Prediction of the Secretome and the Surfaceome: A Strategy to Decipher the Crosstalk between Adipose Tissue and Muscle during Fetal Growth

Abstract
:
1. Introduction
2. Results
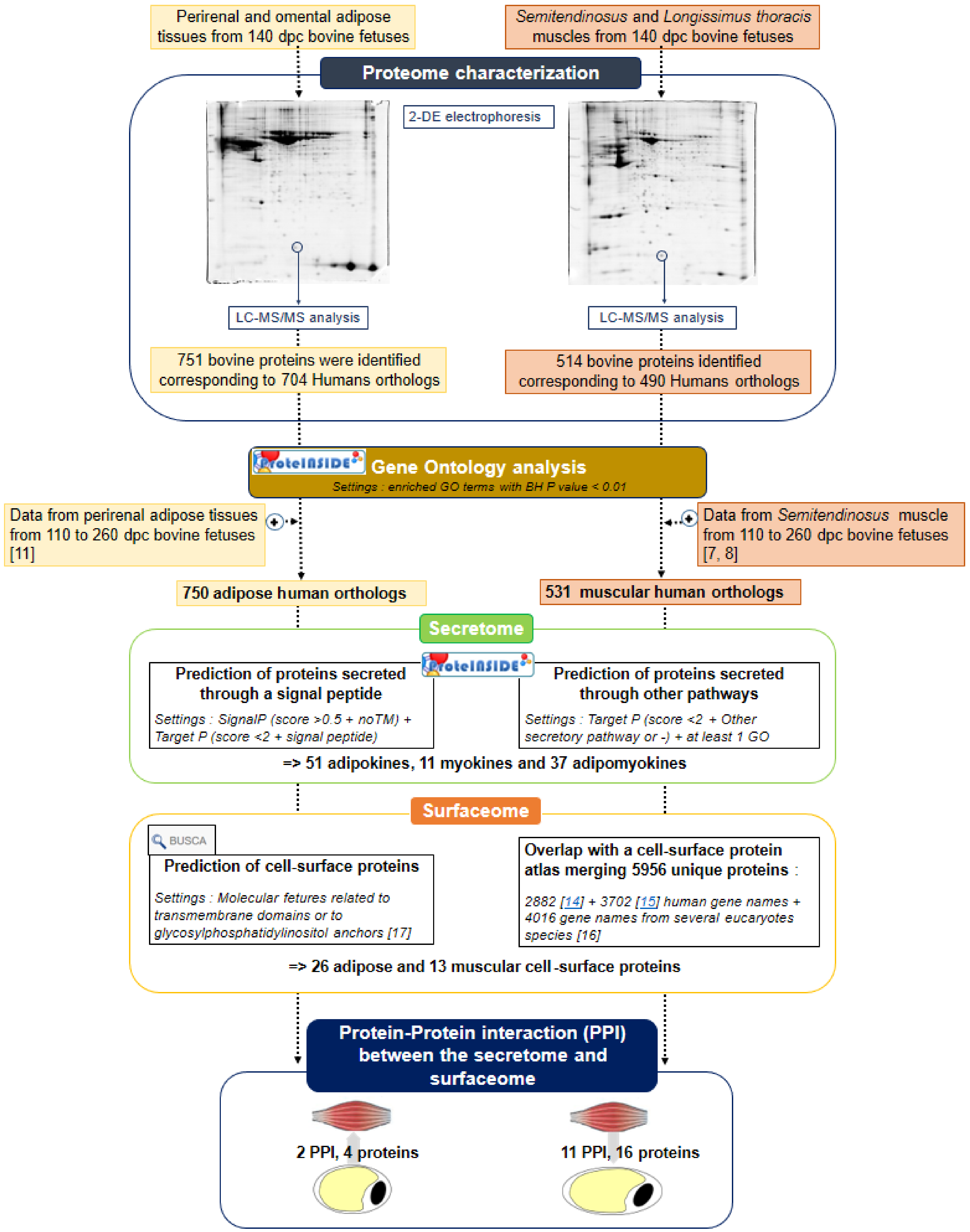
2.1. Proteome Description of the Fetal Bovine Muscle and Adipose Tissues
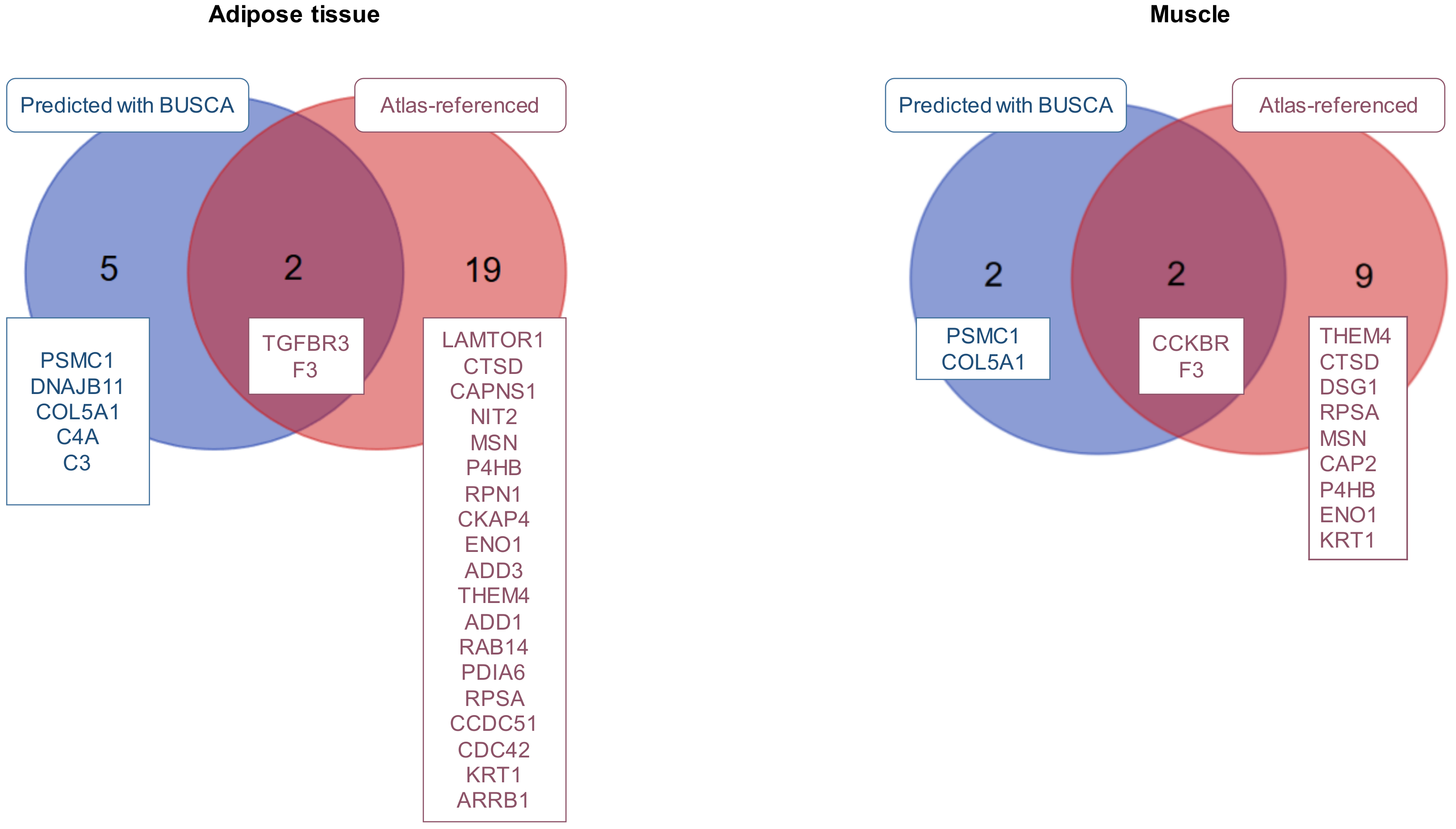
2.2. Prediction of Proteins Secreted by the Fetal Bovine Muscles and Adipose Tissues
2.3. Prediction of Fetal Bovine Muscle and Adipose Tissue Surfaceomes
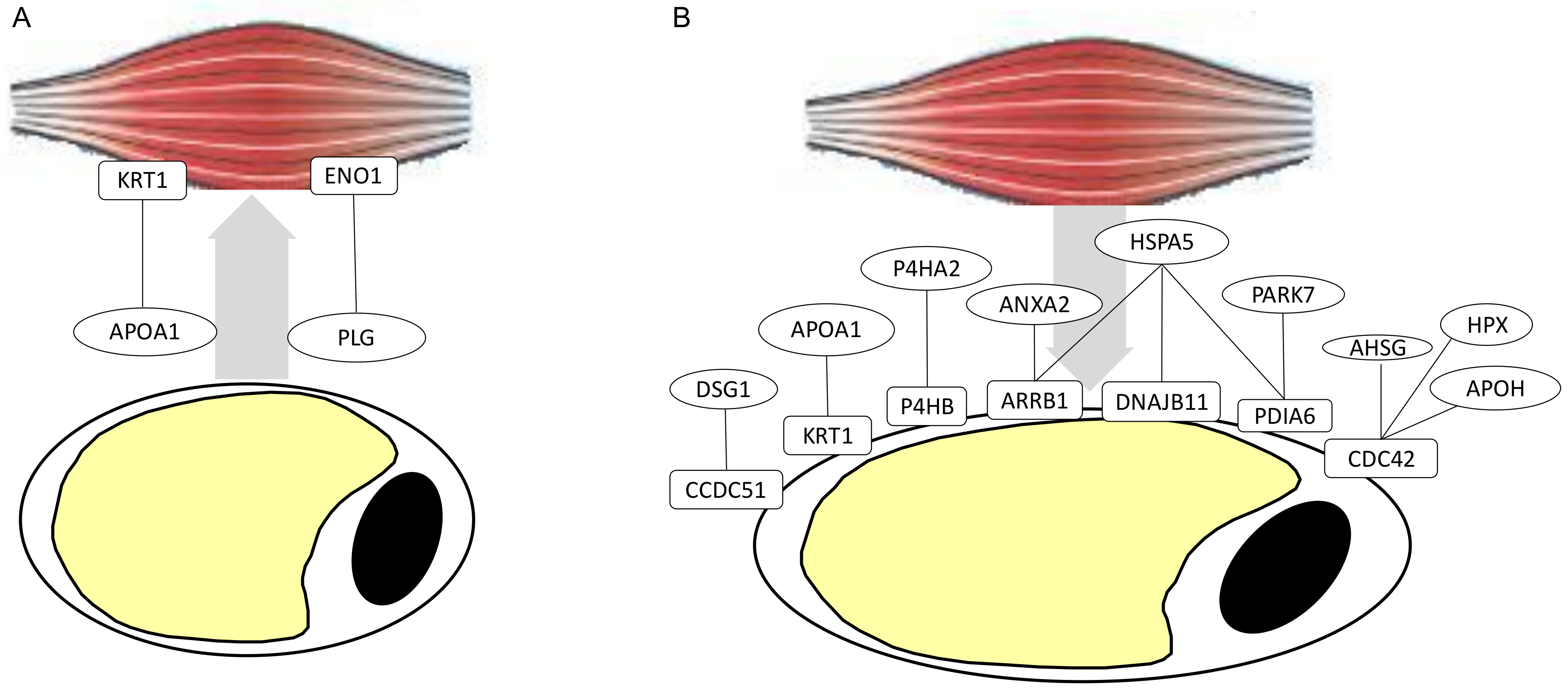
2.4. Predictions of Proteins Interactions between Adipose Tissues and Muscles
3. Discussion
3.1. Pathways that Sustain Fetal Bovine Adipogenesis and Myogenesis
3.2. Secretome and Surfaceome Involvement in Deciphering the Adipose and Musclar Crosstalk
4. Materials and Methods
4.1. Muscles and Adipose Tissues Sampling
4.2. Large-Scale Proteomic Analysis
4.3. Bioinformatic Analysis and Data Mining
4.3.1. Gene Ontology (GO) Analysis
4.3.2. Creation of a Compendium of Fetal Adipose and Muscular Proteomes
4.3.3. Computational Prediction of the Proteins Secreted by Fetal Adipose Tissue or Muscle
4.3.4. Computational Prediction of the Fetal Adipose Tissue or Muscle Surfaceome
4.3.5. Computational Prediction of Protein–Protein Interactions between Cell Surface Proteins and Secreted Proteins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AT | Adipose tissue |
| Da | Dalton |
| dpc | Days post-conception |
| GO | Gene Ontology |
| ID | Identifier |
| IEF | Isoelectrofocalisation |
| PPI | Protein–protein interaction |
References
- Bonnet, M.; Cassar-Malek, I.; Chilliard, Y.; Picard, B. Ontogenesis of muscle and adipose tissues and their interactions in ruminants and other species. Animal 2010, 4, 1093–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hausman, G.J.; Basu, U.; Wei, S.; Hausman, D.B.; Dodson, M.V. Preadipocyte and Adipose Tissue Differentiation in Meat Animals: Influence of Species and Anatomical Location. Annu. Rev. Anim. Biosci. 2014, 2, 323–351. [Google Scholar] [CrossRef]
- Picard, B.; Lefaucheur, L.; Berri, C.; Duclos, M.J. Muscle fibre ontogenesis in farm animal species. Reprod. Nutr. Dev. 2002, 42, 415–431. [Google Scholar] [CrossRef] [Green Version]
- Draeger, A.; Weeds, A.G.; Fitzsimons, R.B. Primary, Secondary and Tertiary Myotubes in Developing Skeletal-Muscle-a New Approach to the Analysis of Human Myogenesis. J. Neurol. Sci. 1987, 81, 19–43. [Google Scholar] [CrossRef]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Naslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Lehnert, S.A.; Reverter, A.; Byrne, K.A.; Wang, Y.; Nattrass, G.S.; Hudson, N.J.; Greenwood, P.L. Gene expression studies of developing bovine longissimus muscle from two different beef cattle breeds. BMC Dev. Biol. 2007, 7, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaze, T.; Meunier, B.; Chambon, C.; Jurie, C.; Picard, B. In vivo proteome dynamics during early bovine myogenesis. Proteomics 2008, 8, 4236–4248. [Google Scholar] [CrossRef]
- Chaze, T.; Meunier, B.; Chambon, C.; Jurie, C.; Picard, B. Proteome dynamics during contractile and metabolic differentiation of bovine foetal muscle. Animal 2009, 3, 980–1000. [Google Scholar] [CrossRef] [Green Version]
- Sudre, K.; Leroux, C.; Pietu, G.; Cassar-Malek, I.; Petit, E.; Listrat, A.; Auffray, C.; Picard, B.; Martin, P.; Hocquette, J.F. Transcriptome analysis of two bovine muscles during ontogenesis. J. Biochem. 2003, 133, 745–756. [Google Scholar] [CrossRef]
- Landis, M.D.; Carstens, G.E.; McPhail, E.G.; Randel, R.D.; Green, K.K.; Slay, L.; Smith, S.B. Ontogenic development of brown adipose tissue in Angus and Brahman fetal calves. J. Anim. Sci. 2002, 80, 591–601. [Google Scholar] [CrossRef]
- Taga, H.; Chilliard, Y.; Meunier, B.; Chambon, C.; Picard, B.; Zingaretti, M.C.; Cinti, S.; Bonnet, M. Cellular and molecular large-scale features of fetal adipose tissue: Is bovine perirenal adipose tissue brown? J. Cell Physiol. 2012, 227, 1688–1700. [Google Scholar] [CrossRef] [PubMed]
- Kaspric, N.; Picard, B.; Reichstadt, M.; Tournayre, J.; Bonnet, M. ProteINSIDE to Easily Investigate Proteomics Data from Ruminants: Application to Mine Proteome of Adipose and Muscle Tissues in Bovine Foetuses. PLoS ONE 2015, 10, e0128086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argiles, J.M.; Orpi, M.; Busquets, S.; Lopez-Soriano, F.J. Myostatin: More than just a regulator of muscle mass. Drug Discov. Today 2012, 17, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Bausch-Fluck, D.; Goldmann, U.; Muller, S.; van Oostrum, M.; Muller, M.; Schubert, O.T.; Wollscheid, B. The in silico human surfaceome. Proc. Natl. Acad. Sci. USA 2018, 115, E10988–E10997. [Google Scholar] [CrossRef] [Green Version]
- Da Cunha, J.P.; Galante, P.A.; de Souza, J.E.; de Souza, R.F.; Carvalho, P.M.; Ohara, D.T.; Moura, R.P.; Oba-Shinja, S.M.; Marie, S.K.; Silva, W.A., Jr.; et al. Bioinformatics construction of the human cell surfaceome. Proc. Natl. Acad. Sci. USA 2009, 106, 16752–16757. [Google Scholar] [CrossRef] [Green Version]
- Pierleoni, A.; Indio, V.; Savojardo, C.; Fariselli, P.; Martelli, P.L.; Casadio, R. MemPype: A pipeline for the annotation of eukaryotic membrane proteins. Nucleic Acids Res. 2011, 39, W375–W380. [Google Scholar] [CrossRef] [Green Version]
- Savojardo, C.; Martelli, P.L.; Fariselli, P.; Profiti, G.; Casadio, R. BUSCA: An integrative web server to predict subcellular localization of proteins. Nucleic Acids Res. 2018, 46, W459–W466. [Google Scholar] [CrossRef]
- Nanjappa, V.; Thomas, J.K.; Marimuthu, A.; Muthusamy, B.; Radhakrishnan, A.; Sharma, R.; Khan, A.A.; Balakrishnan, L.; Sahasrabuddhe, N.A.; Kumar, S.; et al. Plasma Proteome Database as a resource for proteomics research: 2014 update. Nucleic Acids Res. 2014, 42, D959–D965. [Google Scholar] [CrossRef] [Green Version]
- Boudon, S.; Henry-Berger, J.; Cassar-Malek, I. Aggregation of Omic Data and Secretome Prediction Enable the Discovery of Candidate Plasma Biomarkers for Beef Tenderness. Int. J. Mol. Sci. 2020, 21, 664. [Google Scholar] [CrossRef] [Green Version]
- Bonnet, M.; Tournayre, J.; Cassar-Malek, I. Integrated data mining of transcriptomic and proteomic datasets to predict the secretome of adipose tissue and muscle in ruminants. Mol. Biosyst. 2016, 12, 2722–2734. [Google Scholar] [CrossRef]
- Thomas, J.L.; Moncollin, V.; Ravel-Chapuis, A.; Valente, C.; Corda, D.; Mejat, A.; Schaeffer, L. PAK1 and CtBP1 Regulate the Coupling of Neuronal Activity to Muscle Chromatin and Gene Expression. Mol. Cell Biol. 2015, 35, 4110–4120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vernochet, C.; Peres, S.B.; Davis, K.E.; McDonald, M.E.; Qiang, L.; Wang, H.; Scherer, P.E.; Farmer, S.R. C/EBPalpha and the corepressors CtBP1 and CtBP2 regulate repression of select visceral white adipose genes during induction of the brown phenotype in white adipocytes by peroxisome proliferator-activated receptor gamma agonists. Mol. Cell Biol. 2009, 29, 4714–4728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.B.; He, Y.K.; Okutsu, M.; Ong, L.C.; Jin, Y.; Zheng, L.; Chow, P.; Yu, S.; Zhang, M.; Yan, Z. Autophagy is involved in adipogenic differentiation by repressesing proteasome-dependent PPAR gamma 2 degradation. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E530–E539. [Google Scholar] [CrossRef] [Green Version]
- Pal, P.; Lochab, S.; Kanaujiya, J.K.; Kapoor, I.; Sanyal, S.; Behre, G.; Trivedi, A.K. E3 Ubiquitin Ligase E6AP Negatively Regulates Adipogenesis by Downregulating Proadipogenic Factor C/EBPalpha. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding adipocyte differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, G.I.; Griffiths, M.R.; Webster, K.J.; Page, M.J.; Tavare, J.M. Proteome analysis of adipogenesis. Proteomics 2004, 4, 1042–1051. [Google Scholar] [CrossRef]
- Molina, H.; Yang, Y.; Ruch, T.; Kim, J.-W.; Mortensen, P.; Otto, T.; Nalli, A.; Tang, Q.-Q.; Lane, M.D.; Chaerkady, R.; et al. Temporal profiling of the adipocyte proteome during differentiation using a five-plex SILAC based strategy. J. Proteome Res. 2009, 8, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Eirin, A.; Riester, S.M.; Zhu, X.Y.; Tang, H.; Evans, J.M.; O’Brien, D.; van Wijnen, A.J.; Lerman, L.O. MicroRNA and mRNA cargo of extracellular vesicles from porcine adipose tissue-derived mesenchymal stem cells. Gene 2014, 551, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Fromm-Dornieden, C.; von der Heyde, S.; Lytovchenko, O.; Salinas-Riester, G.; Brenig, B.; Beissbarth, T.; Baumgartner, B.G. Novel polysome messages and changes in translational activity appear after induction of adipogenesis in 3T3-L1 cells. BMC Mol. Biol. 2012, 13, 9. [Google Scholar] [CrossRef] [Green Version]
- Picard, B.; Berri, C.; Lefaucheur, L.; Molette, C.; Sayd, T.; Terlouw, C. Skeletal muscle proteomics in livestock production. Brief. Funct. Genom. 2010, 9, 259–278. [Google Scholar] [CrossRef] [PubMed]
- Poleti, M.D.; Regitano, L.C.A.; Souza, G.; Cesar, A.S.M.; Simas, R.C.; Silva-Vignato, B.; Oliveira, G.B.; Andrade, S.C.S.; Cameron, L.C.; Coutinho, L.L. Longissimus dorsi muscle label-free quantitative proteomic reveals biological mechanisms associated with intramuscular fat deposition. J. Proteom. 2018, 179, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.Y.; Zhang, W.; Han, X.; Huang, R.H.; Shi, F.X. Cell proliferation and apoptosis in the fetal and neonatal ovary of guinea pigs. Genet. Mol. Res. 2014, 13, 1570–1578. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, B.; Stratmann, B.; Hess, C.; Tschoepe, D.; Gawlowski, T. Impact of GLO1 knock down on GLUT4 trafficking and glucose uptake in L6 myoblasts. PLoS ONE 2013, 8, e65195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stratmann, B.; Goldstein, B.; Thornalley, P.J.; Rabbani, N.; Tschoepe, D. Intracellular Accumulation of Methylglyoxal by Glyoxalase 1 Knock Down Alters Collagen Homoeostasis in L6 Myoblasts. Int. J. Mol. Sci. 2017, 18, 480. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, Y.; Nahar, S.; Miyagi-Shiohira, C.; Kinjo, T.; Kobayashi, N.; Kitamura, S.; Saitoh, I.; Watanabe, M.; Fujita, J.; Noguchi, H. Identification of Proteins Differentially Expressed by Adipose-derived Mesenchymal Stem Cells Isolated from Immunodeficient Mice. Int. J. Mol. Sci. 2019, 20, 2672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, B.L.; Ma, M.T.; Chen, B.; Li, Z.H.; Abdalla, B.A.; Nie, Q.H.; Zhang, X.Q. MiR-16-5p targets SESN1 to regulate the p53 signaling pathway, affecting myoblast proliferation and apoptosis, and is involved in myoblast differentiation. Cell Death Dis. 2018, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Krstic, J.; Reinisch, I.; Schupp, M.; Schulz, T.J.; Prokesch, A. p53 Functions in Adipose Tissue Metabolism and Homeostasis. Int. J. Mol. Sci. 2018, 19, 2622. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.H.; Han, C.; Huang, H.; Xin, Y.Q.; Xu, Y.M.; Luo, L.; Yin, Z.M. Heat shock protein 70 together with its co-chaperone CHIP inhibits TNF-alpha induced apoptosis by promoting proteasomal degradation of apoptosis signal-regulating kinase1. Apoptosis 2010, 15, 822–833. [Google Scholar] [CrossRef]
- Daugaard, M.; Kirkegaard-Sorensen, T.; Ostenfeld, M.S.; Aaboe, M.; Hoyer-Hansen, M.; Orntoft, T.F.; Rohde, M.; Jaattela, M. Lens epithelium-derived growth factor is an Hsp70-2 regulated guardian of lysosomal stability in human cancer. Cancer Res. 2007, 67, 2559–2567. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Ye, R.; Jung, D.Y.; Barron, E.; Friedline, R.H.; Benoit, V.M.; Hinton, D.R.; Kim, J.K.; Lee, A.S. GRP78 plays an essential role in adipogenesis and postnatal growth in mice. FASEB J. 2013, 27, 955–964. [Google Scholar] [CrossRef] [Green Version]
- Smaldone, G.; Pirone, L.; Capolupo, A.; Vitagliano, L.; Monti, M.C.; Di Gaetano, S.; Pedone, E. The essential player in adipogenesis GRP78 is a novel KCTD15 interactor. Int. J. Biol. Macromol. 2018, 115, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Christodoulides, C.; Lagathu, C.; Sethi, J.K.; Vidal-Puig, A. Adipogenesis and WNT signalling. Trends Endocrinol. Metab. 2009, 20, 16–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, M.; Zhao, J.X.; Yan, X.; Huang, Y.; Nicodemus, L.V.; Yue, W.; McCormick, R.J.; Zhu, M.J. Fetal muscle development, mesenchymal multipotent cell differentiation, and associated signaling pathways. J. Anim. Sci. 2011, 89, 583–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farmer, S.R. Regulation of PPARgamma activity during adipogenesis. Int. J. Obes. 2005, 29 (Suppl. S1), S13–S16. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.H.; Huang, Q.Y.; Qian, X.L.; Duan, J.; Jiao, X.Q.; Wu, L.Y.; Huang, Q.Y.; Li, J.; Lai, X.N.; Shi, Y.B.; et al. Cdc42 Promotes ADSC-Derived IPC Induction, Proliferation, and Insulin Secretion Via Wnt/beta-Catenin Signaling. Diabetes Metab. Syndr. 2019, 12, 2325–2339. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Y.; Daniels, G.; Wang, J.; Zhang, X. TBL1XR1 in physiological and pathological states. Am. J. Clin. Exp. Urol. 2015, 3, 13–23. [Google Scholar]
- Piras, C.; Morittu, V.M.; Spina, A.A.; Soggiu, A.; Greco, V.; Rame, C.; Briant, E.; Mellouk, N.; Tilocca, B.; Bonizzi, L.; et al. Unraveling the Adipose Tissue Proteome of Transition Cows through Severe Negative Energy Balance. Animals 2019, 9, 1013. [Google Scholar] [CrossRef] [Green Version]
- Zachut, M.; Kra, G.; Livshitz, L.; Portnick, Y.; Yakoby, S.; Friedlander, G.; Levin, Y. Seasonal heat stress affects adipose tissue proteome toward enrichment of the Nrf2-mediated oxidative stress response in late-pregnant dairy cows. J. Proteom. 2017, 158, 52–61. [Google Scholar] [CrossRef]
- Maina, F.; Casagranda, F.; Audero, E.; Simeone, A.; Comoglio, P.M.; Klein, R.; Ponzetto, C. Uncoupling of Grb2 from the Met receptor in vivo reveals complex roles in muscle development. Cell 1996, 87, 531–542. [Google Scholar] [CrossRef] [Green Version]
- Woldt, E.; Matz, R.L.; Terrand, J.; Mlih, M.; Gracia, C.; Foppolo, S.; Martin, S.; Bruban, V.; Ji, J.; Velot, E.; et al. Differential Signaling by Adaptor Molecules LRP1 and ShcA Regulates Adipogenesis by the Insulin-like Growth Factor-1 Receptor. J. Biol. Chem. 2011, 286, 16775–16782. [Google Scholar] [CrossRef] [Green Version]
- Arita, Y.; Kihara, S.; Ouchi, N.; Maeda, K.; Kuriyama, H.; Okamoto, Y.; Kumada, M.; Hotta, K.; Nishida, M.; Takahashi, M.; et al. Adipocyte-derived plasma protein adiponectin acts as a platelet-derived growth factor-BB-binding protein and regulates growth factor-induced common postreceptor signal in vascular smooth muscle cell. Circulation 2002, 105, 2893–2898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, A.W.; Barradas, M.; Stone, J.C.; van Aelst, L.; Serrano, M.; Lowe, S.W. Premature senescence involving p53 and p16 is activated in response to constitutive MEK/MAPK mitogenic signaling. Genes Dev. 1998, 12, 3008–3019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, K.Y.; Wang, H.H.; Lai, S.T.; Pan, J.P.; Chiang, A.N. beta(2)-glycoprotein I protects J774A.1 macrophages and human coronary artery smooth muscle cells against apoptosis. J. Cell Biochem. 2005, 94, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Al-Fakhri, N.; Chavakis, T.; Schmidt-Woll, T.; Huang, B.; Cherian, S.M.; Bobryshev, Y.V.; Lord, R.S.; Katz, N.; Preissner, K.T. Induction of apoptosis in vascular cells by plasminogen activator inhibitor-1 and high molecular weight kininogen correlates with their anti-adhesive properties. Biol. Chem. 2003, 384, 423–435. [Google Scholar] [CrossRef]
- Bruneel, A.; Labas, V.; Mailloux, A.; Sharma, S.; Royer, N.; Vinh, J.; Pernet, P.; Vaubourdolle, M.; Baudin, B. Proteomics of human umbilical vein endothelial cells applied to etoposide-induced apoptosis. Proteomics 2005, 5, 3876–3884. [Google Scholar] [CrossRef]
- Helblingleclerc, A.; Zhang, X.; Topaloglu, H.; Cruaud, C.; Tesson, F.; Weissenbach, J.; Tome, F.M.S.; Schwartz, K.; Fardeau, M.; Tryggvason, K.; et al. Mutations in the Laminin Alpha-2-Chain Gene (Lama2) Cause Merosin-Deficient Congenital Muscular-Dystrophy. Nat. Genet. 1995, 11, 216–218. [Google Scholar] [CrossRef]
- Rossignol, P.; Ho-Tin-Noe, B.; Vranckx, R.; Bouton, M.C.; Meilhac, O.; Lijnen, H.R.; Guillin, M.C.; Michel, J.B.; Angles-Cano, E. Protease nexin-1 inhibits plasminogen activation-induced apoptosis of adherent cells. J. Biol. Chem. 2004, 279, 10346–10356. [Google Scholar] [CrossRef] [Green Version]
- Saleh, J.; Summers, L.K.; Cianflone, K.; Fielding, B.A.; Sniderman, A.D.; Frayn, K.N. Coordinated release of acylation stimulating protein (ASP) and triacylglycerol clearance by human adipose tissue in vivo in the postprandial period. J. Lipid Res. 1998, 39, 884–891. [Google Scholar]
- Buraschi, S.; Neill, T.; Goyal, A.; Poluzzi, C.; Smythies, J.; Owens, R.T.; Schaefer, L.; Torres, A.; Iozzo, R.V. Decorin causes autophagy in endothelial cells via Peg3. Proc. Natl. Acad. Sci. USA 2013, 110, E2582–E2591. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, A.; Nieves, E.; Che, F.Y.; Wang, J.; Jin, L.; Murray, J.W.; Gordon, K.; Angeletti, R.H.; Wolkoff, A.W. Proteomic analysis of endocytic vesicles: Rab1a regulates motility of early endocytic vesicles. J. Cell Sci. 2011, 124, 765–775. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Alemany, R.; Suelves, M.; Diaz-Ramos, A.; Vidal, B.; Munoz-Canoves, P. Alpha-enolase plasminogen receptor in myogenesis. Front. Biosci. 2005, 10, 30–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kepser, L.J.; Damar, F.; De Cicco, T.; Chaponnier, C.; Proszynski, T.J.; Pagenstecher, A.; Rust, M.B. CAP2 deficiency delays myofibril actin cytoskeleton differentiation and disturbs skeletal muscle architecture and function. Proc. Natl. Acad. Sci. USA 2019, 116, 8397–8402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velez, E.J.; Azizi, S.; Verheyden, D.; Salmeron, C.; Lutfi, E.; Sanchez-Moya, A.; Navarro, I.; Gutierrez, J.; Capilla, E. Proteolytic systems’ expression during myogenesis and transcriptional regulation by amino acids in gilthead sea bream cultured muscle cells. PLoS ONE 2017, 12, e0187339. [Google Scholar] [CrossRef] [PubMed]
- Ebisui, C.; Tsujinaka, T.; Morimoto, T.; Fujita, J.; Ogawa, A.; Ishidoh, K.; Kominami, E.; Tanaka, K.; Monden, M. Changes of Proteasomes and Cathepsins Activities and Their Expression during Differentiation of C2C12 Myoblasts. J. Biochem. 1995, 117, 1088–1094. [Google Scholar] [CrossRef]
- Moreno-Fortuny, A.; Bragg, L.; Cossu, G.; Roostalu, U. MCAM contributes to the establishment of cell autonomous polarity in myogenic and chondrogenic differentiation. Biol. Open 2017, 6, 1592–1601. [Google Scholar] [CrossRef] [Green Version]
- Baghdadi, M.B.; Castel, D.; Machado, L.; Fukada, S.; Birk, D.E.; Relaix, F.; Tajbakhsh, S.; Mourikis, P. Reciprocal signalling by Notch-Collagen V-CALCR retains muscle stem cells in their niche. Nature 2018, 557, 714–718. [Google Scholar] [CrossRef]
- Santos-Zas, I.; Lodeiro, M.; Gurriaran-Rodriguez, U.; Bouzo-Lorenzo, M.; Mosteiro, C.S.; Casanueva, F.F.; Casabiell, X.; Pazos, Y.; Camina, J.P. beta-Arrestin signal complex plays a critical role in adipose differentiation. Int. J. Biochem. Cell Biol. 2013, 45, 1281–1292. [Google Scholar] [CrossRef]
- Wang, H.C.; Ko, Y.H.; Mersmann, H.J.; Chen, C.L.; Ding, S.T. The expression of genes related to adipocyte differentiation in pigs. J. Anim. Sci. 2006, 84, 1059–1066. [Google Scholar] [CrossRef]
- Titushkin, I.; Sun, S.; Paul, A.; Cho, M. Control of adipogenesis by ezrin, radixin and moesin-dependent biomechanics remodeling. J. Biomech. 2013, 46, 521–526. [Google Scholar] [CrossRef]
- Wang, C.; Zeng, X.; Zhou, Z.; Zhao, J.; Pei, G. β-arrestin-1 contributes to brown fat function and directly interacts with PPARα and PPARγ. Sci. Rep. 2016, 6, 26999. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.Y.; Li, Y.; Teng, W.D.; Du, M.; Li, Y.X.; Sun, B.G. Liensinine Inhibits Beige Adipocytes Recovering to white Adipocytes through Blocking Mitophagy Flux in vitro and in vivo. Nutrients 2019, 11, 1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Greevenbroek, M.M.J. The expanding role of complement in adipose tissue metabolism and lipoprotein function. Curr. Opin. Lipidol. 2009, 20, 353–354. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, P.G.; Lane, W.S.; Bennett, V.; Lienhard, G.E. Association of protein kinase C(lambda) with adducin in 3T3-L1 adipocytes. Biochim. Biophys. Acta 2001, 1539, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Saavedra, P.; Girona, J.; Bosquet, A.; Guaita, S.; Canela, N.; Aragones, G.; Heras, M.; Masana, L. New insights into circulating FABP4: Interaction with cytokeratin 1 on endothelial cell membranes. BBA Mol. Cell Res. 2015, 1853, 2966–2974. [Google Scholar] [CrossRef] [Green Version]
- Reed, S.E.; Hodgson, L.R.; Song, S.; May, M.T.; Kelly, E.E.; McCaffrey, M.W.; Mastick, C.C.; Verkade, P.; Tavare, J.M. A role for Rab14 in the endocytic trafficking of GLUT4 in 3T3-L1 adipocytes. J. Cell Sci. 2013, 126, 1931–1941. [Google Scholar] [CrossRef] [Green Version]
- Lawson, H.A.; Zayed, M.; Wayhart, J.P.; Fabbrini, E.; Love-Gregory, L.; Klein, S.; Semenkovich, C.F. Physiologic and genetic evidence links hemopexin to triglycerides in mice and humans. Int. J. Obes. 2017, 41, 631–638. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, J.M.; Camacho, L.E.; Ebarb, S.M.; Swanson, K.C.; Vonnahme, K.A.; Stelzleni, A.M.; Johnson, S.E. Realimentation of nutrient restricted pregnant beef cows supports compensatory fetal muscle growth. J. Anim. Sci. 2013, 91, 4797–4806. [Google Scholar] [CrossRef]
- Bouley, J.; Chambon, C.; Picard, B. Mapping of bovine skeletal muscle proteins using two-dimensional gel electrophoresis and mass spectrometry. Proteomics 2004, 4, 1811–1824. [Google Scholar] [CrossRef]
- Kaspric, N.; Reichstadt, M.; Picard, B.; Tournayre, J.; Bonnet, M. Protein Function Easily Investigated by Genomics Data Mining Using the ProteINSIDE Online Tool. Genom. Comput. Biol. 2015, 1, e16. [Google Scholar] [CrossRef] [Green Version]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef] [Green Version]
- Caccia, D.; Dugo, M.; Callari, M.; Bongarzone, I. Bioinformatics tools for secretome analysis. Biochim. Biophys. Acta 2013, 1834, 2442–2453. [Google Scholar] [CrossRef]
- UniProt, C. UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar]
- Orchard, S.; Ammari, M.; Aranda, B.; Breuza, L.; Briganti, L.; Broackes-Carter, F.; Campbell, N.H.; Chavali, G.; Chen, C.; del-Toro, N.; et al. The MIntAct project--IntAct as a common curation platform for 11 molecular interaction databases. Nucleic Acids Res. 2014, 42, D358–D363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatr-Aryamontri, A.; Breitkreutz, B.J.; Oughtred, R.; Boucher, L.; Heinicke, S.; Chen, D.; Stark, C.; Breitkreutz, A.; Kolas, N.; O’Donnell, L.; et al. The BioGRID interaction database: 2015 update. Nucleic Acids Res. 2015, 43, D470–D478. [Google Scholar] [CrossRef] [PubMed]
- Orchard, S.; Kerrien, S.; Abbani, S.; Aranda, B.; Bhate, J.; Bidwell, S.; Bridge, A.; Briganti, L.; Brinkman, F.S.; Cesareni, G.; et al. Protein interaction data curation: The International Molecular Exchange (IMEx) consortium. Nat. Methods 2012, 9, 345–350. [Google Scholar] [CrossRef]
- Orchard, S.; Salwinski, L.; Kerrien, S.; Montecchi-Palazzi, L.; Oesterheld, M.; Stumpflen, V.; Ceol, A.; Chatr-aryamontri, A.; Armstrong, J.; Woollard, P.; et al. The minimum information required for reporting a molecular interaction experiment (MIMIx). Nat. Biotechnol. 2007, 25, 894–898. [Google Scholar] [CrossRef]
- Petryszak, R.; Burdett, T.; Fiorelli, B.; Fonseca, N.A.; Gonzalez-Porta, M.; Hastings, E.; Huber, W.; Jupp, S.; Keays, M.; Kryvych, N.; et al. Expression Atlas update--a database of gene and transcript expression from microarray- and sequencing-based functional genomics experiments. Nucleic Acids Res. 2014, 42, D926–D932. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adipose Tissue n = 39 | Muscle Tissue n = 9 | Adipose and Muscle Tissues n = 29 |
|---|---|---|
| A2M | LTF | ADIPOQ |
| AFM | CASQ1 | AFP |
| APOA2 | DPT | AHSG |
| C3 | P4HA3 | AMBP |
| CFI | LAMA2 | APOA1 |
| COL1A2 | C1QTNF3 | APOH |
| COL6A2 | DSG1 | CLEC3B |
| COLGALT1 | P4HA2 | CNPY3 |
| CPB2 | SRL | COL5A1 |
| DCN | COL6A1 | |
| DDOST | CTSB | |
| DNAJB11 | ERLIN2 | |
| ERP29 | FKBP7 | |
| FETUB | HPX | |
| FGG | HSPA5 | |
| FKBP10 | KNG1 | |
| FKBP14 | LUM | |
| GC | MESD | |
| GGH | OGN | |
| GLB1 | OLFML3 | |
| GPX3 | P4HA1 | |
| HSP90B1 | P4HB | |
| ITIH3 | PDIA3 | |
| MFAP4 | POSTN | |
| ORM1 | SERPINA1 | |
| PCOLCE | SERPINF1 | |
| PDIA4 | THBS1 | |
| PLG | TPP1 | |
| PLOD3 | TTR | |
| PRCP | ||
| RCN3 | ||
| RPN1 | ||
| SERPINA3 | ||
| SERPINC1 | ||
| SERPIND1 | ||
| SERPINF2 | ||
| SERPING1 | ||
| SERPINH1 | ||
| TGFBR3 |
| Adipose Tissue n = 12 | Muscle Tissue n = 2 | Adipose and Muscle Tissues n = 8 |
|---|---|---|
| ANP32A | PACSIN3 | ANXA1 |
| APPL2 | SEPT4 | ANXA2 |
| ARRB1 | ANXA4 | |
| CKAP4 | ANXA7 | |
| CYB5A | PARK7 | |
| EMC2 | SNX5 | |
| HNRNPA2B1 | TPT1 | |
| LAMTOR1 | VCP | |
| RAB14 | ||
| SPET5 | ||
| STAM | ||
| TKT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonnet, M.; Kaspric, N.; Vonnahme, K.; Viala, D.; Chambon, C.; Picard, B. Prediction of the Secretome and the Surfaceome: A Strategy to Decipher the Crosstalk between Adipose Tissue and Muscle during Fetal Growth. Int. J. Mol. Sci. 2020, 21, 4375. https://doi.org/10.3390/ijms21124375
Bonnet M, Kaspric N, Vonnahme K, Viala D, Chambon C, Picard B. Prediction of the Secretome and the Surfaceome: A Strategy to Decipher the Crosstalk between Adipose Tissue and Muscle during Fetal Growth. International Journal of Molecular Sciences. 2020; 21(12):4375. https://doi.org/10.3390/ijms21124375
Chicago/Turabian StyleBonnet, Muriel, Nicolas Kaspric, Kimberly Vonnahme, Didier Viala, Christophe Chambon, and Brigitte Picard. 2020. "Prediction of the Secretome and the Surfaceome: A Strategy to Decipher the Crosstalk between Adipose Tissue and Muscle during Fetal Growth" International Journal of Molecular Sciences 21, no. 12: 4375. https://doi.org/10.3390/ijms21124375
APA StyleBonnet, M., Kaspric, N., Vonnahme, K., Viala, D., Chambon, C., & Picard, B. (2020). Prediction of the Secretome and the Surfaceome: A Strategy to Decipher the Crosstalk between Adipose Tissue and Muscle during Fetal Growth. International Journal of Molecular Sciences, 21(12), 4375. https://doi.org/10.3390/ijms21124375