An Updated Review of Pro- and Anti-Inflammatory Properties of Plasma Lysophosphatidylcholines in the Vascular System



Abstract
:1. Introduction
2. The Complex Role of LPC in Vascular Inflammation
2.1. Postulated Pro-Inflammatory Action of LPC on Vascular Reactivity
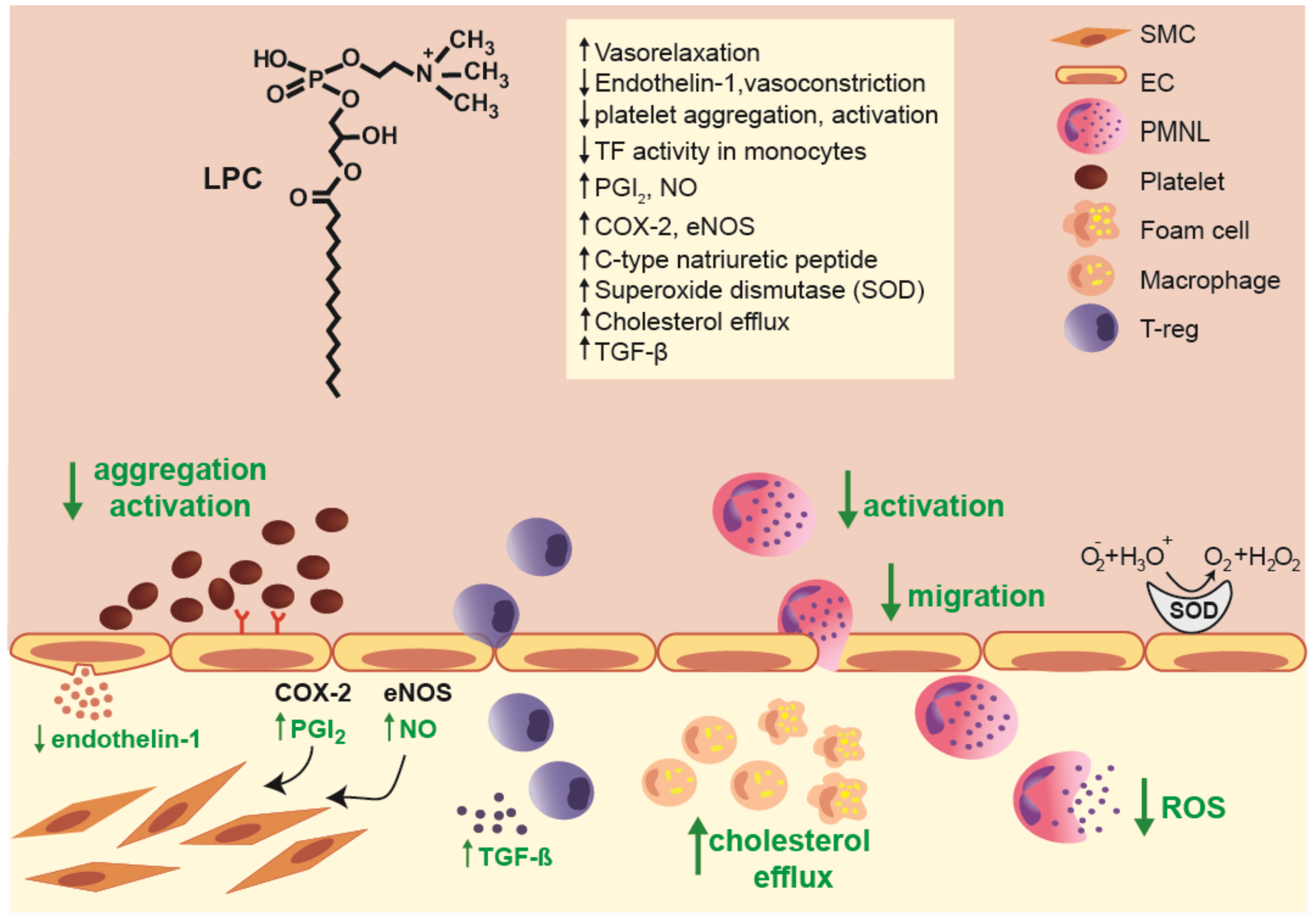
2.2. Postulated Anti-Inflammatory Action of LPCs on Vascular Reactivity
2.3. Investigating the Effects of LPCs on Immune Cells Involved in Vascular Inflammation
2.3.1. Effects of LPCs on Innate Immune Cells
2.3.2. The Proposed Roles of LPC on the Adaptive Immune System
2.4. Additional Anti-Inflammatory Effects of LPC in Vascular Inflammation and Atherosclerosis Development
3. Future Directions of LPC as a Biomarker
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| eNOS | Endothelial Nitric Oxide Synthase |
| HDL | High-Density Lipoprotein |
| LPA | Lysophosphatidic Acid |
| LPC | lysophosphatidylcholine |
| Ox-LDL | Oxidized Low-Density Lipoprotein |
| sPLA2 | Secreted Phospholipase A2 |
References
- Kuzell, W.C.; Davison, R.A. Serum lysolecithin in rheumatoid arthritis, pregnancy, and jaundice and in normal persons. J. Lab. Clin. Med. 1946, 31, 1223–1226. [Google Scholar] [PubMed]
- Marinetti, G.V.; Erbland, J.; Temple, K.; Stotz, E. Hydrolysis of lecithins by venom phospholipase A. I. Structure of the enzymically formed lysolecithins. Biochim. Biophys. Acta 1960, 38, 524–534. [Google Scholar] [CrossRef]
- Sun, C.; Hanasaka, A.; Kashiwagi, H.; Ueno, M. Formation and characterization of phosphatidylethanolamine/lysophosphatidylcholine mixed vesicles. Biochim. Biophys. Acta—Biomembr. 2000, 1467, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Mcmahon, H.T.; Boucrot, E. Membrane curvature at a glance. J. Cell Sci. 2015, 128, 1065–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reman, F.C.; Van Deenen, L.L.M. The action of some synthetic lysolecithins and lecithins on erythrocytes and lipid bilayers. Biochim. Biophys. Acta—Lipids Lipid Metab. 1967, 137, 592–594. [Google Scholar] [CrossRef] [Green Version]
- Flavahan, N.A. Lysophosphatidylcholine modifies G protein-dependent signaling in porcine endothelial cells. Am. J. Physiol. Circ. Physiol. 1993, 264, H722–H727. [Google Scholar] [CrossRef]
- Okajima, F.; Sato, K.; Tomura, H.; Kuwabara, A.; Nochi, H.; Tamoto, K.; Kondo, Y.; Tokumitsu, Y.; Ui, M. Stimulatory and inhibitory actions of lysophosphatidylcholine, depending on its fatty acid residue, on the phospholipase C/Ca2+ system in HL-60 leukaemia cells. Biochem. J. 1998, 336, 491–500. [Google Scholar] [CrossRef] [Green Version]
- Soga, T.; Ohishi, T.; Matsui, T.; Saito, T.; Matsumoto, M.; Takasaki, J.; Matsumoto, S.; Kamohara, M.; Hiyama, H.; Yoshida, S.; et al. Lysophosphatidylcholine enhances glucose-dependent insulin secretion via an orphan G-protein-coupled receptor. Biochem. Biophys. Res. Commun. 2005, 326, 744–751. [Google Scholar] [CrossRef]
- Drzazga, A.; Kristinsson, H.; Sałaga, M.; Zatorski, H.; Koziołkiewicz, M.; Gendaszewska-Darmach, E.; Bergsten, P. Lysophosphatidylcholine and its phosphorothioate analogues potentiate insulin secretion via GPR40 (FFAR1), GPR55 and GPR119 receptors in a different manner. Mol. Cell. Endocrinol. 2018, 472, 117–125. [Google Scholar] [CrossRef]
- Drzazga, A.; Sowinska, A.; Krzeminska, A.; Rytczak, P.; Koziolkiewicz, M.; Gendaszewska-Darmach, E. Lysophosphatidylcholine elicits intracellular calcium signaling in a GPR55-dependent manner. Biochem. Biophys. Res. Commun. 2017, 489, 242–247. [Google Scholar] [CrossRef]
- Zhu, K.; Baudhuin, L.M.; Hong, G.; Williams, F.S.; Cristina, K.L.; Kabarowski, J.H.S.; Witte, O.N.; Xu, Y. Sphingosylphosphorylcholine and Lysophosphatidylcholine Are Ligands for the G Protein-coupled Receptor GPR4. J. Biol. Chem. 2001, 276, 41325–41335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabarowski, J.H.S.; Zhu, K.; Le, L.Q.; Witte, O.N.; Xu, Y. Lysophosphatidylcholine as a Ligand for the Immunoregulatory Receptor G2A. Science 2001, 293, 702–705. [Google Scholar] [CrossRef]
- Witte, O.N.; Kabarowski, J.H.; Xu, Y.; Le, L.Q.; Zhu, K. Retraction. Science 2005, 307, 206b. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, M.-G.; Vanek, M.; Guerini, D.; Gasser, J.A.; Jones, C.E.; Junker, U.; Hofstetter, H.; Wolf, R.M.; Seuwen, K. Proton-sensing G-protein-coupled receptors. Nature 2003, 425, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Sphingosylphosphorylcholine and lysophosphatidylcholine are ligands for the G protein-coupled receptor GPR4. J. Biol. Chem. 2005, 280, 43280.
- Bektas, M.; Barak, L.S.; Jolly, P.S.; Liu, H.; Lynch, K.R.; Lacana, E.; Suhr, K.-B.; Milstien, S.; Spiegel, S. The G Protein-Coupled Receptor GPR4 Suppresses ERK Activation in a Ligand-Independent Manner †. Biochemistry 2003, 42, 12181–12191. [Google Scholar] [CrossRef] [PubMed]
- Drzazga, A.; Okulus, M.; Rychlicka, M.; Biegała, Ł.; Gliszczyńska, A.; Gendaszewska-Darmach, E. Lysophosphatidylcholine containing anisic acid is able to stimulate insulin secretion targeting g protein coupled receptors. Nutrients 2020, 12, 1173. [Google Scholar] [CrossRef]
- Kim, Y.L.; Im, Y.J.; Ha, N.C.; Im, D.S. Albumin inhibits cytotoxic activity of lysophosphatidylcholine by direct binding. Prostaglandins Other Lipid Mediat. 2007, 83, 130–138. [Google Scholar] [CrossRef]
- Gillett, M.P.T.; Besterman, E.M.M. Plasma concentrations of lysolecithin and other phospholipids in the healthy population and in men suffering from atherosclerotic diseases. Atherosclerosis 1975, 22, 111–124. [Google Scholar] [CrossRef]
- Switzer, S.; Eder, H.A. Transport of lysolecithin by albumin in human and rat plasma. J. Lipid Res. 1965, 6, 506–511. [Google Scholar]
- Vuong, T.H.I.D.; Stroes, E.S.G.; Willekes-Koolschijn, N.E.L.; Rabelink, T.O.N.J.; Koomans, H.A.; Joles, J.A. Hypoalbuminemia increases lysophosphatidylcholine in low-density lipoprotein of normocholesterolemic subjects. Kidney Int. 1999, 55, 1005–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, L.A.; Arends, J.; Hodina, A.K.; Unger, C.; Massing, U. Plasma lyso-phosphatidylcholine concentration is decreased in cancer patients with weight loss and activated inflammatory status. Lipids Health Dis. 2007, 6, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreoli, V.M.; Maffei, F.; Tonon, G.C.; Zibetti, A. Significance of plasma lysolecithin in patients with multiple sclerosis: A longitudinal study. J. Neurol. Neurosurg. Psychiatry 1973, 36, 661–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joles, J.A.; Willekes-Koolschun, N.; Koons, H.A. Hypoalbuminemia causes high blood viscosity by increasing red cell lysophosphatidylcholine. Kidney Int. 1997, 52, 761–770. [Google Scholar] [CrossRef] [Green Version]
- Lauber, K.; Bohn, E.; Kröber, S.M.; Xiao, Y.; Blumenthal, S.G.; Lindemann, R.K.; Marini, P.; Wiedig, C.; Zobywalski, A.; Baksh, S.; et al. Apoptotic Cells Induce Migration of Phagocytes via Caspase-3-Mediated Release of a Lipid Attraction Signal. Cell 2003, 113, 717–730. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, B.; Schiller, J. Lysophospholipids: Their generation, physiological role and detection. Are they important disease markers? Mini Rev. Med. Chem. 2009, 9, 368–378. [Google Scholar] [CrossRef]
- Rousset, X.; Vaisman, B.; Amar, M.; Sethi, A.A.; Remaley, A.T. Lecithin: Cholesterol acyltransferase--from biochemistry to role in cardiovascular disease. Curr. Opin. Endocrinol. Diabetes. Obes. 2009, 16, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Santamarina-Fojo, S.; Gonzaález-Navarro, H.; Freeman, L.; Wagner, E.; Nong, Z. Hepatic Lipase, Lipoprotein Metabolism, and Atherogenesis. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1750–1754. [Google Scholar] [CrossRef]
- Gauster, M.; Rechberger, G.; Sovic, A.; Hörl, G.; Steyrer, E.; Sattler, W.; Frank, S. Endothelial lipase releases saturated and unsaturated fatty acids of high density lipoprotein phosphatidylcholine. J. Lipid Res. 2005, 46, 1517–1525. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, T.; Kobayashi, T.; Kamata, K. Role of Lysophosphatidylcholine (LPC) in Atherosclerosis. Curr. Med. Chem. 2007, 14, 3209–3220. [Google Scholar] [CrossRef] [Green Version]
- Seilhamer, J.J.; Pruzanski, W.; Vadas, P.; Plant, S.; Miller, J.A.; Kloss, J.; Johnson, L.K. Cloning and recombinant expression of phospholipase A2 present in rheumatoid arthritic synovial fluid. J. Biol. Chem. 1989, 264, 5335–5338. [Google Scholar] [PubMed]
- Murakami, M.; Sato, H.; Miki, Y.; Yamamoto, K.; Taketomi, Y. A new era of secreted phospholipase A₂. J. Lipid Res. 2015, 56, 1248–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanna, V.S.; Hafez, E.A.A. Synopsis of arachidonic acid metabolism: A review. J. Adv. Res. 2018, 11, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Schiller, J.; Wagner, U.; Häntzschel, H.; Arnold, K. The phosphatidylcholine/lysophosphatidylcholine ratio in human plasma is an indicator of the severity of rheumatoid arthritis: Investigations by 31P NMR and MALDI-TOF MS. Clin. Biochem. 2005, 38, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Barber, M.N.; Risis, S.; Yang, C.; Meikle, P.J.; Staples, M.; Febbraio, M.A.; Bruce, C.R. Plasma Lysophosphatidylcholine Levels Are Reduced in Obesity and Type 2 Diabetes. PLoS ONE 2012, 7, e41456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Cheng, S.L.; Fei, Q.; Gu, H.; Raftery, D.; Cao, B.; Sun, X.; Yan, J.; Zhang, C.; Wang, J. Metabolic profiling identifies phospholipids as potential serum biomarkers for schizophrenia. Psychiatry Res. 2019, 272, 18–29. [Google Scholar] [CrossRef]
- Sun, Z.; Chang, H.M.; Wang, A.; Song, J.; Zhang, X.; Guo, J.; Leung, P.C.K.; Lian, F. Identification of potential metabolic biomarkers of polycystic ovary syndrome in follicular fluid by SWATH mass spectrometry. Reprod. Biol. Endocrinol. 2019, 17, 45. [Google Scholar] [CrossRef]
- Jiang, Y.; Qi, J.; Xue, X.; Huang, R.; Zheng, J.; Liu, W.; Yin, H.; Li, S. Ceramide subclasses identified as novel lipid biomarker elevated in women with polycystic ovary syndrome: A pilot study employing shotgun lipidomics. Gynecol. Endocrinol. 2019, 1–5. [Google Scholar] [CrossRef]
- Grimm, M.O.W.; Grösgen, S.; Riemenschneider, M.; Tanila, H.; Grimm, H.S.; Hartmann, T. From brain to food: Analysis of phosphatidylcholins, lyso-phosphatidylcholins and phosphatidylcholin–plasmalogens derivates in Alzheimer’s disease human post mortem brains and mice model via mass spectrometry. J. Chromatogr. A 2011, 1218, 7713–7722. [Google Scholar] [CrossRef]
- Mulder, C.; Wahlund, L.-O.; Teerlink, T.; Blomberg, M.; Veerhuis, R.; van Kamp, G.J.; Scheltens, P.; Scheffer, P.G. Decreased lysophosphatidylcholine/phosphatidylcholine ratio in cerebrospinal fluid in Alzheimer?s disease. J. Neural Transm. 2003, 110, 949–955. [Google Scholar] [CrossRef]
- Chen, C.; Luo, F.; Wu, P.; Huang, Y.; Das, A.; Chen, S.; Chen, J.; Hu, X.; Li, F.; Fang, Z.; et al. Metabolomics reveals metabolite changes of patients with pulmonary arterial hypertension in China. J. Cell. Mol. Med. 2020, 24, 2484–2496. [Google Scholar] [CrossRef] [PubMed]
- Semba, R.D.; Zhang, P.; Adelnia, F.; Sun, K.; Gonzalez-Freire, M.; Salem, N.; Brennan, N.; Spencer, R.G.; Fishbein, K.; Khadeer, M.; et al. Low plasma lysophosphatidylcholines are associated with impaired mitochondrial oxidative capacity in adults in the Baltimore Longitudinal Study of Aging. Aging Cell 2019, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ried, J.S.; Baurecht, H.; Stückler, F.; Krumsiek, J.; Gieger, C.; Heinrich, J.; Kabesch, M.; Prehn, C.; Peters, A.; Rodriguez, E.; et al. Integrative genetic and metabolite profiling analysis suggests altered phosphatidylcholine metabolism in asthma. Allergy Eur. J. Allergy Clin. Immunol. 2013, 68, 629–636. [Google Scholar] [CrossRef]
- Krautbauer, S.; Eisinger, K.; Wiest, R.; Liebisch, G.; Buechler, C. Systemic saturated lysophosphatidylcholine is associated with hepatic function in patients with liver cirrhosis. Prostaglandins Other Lipid Mediat. 2016, 124, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Gimbrone, M.A.; García-Cardeña, G. Endothelial Cell Dysfunction and the Pathobiology of Atherosclerosis. Circ. Res. 2016, 118, 620–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, M.; Hirata, K.; Miyake, R.; Akita, H.; Ishikawa, Y.; Fukuzaki, H. Lysophosphatidylcholine: Essential role in the inhibition of endothelium-dependent vasorelaxation by oxidized low density lipoprotein. Biochem. Biophys. Res. Commun. 1990, 168, 301–308. [Google Scholar] [CrossRef]
- Kugiyama, K.; Kerns, S.A.; Morrisett, J.D.; Roberts, R.; Henry, P.D. Impairment of endothelium-dependent arterial relaxation by lysolecithin in modified low-density lipoproteins. Nature 1990, 344, 160–162. [Google Scholar] [CrossRef]
- Mahfouz, M.; Kummerow, F. Oxidized low density lipoprotein inhibits prostacyclin generation by rat aorta in vitro: A key role of lysolecithin. Prostaglandins Other Lipid Mediat. 2001, 66, 283–304. [Google Scholar] [CrossRef]
- Stoll, L.L.; Oskarsson, H.J.; Spector, A.A. Interaction of lysophosphatidylcholine with aortic endothelial cells. Am. J. Physiol. Circ. Physiol. 1992, 262, H1853–H1860. [Google Scholar] [CrossRef]
- Froese, D.E.; McMaster, J.; Man, R.Y.; Choy, P.C.; Kroeger, E.A. Inhibition of endothelium-dependent vascular relaxation by lysophosphatidylcholine: Impact of lysophosphatidylcholine on mechanisms involving endothelium-derived nitric oxide and endothelium derived hyperpolarizing factor. Mol. Cell. Biochem. 1999, 197, 1–6. [Google Scholar] [CrossRef]
- Okamoto, Y.; Kawahara, Y.; Yokoyama, M. Inhibition by lysophosphatidylcholine of nitric oxide production in interleukin 1 beta-stimulated vascular smooth muscle cells. Kobe J. Med. Sci. 1998, 44, 169–189. [Google Scholar] [PubMed]
- Rao, S.P.; Riederer, M.; Lechleitner, M.; Hermansson, M.; Desoye, G.; Hallström, S.; Graier, W.F.; Frank, S. Acyl Chain-Dependent Effect of Lysophosphatidylcholine on Endothelium-Dependent Vasorelaxation. PLoS ONE 2013, 8, e65155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murohara, T.; Kugiyama, K.; Ohgushi, M.; Sugiyama, S.; Ohta, Y.; Yasue, H. LPC in oxidized LDL elicits vasocontraction and inhibits endothelium- dependent relaxation. Am. J. Physiol. Circ. Physiol. 1994, 267, H2441–H2449. [Google Scholar] [CrossRef] [PubMed]
- Galle, J.; Mameghani, A.; Bolz, S.-S.; Gambaryan, S.; Görg, M.; Quaschning, T.; Raff, U.; Barth, H.; Seibold, S.; Wanner, C.; et al. Oxidized LDL and its compound lysophosphatidylcholine potentiate AngII-induced vasoconstriction by stimulation of RhoA. J. Am. Soc. Nephrol. 2003, 14, 1471–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bing, R.J.; Saeed, M. The role of lysolecithin in the relaxation of vascular smooth muscle. Biosci. Rep. 1987, 7, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Wolf, A.; Menon, N.K.; Saeed, M.; Alves, C.; Bing, R.J. Lysolecithins as endothelium-dependent vascular smooth muscle relaxants that differ from endothelium-derived relaxing factor (nitric oxide). Proc. Natl. Acad. Sci. USA 1988, 85, 8246–8250. [Google Scholar] [CrossRef] [Green Version]
- Jougasaki, M.; Kugiyama, K.; Saito, Y.; Nakao, K.; Imura, H.; Yasue, H. Suppression of endothelin-1 secretion by lysophosphatidylcholine in oxidized low density lipoprotein in cultured vascular endothelial cells. Circ. Res. 1992, 71, 614–619. [Google Scholar] [CrossRef] [Green Version]
- Zembowicz, A.; Jones, S.L.; Wu, K.K. Induction of cyclooxygenase-2 in human umbilical vein endothelial cells by lysophosphatidylcholine. J. Clin. Invest. 1995, 96, 1688–1692. [Google Scholar] [CrossRef]
- Brkić, L.; Riederer, M.; Graier, W.F.; Malli, R.; Frank, S. Acyl chain-dependent effect of lysophosphatidylcholine on cyclooxygenase (COX)-2 expression in endothelial cells. Atherosclerosis 2012, 224, 348–354. [Google Scholar] [CrossRef] [Green Version]
- Zembowicz, A.; Tang, J.L.; Wu, K.K. Transcriptional induction of endothelial nitric oxide synthase type III by lysophosphatidylcholine. J. Biol. Chem. 1995, 270, 17006–17010. [Google Scholar] [CrossRef] [Green Version]
- Xing, F.; Liu, J.; Mo, Y.; Liu, Z.; Qin, Q.; Wang, J.; Fan, Z.; Long, Y.; Liu, N.; Zhao, K.; et al. Lysophosphatidylcholine up-regulates human endothelial nitric oxide synthase gene transactivity by c-Jun N-terminal kinase signalling pathway. J. Cell. Mol. Med. 2009, 13, 1136–1148. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.M.; Libby, P.; Barton, M. Linking regulation of nitric oxide to endothelin-1: The Yin and Yang of vascular tone in the atherosclerotic plaque. Atherosclerosis 2020, 292, 201–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radulović, S.; Gottschalk, B.; Hörl, G.; Zardoya-Laguardia, P.; Schilcher, I.; Hallström, S.; Vujić, N.; Schmidt, K.; Trieb, M.; Graier, W.F.; et al. Endothelial lipase increases eNOS activating capacity of high-density lipoprotein. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2020, 1865. [Google Scholar] [CrossRef] [PubMed]
- Schilcher, I.; Ledinski, G.; Radulović, S.; Hallström, S.; Eichmann, T.; Madl, T.; Zhang, F.; Leitinger, G.; Kolb-Lenz, D.; Darnhofer, B.; et al. Endothelial lipase increases antioxidative capacity of high-density lipoprotein. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2019, 1864, 1363–1374. [Google Scholar] [CrossRef]
- Riederer, M.; Ojala, P.J.; Hrzenjak, A.; Graier, W.F.; Malli, R.; Tritscher, M.; Hermansson, M.; Watzer, B.; Schweer, H.; Desoye, G.; et al. Acyl chain-dependent effect of lysophosphatidylcholine on endothelial prostacyclin production. J. Lipid Res. 2010, 51, 2957–2966. [Google Scholar] [CrossRef] [Green Version]
- Wolf, A.; Saito, T.; Menon, N.K.; Zehetgruber, M.; Bing, R.J. Effect of lysophosphatidylcholine on atherosclerotic rabbit arteries. Atherosclerosis 1989, 80, 81–89. [Google Scholar] [CrossRef]
- Wolf, A.; Saito, T.; Dudek, R.; Bing, R.J. The effect of lysophosphatidylcholine on coronary and renal circulation in the rabbit. Lipids 1991, 26, 223–226. [Google Scholar] [CrossRef]
- Hansson, G.K.; Libby, P. The immune response in atherosclerosis: A double-edged sword. Nat. Rev. Immunol. 2006, 6, 508–519. [Google Scholar] [CrossRef]
- Hansson, G.K.; Hermansson, A. The immune system in atherosclerosis. Nat. Immunol. 2011, 12, 204–212. [Google Scholar] [CrossRef]
- Galkina, E.; Ley, K. Immune and Inflammatory Mechanisms of Atherosclerosis. Annu. Rev. Immunol. 2009, 27, 165–197. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, S.; Kugiyama, K.; Ohgushi, M.; Fujimoto, K.; Yasue, H. Lysophosphatidylcholine in oxidized low-density lipoprotein increases endothelial susceptibility to polymorphonuclear leukocyte-induced endothelial dysfunction in porcine coronary arteries. Role of protein kinase C. Circ. Res. 1994, 74, 565–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Lin, J.H.; Liao, H.L.; Verna, L.; Stemerman, M.B. Activation of ICAM-1 promoter by lysophosphatidylcholine: Possible involvement of protein tyrosine kinases. Biochim. Biophys. Acta 1997, 1345, 93–98. [Google Scholar] [CrossRef]
- Zou, Y.; Kim, C.H.; Chung, J.H.; Kim, J.Y.; Chung, S.W.; Kim, M.K.; Im, D.S.; Lee, J.; Yu, B.P.; Chung, H.Y. Upregulation of endothelial adhesion molecules by lysophosphatidylcholine. FEBS J. 2007, 274, 2573–2584. [Google Scholar] [CrossRef] [PubMed]
- Murohara, T.; Scalia, R.; Lefer, A.M. Lysophosphatidylcholine promotes P-selectin expression in platelets and endothelial cells. Possible involvement of protein kinase C activation and its inhibition by nitric oxide donors. Circ. Res. 1996, 78, 780–789. [Google Scholar] [CrossRef]
- Li, X.; Wang, L.; Fang, P.; Sun, Y.; Jiang, X.; Wang, H.; Yang, X.-F. Lysophospholipids induce innate immune transdifferentiation of endothelial cells, resulting in prolonged endothelial activation. J. Biol. Chem. 2018, 293, 11033–11045. [Google Scholar] [CrossRef] [Green Version]
- Jing, Q.; Xin, S.M.; Zhang, W.B.; Wang, P.; Qin, Y.W.; Pei, G. Lysophosphatidylcholine activates p38 and p42/44 mitogen-activated protein kinases in monocytic THP-1 cells, but only p38 activation is involved in its stimulated chemotaxis. Circ. Res. 2000, 87, 52–59. [Google Scholar] [CrossRef]
- Quinn, M.T.; Parthasarathy, S.; Steinberg, D. Lysophosphatidylcholine: A chemotactic factor for human monocytes and its potential role in atherogenesis. Proc. Natl. Acad. Sci. USA 1988, 85, 2805–2809. [Google Scholar] [CrossRef] [Green Version]
- Radu, C.G.; Yang, L.V.; Riedinger, M.; Au, M.; Witte, O.N. T cell chemotaxis to lysophosphatidylcholine through the G2A receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Damaj, B.B.; Maghazachi, A.A. Human resting CD16-, CD16+ and IL-2-, IL-12-, IL-15- or IFN-α-activated natural killer cells differentially respond to sphingosylphosphorylcholine, lysophosphatidylcholine and platelet-activating factor. Eur. J. Immunol. 2005, 35, 2699–2708. [Google Scholar] [CrossRef]
- Gisterå, A.; Hansson, G.K. The immunology of atherosclerosis. Nat. Rev. Nephrol. 2017, 13, 368–380. [Google Scholar] [CrossRef]
- Ngwenya, B.Z.; Yamamoto, N. Activation of peritoneal macrophages by lysophosphatidylcholine. Biochim. Biophys. Acta 1985, 839, 9–15. [Google Scholar] [CrossRef]
- Qin, X.; Qiu, C.; Zhao, L. Lysophosphatidylcholine perpetuates macrophage polarization toward classically activated phenotype in inflammation. Cell. Immunol. 2014, 289, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Oestvang, J.; Anthonsen, M.W.; Johansen, B. LysoPC and PAF Trigger Arachidonic Acid Release by Divergent Signaling Mechanisms in Monocytes. J. Lipids 2011, 2011, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, M.H.; Lee, S.M.; Jung, J. Lysophosphatidylcholine induces expression of genes involved in cholesterol biosynthesis in THP-1 derived macrophages. Steroids 2018, 139, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Rolin, J.; Vego, H.; Maghazachi, A. Oxidized Lipids and Lysophosphatidylcholine Induce the Chemotaxis, Up-Regulate the Expression of CCR9 and CXCR4 and Abrogate the Release of IL-6 in Human Monocytes. Toxins (Basel) 2014, 6, 2840–2856. [Google Scholar] [CrossRef] [Green Version]
- Hourton, D.; Stengel, D.; Chapman, M.J.; Ninio, E. Oxidized low density lipoproteins downregulate LPS-induced platelet-activating factor receptor expression in human monocyte-derived macrophages: Implications for LPS-induced nuclear factor-kappaB binding activity. Eur. J. Biochem. 2001, 268, 4489–4496. [Google Scholar] [CrossRef]
- Carneiro, A.B.; Iaciura, B.M.F.; Nohara, L.L.; Lopes, C.D.; Veas, E.M.C.; Mariano, V.S.; Bozza, P.T.; Lopes, U.G.; Atella, G.C.; Almeida, I.C.; et al. Lysophosphatidylcholine Triggers TLR2- and TLR4-Mediated Signaling Pathways but Counteracts LPS-Induced NO Synthesis in Peritoneal Macrophages by Inhibiting NF-κB Translocation and MAPK/ERK Phosphorylation. PLoS ONE 2013, 8, e76233. [Google Scholar] [CrossRef] [Green Version]
- Hara, S.; Shike, T.; Takasu, N.; Mizui, T. Lysophosphatidylcholine promotes cholesterol efflux from mouse macrophage foam cells. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 1258–1266. [Google Scholar] [CrossRef]
- Hou, M.; Xia, M.; Zhu, H.; Wang, Q.; Li, Y.; Xiao, Y.; Zhao, T.; Tang, Z.; Ma, J.; Ling, W. Lysophosphatidylcholine promotes cholesterol efflux from mouse macrophage foam cells via PPARγ-LXRα-ABCA1-dependent pathway associated with apoE. Cell Biochem. Funct. 2007, 25, 33–44. [Google Scholar] [CrossRef]
- O’Neil, L.J.; Kaplan, M.J.; Carmona-Rivera, C. The Role of Neutrophils and Neutrophil Extracellular Traps in Vascular Damage in Systemic Lupus Erythematosus. J. Clin. Med. 2019, 8, 1325. [Google Scholar] [CrossRef] [Green Version]
- Bot, I.; Shi, G.P.; Kovanen, P.T. Mast cells as effectors in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 265–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marx, C.; Novotny, J.; Salbeck, D.; Zellner, K.R.; Nicolai, L.; Pekayvaz, K.; Kilani, B.; Stockhausen, S.; Bürgener, N.; Kupka, D.; et al. Eosinophil-platelet interactions promote atherosclerosis and stabilize thrombosis with eosinophil extracellular traps. Blood 2019, 134, 1859–1872. [Google Scholar] [CrossRef] [Green Version]
- Englberger, W.; Bitter-Suermann, D.; Hadding, U. Influence of lysophospholipids and PAF on the oxidative burst of PMNL. Int. J. Immunopharmacol. 1987, 9, 275–282. [Google Scholar] [CrossRef]
- Lin, P.; Welch, E.J.; Gao, X.-P.; Malik, A.B.; Ye, R.D. Lysophosphatidylcholine modulates neutrophil oxidant production through elevation of cyclic AMP. J. Immunol. 2005, 174, 2981–2989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curcic, S.; Holzer, M.; Frei, R.; Pasterk, L.; Schicho, R.; Heinemann, A.; Marsche, G. Neutrophil effector responses are suppressed by secretory phospholipase A2 modified HDL. Biochim. Biophys. Acta 2015, 1851, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, O.; Kume, H.; Kondo, M.; Ito, Y.; Ito, M.; Yamaki, K. Role of lysophosphatidylcholine in eosinophil infiltration and resistance in airways. Clin. Exp. Pharmacol. Physiol. 2004, 31, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Learoyd, J.; Butt, S.; Zhu, L.; Usatyuk, P.V.; Natarajan, V.; Munoz, N.M.; Leff, A.R. Regulation of eosinophil adhesion by lysophosphatidylcholine via a non-store-operated Ca2+ channel. Am. J. Respir. Cell Mol. Biol. 2007, 36, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Knuplez, E.; Curcic, S.; Theiler, A.; Bärnthaler, T.; Trakaki, A.; Trieb, M.; Holzer, M.; Heinemann, A.; Zimmermann, R.; Sturm, E.M.; et al. Lysophosphatidylcholines inhibit human eosinophil activation and suppress eosinophil migration in vivo. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2020, 1865, 158686. [Google Scholar] [CrossRef]
- Marquardt, D.L.; Walker, L.L. Lysophosphatidylcholine induces mast cell secretion and protein kinase C activation. J. Allergy Clin. Immunol. 1991, 88, 721–730. [Google Scholar] [CrossRef]
- Mio, M.; Ikeda, A.; Akagi, M.; Tasaka, K. Inhibitory effect of lysophosphatidylcholine on the histamine release from rat peritoneal mast cells. Agents Actions 1985, 16, 113–117. [Google Scholar] [CrossRef]
- Hansson, G.K.; Libby, P.; Schönbeck, U.; Yan, Z.Q. Innate and adaptive immunity in the pathogenesis of atherosclerosis. Circ. Res. 2002, 91, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Stemme, S.; Faber, B.; Holm, J.; Wiklund, O.; Witztum, J.L.; Hansson, G.K. T lymphocytes from human atherosclerotic plaques recognize oxidized low density lipoprotein. Proc. Natl. Acad. Sci. USA 1995, 92, 3893–3897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulsson, G.; Zhou, X.; Törnquist, E.; Hansson, G.K. Oligoclonal T cell expansions in atherosclerotic lesions of apolipoprotein E-deficient mice. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 10–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asaoka, Y.; Oka, M.; Yoshida, K.; Sasaki, Y.; Nishizuka, Y. Role of lysophosphatidylcholine in T-lymphocyte activation: Involvement of phospholipase A2 in signal transduction through protein kinase C. Proc. Natl. Acad. Sci. USA 1992, 89, 6447–6451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, Y.; Kusumi, Y.; Mitsumata, M.; Li, X.-K.; Fujino, M. Lysophosphatidylcholine upregulates LOX-1, chemokine receptors, and activation-related transcription factors in human T-cell line Jurkat. J. Thromb. Thrombolysis 2008, 26, 113–118. [Google Scholar] [CrossRef]
- Im, Y.; Lee, Y.; Chung, H.; Im, D. Multiple actions of lysophosphatidylcholine in human Jurkat T cells1. Acta Pharmacol. Sin. 2006, 27, 700–707. [Google Scholar] [CrossRef] [Green Version]
- Sakata-Kaneko, S.; Wakatsuki, Y.; Usui, T.; Matsunaga, Y.; Itoh, T.; Nishi, E.; Kume, N.; Kita, T. Lysophosphatidylcholine upregulates CD40 ligand expression in newly activated human CD4+ T cells. FEBS Lett. 1998, 433, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Han, K.H.; Hong, K.H.; Ko, J.; Rhee, K.S.; Hong, M.K.; Kim, J.J.; Kim, Y.H.; Park, S.J. Lysophosphatidylcholine up-regulates CXCR4 chemokine receptor expression in human CD4 T cells. J. Leukoc. Biol. 2004, 76, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Nishi, E.; Kume, N.; Ueno, Y.; Ochi, H.; Moriwaki, H.; Kita, T. Lysophosphatidylcholine enhances cytokine-induced interferon gamma expression in human T lymphocytes. Circ. Res. 1998, 83, 508–515. [Google Scholar] [CrossRef]
- Hasegawa, H.; Lei, J.; Matsumoto, T.; Onishi, S.; Suemori, K.; Yasukawa, M. Lysophosphatidylcholine enhances the suppressive function of human naturally occurring regulatory T cells through TGF-β production. Biochem. Biophys. Res. Commun. 2011, 415, 526–531. [Google Scholar] [CrossRef]
- Ait-Oufella, H.; Salomon, B.L.; Potteaux, S.; Robertson, A.K.L.; Gourdy, P.; Zoll, J.; Merval, R.; Esposito, B.; Cohen, J.L.; Fisson, S.; et al. Natural regulatory T cells control the development of atherosclerosis in mice. Nat. Med. 2006, 12, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Planer, D.; Luboshits, G.; Afek, A.; Metzger, S.; Chajek-Shaul, T.; Keren, G.; George, J. Role of naturally occurring CD4+CD25+ regulatory T cells in experimental atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 893–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grainger, D.J.; Kemp, P.R.; Metcalfe, J.C.; Liu, A.C.; Lawn, R.M.; Williams, N.R.; Grace, A.A.; Schofield, P.M.; Chauhan, A. The serum concentration of active transforming growth factor-β is severely depressed in advanced atherosclerosis. Nat. Med. 1995, 1, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Mallat, Z.; Gojova, A.; Marchiol-Fournigault, C.; Esposito, B.; Kamaté, C.; Merval, R.; Fradelizi, D.; Tedgui, A. Inhibition of transforming growth factor-β signaling accelerates atherosclerosis and induces an unstable plaque phenotype in mice. Circ. Res. 2001, 89, 930–934. [Google Scholar] [CrossRef] [Green Version]
- Nording, H.M.; Seizer, P.; Langer, H.F. Platelets in inflammation and atherogenesis. Front. Immunol. 2015, 6, 98. [Google Scholar] [CrossRef] [Green Version]
- Weber, C. Platelets and chemokines in atherosclerosis: Partners in crime. Circ. Res. 2005, 96, 612–616. [Google Scholar] [CrossRef] [Green Version]
- Kirschmann, C.; Aloof, S.; Devries, A. Action of Lysolecithin on Blood Platelets. Thromb. Diath. Haemorrh. 1963, 18, 512–524. [Google Scholar] [CrossRef]
- Yuan, Y.; Jackson, S.P.; Newnham, H.H.; Mitchell, C.A.; Salem, H.H. An essential role for lysophosphatidylcholine in the inhibition of platelet aggregation by secretory phospholipase A2. Blood 1995, 86, 4166–4174. [Google Scholar] [CrossRef]
- Curcic, S.; Holzer, M.; Pasterk, L.; Knuplez, E.; Eichmann, T.O.; Frank, S.; Zimmermann, R.; Schicho, R.; Heinemann, A.; Marsche, G. Secretory phospholipase A2 modified HDL rapidly and potently suppresses platelet activation. Sci. Rep. 2017, 7, 8030. [Google Scholar] [CrossRef]
- Revel-Vilk, S.; Rand, M.L.; Israels, S.J. Primary and Secondary Hemostasis, Regulators of Coagulation, and Fibrinolysis: Understanding the Basics. In SickKids Handbook of Pediatric Thrombosis and Hemostasis; KARGER: Basel, Switzerland, 2013; pp. 5–13. [Google Scholar]
- Engelmann, B.; Zieseniss, S.; Brand, K.; Page, S.; Lentschat, A.; Ulmer, A.J.; Gerlach, E. Tissue factor expression of human monocytes is suppressed by lysophosphatidylcholine. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Hara, H.; Adachi, T. The expression of extracellular-superoxide dismutase is increased by lysophosphatidylcholine in human monocytic U937 cells. Atherosclerosis 2002, 163, 223–228. [Google Scholar] [CrossRef]
- Mendonça, M.C.; Rezende, A.; Doi, S.Q.; Sellitti, D.F. Lysophosphatidylcholine increases C-type natriuretic peptide expression in human vascular smooth muscle cells via membrane distortion. Vascul. Pharmacol. 2009, 51, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.-K.; Hartvigsen, K.; Ryu, J.; Kim, Y.; Han, K. The pro-atherogenic effects of macrophages are reduced upon formation of a complex between C-reactive protein and lysophosphatidylcholine. J. Inflamm. 2012, 9, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-M.; Han, H.-J.; Hur, Y.-H.; Quan, H.; Kwak, S.-H.; Choi, J.-I.; Bae, H.-B. Stearoyl lysophosphatidylcholine prevents lipopolysaccharide-induced extracellular release of high mobility group box-1 through AMP-activated protein kinase activation. Int. Immunopharmacol. 2015, 28, 540–545. [Google Scholar] [CrossRef]
- Coutant, F.; Perrin-Cocon, L.; Agaugué, S.; Delair, T.; André, P.; Lotteau, V. Mature dendritic cell generation promoted by lysophosphatidylcholine. J. Immunol. 2002, 169, 1688–1695. [Google Scholar] [CrossRef] [Green Version]
- Bühligen, J.; Himmel, M.; Gebhardt, C.; Simon, J.C.; Ziegler, W.; Averbeck, M. Lysophosphatidylcholine-mediated functional inactivation of syndecan-4 results in decreased adhesion and motility of dendritic cells. J. Cell. Physiol. 2010, 225, 905–914. [Google Scholar] [CrossRef]
- Piccirillo, A.R.; Hyzny, E.J.; Beppu, L.Y.; Menk, A.V.; Wallace, C.T.; Hawse, W.F.; Buechel, H.M.; Wong, B.H.; Foo, J.C.; Cazenave-Gassiot, A.; et al. The Lysophosphatidylcholine Transporter MFSD2A Is Essential for CD8 + Memory T Cell Maintenance and Secondary Response to Infection. J. Immunol. 2019, 203, 117–126. [Google Scholar] [CrossRef]
- Whalen, M.M.; Doshi, R.N.; Bader, B.W.; Bankhurst, A.D. Lysophosphatidylcholine and Arachidonic Acid Are Required in the Cytotoxic Response of Human Natural Killer Cells to Tumor Target Cells. Cell. Physiol. Biochem. 1999, 9, 297–309. [Google Scholar] [CrossRef]
- Hong, C.W.; Kim, T.K.; Ham, H.Y.; Nam, J.S.; Kim, Y.H.; Zheng, H.; Pang, B.; Min, T.K.; Jung, J.S.; Lee, S.N.; et al. Lysophosphatidylcholine Increases Neutrophil Bactericidal Activity by Enhancement of Azurophil Granule-Phagosome Fusion via Glycine{middle dot}GlyR 2/TRPM2/p38 MAPK Signaling. J. Immunol. 2010, 184, 4401–4413. [Google Scholar] [CrossRef]
- Ginsburg, I.; Ward, P.A.; Varani, J. Lysophosphatides enhance superoxide responses of stimulated human neutrophils. Inflammation 1989, 13, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Butterworth, A.E.; Cater, D.B. Effect of lysolecithin on the oxygen uptake of tumour cells, polymorphonuclear leucocytes, lymphocytes and macrophages in vitro. Br. J. Cancer 1967, 21, 373–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, T.; Heuter, S.; Schlesinger, M.; Jakubzig, B.; Raynor, A.; Massing, U.; Bendas, G. Lysophosphatidylcholine attenuates melanoma cell adhesion and migration dependent on the degree of fatty acid saturation. Int. J. Clin. Pharmacol. Ther. 2015, 53, 1049–1051. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Jin, M.; Brietzke, E.; McIntyre, R.S.; Wang, D.; Rosenblat, J.D.; Ragguett, R.M.; Zhang, C.; Sun, X.; Rong, C.; et al. Serum metabolic profiling using small molecular water-soluble metabolites in individuals with schizophrenia: A longitudinal study using a pre–post-treatment design. Psychiatry Clin. Neurosci. 2019, 73, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Park, D.W.; Kwak, D.S.; Park, Y.Y.; Chang, Y.; Huh, J.W.; Lim, C.-M.; Koh, Y.; Song, D.-K.; Hong, S.-B. Impact of serial measurements of lysophosphatidylcholine on 28-day mortality prediction in patients admitted to the intensive care unit with severe sepsis or septic shock. J. Crit. Care 2014, 29, 882.e5-882.e11. [Google Scholar] [CrossRef] [PubMed]
- Drobnik, W.; Liebisch, G.; Audebert, F.-X.; Fröhlich, D.; Glück, T.; Vogel, P.; Rothe, G.; Schmitz, G. Plasma ceramide and lysophosphatidylcholine inversely correlate with mortality in sepsis patients. J. Lipid Res. 2003, 44, 754–761. [Google Scholar] [CrossRef] [Green Version]
- Cho, W.H.; Yeo, H.J.; Yoon, S.H.; Lee, S.E.; Jeon, D.S.; Kim, Y.S.; Lee, S.J.; Jo, E.J.; Mok, J.H.; Kim, M.H.; et al. Lysophosphatidylcholine as a prognostic marker in community-acquired pneumonia requiring hospitalization: A pilot study. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Van der Kolk, B.W.; Vogelzangs, N.; Jocken, J.W.E.; Valsesia, A.; Hankemeier, T.; Astrup, A.; Saris, W.H.M.; Arts, I.C.W.; van Greevenbroek, M.M.J.; Blaak, E.E. Plasma lipid profiling of tissue-specific insulin resistance in human obesity. Int. J. Obes. 2019, 43, 989–998. [Google Scholar] [CrossRef]
- Wang, S.; Yu, X.; Zhang, W.; Ji, F.; Wang, M.; Yang, R.; Li, H.; Chen, W.; Dong, J. Association of serum metabolites with impaired fasting glucose/diabetes and traditional risk factors for metabolic disease in Chinese adults. Clin. Chim. Acta 2018, 487, 60–65. [Google Scholar] [CrossRef]
- Razquin, C.; Toledo, E.; Clish, C.B.; Ruiz-Canela, M.; Dennis, C.; Corella, D.; Papandreou, C.; Ros, E.; Estruch, R.; Guasch-Ferré, M.; et al. Plasma lipidomic profiling and risk of type 2 diabetes in the PREDIMED trial. Diabetes Care 2018, 41, 2617–2624. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Mtintsilana, A.; Goedecke, J.H.; Micklesfield, L.K.; Olsson, T.; Chorell, E. Alterations in the metabolism of phospholipids, bile acids and branched-chain amino acids predicts development of type 2 diabetes in black South African women: A prospective cohort study. Metabolism. 2019, 95, 57–64. [Google Scholar] [CrossRef]
- Diamanti, K.; Cavalli, M.; Pan, G.; Pereira, M.J.; Kumar, C.; Skrtic, S.; Grabherr, M.; Risérus, U.; Eriksson, J.W.; Komorowski, J.; et al. Intra- and inter-individual metabolic profiling highlights carnitine and lysophosphatidylcholine pathways as key molecular defects in type 2 diabetes. Sci. Rep. 2019, 9, 9653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duranton, F.; Laget, J.; Gayrard, N.; Saulnier-Blache, J.S.; Lundin, U.; Schanstra, J.P.; Mischak, H.; Weinberger, K.M.; Servel, M.F.; Argilés, A. The CKD plasma lipidome varies with disease severity and outcome. J. Clin. Lipidol. 2019, 13, 176–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, S.; Yang, L.; Li, L.; Bai, Y.; Cai, C.; Liu, H. A plasma lipidomics strategy reveals perturbed lipid metabolic pathways and potential lipid biomarkers of human colorectal cancer. J. Chromatogr. B 2017, 1068–1069, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Xiao, Y.; Elson, P.; Tan, H.; Plummer, S.J.; Berk, M.; Aung, P.P.; Lavery, I.C.; Achkar, J.P.; Li, L.; et al. Plasma Lysophosphatidylcholine Levels: Potential Biomarkers for Colorectal Cancer. J. Clin. Oncol. 2007, 25, 2696–2701. [Google Scholar] [CrossRef]
- Geijsen, A.J.M.R.; van Roekel, E.H.; van Duijnhoven, F.J.B.; Achaintre, D.; Bachleitner-Hofmann, T.; Baierl, A.; Bergmann, M.M.; Boehm, J.; Bours, M.J.L.; Brenner, H.; et al. Plasma metabolites associated with colorectal cancer stage: Findings from an international consortium. Int. J. Cancer 2020, 146, 3256–3266. [Google Scholar] [CrossRef] [Green Version]
- Patterson, A.D.; Maurhofer, O.; Beyoglu, D.; Lanz, C.; Krausz, K.W.; Pabst, T.; Gonzalez, F.J.; Dufour, J.-F.; Idle, J.R. Aberrant Lipid Metabolism in Hepatocellular Carcinoma Revealed by Plasma Metabolomics and Lipid Profiling. Cancer Res. 2011, 71, 6590–6600. [Google Scholar] [CrossRef] [Green Version]
- Sutphen, R.; Xu, Y.; Wilbanks, G.D.; Fiorica, J.; Grendys, E.C.; LaPolla, J.P.; Arango, H.; Hoffman, M.S.; Martino, M.; Wakeley, K.; et al. Lysophospholipids are potential biomarkers of ovarian cancer. Cancer Epidemiol. Biomarkers Prev. 2004, 13, 1185–1191. [Google Scholar]
- Yagi, T.; Kuschner, C.E.; Shoaib, M.; Choudhary, R.C.; Becker, L.B.; Lee, A.T.; Kim, J. Relative ratios enhance the diagnostic power of phospholipids in distinguishing benign and cancerous ovarian masses. Cancers (Basel) 2020, 12, 72. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-H.; Joo, J.; Park, B.; Park, S.-J.; Lee, W.J.; Han, S.-S.; Kim, T.H.; Hong, E.K.; Woo, S.M.; Yoo, B.C. Reduced levels of N’-methyl-2-pyridone-5-carboxamide and lysophosphatidylcholine 16:0 in the serum of patients with intrahepatic cholangiocarcinoma, and the correlation with recurrence-free survival. Oncotarget 2017, 8, 112598–112609. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Yu, S.E.; Kim, K.H.; Yu, M.H.; Jeong, I.H.; Cho, J.Y.; Park, S.J.; Lee, W.J.; Han, S.S.; Kim, T.H.; et al. Individualized metabolic profiling stratifies pancreatic and biliary tract cancer: A useful tool for innovative screening programs and predictive strategies in healthcare. EPMA J. 2018, 9, 287–297. [Google Scholar] [CrossRef] [Green Version]
- Yin, M.; Tan, S.; Li, X.; Hou, Y.; Cao, G.; Li, K.; Kou, J.; Lou, G. Identification of phosphatidylcholine and lysophosphatidylcholine as novel biomarkers for cervical cancers in a prospective cohort study. Tumor Biol. 2016, 37, 5485–5492. [Google Scholar] [CrossRef]
- Goto, T.; Terada, N.; Inoue, T.; Kobayashi, T.; Nakayama, K.; Okada, Y.; Yoshikawa, T.; Miyazaki, Y.; Uegaki, M.; Utsunomiya, N.; et al. Decreased expression of lysophosphatidylcholine (16:0/OH) in high resolution imaging mass spectrometry independently predicts biochemical recurrence after surgical treatment for prostate cancer. Prostate 2015, 75, 1821–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Song, Z.; Chen, Z.; Guo, Z.; Jin, H.; Ding, C.; Hong, Y.; Cai, Z. Metabolic and lipidomic characterization of malignant pleural effusion in human lung cancer. J. Pharm. Biomed. Anal. 2020, 180. [Google Scholar] [CrossRef] [PubMed]
- Kühn, T.; Floegel, A.; Sookthai, D.; Johnson, T.; Rolle-Kampczyk, U.; Otto, W.; von Bergen, M.; Boeing, H.; Kaaks, R. Higher plasma levels of lysophosphatidylcholine 18:0 are related to a lower risk of common cancers in a prospective metabolomics study. BMC Med. 2016, 14, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, J.A.; Fensom, G.K.; Rinaldi, S.; Scalbert, A.; Appleby, P.N.; Achaintre, D.; Gicquiau, A.; Gunter, M.J.; Ferrari, P.; Kaaks, R.; et al. Patterns in metabolite profile are associated with risk of more aggressive prostate cancer: A prospective study of 3,057 matched case–control sets from EPIC. Int. J. Cancer 2020, 146, 720–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.-M.; Kim, M.-J.; Noh, J.-Y.; Yun, T.G.; Kang, M.-J.; Lee, S.-G.; Yoo, B.C.; Pyun, J.-C. Simultaneous Analysis of Multiple Cancer Biomarkers Using MALDI-TOF Mass Spectrometry Based on a Parylene-Matrix Chip. J. Am. Soc. Mass Spectrom. 2020, 31, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Ivandic, B.; Castellani, L.W.; Wang, X.P.; Qiao, J.H.; Mehrabian, M.; Navab, M.; Fogelman, A.M.; Grass, D.S.; Swanson, M.E.; de Beer, M.C.; et al. Role of group II secretory phospholipase A2 in atherosclerosis: 1. Increased atherogenesis and altered lipoproteins in transgenic mice expressing group IIa phospholipase A2. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 1284–1290. [Google Scholar] [CrossRef] [Green Version]
- Hartford, M.; Wiklund, O.; Hultén, L.M.; Perers, E.; Person, A.; Herlitz, J.; Hurt-Camejo, E.; Karlsson, T.; Caidahl, K. CRP, interleukin-6, secretory phospholipase A2 group IIA, and intercellular adhesion molecule-1 during the early phase of acute coronary syndromes and long-term follow-up. Int. J. Cardiol. 2006, 108, 55–62. [Google Scholar] [CrossRef]
- Akinkuolie, A.O.; Lawler, P.R.; Chu, A.Y.; Caulfield, M.; Mu, J.; Ding, B.; Nyberg, F.; Glynn, R.J.; Ridker, P.M.; Hurt-Camejo, E.; et al. Group IIA Secretory Phospholipase A2, Vascular Inflammation, and Incident Cardiovascular Disease: An Analysis from the JUPITER Trial. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1182–1190. [Google Scholar] [CrossRef] [Green Version]
- Ghesquiere, S.A.I.; Gijbels, M.J.J.; Anthonsen, M.; Van Gorp, P.J.J.; Van Der Made, I.; Johansen, B.; Hofker, M.H.; De Winther, M.P.J. Macrophage-specific overexpression of group IIa sPLA2 increases atherosclerosis and enhances collagen deposition. J. Lipid Res. 2005, 46, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Koenig, W.; Khuseyinova, N. Lipoprotein-associated and secretory phospholipase A2 in cardiovascular disease: The epidemiological evidence. Cardiovasc. Drugs Ther. 2009, 23, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, S.J.; Kastelein, J.J.P.; Schwartz, G.G.; Bash, D.; Rosenson, R.S.; Cavender, M.A.; Brennan, D.M.; Koenig, W.; Jukema, J.W.; Nambi, V.; et al. Varespladib and Cardiovascular Events in Patients With an Acute Coronary Syndrome. JAMA 2014, 311, 252. [Google Scholar] [CrossRef] [PubMed]
- Bowton, D.L.; Dmitrienko, A.A.; Israel, E.; Zeiher, B.G.; Sides, G.D. Impact of a soluble phospholipase A2 inhibitor on inhaled allergen challenge in subjects with asthma. J. Asthma 2005, 42, 65–71. [Google Scholar] [CrossRef]
- Bradley, J.D.; Dmitrienko, A.A.; Kivitz, A.J.; Gluck, O.S.; Weaver, A.L.; Wiesenhutter, C.; Myers, S.L.; Sides, G.D. A randomized, double-blinded, placebo-controlled clinical trial of LY333013, a selective inhibitor of group II secretory phospholipase A2, in the treatment of rheumatoid arthritis. J. Rheumatol. 2005, 32, 417–423. [Google Scholar] [PubMed]
- Marathe, G.K.; Silva, A.R.; de Castro Faria Neto, H.C.; Tjoelker, L.W.; Prescott, S.M.; Zimmerman, G.A.; McIntyre, T.M. Lysophosphatidylcholine and lyso-PAF display PAF-like activity derived from contaminating phospholipids. J. Lipid Res. 2001, 42, 1430–1437. [Google Scholar]
- Besterman, J.M.; Domanico, P.L. Association and metabolism of exogenously-derived lysophosphatidylcholine by cultured mammalian cells: Kinetics and mechanisms. Biochemistry 1992, 31, 2046–2056. [Google Scholar] [CrossRef]
- Law, S.H.; Chan, M.L.; Marathe, G.K.; Parveen, F.; Chen, C.H.; Ke, L.Y. An updated review of lysophosphatidylcholine metabolism in human diseases. Int. J. Mol. Sci. 2019, 20, 1149. [Google Scholar] [CrossRef] [Green Version]
- Perrakis, A.; Moolenaar, W.H. Autotaxin: Structure-function and signaling. J. Lipid Res. 2014, 55, 1010–1018. [Google Scholar] [CrossRef] [Green Version]
- Barbayianni, E.; Magrioti, V.; Moutevelis-Minakakis, P.; Kokotos, G. Autotaxin inhibitors: A patent review. Expert Opin. Ther. Pat. 2013, 23, 1123–1132. [Google Scholar] [CrossRef]
- Bao, L.; Qi, J.; Wang, Y.; Xi, Q.; Tserennadmid, T.; Zhao, P.; Qi, J.; Damirin, A. The atherogenic actions of LPC on vascular smooth muscle cells and its LPA receptor mediated mechanism. Biochem. Biophys. Res. Commun. 2018, 503, 1911–1918. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, J.; Deng, X.; Liu, Y.; Yang, X.; Wu, Q.; Yu, C. Lysophosphatidic acid directly induces macrophage-derived foam cell formation by blocking the expression of SRBI. Biochem. Biophys. Res. Commun. 2017, 491, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Zou, J.; Li, F.; Zhang, T.; Guo, T. Lysophosphatidic acid enhances neointimal hyperplasia following vascular injury through modulating proliferation, autophagy, inflammation and oxidative stress. Mol. Med. Rep. 2018, 18, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Yang, B.; Li, X.; Liu, H.; Lei, G. Lysophosphatidic Acid Promotes Expression and Activation of Matrix Metalloproteinase 9 (MMP9) in THP-1 Cells via Toll-Like Receptor 4/Nuclear Factor-κB (TLR4/NF-κB) Signaling Pathway. Med. Sci. Monit. 2018, 24, 4861–4868. [Google Scholar] [CrossRef] [PubMed]
- Aldi, S.; Matic, L.P.; Hamm, G.; van Keulen, D.; Tempel, D.; Holmstrøm, K.; Szwajda, A.; Nielsen, B.S.; Emilsson, V.; Ait-Belkacem, R.; et al. Integrated Human Evaluation of the Lysophosphatidic Acid Pathway as a Novel Therapeutic Target in Atherosclerosis. Mol. Ther.—Methods Clin. Dev. 2018, 10, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matralis, A.N.; Afantitis, A.; Aidinis, V. Development and therapeutic potential of autotaxin small molecule inhibitors: From bench to advanced clinical trials. Med. Res. Rev. 2018, 39, 976–1013. [Google Scholar] [CrossRef]


{kind=link}
| Function/Action | Tissue/Cell Type Studied | LPC Species Examined |
|---|---|---|
| Inhibition of platelet aggregation [118,119] | platelets | Mixture [118]; LPC 16:0, LPC 18:0, LPC 18:1, LPC 18:2 [119] |
| Decreased tissue factor activity and NF-κB expression [121]; Increased expression of extracellular superoxide dismutase [122]; Suppression of IL-6 release following lipopolysaccharide (LPS) stimulation [85]; Down-regulation of platelet activating factor (PAF) receptor expression and NF-κB translocation to nucleus [86]; Decreased high-mobility group protein 1 (HMGB-1) production [125] | monocytes | LPC 16:0 [121]; Mixture [122]; Not listed [85]; Mixture [86]; LPC 18:0 [125] |
| Increase in cholesterol efflux [88,89] | macrophage foam cells | LPC 14:0, LPC 16:0, LPC 18:0 [88]; Not listed [89] |
| Vascular smooth muscle relaxation [55,56]; Decrease in mean arterial pressure and coronary, renal and total vascular resistance [67] | rabbit aortic strip [55,56] in vivo application in rabbit [67] | Mixture [55]; LPC 10:0, LPC 14:0, LPC 16:0, LPC 18:0, LPC 18:1 [56]; Not listed [67] |
| Suppression of endothelin-1 secretion [57]; Increased prostacyclin production [58,65]; Increase in NO production [60,61] | endothelial cells | LPC 16:0 [57,58,60]; LPC 16:0, LPC 18:1, LPC 20:4 [65]; Not listed [61] |
| Increased C-type natriuretic peptide expression [123] | vascular smooth muscle cells | Mixture |
| Promotion of dendritic cell maturation [126]; Reduction of cell motility and adhesion [127] | dendritic cells | Mixture [126]; LPC 18:0 [127] |
| (Potentiated) T-cell activation [104,105,106,107,108,109]; Maintenance of T-cell homeostatic turnover [128]; Enhanced suppressive function [110] | T cells [104,109] CD4+ T cells [105,106,107,108] CD8+ T cells [128] regulatory T cells [110] | Mixture [104,105]; LPC 16:0 > LPC 18:0 > LPC 14:0 > LPC 18:1 [106]; LPC 16:0 [107,108,109]; LPC 11:0 [128]; Not listed [110] |
| Increased cytotoxic activity towards tumor cells [129] | NK cells | Not listed [129] |
| Inhibition of histamine release [100] | mast cells | LPC 16:0 |
| Increased bactericidal activity [130], increased reactive oxygen species (ROS) production [93,131]; Decreased ROS production [94]; Inhibition of activation and effector functions [95] | neutrophils (PMNL) | Mixture [93]; LPC 18:0 [130]; Mixture, LPC 14:0, LPC 16:0, LPC 18:0 [131]; LPC 16:0, LPC 18:0, LPC 18:1 [94]; LPC 18:0 [95] |
| Inhibition of migration and effector functions [98] | eosinophils | LPC 16:0, LPC 18:0 [98] |
| Tumor cell apoptosis [132]; Reduction in tumor cell migration and adhesion [133] | tumor cells | Mixture [132]; LPC 16:0, LPC 18:0 [133] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knuplez, E.; Marsche, G. An Updated Review of Pro- and Anti-Inflammatory Properties of Plasma Lysophosphatidylcholines in the Vascular System. Int. J. Mol. Sci. 2020, 21, 4501. https://doi.org/10.3390/ijms21124501
Knuplez E, Marsche G. An Updated Review of Pro- and Anti-Inflammatory Properties of Plasma Lysophosphatidylcholines in the Vascular System. International Journal of Molecular Sciences. 2020; 21(12):4501. https://doi.org/10.3390/ijms21124501
Chicago/Turabian StyleKnuplez, Eva, and Gunther Marsche. 2020. "An Updated Review of Pro- and Anti-Inflammatory Properties of Plasma Lysophosphatidylcholines in the Vascular System" International Journal of Molecular Sciences 21, no. 12: 4501. https://doi.org/10.3390/ijms21124501
APA StyleKnuplez, E., & Marsche, G. (2020). An Updated Review of Pro- and Anti-Inflammatory Properties of Plasma Lysophosphatidylcholines in the Vascular System. International Journal of Molecular Sciences, 21(12), 4501. https://doi.org/10.3390/ijms21124501