Sharing Marks: H3K4 Methylation and H2B Ubiquitination as Features of Meiotic Recombination and Transcription



Abstract
:1. Overview of Some Molecular Events Occurring during Meiosis in Yeast
1.1. Transcriptional Events Leading to Meiosis
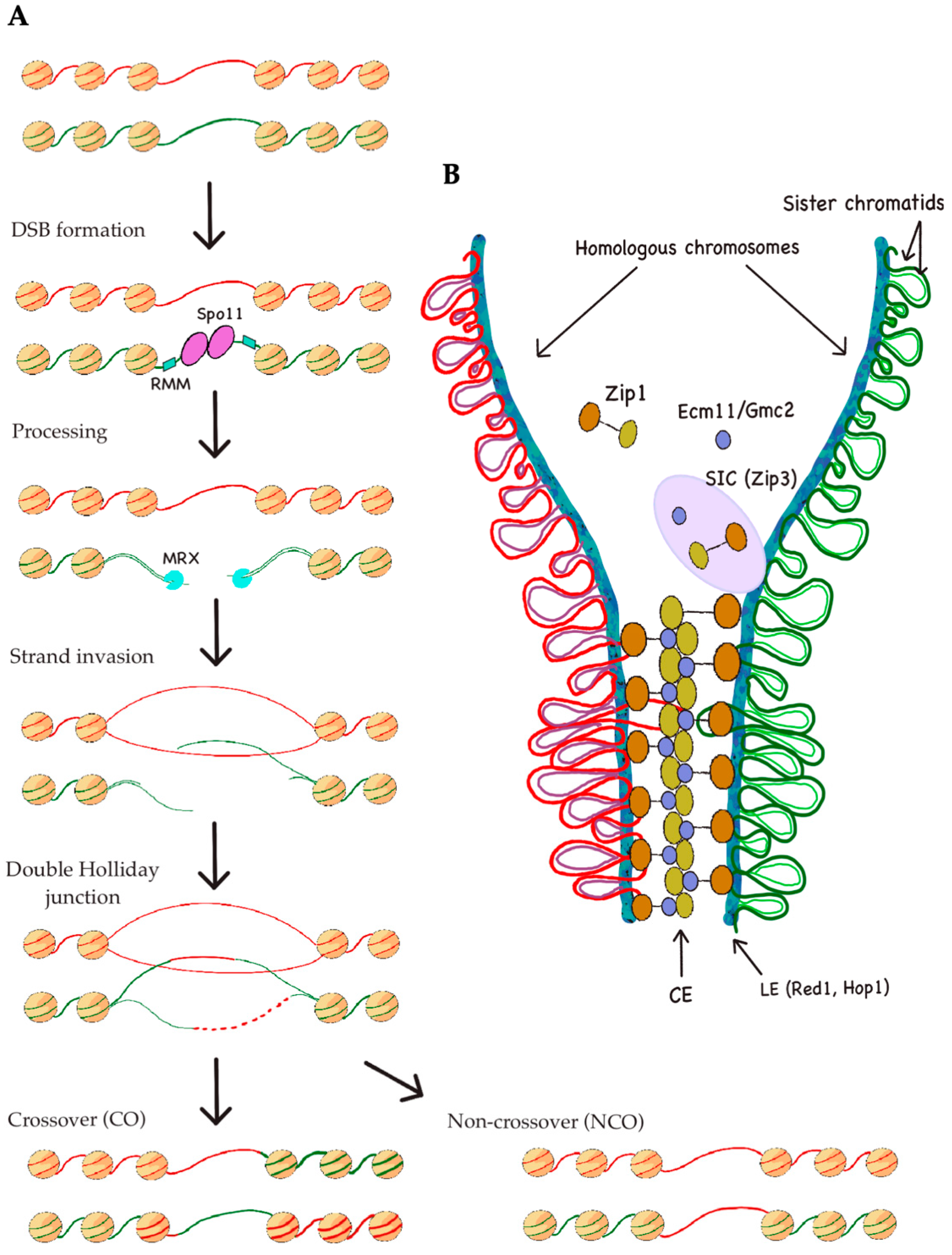
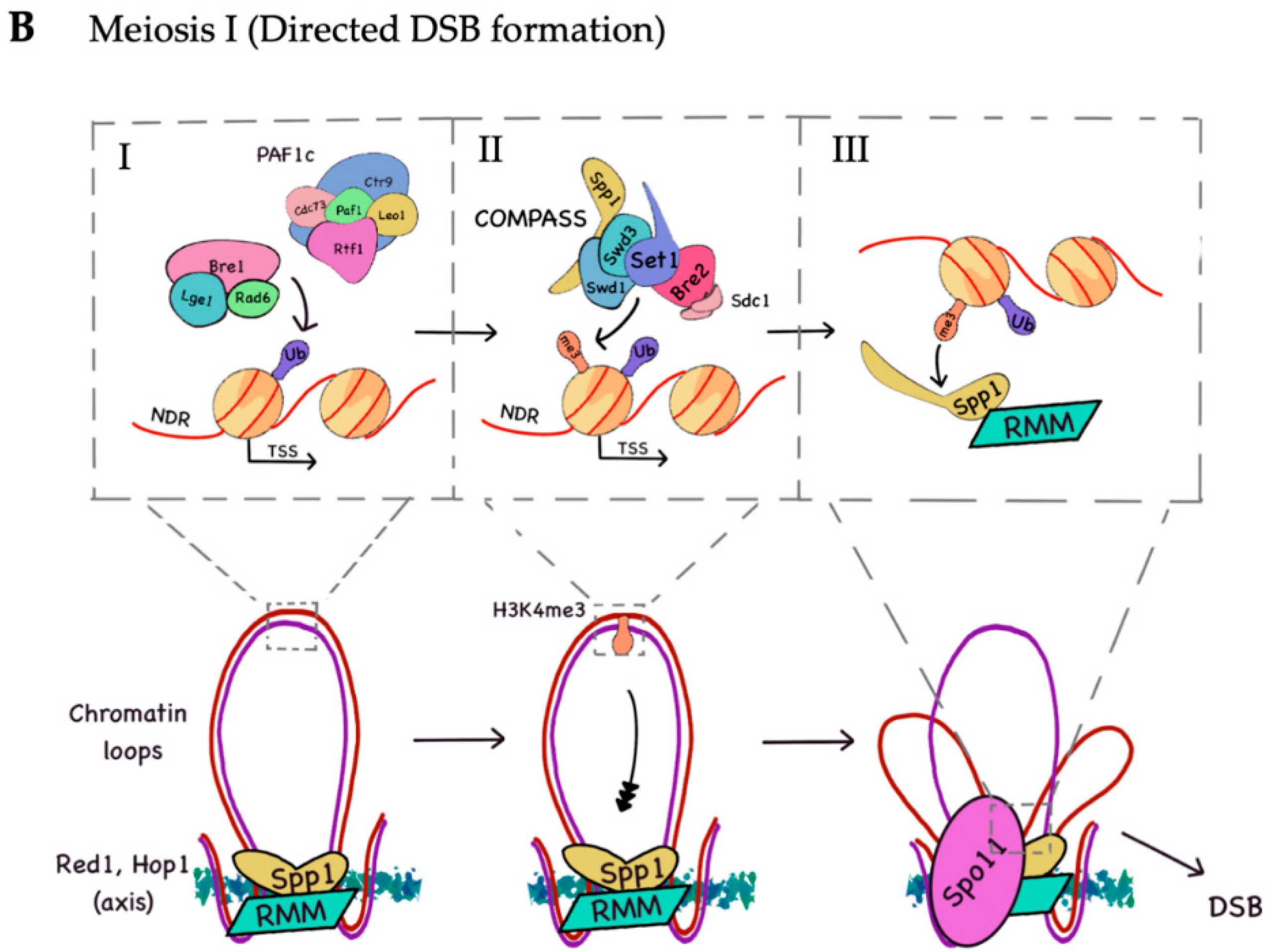
1.2. Meiotic Recombination: DSB Formation, Repair, and Distribution
2. Different Histone Modifications That Are Linked to Meiotic Events
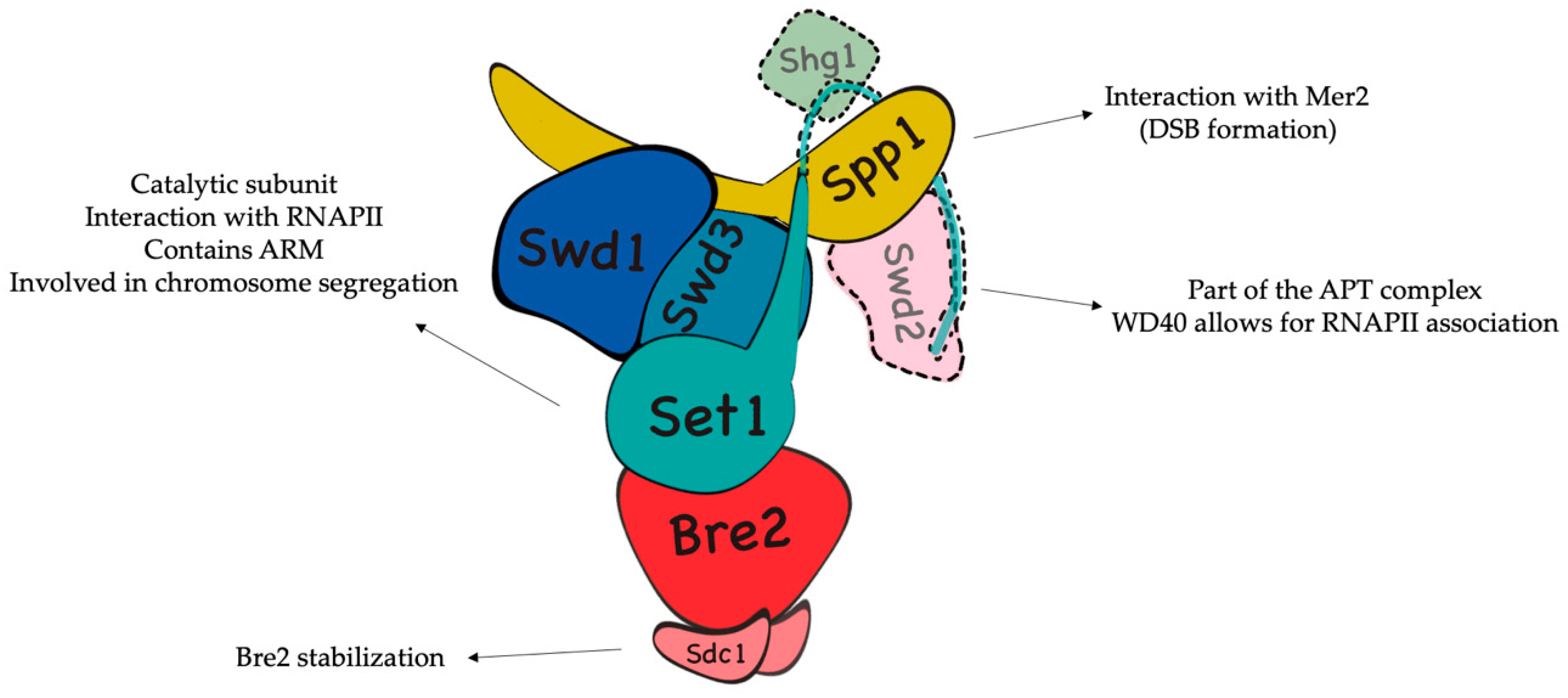
2.1. H3K4 Trimethylation by the COMPASS Complex Plays an Important Role in Recombination Initiation during Meiosis and DSB Generation
2.2. H2B Ubiquitination Is Important for DSB Formation
3. H3K4me and H2Bub Are Crucial Histone Marks during Transcription: Structural Insights into the Molecular Mechanism behind H3K4me/H2Bub Coordination
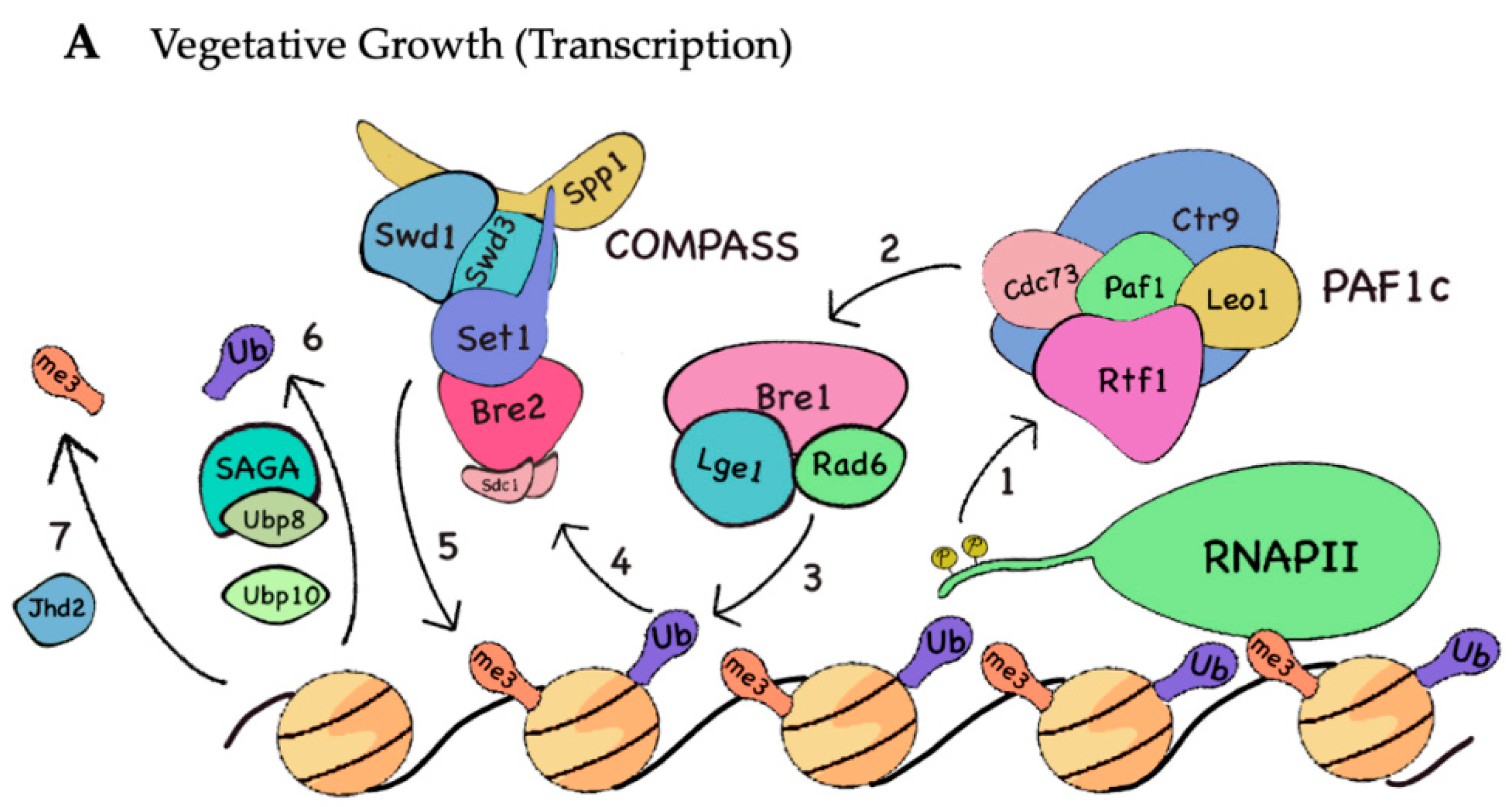
3.1. H3K4me during Transcription
3.2. H2B Monoubiquitination during Transcription
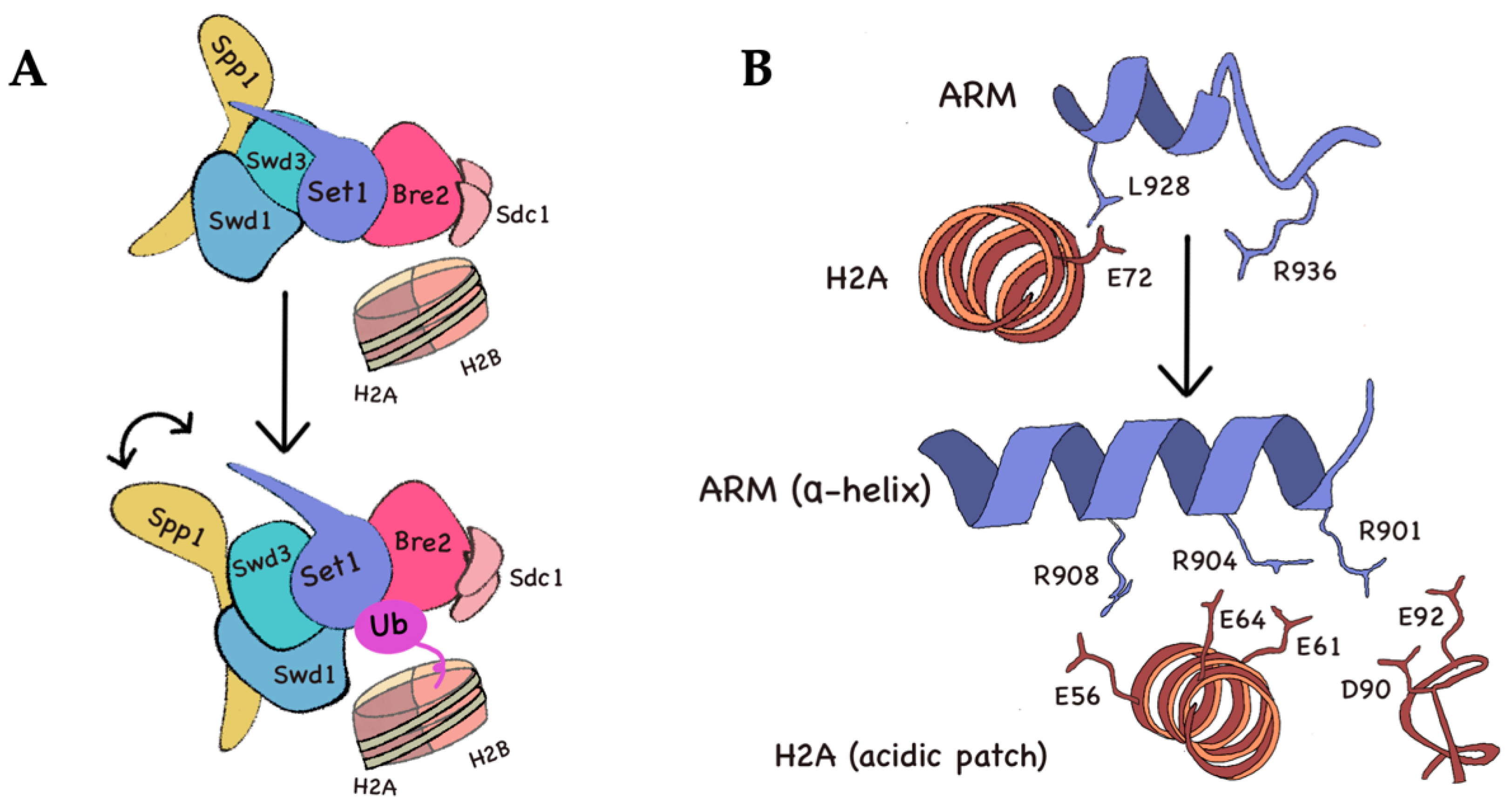
3.3. Structural Overview of COMPASS Activation upon H2B Ubiquitination
4. Human Diseases Associated to Defects in H3K4me and H2Bub Machineries
5. Future Directions and Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DSB | Double-strand break |
| PTM | Posttranslational modification |
| COMPASS | Complex of Proteins Associated with Set1 |
| PAF1c | RNA polymerase II associated factor 1 complex |
| cryo-EM | Cryo-electron microscopy |
| RRE | Rme1 repressor element |
| lncRNA | Long non-coding RNA |
| APC/C | Anaphase-promoting complex/cyclosome |
| SAGA | Spt-Ada-Gcn5 acetyltransferase |
| CO | Crossover |
| NCO | Non-crossover |
| RMM | Mer2-Mei4-Rec114 |
| MRX | Mre11-Rad50-Xrs2 |
| SC | Synaptonemal complex |
| CE | Central element |
| LE | Lateral elements |
| SIC | Synapsis initiation complex |
| SPB | Spindle pole body |
| NDR | Nucleosome-depleted region |
| SAM | S-adenosine-methionine |
| CM | Catalytic module |
| RRM | RNA-recognition motif |
| APT | Associated with Pta1 |
| PHD | Plant homeodomain |
| DUBm | Deubiquitinating module |
| FACT | Facilitates chromatin transcription |
| CENP-A | Centromere protein A |
| TSS | Transcription start site |
| TFIID | Transcription factor II D |
| RNAPII | RNA polymerase II |
| CTD | C-terminal domain |
| HMD | Histone modification domain |
| ARM | Arginine-rich motif |
| ESC | Embryonic stem cell |
| iPSC | Induced pluripotent stem cell |
| PC | Parathyroid carcinoma |
| ROS | Reactive oxygen species |
| PD2 | Pancreatic differentiation 2 |
References
- Piekarska, I.; Rytka, J.; Rempola, B. Regulation of sporulation in the yeast Saccharomyces cerevisiae. Acta Biochim. Pol. 2010, 57, 241–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neiman, A.M. Sporulation in the Budding Yeast Saccharomyces cerevisiae. Genetics 2011, 189, 737–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owens, S.; Tang, S.; Hunter, N. Monitoring Recombination During Meiosis in Budding Yeast. Methods Enzymol. 2018, 601, 275–307. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Neiman, A.M. Post-Transcriptional regulation in budding yeast meiosis. Curr. Genet. 2015, 62, 313–315. [Google Scholar] [CrossRef] [Green Version]
- Kassir, Y.; Simchen, G. Regulation of Mating and Meiosis in Yeast by the Mating-Type Region. Genetics 1976, 82, 187–206. [Google Scholar]
- Mitchell, A.P.; Herskowitz, I. Activation of meiosis and sporulation by repression of the RME1 product in yeast. Nature 1986, 319, 738–742. [Google Scholar] [CrossRef]
- Sagee, S.; Sherman, A.; Shenhar, G.; Robzyk, K.; Ben-Doy, N.; Simchen, G.; Kassir, Y. Multiple and Distinct Activation and Repression Sequences Mediate the Regulated Transcription of IME1, a Transcriptional Activator of Meiosis-Specific Genes inSaccharomyces cerevisiae. Mol. Cell. Biol. 1998, 18, 1985–1995. [Google Scholar] [CrossRef] [Green Version]
- Van Werven, F.J.; Neuert, G.; Hendrick, N.; Lardenois, A.; Buratowski, S.; Van Oudenaarden, A.; Primig, M.; Amon, A. Transcription of Two Long Noncoding RNAs Mediates Mating-Type Control of Gametogenesis in Budding Yeast. Cell 2012, 150, 1170–1181. [Google Scholar] [CrossRef] [Green Version]
- Clancy, M.J.; Shambaugh, M.; Timpte, C.; Bokar, J. Induction of sporulation in Saccharomyces cerevisiae leads to the formation of N6-methyladenosine in mRNA: A potential mechanism for the activity of the IME4 gene. Nucleic Acids Res. 2002, 30, 4509–4518. [Google Scholar] [CrossRef] [Green Version]
- Bodi, Z.; Button, J.D.; Grierson, D.; Fray, R. Yeast targets for mRNA methylation. Nucleic Acids Res. 2010, 38, 5327–5335. [Google Scholar] [CrossRef] [Green Version]
- Hongay, C.F.; Grisafi, P.L.; Galitski, T.; Fink, G.R. Antisense Transcription Controls Cell Fate in Saccharomyces cerevisiae. Cell 2006, 127, 735–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govin, J.; Berger, S.L. Genome reprogramming during sporulation. Int. J. Dev. Biol. 2009, 53, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Strich, R.; Steber, C.; Messenguy, F.; Surosky, R.T.; Dubois, E.; Esposito, R.E. UME6 is a key regulator of nitrogen repression and meiotic development. Genes Dev. 1994, 8, 796–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowdish, K.S.; Mitchell, A.P. Bipartite structure of an early meiotic upstream activation sequence from Saccharomyces cerevisiae. Mol. Cell. Biol. 1993, 13, 2172–2181. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S.F.; Steber, C.M.; Esposito, R.E.; Coleman, J.E. UME6, a negative regulator of meiosis insaccharomyces cerevisiae, contains a C-terminal Zn2Cys6binuclear cluster that binds the URS1 DNA sequence in a zinc-dependent manner. Protein Sci. 1995, 4, 1832–1843. [Google Scholar] [CrossRef] [Green Version]
- Kadosh, D.; Struhl, K. Repression by Ume6 Involves Recruitment of a Complex Containing Sin3 Corepressor and Rpd3 Histone Deacetylase to Target Promoters. Cell 1997, 89, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Goldmark, J.P.; Fazzio, T.G.; Estep, P.W.; Church, G.M.; Tsukiyama, T. The Isw2 Chromatin Remodeling Complex Represses Early Meiotic Genes upon Recruitment by Ume6p. Cell 2000, 103, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Yukawa, M.; Yo, K.; Hasegawa, H.; Ueno, M.; Tsuchiya, E. The Rpd3/HDAC Complex Is Present at the URS1cis-Element with Hyperacetylated Histone H3. Biosci. Biotechnol. Biochem. 2009, 73, 378–384. [Google Scholar] [CrossRef]
- Washburn, B.K.; Esposito, R.E. Identification of the Sin3-Binding Site in Ume6 Defines a Two-Step Process for Conversion of Ume6 from a Transcriptional Repressor to an Activator in Yeast. Mol. Cell. Biol. 2001, 21, 2057–2069. [Google Scholar] [CrossRef] [Green Version]
- Mallory, M.J.; Cooper, K.F.; Strich, R. Meiosis-Specific Destruction of the Ume6p Repressor by the Cdc20-Directed APC/C. Mol. Cell 2007, 27, 951–961. [Google Scholar] [CrossRef] [Green Version]
- Grant, P.A.; Duggan, L.; Côté, J.; Roberts, S.M.; Brownell, J.E.; Candau, R.; Ohba, R.; Owen-Hughes, T.; Allis, C.D.; Winston, F.; et al. Yeast Gcn5 functions in two multisubunit complexes to acetylate nucleosomal histones: Characterization of an Ada complex and the SAGA (Spt/Ada) complex. Genome Res. 1997, 11, 1640–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallory, M.J.; Law, M.J.; Sterner, D.E.; Berger, S.L.; Strich, R. Gcn5p-dependent acetylation induces degradation of the meiotic transcriptional repressor Ume6p. Mol. Biol. Cell 2012, 23, 1609–1617. [Google Scholar] [CrossRef] [PubMed]
- Law, M.J.; Mallory, M.J.; Dunbrack, R.L.; Strich, R. Acetylation of the Transcriptional Repressor Ume6p Allows Efficient Promoter Release and Timely Induction of the Meiotic Transient Transcription Program in Yeast. Mol. Cell. Biol. 2013, 34, 631–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Primig, M.; Williams, R.M.; Winzeler, E.A.; Tevzadze, G.G.; Conway, A.R.; Hwang, S.Y.; Davis, R.W.; Esposito, R.E. The core meiotic transcriptome in budding yeasts. Nat. Genet. 2000, 26, 415–423. [Google Scholar] [CrossRef]
- Slekar, K.H.; Henry, S.A. SIN3 works through two different promoter elements to regulate INO1 gene expression in yeast. Nucleic Acids Res. 1995, 23, 1964–1969. [Google Scholar] [CrossRef] [Green Version]
- Petronczki, M.; Siomos, M.F.; Nasmyth, K. Un menage a quatre: The molecular biology of chromosome segregation in meiosis. Cell 2003, 112, 423–440. [Google Scholar] [CrossRef] [Green Version]
- Lichten, M.; De Massy, B. The Impressionistic Landscape of Meiotic Recombination. Cell 2011, 147, 267–270. [Google Scholar] [CrossRef] [Green Version]
- Lane, S.I.R.; Kauppi, L. Meiotic spindle assembly checkpoint and aneuploidy in males versus females. Cell. Mol. Life Sci. 2018, 76, 1135–1150. [Google Scholar] [CrossRef] [Green Version]
- Martini, E.; Borde, V.; Legendre, M.; Audic, S.; Regnault, B.; Soubigou, G.; Dujon, B.; Llorente, B. Genome-Wide Analysis of Heteroduplex DNA in Mismatch Repair–Deficient Yeast Cells Reveals Novel Properties of Meiotic Recombination Pathways. PLoS Genet. 2011, 7, e1002305. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Sasaki, M.; Kniewel, R.; Murakami, H.; Blitzblau, H.G.; Tischfield, S.E.; Zhu, X.; Neale, M.J.; Jasin, M.; Socci, N.D.; et al. A Hierarchical Combination of Factors Shapes the Genome-wide Topography of Yeast Meiotic Recombination Initiation. Cell 2011, 144, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Allers, T.; Lichten, M. Differential Timing and Control of Noncrossover and Crossover Recombination during Meiosis. Cell 2001, 106, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Bergerat, A.; De Massy, B.; Gadelle, D.; Varoutas, P.-C.; Nicolas, A.; Forterre, P. An atypical topoisomerase II from archaea with implications for meiotic recombination. Nature 1997, 386, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Keeney, S.; Giroux, C.N.; Kleckner, N. Meiosis-Specific DNA Double-Strand Breaks Are Catalyzed by Spo11, a Member of a Widely Conserved Protein Family. Cell 1997, 88, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Panizza, S.; Mendoza-Parra, M.A.; Berlinger, M.; Huang, L.; Nicolas, A.; Shirahige, K.; Klein, F. Spo11-Accessory Proteins Link Double-Strand Break Sites to the Chromosome Axis in Early Meiotic Recombination. Cell 2011, 146, 372–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, I.; Keeney, S. Mechanism and Regulation of Meiotic Recombination Initiation. Cold Spring Harb. Perspect. Biol. 2014, 7, a016634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Székvölgyi, L.; Ohta, K.; Nicolas, A. Initiation of Meiotic Homologous Recombination: Flexibility, Impact of Histone Modifications, and Chromatin Remodeling. Cold Spring Harb. Perspect. Biol. 2015, 7, a016527. [Google Scholar] [CrossRef] [Green Version]
- Maleki, S.; Neale, M.J.; Arora, C.; Henderson, K.A.; Keeney, S. Interactions between Mei4, Rec114, and other proteins required for meiotic DNA double-strand break formation in Saccharomyces cerevisiae. Chromosoma 2007, 116, 471–486. [Google Scholar] [CrossRef] [Green Version]
- Voelkel-Meiman, K.; Moustafa, S.S.; Lefrançois, P.; Villeneuve, A.M.; MacQueen, A.J. Full-Length Synaptonemal Complex Grows Continuously during Meiotic Prophase in Budding Yeast. PLoS Genet. 2012, 8, e1002993. [Google Scholar] [CrossRef]
- Pyatnitskaya, A.; Borde, V.; De Muyt, A. Crossing and zipping: Molecular duties of the ZMM proteins in meiosis. Chromosoma 2019, 128, 181–198. [Google Scholar] [CrossRef] [Green Version]
- Klein, F.; Mahr, P.; Galova, M.; Buonomo, S.C.B.; Michaelis, C.; Nairz, K.; Nasmyth, K. A Central Role for Cohesins in Sister Chromatid Cohesion, Formation of Axial Elements, and Recombination during Yeast Meiosis. Cell 1999, 98, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Ma, O.X.; Cai, S.; Shi, J.; Gan, L. Cryo-ET analysis of budding yeast synaptonemal complexes in situ. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Humphryes, N.; Leung, W.-K.; Argunhan, B.; Terentyev, Y.; Dvorácková, M.; Tsubouchi, H. The Ecm11-Gmc2 Complex Promotes Synaptonemal Complex Formation through Assembly of Transverse Filaments in Budding Yeast. PLoS Genet. 2013, 9, e1003194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zickler, D.; Kleckner, N. Recombination, Pairing, and Synapsis of Homologs during Meiosis. Cold Spring Harb. Perspect. Biol. 2015, 7, a016626. [Google Scholar] [CrossRef] [Green Version]
- Voelkel-Meiman, K.; Johnston, C.; Thappeta, Y.; Subramanian, V.V.; Hochwagen, A.; MacQueen, A.J. Separable Crossover-Promoting and Crossover-Constraining Aspects of Zip1 Activity during Budding Yeast Meiosis. PLoS Genet. 2015, 11, e1005335. [Google Scholar] [CrossRef] [Green Version]
- Voelkel-Meiman, K.; Cheng, S.-Y.; Morehouse, S.J.; MacQueen, A.J. Synaptonemal Complex Proteins of Budding Yeast Define Reciprocal Roles in MutSγ-Mediated Crossover Formation. Genetics 2016, 203, 1091–1103. [Google Scholar] [CrossRef] [Green Version]
- Cavanaugh, A.M.; Jaspersen, S.L. Big Lessons from Little Yeast: Budding and Fission Yeast Centrosome Structure, Duplication, and Function. Annu. Rev. Genet. 2017, 51, 361–383. [Google Scholar] [CrossRef] [PubMed]
- Jaspersen, S.L.; Winey, M. The budding yeast spindle pole body: Structure, duplication, and function. Annu. Rev. Cell Dev. Biol. 2004, 20, 1–28. [Google Scholar] [CrossRef]
- Jenni, S.; Harrison, S.C. Structure of the DASH/Dam1 complex shows its role at the yeast kinetochore-microtubule interface. Science 2018, 360, 552–558. [Google Scholar] [CrossRef] [Green Version]
- Villoria, M.T.; Ramos, F.; Dueñas, E.; Faull, P.; Cutillas, P.R.; Clemente-Blanco, A. Stabilization of the metaphase spindle by Cdc14 is required for recombinational DNA repair. EMBO J. 2016, 36, 79–101. [Google Scholar] [CrossRef]
- Storey, A.J.; Wang, H.-P.; Protacio, R.U.; Davidson, M.K.; Tackett, A.J.; Wahls, W.P. Chromatin-mediated regulators of meiotic recombination revealed by proteomics of a recombination hotspot. Epigenet. Chromatin 2018, 11, 64. [Google Scholar] [CrossRef] [Green Version]
- Wahls, W.P.; Davidson, M.K. Discrete DNA sites regulate global distribution of meiotic recombination. Trends Genet. 2010, 26, 202–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouzarides, T. Chromatin Modifications and Their Function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornberg, R.D. Chromatin Structure: A Repeating Unit of Histones and DNA. Science 1974, 184, 868–871. [Google Scholar] [CrossRef] [PubMed]
- Luense, L.J.; Wang, X.; Schon, S.B.; Weller, A.H.; Lin-Shiao, E.; Bryant, J.; Bartolomei, M.S.; Coutifaris, C.; Garcia, B.A.; Berger, S.L. Comprehensive analysis of histone post-translational modifications in mouse and human male germ cells. Epigenet. Chromatin 2016, 9, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Xu, Z.; Khawar, M.B.; Liu, C.; Li, W. The histone codes for meiosis. Reproduction 2017, 154, R65–R79. [Google Scholar] [CrossRef]
- Yamada, S.; Ohta, K.; Yamada, T. Acetylated Histone H3K9 is associated with meiotic recombination hotspots, and plays a role in recombination redundantly with other factors including the H3K4 methylase Set1 in fission yeast. Nucleic Acids Res. 2013, 41, 3504–3517. [Google Scholar] [CrossRef] [Green Version]
- Cavero, S.; Herruzo, E.; Ontoso, D.; San-Segundo, P.A. Impact of histone H4K16 acetylation on the meiotic recombination checkpoint in Saccharomyces cerevisiae. Microb. Cell 2016, 3, 606–620. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Donahue, G.; Dorsey, J.; Govin, J.; Yuan, Z.; Garcia, B.A.; Shah, P.P.; Berger, S.L. H4K44 Acetylation Facilitates Chromatin Accessibility during Meiosis. Cell Rep. 2015, 13, 1772–1780. [Google Scholar] [CrossRef] [Green Version]
- Acquaviva, L.; Székvölgyi, L.; Dichtl, B.; Dichtl, B.S.; de la Andre, C.R.S.; Nicolas, A.; Géli, V. The COMPASS Subunit Spp1 Links Histone Methylation to Initiation of Meiotic Recombination. Science 2012, 339, 215–218. [Google Scholar] [CrossRef]
- Sommermeyer, V.; Béneut, C.; Chaplais, E.; Serrentino, M.E.; Borde, V. Spp1, a Member of the Set1 Complex, Promotes Meiotic DSB Formation in Promoters by Tethering Histone H3K4 Methylation Sites to Chromosome Axes. Mol. Cell 2013, 49, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Buard, J.; Barthès, P.; Grey, C.; De Massy, B. Distinct histone modifications define initiation and repair of meiotic recombination in the mouse. EMBO J. 2009, 28, 2616–2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, K.; Shinohara, M.; Shinohara, A. Rad6-Bre1-mediated histone H2B ubiquitylation modulates the formation of double-strand breaks during meiosis. Proc. Natl. Acad. Sci. USA 2004, 101, 11380–11385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, Y. Geometry and force behind kinetochore orientation: Lessons from meiosis. Nat. Rev. Mol. Cell Biol. 2012, 13, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ma, H.; Pugh, B.F. Stable and dynamic nucleosome states during a meiotic developmental process. Genome Res. 2011, 21, 875–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, L.M.; Radman-Livaja, M.; Lin, S.G.; Rando, O.J.; Buratowski, S. Feedback control of Set1 protein levels is important for proper H3K4 methylation patterns. Cell Rep. 2014, 6, 961–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruthenburg, A.J.; Allis, C.D.; Wysocka, J. Methylation of Lysine 4 on Histone H3: Intricacy of Writing and Reading a Single Epigenetic Mark. Mol. Cell 2007, 25, 15–30. [Google Scholar] [CrossRef]
- Shilatifard, A. The COMPASS family of histone H3K4 methylases: Mechanisms of regulation in development and disease pathogenesis. Annu. Rev. Biochem. 2012, 81, 65–95. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, Y.-H.; Shilatifard, A. Structural basis for H3K4 trimethylation by yeast Set1/COMPASS. Adv. Enzym. Regul. 2009, 50, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Miller, T.; Krogan, N.J.; Dover, J.; Erdjument-Bromage, H.; Tempst, P.; Johnston, M.; Greenblatt, J.F.; Shilatifard, A. Compass: A complex of proteins associated with a trithorax-related SET domain protein. Proc. Natl. Acad. Sci. USA 2001, 98, 12902–12907. [Google Scholar] [CrossRef] [Green Version]
- Roguev, A.; Schaft, D.; Shevchenko, A.; Pijnappel, W.; Wilm, M.; Aasland, R.; Stewart, A.F. The Saccharomyces cerevisiae Set1 complex includes an Ash2 homologue and methylates histone 3 lysine 4. EMBO J. 2001, 20, 7137–7148. [Google Scholar] [CrossRef]
- Ramakrishnan, S.; Pokhrel, S.; Palani, S.; Pflueger, C.; Parnell, T.J.; Cairns, B.R.; Bhaskara, S.; Chandrasekharan, M.B. Counteracting H3K4 methylation modulators Set1 and Jhd2 co-regulate chromatin dynamics and gene transcription. Nat. Commun. 2016, 7, 11949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, R.; Singh, S.; Arumugam, S.; Roguev, A.; Stewart, A.F. The Set1 complex is dimeric and acts with Jhd2 demethylation to convey symmetrical H3K4 trimethylation. Genome Res. 2019, 33, 550–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, P.L.; Li, H.; Lau, H.-T.; Leonen, C.; Dhall, A.; Ong, S.-E.; Chatterjee, C.; Zheng, N. Crystal Structure of the COMPASS H3K4 Methyltransferase Catalytic Module. Cell 2018, 174, 1106–1116.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, Q.; Takahashi, Y.-H.; Yang, Y.; Hu, H.; Zhang, Y.; Brunzelle, J.S.; Couture, J.-F.; Shilatifard, A.; Skiniotis, G. Structure and Conformational Dynamics of a COMPASS Histone H3K4 Methyltransferase Complex. Cell 2018, 174, 1117–1126.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kim, J.-A.; McGinty, R.K.; Nguyen, U.T.T.; Muir, T.W.; Allis, C.D.; Roeder, R.G. The n-SET domain of Set1 regulates H2B ubiquitylation-dependent H3K4 methylation. Mol. Cell 2013, 49, 1121–1133. [Google Scholar] [CrossRef] [PubMed]
- Dehe, P.-M.; Dichtl, B.; Schaft, D.; Roguev, A.; Pamblanco, M.; Lebrun, R.; Gil, A.R.; Mkandawire, M.; Landsberg, K.; Shevchenko, A.; et al. Protein Interactions within the Set1 Complex and Their Roles in the Regulation of Histone 3 Lysine 4 Methylation. J. Biol. Chem. 2006, 281, 35404–35412. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; He, X.; Moore, C.L. The Essential WD Repeat Protein Swd2 Has Dual Functions in RNA Polymerase II Transcription Termination and Lysine 4 Methylation of Histone H3. Mol. Cell. Biol. 2004, 24, 2932–2943. [Google Scholar] [CrossRef] [Green Version]
- Dichtl, B.; Aasland, R.; Keller, W. Functions for S. cerevisiae Swd2p in 3′ end formation of specific mRNAs and snoRNAs and global histone 3 lysine 4 methylation. RNA 2004, 10, 965–977. [Google Scholar] [CrossRef] [Green Version]
- Nedea, E.; He, X.; Kim, M.; Pootoolal, J.; Zhong, G.; Canadien, V.; Hughes, T.; Buratowski, S.; Moore, C.L.; Greenblatt, J.F. Organization and Function of APT, a Subcomplex of the Yeast Cleavage and Polyadenylation Factor Involved in the Formation of mRNA and Small Nucleolar RNA 3′-Ends. J. Biol. Chem. 2003, 278, 33000–33010. [Google Scholar] [CrossRef] [Green Version]
- Acquaviva, L.; Drogat, J.; Dehe, P.-M.; de la Saint-André, C.R.; Géli, V. Spp1 at the crossroads of H3K4me3 regulation and meiotic recombination. Epigenetics 2013, 8, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Adam, C.; Guerois, R.; Citarella, A.; Verardi, L.; Adolphe, F.; Beneut, C.; Sommermeyer, V.; Ramus, C.; Govin, J.; Couté, Y.; et al. The PHD finger protein Spp1 has distinct functions in the Set1 and the meiotic DSB formation complexes. PLoS Genet. 2018, 14, e1007223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karányi, Z.; Halász, L.; Acquaviva, L.; Jónás, D.; Hetey, S.; Boros-Oláh, B.; Peng, F.; Chen, D.; Klein, F.; Geli, V.; et al. Nuclear dynamics of the Set1C subunit Spp1 prepares meiotic recombination sites for break formation. J. Cell Biol. 2018, 217, 3398–3415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borde, V.; Robine, N.; Lin, W.; Bonfils, S.; Géli, V.; Nicolas, A. Histone H3 lysine 4 trimethylation marks meiotic recombination initiation sites. EMBO J. 2008, 28, 99–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Suzuki, T.; Li, K.; Gothwal, S.K.; Shinohara, M.; Shinohara, A. Genetic Interactions of Histone Modification Machinery Set1 and PAF1C with the Recombination Complex Rec114-Mer2-Mei4 in the Formation of Meiotic DNA Double-Strand Breaks. Int. J. Mol. Sci. 2020, 21, 2679. [Google Scholar] [CrossRef] [Green Version]
- Robine, N.; Uematsu, N.; Amiot, F.; Gidrol, X.; Barillot, E.; Nicolas, A.; Borde, V. Genome-Wide Redistribution of Meiotic Double-Strand Breaks in Saccharomyces cerevisiae. Mol. Cell. Biol. 2006, 27, 1868–1880. [Google Scholar] [CrossRef] [Green Version]
- Baudat, F.; Buard, J.; Grey, C.; Fledel-Alon, A.; Ober, C.; Przeworski, M.; Coop, G.; De Massy, B. PRDM9 Is a Major Determinant of Meiotic Recombination Hotspots in Humans and Mice. Science 2009, 327, 836–840. [Google Scholar] [CrossRef] [Green Version]
- Robzyk, K.; Recht, J.; Osley, M.A. Rad6-Dependent Ubiquitination of Histone H2B in Yeast. Science 2000, 287, 501–504. [Google Scholar] [CrossRef]
- Wood, A.; Krogan, N.J.; Dover, J.; Schneider, J.; Heidt, J.; Boateng, M.A.; Dean, K.; Golshani, A.; Zhang, Y.; Greenblatt, J.F.; et al. Bre1, an E3 Ubiquitin Ligase Required for Recruitment and Substrate Selection of Rad6 at a Promoter. Mol. Cell 2003, 11, 267–274. [Google Scholar] [CrossRef]
- Schulze, J.M.; Hentrich, T.; Nakanishi, S.; Gupta, A.; Emberly, E.; Shilatifard, A.; Kobor, M.S. Splitting the task: Ubp8 and Ubp10 deubiquitinate different cellular pools of H2BK123. Genes Dev. 2011, 25, 2242–2247. [Google Scholar] [CrossRef] [Green Version]
- Koutelou, E.; Hirsch, C.L.; Dent, S.Y. Multiple faces of the SAGA complex. Curr. Opin. Cell Biol. 2010, 22, 374–382. [Google Scholar] [CrossRef] [Green Version]
- Wood, A.; Schneider, J.; Dover, J.; Johnston, M.; Shilatifard, A. The Paf1 Complex Is Essential for Histone Monoubiquitination by the Rad6-Bre1 Complex, Which Signals for Histone Methylation by COMPASS and Dot1p. J. Biol. Chem. 2003, 278, 34739–34742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, H.H.; Dole, S.; Struhl, K.; Gavrilova, O.; Haluzík, M.; Matsusue, K.; Cutson, J.J.; Johnson, L.; Dietz, K.R.; Nicol, C.J.; et al. The Rtf1 Component of the Paf1 Transcriptional Elongation Complex Is Required for Ubiquitination of Histone H2B. J. Biol. Chem. 2003, 278, 33625–33628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavri, R.; Zhu, B.; Li, G.; Trojer, P.; Mandal, S.; Shilatifard, A.; Reinberg, D. Histone H2B Monoubiquitination Functions Cooperatively with FACT to Regulate Elongation by RNA Polymerase II. Cell 2006, 125, 703–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Song, Z.; Li, G.; Tu, H.; Liu, W.; Liu, Y.; Wang, P.; Wang, Y.; Cui, X.; Liu, C.; et al. H2B ubiquitination regulates meiotic recombination by promoting chromatin relaxation. Nucleic Acids Res. 2016, 44, 9681–9697. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Cao, C.; Wang, F.; Zhao, J.; Li, W. H2B ubiquitination: Conserved molecular mechanism, diverse physiologic functions of the E3 ligase during meiosis. Nucleus 2017, 8, 461–468. [Google Scholar] [CrossRef] [Green Version]
- Fierz, B.; Chatterjee, C.; McGinty, R.K.; Bar-Dagan, M.; Raleigh, D.P.; Muir, T.W. Histone H2B ubiquitylation disrupts local and higher-order chromatin compaction. Nat. Methods 2011, 7, 113–119. [Google Scholar] [CrossRef]
- Roest, H.; Van Klaveren, J.; De Wit, J.; Van Gurp, C.; Koken, M.; Vermey, M.; Van Roijen, J.; Hoogerbrugge, J.; Vreeburg, J.; Baarends, W.; et al. Inactivation of the HR6B Ubiquitin-Conjugating DNA Repair Enzyme in Mice Causes Male Sterility Associated with Chromatin Modification. Cell 1996, 86, 799–810. [Google Scholar] [CrossRef] [Green Version]
- Jordan, P.W.; Klein, F.; Leach, D.R.F. Novel Roles for Selected Genes in Meiotic DNA Processing. PLoS Genet. 2007, 3, e222. [Google Scholar] [CrossRef] [Green Version]
- Gothwal, S.K.; Patel, N.J.; Colletti, M.M.; Sasanuma, H.; Shinohara, M.; Hochwagen, A.; Shinohara, A. The Double-Strand Break Landscape of Meiotic Chromosomes Is Shaped by the Paf1 Transcription Elongation Complex in Saccharomyces cerevisiae. Genetics 2015, 202, 497–512. [Google Scholar] [CrossRef] [Green Version]
- Latham, J.A.; Chosed, R.; Wang, S.; Dent, S.Y. Chromatin signaling to kinetochores: Transregulation of Dam1 methylation by histone H2B ubiquitination. Cell 2011, 146, 709–719. [Google Scholar] [CrossRef] [Green Version]
- Canzonetta, C.; Vernarecci, S.; Iuliani, M.; Marracino, C.; Belloni, C.; Ballario, P.; Filetici, P. SAGA DUB-Ubp8 Deubiquitylates Centromeric Histone Variant Cse4. G3 (Bethesda) 2015, 6, 287–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, M.; Mehta, G.; Ghosh, S.K. Role of Ctf3 and COMA subcomplexes in meiosis: Implication in maintaining Cse4 at the centromere and numeric spindle poles. Biochim. Biophys. Acta 2015, 1853, 671–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pray-Grant, M.G.; Schieltz, D.; McMahon, S.J.; Wood, J.M.; Kennedy, E.L.; Cook, R.G.; Workman, J.L.; Yates, J.R.; Grant, P.A.; Iii, J.R.Y. The Novel SLIK Histone Acetyltransferase Complex Functions in the Yeast Retrograde Response Pathway. Mol. Cell. Biol. 2002, 22, 8774–8786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barski, A.; Cuddapah, S.; Cui, K.; Roh, T.-Y.; Schones, D.E.; Wang, Z.; Wei, G.; Chepelev, I.; Zhao, K. High-Resolution Profiling of Histone Methylations in the Human Genome. Cell 2007, 129, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Soares, L.M.; He, P.C.; Chun, Y.; Suh, H.; Kim, T.; Buratowski, S. Determinants of Histone H3K4 Methylation Patterns. Mol. Cell 2017, 68, 773–785.e6. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, M.; Eberl, H.C.; Matarese, F.; Marks, H.; Denissov, S.; Butter, F.; Lee, K.K.; Olsen, J.V.; Hyman, A.A.; Stunnenberg, H.G.; et al. Quantitative Interaction Proteomics and Genome-wide Profiling of Epigenetic Histone Marks and Their Readers. Cell 2010, 142, 967–980. [Google Scholar] [CrossRef] [Green Version]
- Margaritis, T.; Oreal, V.; Brabers, N.; Maestroni, L.; Vitaliano-Prunier, A.; Benschop, J.J.; Van Hooff, S.; Van Leenen, D.; Dargemont, C.; Géli, V.; et al. Two Distinct Repressive Mechanisms for Histone 3 Lysine 4 Methylation through Promoting 3′-End Antisense Transcription. PLoS Genet. 2012, 8, e1002952. [Google Scholar] [CrossRef]
- Lee, J.-H.; Skalnik, D.G. Wdr82 Is a C-Terminal Domain-Binding Protein That Recruits the Setd1A Histone H3-Lys4 Methyltransferase Complex to Transcription Start Sites of Transcribed Human Genes. Mol. Cell. Biol. 2007, 28, 609–618. [Google Scholar] [CrossRef] [Green Version]
- Krogan, N.J.; Kim, M.; Tong, A.; Golshani, A.; Cagney, G.; Canadien, V.; Richards, D.P.; Beattie, B.K.; Emili, A.; Boone, C.; et al. Methylation of Histone H3 by Set2 in Saccharomyces cerevisiae Is Linked to Transcriptional Elongation by RNA Polymerase II. Mol. Cell. Biol. 2003, 23, 4207–4218. [Google Scholar] [CrossRef] [Green Version]
- Ng, H.H.; Robert, F.; Young, R.A.; Struhl, K. Targeted Recruitment of Set1 Histone Methylase by Elongating Pol II Provides a Localized Mark and Memory of Recent Transcriptional Activity. Mol. Cell 2003, 11, 709–719. [Google Scholar] [CrossRef]
- Bae, H.J.; Dubarry, M.; Jeon, J.; Soares, L.M.; Dargemont, C.; Kim, J.; Geli, V.; Buratowski, S. The Set1 N-terminal domain and Swd2 interact with RNA polymerase II CTD to recruit COMPASS. Nat. Commun. 2020, 11, 2181. [Google Scholar] [CrossRef] [PubMed]
- Howe, F.S.; Fischl, H.; Murray, S.C.; Mellor, J. Is H3K4me3 instructive for transcription activation? BioEssays 2016, 39, e201600095. [Google Scholar] [CrossRef] [PubMed]
- Pokholok, D.K.; Harbison, C.T.; Levine, S.S.; Cole, M.; Hannett, N.M.; Lee, T.I.; Bell, G.W.; Walker, K.; Rolfe, P.A.; Herbolsheimer, E.; et al. Genome-wide Map of Nucleosome Acetylation and Methylation in Yeast. Cell 2005, 122, 517–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luciano, P.; Jeon, J.; El-Kaoutari, A.; Challal, D.; Bonnet, A.; Barucco, M.; Candelli, T.; Jourquin, F.; Lesage, P.; Kim, J.; et al. Binding to RNA regulates Set1 function. Cell Discov. 2017, 3, 17040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglia, S.; Lidschreiber, M.; Baejen, C.; Torkler, P.; Vos, S.; Cramer, P. RNA-dependent chromatin association of transcription elongation factors and Pol II CTD kinases. ELife 2017, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shema, E.; Nikolov, M.; Haj-Yahya, M.; Siman, P.; Allemand, E.; Yamaguchi, Y.; Muchardt, C.; Urlaub, H.; Brik, A.; Oren, M.; et al. Systematic Identification of Proteins Binding to Chromatin-Embedded Ubiquitylated H2B Reveals Recruitment of SWI/SNF to Regulate Transcription. Cell Rep. 2013, 4, 601–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, A.B.; Kao, C.-F.; Hillyer, C.; Pikaart, M.; Osley, M.A. H2B Ubiquitylation Plays a Role in Nucleosome Dynamics during Transcription Elongation. Mol. Cell 2008, 31, 57–66. [Google Scholar] [CrossRef]
- Mao, P.; Meas, R.; Dorgan, K.M.; Smerdon, M.J. UV damage-induced RNA polymerase II stalling stimulates H2B deubiquitylation. Proc. Natl. Acad. Sci. USA 2014, 111, 12811–12816. [Google Scholar] [CrossRef] [Green Version]
- Jaehning, J.A. The Paf1 complex: Platform or player in RNA polymerase II transcription? Biochim. Biophys. Acta 2010, 1799, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Van Oss, S.B.; Cucinotta, C.E.; Arndt, K.M. Emerging Insights into the Roles of the Paf1 Complex in Gene Regulation. Trends Biochem. Sci. 2017, 42, 788–798. [Google Scholar] [CrossRef]
- Crisucci, E.M.; Arndt, K.M. The Roles of the Paf1 Complex and Associated Histone Modifications in Regulating Gene Expression. Genet. Res. Int. 2011, 2011, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Dover, J.; Schneider, J.; Tawiah-Boateng, M.A.; Wood, A.; Dean, K.; Johnston, M.; Shilatifard, A. Methylation of Histone H3 by COMPASS Requires Ubiquitination of Histone H2B by Rad6. J. Biol. Chem. 2002, 277, 28368–28371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.L.; Kaplan, T.; Kim, M.; Buratowski, S.; Schreiber, S.L.; Friedman, N.; Rando, O.J. Single-Nucleosome Mapping of Histone Modifications in S. cerevisiae. PLoS Biol. 2005, 3, e328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halbach, A.; Zhang, H.; Wengi, A.; Jablonska, Z.; Gruber, I.M.L.; Halbeisen, R.E.; Dehe, P.-M.; Kemmeren, P.; Holstege, F.; Géli, V.; et al. Cotranslational assembly of the yeast SET1C histone methyltransferase complex. EMBO J. 2009, 28, 2959–2970. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.L.; Shi, H.; Leonen, C.; Kang, J.; Chatterjee, C.; Zheng, N. Structural Basis of H2B Ubiquitination-Dependent H3K4 Methylation by COMPASS. Mol. Cell 2019, 76, 712–723.e4. [Google Scholar] [CrossRef]
- Winget, J.M.; Mayor, T. The Diversity of Ubiquitin Recognition: Hot Spots and Varied Specificity. Mol. Cell 2010, 38, 627–635. [Google Scholar] [CrossRef]
- Worden, E.J.; Zhang, X.; Wolberger, C. Structural basis for COMPASS recognition of an H2B-Ubiquitinated nucleosome. ELife 2020, 9. [Google Scholar] [CrossRef]
- McGinty, R.K.; Tan, S. Recognition of the nucleosome by chromatin factors and enzymes. Curr. Opin. Struct. Biol. 2016, 37, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Ford, D.J.; Dingwall, A.K. The cancer COMPASS: Navigating the functions of MLL complexes in cancer. Cancer Genet. 2015, 208, 178–191. [Google Scholar] [CrossRef]
- Meeks, J.J.; Shilatifard, A. Multiple Roles for the MLL/COMPASS Family in the Epigenetic Regulation of Gene Expression and in Cancer. Annu. Rev. Cancer Biol. 2017, 1, 425–446. [Google Scholar] [CrossRef]
- Michalak, E.; Burr, M.L.; Bannister, A.J.; Dawson, M.A. The roles of DNA, RNA and histone methylation in ageing and cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 573–589. [Google Scholar] [CrossRef] [PubMed]
- Jeusset, L.M.-P.; McManus, K.J. Developing Targeted Therapies That Exploit Aberrant Histone Ubiquitination in Cancer. Cells 2019, 8, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Dent, S.Y. Functions of SAGA in development and disease. Epigenomics 2014, 6, 329–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suresh, B.; Lee, J.; Hong, S.-H.; Kim, K.-S.; Ramakrishna, S. The role of deubiquitinating enzymes in spermatogenesis. Cell. Mol. Life Sci. 2015, 72, 4711–4720. [Google Scholar] [CrossRef]
- Cuvertino, S.; Hartill, V.; Colyer, A.; Garner, T.; Nair, N.; Al-Gazali, L.; Canham, N.; Faundes, V.; Flinter, F.; Genomics England Research Consortium; et al. A restricted spectrum of missense KMT2D variants cause a multiple malformations disorder distinct from Kabuki syndrome. Genet. Med. 2020, 22, 867–877. [Google Scholar] [CrossRef] [Green Version]
- Antunes, E.T.; Ottersbach, K. The MLL/SET family and haematopoiesis. Biochim. Biophys. Acta 2020, 1863, 194579. [Google Scholar] [CrossRef]
- Cardoso, L.; Stevenson, M.; Thakker, R.V. Molecular genetics of syndromic and non-syndromic forms of parathyroid carcinoma. Hum. Mutat. 2017, 38, 1621–1648. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Paszkowski-Rogacz, M.; Nitzsche, A.; Slabicki, M.M.; Heninger, A.-K.; De Vries, I.; Kittler, R.; Junqueira, M.; Shevchenko, A.; Schulz, H.; et al. A Genome-Scale RNAi Screen for Oct4 Modulators Defines a Role of the Paf1 Complex for Embryonic Stem Cell Identity. Cell Stem Cell 2009, 4, 403–415. [Google Scholar] [CrossRef] [Green Version]
- Shu, W.-J.; Zhao, M.-J.; Klionsky, D.J.; Du, H.-N. Old factors, new players: Transcriptional regulation of autophagy. Autophagy 2020, 16, 1–3. [Google Scholar] [CrossRef]
- Paigen, K.; Petkov, P.M. PRDM9 and Its Role in Genetic Recombination. Trends Genet. 2018, 34, 291–300. [Google Scholar] [CrossRef]
- Bhattacharyya, T.; Walker, M.; Powers, N.R.; Brunton, C.; Fine, A.D.; Petkov, P.M.; Handel, M.A. Prdm9 and Meiotic Cohesin Proteins Cooperatively Promote DNA Double-Strand Break Formation in Mammalian Spermatocytes. Curr. Biol. 2019, 29, 1002–1018.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somasagara, R.R.; Spencer, S.M.; Tripathi, K.; Clark, D.W.; Mani, C.; Da Silva, L.M.; Scalici, J.; Kothayer, H.; Westwell, A.D.; Rocconi, R.P.; et al. RAD6 promotes DNA repair and stem cell signaling in ovarian cancer and is a promising therapeutic target to prevent and treat acquired chemoresistance. Oncogene 2017, 36, 6680–6690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Kim, D.-H.; Lee, S.; Yang, Q.-H.; Lee, D.K.; Lee, S.-K.; Roeder, R.G.; Lee, J.W. A tumor suppressive coactivator complex of p53 containing ASC-2 and histone H3-lysine-4 methyltransferase MLL3 or its paralogue MLL4. Proc. Natl. Acad. Sci. USA 2009, 106, 8513–8518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, H.; Kurata, T.; Takiguchi, Y.; Arai, M.; Takeda, K.; Akiyoshi, K.; Matsumoto, K.; Onoe, T.; Mukai, H.; Matsubara, N.; et al. Randomized Phase II Trial Comparing Site-Specific Treatment Based on Gene Expression Profiling With Carboplatin and Paclitaxel for Patients With Cancer of Unknown Primary Site. J. Clin. Oncol. 2019, 37, 570–579. [Google Scholar] [CrossRef]
- Butler, J.S.; Qiu, Y.H.; Zhang, N.; Yoo, S.-Y.; Coombes, K.R.; Dent, S.Y.; Kornblau, S.M. Low expression of ASH2L protein correlates with a favorable outcome in acute myeloid leukemia. Leuk. Lymphoma 2016, 58, 1207–1218. [Google Scholar] [CrossRef]
- Wu, Y.-J.; Li, L.-X.; Liu, L.; Zhao, S.-S.; Qiu, H.-R.; Wang, H. ASH2L-Promoted HOXC8 Gene Expression Plays a Role in Mixed Lineage Leukemia-Rearranged Acute Leukemia. Onco Targets Ther. 2020, 13, 381–387. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Wang, L.; Volk, A.; Birch, N.W.; Stoltz, K.L.; Bartom, E.T.; Marshall, S.A.; Rendleman, E.J.; Nestler, C.M.; Shilati, J.; et al. Regulation of MLL/COMPASS stability through its proteolytic cleavage by taspase1 as a possible approach for clinical therapy of leukemia. Genes Dev. 2018, 33, 61–74. [Google Scholar] [CrossRef]
- Davalli, P.; Mitic, T.; Caporali, A.; Lauriola, A.; D’Arca, D. ROS, Cell Senescence, and Novel Molecular Mechanisms in Aging and Age-Related Diseases. Oxid. Med. Cell. Longev. 2016, 2016, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Bazopoulou, D.; Knoefler, D.; Zheng, Y.; Ulrich, K.; Oleson, B.J.; Xie, L.; Kim, M.; Kaufmann, A.; Lee, Y.-T.; Dou, Y.; et al. Developmental ROS individualizes organismal stress resistance and lifespan. Nature 2019, 576, 301–305. [Google Scholar] [CrossRef]
- Yang, Y.; Joshi, M.; Takahashi, Y.-H.; Ning, Z.; Qu, Q.; Brunzelle, J.S.; Skiniotis, G.; Figeys, D.; Shilatifard, A.; Couture, J.-F. A non-canonical monovalent zinc finger stabilizes the integration of Cfp1 into the H3K4 methyltransferase complex COMPASS. Nucleic Acids Res. 2020, 48, 421–431. [Google Scholar] [CrossRef]
- Sha, Q.-Q.; Jiang, Y.; Yu, C.; Xiang, Y.; Dai, X.-X.; Jiang, J.-C.; Ou, X.-H.; Fan, H.-Y. CFP1-dependent histone H3K4 trimethylation in murine oocytes facilitates ovarian follicle recruitment and ovulation in a cell-nonautonomous manner. Cell. Mol. Life Sci. 2019, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Billings, T.; Petkov, P.M. CXXC1 is not essential for normal DNA double-strand break formation and meiotic recombination in mouse. PLoS Genet. 2018, 14, e1007657. [Google Scholar] [CrossRef] [PubMed]
- Albert, T.K.; Antrecht, C.; Kremmer, E.; Meisterernst, M. The Establishment of a Hyperactive Structure Allows the Tumour Suppressor Protein p53 to Function through P-TEFb during Limited CDK9 Kinase Inhibition. PLoS ONE 2016, 11, e0146648. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.F.; Tan, M.-H.; Zhang, C.; Morreau, H.; Teh, B.T. HRPT2, a Tumor Suppressor Gene for Hyperparathyroidism-Jaw Tumor Syndrome. Horm. Metab. Res. 2005, 37, 380–383. [Google Scholar] [CrossRef]
- Hanks, S.; Perdeaux, E.; Seal, S.; Ruark, E.; Mahamdallie, S.S.; Murray, A.; Ramsay, E.; Duarte, S.D.V.; Zachariou, A.; De Souza, B.; et al. Germline mutations in the PAF1 complex gene CTR9 predispose to Wilms tumour. Nat. Commun. 2014, 5, 4398. [Google Scholar] [CrossRef] [Green Version]
- Karmakar, S.; Dey, P.; Vaz, A.P.; Bhaumik, S.R.; Ponnusamy, M.P.; Batra, S.K. PD2/PAF1 at the Crossroads of the Cancer Network. Cancer Res. 2018, 78, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Soloveychik, M.; Ranger, M.; Schertzberg, M.; Shah, Z.; Raisner, R.; Venkatasubrahmanyan, S.; Tsui, K.; Gebbia, M.; Hughes, T.; et al. Timing of transcriptional quiescence during gametogenesis is controlled by global histone H3K4 demethylation. Dev. Cell 2012, 23, 1059–1071. [Google Scholar] [CrossRef] [Green Version]
- Verzijlbergen, K.F.; Marston, A.L. A JARID Family Demethylase Controls Differentiation Timing through Global Effects on Transcription. Mol. Cell 2012, 48, 489–490. [Google Scholar] [CrossRef] [Green Version]
- Henry, K.W.; Wyce, A.; Lo, W.-S.; Duggan, L.J.; Emre, N.T.; Kao, C.-F.; Pillus, L.; Shilatifard, A.; Osley, M.A.; Berger, S.L. Transcriptional activation via sequential histone H2B ubiquitylation and deubiquitylation, mediated by SAGA-associated Ubp8. Genome Res. 2003, 17, 2648–2663. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.K.; Florens, L.; Swanson, S.K.; Washburn, M.P.; Workman, J.L. The Deubiquitylation Activity of Ubp8 Is Dependent upon Sgf11 and Its Association with the SAGA Complex. Mol. Cell. Biol. 2005, 25, 1173–1182. [Google Scholar] [CrossRef] [Green Version]
- Ingvarsdottir, K.; Krogan, N.J.; Emre, N.C.T.; Wyce, A.; Thompson, N.J.; Emili, A.; Hughes, T.R.; Greenblatt, J.F.; Berger, S.L. H2B Ubiquitin Protease Ubp8 and Sgf11 Constitute a Discrete Functional Module within the Saccharomyces cerevisiae SAGA Complex. Mol. Cell. Biol. 2005, 25, 1162–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmlinger, D.; Marguerat, S.; Villén, J.; Gygi, S.P.; Bahler, J.; Winston, F. The S. pombe SAGA complex controls the switch from proliferation to sexual differentiation through the opposing roles of its subunits Gcn5 and Spt8. Genome Res. 2008, 22, 3184–3195. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complex. | Pathologies Associated | Reference |
|---|---|---|
| COMPASS/Set1 | Lymphoblastic and acute myeloid leukemia | [130] |
| Kabuki syndrome 1 | [135] | |
| Hematopoiesis | [136] | |
| Embryonic stem cell (ESC), induced pluripotent stem cell (iPSC) and neuronal stem cell proliferation | [130] | |
| Embryogenesis | [130] | |
| PAF1c | Parathyroid carcinoma (PC) | [137] |
| ESC pluripotency maintenance | [138] | |
| Mitochondrial autophagy | [139] | |
| PRDM9 | Defective synapsis and male infertility | [140,141] |
| Rad6 | Male infertility | [97,134] |
| Ovarian cancer | [142] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serrano-Quílez, J.; Roig-Soucase, S.; Rodríguez-Navarro, S. Sharing Marks: H3K4 Methylation and H2B Ubiquitination as Features of Meiotic Recombination and Transcription. Int. J. Mol. Sci. 2020, 21, 4510. https://doi.org/10.3390/ijms21124510
Serrano-Quílez J, Roig-Soucase S, Rodríguez-Navarro S. Sharing Marks: H3K4 Methylation and H2B Ubiquitination as Features of Meiotic Recombination and Transcription. International Journal of Molecular Sciences. 2020; 21(12):4510. https://doi.org/10.3390/ijms21124510
Chicago/Turabian StyleSerrano-Quílez, Joan, Sergi Roig-Soucase, and Susana Rodríguez-Navarro. 2020. "Sharing Marks: H3K4 Methylation and H2B Ubiquitination as Features of Meiotic Recombination and Transcription" International Journal of Molecular Sciences 21, no. 12: 4510. https://doi.org/10.3390/ijms21124510
APA StyleSerrano-Quílez, J., Roig-Soucase, S., & Rodríguez-Navarro, S. (2020). Sharing Marks: H3K4 Methylation and H2B Ubiquitination as Features of Meiotic Recombination and Transcription. International Journal of Molecular Sciences, 21(12), 4510. https://doi.org/10.3390/ijms21124510