. 2009, 6, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Hamao, M.; Matsuda, H.; Nakamura, S.; Nakashima, S.; Semura, S.; Maekubo, S.; Wakasugi, S.; Yoshikawa, M. Anti-obesity effects of the methanolic extract and chakasaponins from the flower buds of camellia sinensis in mice. Bioorgan. Med. Chem. 2011, 19, 6033–6041. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.M.; Ren, N.; Rankin, G.O.; Li, B.; Rojanasakul, Y.; Tu, Y.Y.; Chen, Y.C. Anti-proliferative effect and cell cycle arrest induced by saponins extracted from tea (camellia sinensis) flower in human ovarian cancer cells. J. Funct. Foods 2017, 37, 310–321. [Google Scholar] [CrossRef]
- Zamzami, N.; Kroemer, G. Apoptosis: Mitochondrial membrane permeabilization—The (w) hole story? Curr. Biol. 2003, 13, R71–R73. [Google Scholar] [CrossRef] [Green Version]
- Green, D.R.; Reed, J.C. Mitochondria and apoptosis. Science 1998, 281, 1309–1312. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. The bcl-2 protein family: Arbiters of cell survival. Science 1998, 281, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.I.; Kim, J.H.; Lee, K.T.; Choi, J.H. Costunolide induces apoptosis in platinum-resistant human ovarian cancer cells by generating reactive oxygen species. Gynecol. Oncol. 2011, 123, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Wahl, H.; Tan, L.; Griffith, K.; Choi, M.; Liu, J.R. Curcumin enhances apo2l/trail-induced apoptosis in chemoresistant ovarian cancer cells. Gynecol. Oncol. 2007, 105, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Garziera, M.; Cecchin, E.; Giorda, G.; Sorio, R.; Scalone, S.; De Mattia, E.; Roncato, R.; Gagno, S.; Poletto, E.; Romanato, L.; et al. Clonal evolution of tp53 c.375+ 1G> A mutation in pre- and post- neo-adjuvant chemotherapy (nact) tumor samples in high-grade serous ovarian cancer (hgsoc). Cells 2019, 8, 1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garziera, M.; Roncato, R.; Montico, M.; De Mattia, E.; Gagno, S.; Poletto, E.; Scalone, S.; Canzonieri, V.; Giorda, G.; Sorio, R.; et al. New challenges in tumor mutation heterogeneity in advanced ovarian cancer by a targeted next-generation sequencing (ngs) approach. Cells 2019, 8, 584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, A.J.; Hu, W.; Feng, Z. The p53 pathway: What questions remain to be explored? Curr. Biol. 2003, 13, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Giaccia, A.J.; Kastan, M.B. The complexity of p53 modulation: Emerging patterns from divergent signals. Genes Dev. 1998, 12, 2973–2983. [Google Scholar] [CrossRef] [Green Version]
- Gudkov, A.V.; Komarova, E.A. The role of p53 in determining sensitivity to radiotherapy. Nat. Rev. Cancer 2003, 3, 117–129. [Google Scholar] [CrossRef]
- Pallikkathayil, L.; Crighton, F.; Aaronson, L.S. Balancing ethical quandaries with scientific rigor: Part 1. West J. Nurs. Res. 1998, 20, 388–393. [Google Scholar] [CrossRef]
- Morales, J.; Li, L.; Fattah, F.J.; Dong, Y.; Bey, E.A.; Patel, M.; Gao, J.; Boothman, D.A. Review of poly (adp-ribose) polymerase (parp) mechanisms of action and rationale for targeting in cancer and other diseases. Crit. Rev. Eukaryot. Gene Expr. 2014, 24, 15–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tibbetts, R.S.; Brumbaugh, K.M.; Williams, J.M.; Sarkaria, J.N.; Cliby, W.A.; Shieh, S.Y.; Taya, Y.; Prives, C.; Abraham, R.T. A role for atr in the DNA damage-induced phosphorylation of p53. Genes Dev. 1999, 13, 152–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abedini, M.R.; Muller, E.J.; Bergeron, R.; Gray, D.A.; Tsang, B.K. Akt promotes chemoresistance in human ovarian cancer cells by modulating cisplatin-induced, p53-dependent ubiquitination of flice-like inhibitory protein. Oncogene 2010, 29, 11–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Wu, Q.-S.; Meng, F.-C.; Tang, Z.-H.; Chen, X.; Lin, L.-G.; Chen, P.; Qiang, W.-A.; Wang, Y.-T.; Zhang, Q.-W.; et al. Chikusetsusaponin iva methyl ester induces g1 cell cycle arrest, triggers apoptosis and inhibits migration and invasion in ovarian cancer cells. Phytomedicine 2016, 23, 1555–1565. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X. Paris saponin ii of rhizoma paridis—A novel inducer of apoptosis in human ovarian cancer cells. Biosci. Trends 2012, 6, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Celis, J.E.; Bravo, R. Synthesis of the nuclear protein cyclin in growing, senescent and morphologically transformed human skin fibroblasts. FEBS Lett. 1984, 165, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Kubben, F.J.; Peeters-Haesevoets, A.; Engels, L.G.; Baeten, C.G.; Schutte, B.; Arends, J.W.; Stockbrugger, R.W.; Blijham, G.H. Proliferating cell nuclear antigen (pcna): A new marker to study human colonic cell proliferation. Gut 1994, 35, 530–535. [Google Scholar] [CrossRef] [Green Version]
- Miyachi, K.; Fritzler, M.J.; EM, T. Auroantibody to a nuclear antigen in proliferating cells. J. Immunol. 1978, 121, 2228–2234. [Google Scholar]
- Blagosklonny, M.V. Prospective strategies to enforce selectively cell death in cancer cells. Oncogene 2004, 23, 2967–2975. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Zamzami, N.; de La Motte Rouge, T.; Lemaire, C.; Brenner, C.; Kroemer, G. Methods for the assessment of mitochondrial membrane permeabilization in apoptosis. Apoptosis 2007, 12, 803–813. [Google Scholar] [CrossRef] [Green Version]
- Metivier, D.; Dallaporta, B.; Zamzami, N.; Larochette, N.; Susin, S.A.; Marzo, I.; Kroemer, G. Cytofluorometric detection of mitochondrial alterations in early cd95/fas/apo-1-triggered apoptosis of jurkat t lymphoma cells. Comparison of seven mitochondrion-specific fluorochromes. Immunol. Lett. 1998, 61, 157–163. [Google Scholar] [CrossRef]
- Garrido, C.; Galluzzi, L.; Brunet, M.; Puig, P.E.; Didelot, C.; Kroemer, G. Mechanisms of cytochrome c release from mitochondria. Cell Death Differ. 2006, 13, 1423–1433. [Google Scholar] [CrossRef] [Green Version]
- Nair, J.; van Staden, J. Phenanthridone alkaloids of the amaryllidaceae as activators of the apoptosis-related proteolytic enzymes, caspases. Nat. Prod. Commun. 2018, 13, 1375–1380. [Google Scholar] [CrossRef] [Green Version]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef] [PubMed]
- Day, T.W.; Huang, S.; Safa, A.R. C-flip knockdown induces ligand-independent dr5-, fadd-, caspase-8-, and caspase-9-dependent apoptosis in breast cancer cells. Biochem. Pharmacol. 2008, 76, 1694–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaVallee, T.M.; Zhan, X.H.; Johnson, M.S.; Herbstritt, C.J.; Swartz, G.; Williams, M.S.; Hembrough, W.A.; Green, S.J.; Pribluda, V.S. 2-methoxyestradiol up-regulates death receptor 5 and induces apoptosis through activation of the extrinsic pathway. Cancer Res. 2003, 63, 468–475. [Google Scholar] [PubMed]
- Darcy, K.M.; Brady, W.E.; McBroom, J.W.; Bell, J.G.; Young, R.C.; McGuire, W.P.; Linnolia, R.I.; Hendricks, D.; Bonome, T.; Farley, J.H. Associations between p53 overexpression and multiple measures of clinical outcome in high-risk, early stage or suboptimally-resected, advanced stage epithelial ovarian cancers: A gynecologic oncology group study. Gynecol. Oncol. 2008, 111, 487–495. [Google Scholar] [CrossRef] [Green Version]
- Tu, Y.Y.; Kim, E.; Gao, Y.; Rankin, G.O.; Li, B.; Chen, Y.C. Theaflavin-3, 3′-digallate induces apoptosis and g2 cell cycle arrest through the akt/mdm2/p53 pathway in cisplatin-resistant ovarian cancer a2780/cp70 cells. Int. J. Oncol. 2016, 48, 2657–2665. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.B.; Wang, F.; Rankin, G.O.; Rojanasakul, Y.; Tu, Y.Y.; Chen, Y.C. Inhibitory effect of black tea pigments, theaflavin-3/3′-gallate against cisplatin-resistant ovarian cancer cells by inducing apoptosis and g1 cell cycle arrest. Int. J. Oncol. 2017, 51, 1508–1520. [Google Scholar] [CrossRef] [Green Version]
- Kong, D.; Ma, S.; Liang, B.; Yi, H.; Zhao, Y.; Xin, R.; Cui, L.; Jia, L.; Liu, X.; Liu, X. The different regulatory effects of p53 status on multidrug resistance are determined by autophagy in ovarian cancer cells. Biomed. Pharmacother. 2012, 66, 271–278. [Google Scholar] [CrossRef]
- Kao, C.L.; Hsu, H.S.; Chen, H.W.; Cheng, T.H. Rapamycin increases the p53/mdm2 protein ratio and p53-dependent apoptosis by translational inhibition of mdm2 in cancer cells. Cancer Lett. 2009, 286, 250–259. [Google Scholar] [CrossRef]
- Li, B.; Gao, Y.; Rankin, G.O.; Rojanasakul, Y.; Cutler, S.J.; Tu, Y.Y.; Chen, Y.C. Chaetoglobosin k induces apoptosis and g2 cell cycle arrest through p53-dependent pathway in cisplatin-resistant ovarian cancer cells. Cancer Lett. 2015, 356, 418–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzon, I.; Pellarin, I.; Pellizzari, I.; D’Andrea, S.; Belletti, B.; Sonego, M.; Baldassarre, G.; Schiappacassi, M. Identification and characterization of a new platinum-induced tp53 mutation in mdah ovarian cancer cells. Cells 2019, 9, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xirodimas, D.P.; Saville, M.K.; Bourdon, J.C.; Hay, R.T.; Lane, D.P. Mdm2-mediated nedd8 conjugation of p53 inhibits its transcriptional activity. Cell 2004, 118, 83–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, D.L.; Schafer, S.A.; Nordquist, R.; Payton, M.E.; Dickey, D.T.; Bartels, K.E. Comparison of a high power diode laser with the Nd: YAG laser using in situ wound strength analysis of healing cutaneous incisions. Lasers Surg. Med. 1997, 21, 248–254. [Google Scholar] [CrossRef]
- Xu, Y. Regulation of p53 responses by post-translational modifications. Cell Death Differ. 2003, 10, 400–403. [Google Scholar] [CrossRef]
- Lee, J.H.; Paull, T.T. Activation and regulation of atm kinase activity in response to DNA double-strand breaks. Oncogene 2007, 26, 7741–7748. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, S.; Rotman, G.; Ogawa, A.; Shiloh, Y.; Tamai, K.; Elledge, S.J. Ataxia telangiectasia-mutated phosphorylates chk2 in vivo and in vitro. Proc. Natl. Acad. Sci. USA 2000, 97, 10389–10394. [Google Scholar] [CrossRef] [Green Version]
- Melchionna, R.; Chen, X.B.; Blasina, A.; McGowan, C.H. Threonine 68 is required for radiation-induced phosphorylation and activation of cds1. Nat. Cell Biol. 2000, 2, 762–765. [Google Scholar] [CrossRef]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA double-stranded breaks induce histone h2ax phosphorylation on serine 139. J. Biol. Chem. 1998, 273, 5858–5868. [Google Scholar] [CrossRef] [Green Version]
- Burma, S.; Chen, B.P.; Murphy, M.; Kurimasa, A.; Chen, D.J. Atm phosphorylates histone h2ax in response to DNA double-strand breaks. J. Biol. Chem. 2001, 276, 42462–42467. [Google Scholar] [CrossRef] [Green Version]
- Shieh, S.Y.; Ikeda, M.; Taya, Y.; Prives, C. DNA damage-induced phosphorylation of p53 alleviates inhibition by mdm2. Cell 1997, 91, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Sahu, R.P.; Batra, S.; Srivastava, S.K. Activation of atm/chk1 by curcumin causes cell cycle arrest and apoptosis in human pancreatic cancer cells. Br. J. Cancer 2009, 100, 1425–1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Man, S.; Gao, W.; Zhang, Y.; Huang, L.; Liu, C. Chemical study and medical application of saponins as anti-cancer agents. Fitoterapia 2010, 81, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yuan, L.; Xiao, H.; Xiao, C.; Wang, Y.; Liu, X. Momordin ic induces hepg2 cell apoptosis through mapk and pi3k/akt-mediated mitochondrial pathways. Apoptosis 2013, 18, 751–765. [Google Scholar] [CrossRef]
- Wang, J.; Han, Y.; Wang, M.; Zhao, Q.Q.; Chen, X.F.; Liu, X.B. Natural triterpenoid saponin momordin ic suppresses hepg2 cell invasion via cox-2 inhibition and ppar gamma activation. Toxicol. Vitro 2020, 65, 9. [Google Scholar] [CrossRef]
- Fu, R.; Zhang, L.; Li, Y.; Li, B.; Ming, Y.; Li, Z.; Xing, H.; Chen, J. Saikosaponin d inhibits autophagosome-lysosome fusion and induces autophagy-independent apoptosis in mda-mb-231 breast cancer cells. Mol. Med. Rep. 2020. [Google Scholar] [CrossRef]
- Guan, Y.Y.; Liu, H.J.; Luan, X.; Xu, J.R.; Lu, Q.; Liu, Y.R.; Gao, Y.G.; Zhao, M.; Chen, H.Z.; Fang, C. Raddeanin a, a triterpenoid saponin isolated from anemone raddeana, suppresses the angiogenesis and growth of human colorectal tumor by inhibiting vegfr2 signaling. Phytomedicine 2015, 22, 103–110. [Google Scholar] [CrossRef]
- Yu, K.; Chen, F.; Li, C. Absorption, disposition, and pharmacokinetics of saponins from Chinese medicinal herbs: What do we know and what do we need to know more? Curr. Drug Metab. 2012, 13, 577–598. [Google Scholar] [CrossRef]
- Hu, X.; Lin, M.; Zhu, W.; Zheng, Y.; Zhang, Q.; Wu, G.; Qiu, Y. Potential cytotoxicity, pharmacokinetics, and excretion properties of sapindoside b from the seeds of nigella sativa var. Hispidula. Planta Med. 2020, 86, 356–363. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Retention Time (Min) | [M-H]− | MS2 | Formula | Peak Identity | % |
|---|---|---|---|---|---|---|
| 1 | 50.52 | 1173.5714 | 1042,993,909,569 | C57H90O25 | Chakasaponin IV | 8.1 |
| 2 | 51.57 | 1215.5828 | 1083,1035,951,933,611 | C59H92O26 | Chakasaponin I | 80.1 |
| 3 | 62.43 | 1215.5949 | 1083,1035,951,933,611 | C59H92O26 | Floratheasaponin A | 1.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, N.; Chen, L.; Li, B.; Rankin, G.O.; Chen, Y.C.; Tu, Y. Purified Tea (Camellia sinensis (L.) Kuntze) Flower Saponins Induce the p53-Dependent Intrinsic Apoptosis of Cisplatin-Resistant Ovarian Cancer Cells. Int. J. Mol. Sci. 2020, 21, 4324. https://doi.org/10.3390/ijms21124324
Ren N, Chen L, Li B, Rankin GO, Chen YC, Tu Y. Purified Tea (Camellia sinensis (L.) Kuntze) Flower Saponins Induce the p53-Dependent Intrinsic Apoptosis of Cisplatin-Resistant Ovarian Cancer Cells. International Journal of Molecular Sciences. 2020; 21(12):4324. https://doi.org/10.3390/ijms21124324
Chicago/Turabian StyleRen, Ning, Lianfu Chen, Bo Li, Gary O. Rankin, Yi Charlie Chen, and Youying Tu. 2020. "Purified Tea (Camellia sinensis (L.) Kuntze) Flower Saponins Induce the p53-Dependent Intrinsic Apoptosis of Cisplatin-Resistant Ovarian Cancer Cells" International Journal of Molecular Sciences 21, no. 12: 4324. https://doi.org/10.3390/ijms21124324
APA StyleRen, N., Chen, L., Li, B., Rankin, G. O., Chen, Y. C., & Tu, Y. (2020). Purified Tea (Camellia sinensis (L.) Kuntze) Flower Saponins Induce the p53-Dependent Intrinsic Apoptosis of Cisplatin-Resistant Ovarian Cancer Cells. International Journal of Molecular Sciences, 21(12), 4324. https://doi.org/10.3390/ijms21124324