Connexin Hemichannels Contribute to the Activation of cAMP Signaling Pathway and Renin Production

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
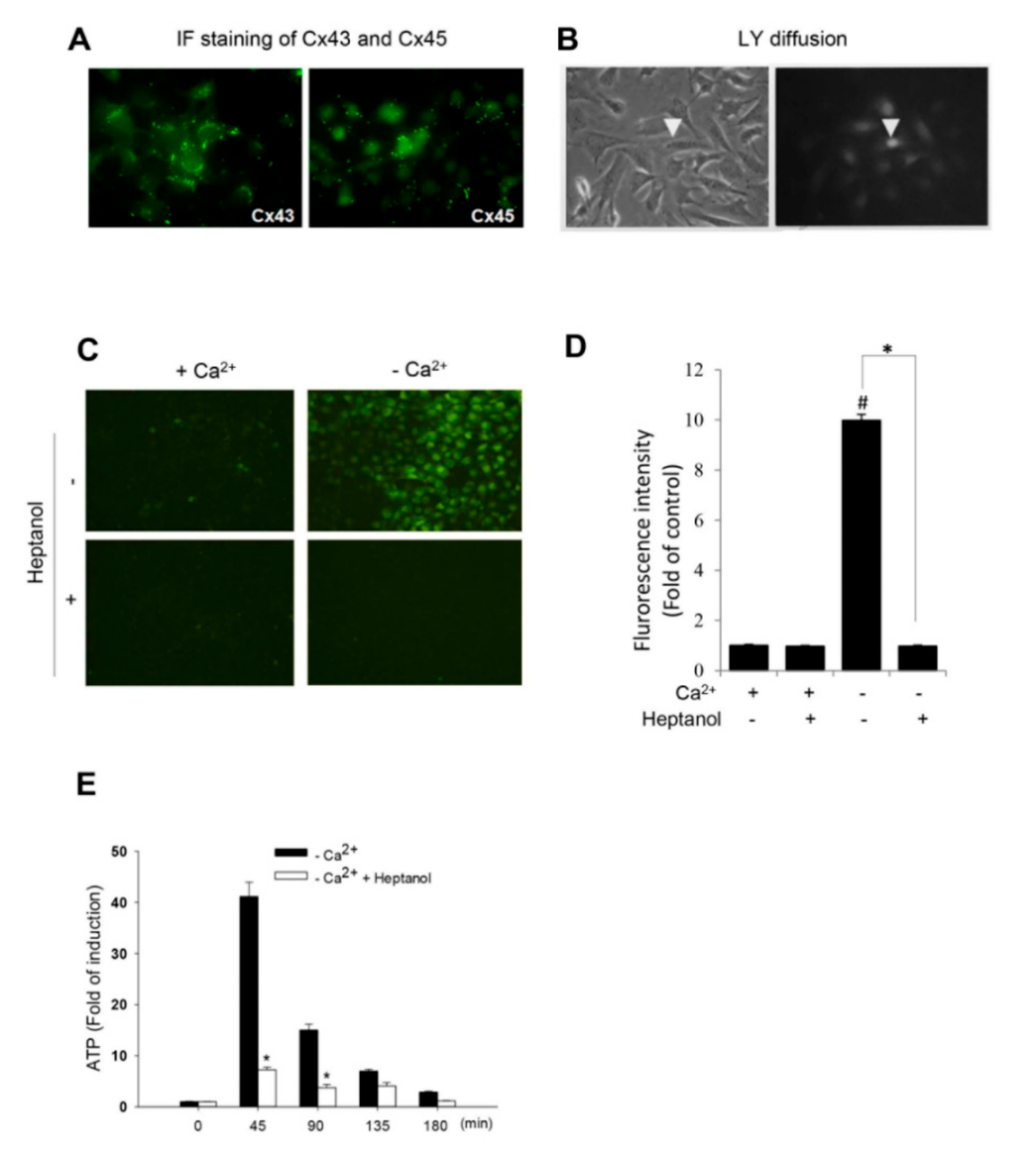
2.1. Removing Extracellular Ca2+ Activates Hemichannels in As4.1 Cells
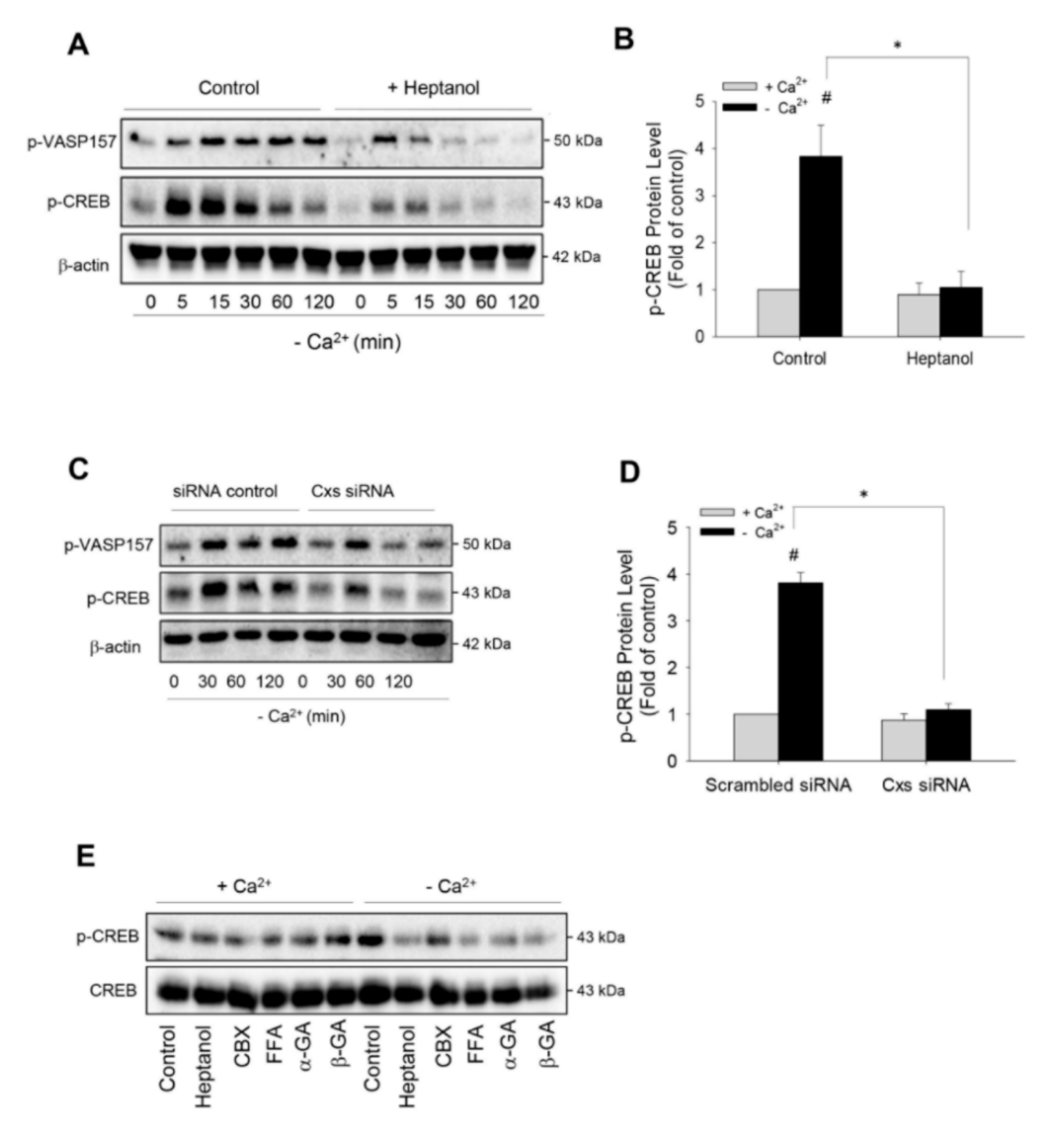
2.2. Hemichannels Mediate Low Ca2+-Triggered Activation of cAMP Signaling Pathway
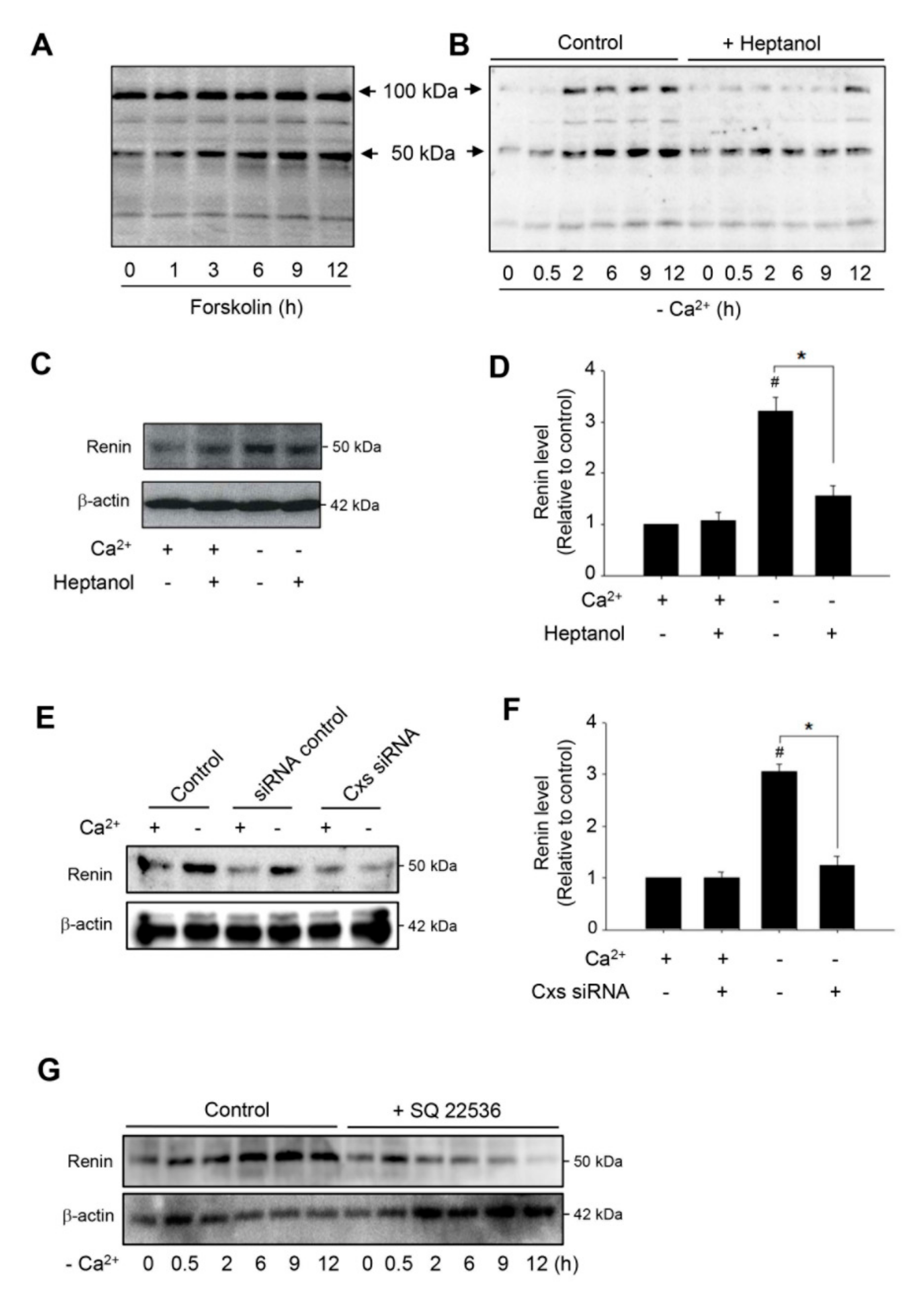
2.3. Low Ca2+-Induced Activation of cAMP Signaling is Mediated by Adenylyl Cyclase
2.4. Hemichannels Contribute to Low Ca2+-Elicited Expression of Renin
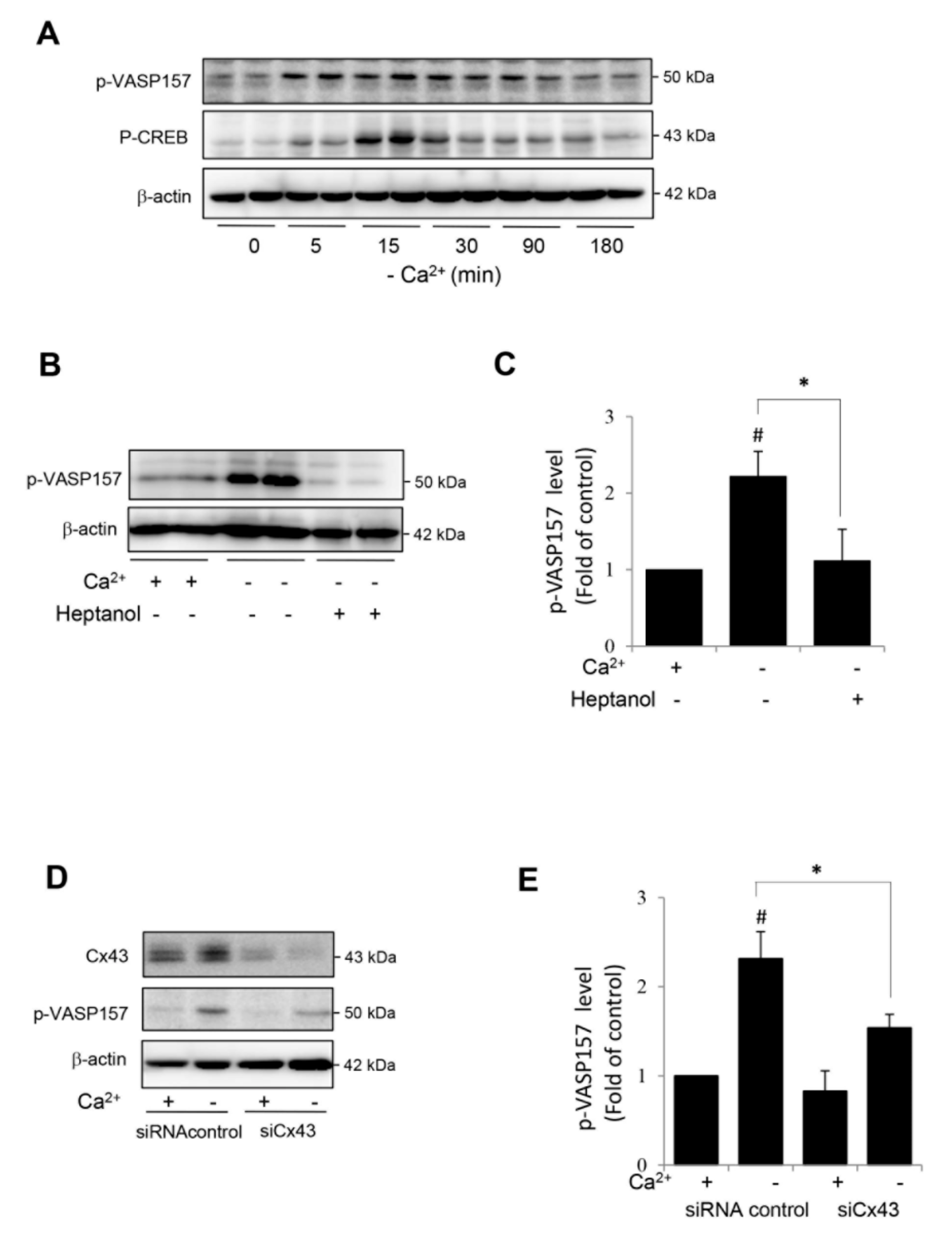
2.5. Hemichannels Mediate Low Ca2+-Elicited Activation of cAMP Signaling Pathway in Rat Renal Tubular Epithelial Cell NRK
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. Immunocytochemical Analysis
4.4. Evaluation of GJIC by Microinjection of LY
4.5. Dye Uptake Assay
4.6. ATP Measurement
4.7. Western Blot Analysis
4.8. Transient Transfection of As4.1 Cells with siRNA
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ATP | adenosine triphosphate |
| Ca2+ | calcium |
| CREB | cAMP response element binding protein |
| Cx | connexin |
| GJ | gap junction |
| JG | juxtaglomerular |
| NAD+ | nicotinamide adenine dinucleotide |
| NRK | normal rat kidney |
| PGE2 | prostaglandin E2 |
| PKA | protein kinase A |
| LY | lucifer yellow |
| VASP | vasodilator-stimulated phosphoprotein |
References
- Ortiz-Capisano, M.C.; Ortiz, P.A.; Harding, P.; Garvin, J.L.; Beierwaltes, W.H. Adenylyl cyclase isoform v mediates renin release from juxtaglomerular cells. Hypertension 2007, 49, 618–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz-Capisano, M.C.; Ortiz, P.A.; Harding, P.; Garvin, J.L.; Beierwaltes, W.H. Decreased intracellular calcium stimulates renin release via calcium-inhibitable adenylyl cyclase. Hypertension 2007, 49, 162–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atchison, D.K.; Ortiz-Capisano, M.C.; Beierwaltes, W.H. Acute activation of the calcium-sensing receptor inhibits plasma renin activity in vivo. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R1020–R1026. [Google Scholar] [CrossRef] [Green Version]
- Klar, J.; Sigl, M.; Obermayer, B.; Schweda, F.; Kramer, B.K.; Kurtz, A. Calcium inhibits renin gene expression by transcriptional and posttranscriptional mechanisms. Hypertension 2005, 46, 1340–1346. [Google Scholar] [CrossRef] [Green Version]
- Grunberger, C.; Obermayer, B.; Klar, J.; Kurtz, A.; Schweda, F. The calcium paradoxon of renin release: Calcium suppresses renin exocytosis by inhibition of calcium-dependent adenylate cyclases AC5 and AC6. Circ. Res. 2006, 99, 1197–1206. [Google Scholar] [CrossRef] [Green Version]
- Schweda, F.; Kurtz, A. Cellular mechanism of renin release. Acta Physiol. Scand. 2004, 181, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Suwa, M.; Li, B.; Kawamura, K.; Morioka, T.; Oite, T. ATP-dependent mechanism for coordination of intercellular Ca2+ signaling and renin secretion in rat juxtaglomerular cells. Circ. Res. 2003, 93, 338–345. [Google Scholar] [CrossRef] [Green Version]
- Beierwaltes, W.H. The role of calcium in the regulation of renin secretion. Am. J. Physiol. Ren. Physiol. 2010, 298, F1–F11. [Google Scholar] [CrossRef] [Green Version]
- Klar, J.; Sandner, P.; Muller, M.W.; Kurtz, A. Cyclic AMP stimulates renin gene transcription in juxtaglomerular cells. Pflug. Arch. 2002, 444, 335–344. [Google Scholar] [CrossRef]
- Schweda, F.; Klar, J.; Narumiya, S.; Nusing, R.M.; Kurtz, A. Stimulation of renin release by prostaglandin E2 is mediated by EP2 and EP4 receptors in mouse kidneys. Am. J. Physiol. Ren. Physiol. 2004, 287, F427–F433. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, A.A.; Salinas-Parra, N.; Leach, D.; Navar, L.G.; Prieto, M.C. PGE2 upregulates renin through E-prostanoid receptor 1 via PKC/cAMP/CREB pathway in M-1 cells. Am. J. Physiol. Ren. Physiol. 2017, 313, F1038–F1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackermann, M.; Ritthaler, T.; Riegger, G.; Kurtz, A.; Kramer, B.K. Endothelin inhibits cAMP-induced renin release from isolated renal juxtaglomerular cells. J. Cardiovasc. Pharm. 1995, 26, S135–S137. [Google Scholar] [CrossRef]
- Lu, M.; Liu, Y.H.; Goh, H.S.; Wang, J.J.; Yong, Q.C.; Wang, R.; Bian, J.S. Hydrogen sulfide inhibits plasma renin activity. J. Am. Soc. Nephrol. 2010, 21, 993–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.; Zhu, Y.; Morioka, T.; Oite, T.; Kitamura, M. Pathophysiological roles of gap junction in glomerular mesangial cells. J. Membr. Biol. 2007, 217, 123–130. [Google Scholar] [CrossRef]
- Yao, J.; Oite, T.; Kitamura, M. Gap junctional intercellular communication in the juxtaglomerular apparatus. Am. J. Physiol. Ren. Physiol. 2009, 296, F939–F946. [Google Scholar] [CrossRef] [Green Version]
- Bennett, M.V.; Contreras, J.E.; Bukauskas, F.F.; Saez, J.C. New roles for astrocytes: Gap junction hemichannels have something to communicate. Trends Neurosci. 2003, 26, 610–617. [Google Scholar] [CrossRef] [Green Version]
- Wagner, C. Function of connexins in the renal circulation. Kidney Int. 2008, 73, 547–555. [Google Scholar] [CrossRef] [Green Version]
- Peti-Peterdi, J. Calcium wave of tubuloglomerular feedback. Am. J. Physiol. Ren. Physiol. 2006, 291, F473–F480. [Google Scholar] [CrossRef] [Green Version]
- Wagner, C.; de Wit, C.; Kurtz, L.; Grunberger, C.; Kurtz, A.; Schweda, F. Connexin40 is essential for the pressure control of renin synthesis and secretion. Circ. Res. 2007, 100, 556–563. [Google Scholar] [CrossRef]
- Schweda, F.; Kurtz, L.; de Wit, C.; Janssen-Bienhold, U.; Kurtz, A.; Wagner, C. Substitution of connexin40 with connexin45 prevents hyperreninemia and attenuates hypertension. Kidney Int. 2009, 75, 482–489. [Google Scholar] [CrossRef] [Green Version]
- Lubkemeier, I.; Machura, K.; Kurtz, L.; Neubauer, B.; Dobrowolski, R.; Schweda, F.; Wagner, C.; Willecke, K.; Kurtz, A. The connexin 40 A96S mutation causes renin-dependent hypertension. J. Am. Soc. Nephrol. 2011, 22, 1031–1040. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Huang, T.; Zhu, Y.; Yan, Q.; Chi, Y.; Jiang, J.X.; Wang, P.; Matsue, H.; Kitamura, M.; Yao, J. Connexin43 hemichannels contribute to cadmium-induced oxidative stress and cell injury. Antioxid. Redox Signal. 2011, 14, 2427–2439. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Chi, Y.; Gao, K.; Yan, Q.; Matsue, H.; Takeda, M.; Kitamura, M.; Yao, J. Connexin43 hemichannel-mediated regulation of connexin43. PLoS ONE 2013, 8, e58057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siller-Jackson, A.J.; Burra, S.; Gu, S.; Xia, X.; Bonewald, L.F.; Sprague, E.; Jiang, J.X. Adaptation of connexin 43-hemichannel prostaglandin release to mechanical loading. J. Biol. Chem. 2008, 283, 26374–26382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burra, S.; Jiang, J.X. Regulation of cellular function by connexin hemichannels. Int. J. Biochem. Mol. Biol. 2011, 2, 119. [Google Scholar] [PubMed]
- Jiang, L.; Foster, F.M.; Ward, P.; Tasevski, V.; Luttrell, B.M.; Conigrave, A.D. Extracellular ATP triggers cyclic AMP-dependent differentiation of HL-60 cells. Biochem. Biophys. Res. Commun. 1997, 232, 626–630. [Google Scholar] [CrossRef] [PubMed]
- Stout, C.E.; Costantin, J.L.; Naus, C.C.; Charles, A.C. Intercellular calcium signaling in astrocytes via ATP release through connexin hemichannels. J. Biol. Chem. 2002, 277, 10482–10488. [Google Scholar] [CrossRef] [Green Version]
- Stridh, M.H.; Tranberg, M.; Weber, S.G.; Blomstrand, F.; Sandberg, M. Stimulated efflux of amino acids and glutathione from cultured hippocampal slices by omission of extracellular calcium: Likely involvement of connexin hemichannels. J. Biol. Chem. 2008, 283, 10347–10356. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.; Zhang, X.; Mitsui, T.; Kamiyama, M.; Takeda, M.; Yao, J. Connexin43 hemichannels contributes to the disassembly of cell junctions through modulation of intracellular oxidative status. Redox Biol. 2016, 9, 198–209. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.; Gao, K.; Zhang, H.; Takeda, M.; Yao, J. Suppression of cell membrane permeability by suramin: Involvement of its inhibitory actions on connexin 43 hemichannels. Br. J. Pharm. 2014, 171, 3448–3462. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.; Gao, K.; Li, K.; Nakajima, S.; Kira, S.; Takeda, M.; Yao, J. Purinergic control of AMPK activation by ATP released through connexin 43 hemichannels–pivotal roles in hemichannel-mediated cell injury. J. Cell Sci. 2014, 127, 1487–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russ, U.; Rauch, U.; Quast, U. Pharmacological evidence for a KATP channel in renin-secreting cells from rat kidney. J. Physiol. 1999, 517, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.L.; Skott, O. Renin release from permeabilized juxtaglomerular cells is stimulated by chloride but not by low calcium. Am. J. Physiol. 1994, 266, F604–F611. [Google Scholar] [CrossRef] [PubMed]
- Friis, U.G.; Madsen, K.; Svenningsen, P.; Hansen, P.B.; Gulaveerasingam, A.; Jorgensen, F.; Aalkjaer, C.; Skott, O.; Jensen, B.L. Hypotonicity-induced Renin exocytosis from juxtaglomerular cells requires aquaporin-1 and cyclooxygenase-2. J. Am. Soc. Nephrol. 2009, 20, 2154–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, M.J.; Liu, B.; Herbowy, M.T.; Gross, K.W.; Hajduczok, G. Intercellular communication between renin expressing As4.1 cells, endothelial cells and smooth muscle cells. Life Sci. 2003, 72, 1289–1301. [Google Scholar] [CrossRef]
- Halbrugge, M.; Friedrich, C.; Eigenthaler, M.; Schanzenbacher, P.; Walter, U. Stoichiometric and reversible phosphorylation of a 46-kDa protein in human platelets in response to cGMP- and cAMP-elevating vasodilators. J. Biol. Chem. 1990, 265, 3088–3093. [Google Scholar]
- Cherian, P.P.; Siller-Jackson, A.J.; Gu, S.; Wang, X.; Bonewald, L.F.; Sprague, E.; Jiang, J.X. Mechanical strain opens connexin 43 hemichannels in osteocytes: A novel mechanism for the release of prostaglandin. Mol. Biol. Cell 2005, 16, 3100–3106. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.; Black, T.A.; Shi, Q.; Jones, C.A.; Petrovic, N.; Loudon, J.; Kane, C.; Sigmund, C.D.; Gross, K.W. Critical roles of a cyclic AMP responsive element and an E-box in regulation of mouse renin gene expression. J. Biol. Chem. 2001, 276, 45530–45538. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Huang, T.; Fang, X.; Chi, Y.; Zhu, Y.; Wan, Y.; Matsue, H.; Kitamura, M. Disruption of gap junctions attenuates aminoglycoside-elicited renal tubular cell injury. Br. J. Pharm. 2010, 160, 2055–2068. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Liu, Y.H.; Ho, C.Y.; Tiong, C.X.; Bian, J.S. Hydrogen sulfide regulates cAMP homeostasis and renin degranulation in As4.1 and rat renin-rich kidney cells. Am. J. Physiol. Cell Physiol. 2012, 302, C59–C66. [Google Scholar] [CrossRef] [Green Version]
- Puceat, M.; Bony, C.; Jaconi, M.; Vassort, G. Specific activation of adenylyl cyclase V by a purinergic agonist. FEBS Lett. 1998, 431, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Torres, B.; Zambon, A.C.; Insel, P.A. P2Y11 receptors activate adenylyl cyclase and contribute to nucleotide-promoted cAMP formation in MDCK-D(1) cells. A mechanism for nucleotide-mediated autocrine-paracrine regulation. J. Biol. Chem. 2002, 277, 7761–7765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwamoto, T.; Nakamura, T.; Doyle, A.; Ishikawa, M.; de Vega, S.; Fukumoto, S.; Yamada, Y. Pannexin 3 regulates intracellular ATP/cAMP levels and promotes chondrocyte differentiation. J. Biol. Chem. 2010, 285, 18948–18958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anselmi, F.; Hernandez, V.H.; Crispino, G.; Seydel, A.; Ortolano, S.; Roper, S.D.; Kessaris, N.; Richardson, W.; Rickheit, G.; Filippov, M.A.; et al. ATP release through connexin hemichannels and gap junction transfer of second messengers propagate Ca2+ signals across the inner ear. Proc. Natl. Acad. Sci. USA 2008, 105, 18770–18775. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, L.I. Connexin 43 hemichannels and intracellular signaling in bone cells. Front. Physiol. 2014, 5, 131. [Google Scholar] [CrossRef] [Green Version]
- Mitropoulou, G.; Bruzzone, R. Modulation of perch connexin35 hemi-channels by cyclic AMP requires a protein kinase A phosphorylation site. J. Neurosci. Res. 2003, 72, 147–157. [Google Scholar] [CrossRef]
- Retamal, M.A.; Yin, S.; Altenberg, G.A.; Reuss, L. Modulation of Cx46 hemichannels by nitric oxide. Am. J. Physiol. Cell Physiol. 2009, 296, C1356–C1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.; Hiramatsu, N.; Zhu, Y.; Morioka, T.; Takeda, M.; Oite, T.; Kitamura, M. Nitric oxide-mediated regulation of connexin43 expression and gap junctional intercellular communication in mesangial cells. J. Am. Soc. Nephrol. 2005, 16, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Spray, D.C. Hypertension in connexin40-null mice: A renin disorder. Kidney Int. 2007, 72, 781–782. [Google Scholar] [CrossRef] [Green Version]
- Gomez, R.A.; Sequeira Lopez, M.L. Who and where is the renal baroreceptor?: The connexin hypothesis. Kidney Int. 2009, 75, 460–462. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, J.; Yao, J. Connexin Hemichannels Contribute to the Activation of cAMP Signaling Pathway and Renin Production. Int. J. Mol. Sci. 2020, 21, 4462. https://doi.org/10.3390/ijms21124462
Hong J, Yao J. Connexin Hemichannels Contribute to the Activation of cAMP Signaling Pathway and Renin Production. International Journal of Molecular Sciences. 2020; 21(12):4462. https://doi.org/10.3390/ijms21124462
Chicago/Turabian StyleHong, Jingru, and Jian Yao. 2020. "Connexin Hemichannels Contribute to the Activation of cAMP Signaling Pathway and Renin Production" International Journal of Molecular Sciences 21, no. 12: 4462. https://doi.org/10.3390/ijms21124462
APA StyleHong, J., & Yao, J. (2020). Connexin Hemichannels Contribute to the Activation of cAMP Signaling Pathway and Renin Production. International Journal of Molecular Sciences, 21(12), 4462. https://doi.org/10.3390/ijms21124462