Understanding the Biology of Human Interstitial Cells of Cajal in Gastrointestinal Motility




Abstract
:1. Introduction
2. Anatomical Locations of ICC
ICC Subtypes
- Myenteric ICC (ICC-MY) are located surrounding the circumference of the myenteric plexus. They are multipolar cells with branched processes that form a network around the myenteric plexus. ICC-MY have been suggested to be the primary pacemaker cells of the stomach and small intestine muscles, involved in generating and propagating slow-wave activity through smooth muscle cells [4].
- Intramuscular ICC (ICC-IM) are found within the circular and longitudinal muscle. They are bipolar or spindle-shaped cells that align with the long axis of surrounding smooth muscle cells. ICC-IM play a key role in mediating enteric neurotransmission [36].
- ICC of the deep myenteric plexus (ICC-DMP) are found exclusively in the small intestine. They are multipolar cells that are closely associated with nerve bundles of the DMP. Similar to ICC-IM, ICC-DMP also mediate enteric neurotransmission [36]. Neurokinin-1 receptor (NK1R) has been used as a marker to identify and isolate murine Kit+ ICC-DMP [28].
- Other ICC subtypes have been described, including submucosal ICC (ICC-SM; located at the interface between the submucosa and circular muscle of the stomach), subserosal ICC (ICC-SS; found between the serosa and longitudinal muscle of small intestine and colon) and septal ICC (ICC-SEP; located between and surrounding muscle bundles, particularly in larger animals like humans). ICC-SEP have also been implicated in propagating pacemaker activity into muscle bundles of the human jejunum [45]. More recently, a small stem cell-like population of ‘ICC progenitors’ were defined as ‘KitlowCD44+CD34+Insr+Igf1r+’ within the mouse stomach [38].
3. Potential Sources of ICC
3.1. Murine GI Tissue
3.2. Human GI Tissue
3.3. Bioengineered GI Tissue
4. ICC and Molecular Characteristics
4.1. Murine ICC
4.2. Human ICC
5. ICC and GI Disorders
5.1. ICC and Ageing
5.2. ICC and Diabetic Gastroparesis
5.3. ICC and Other GI Motility Disorders
5.4. ICC and Gastrointestinal Stromal Tumours (GISTs)
6. ICC and GI Therapy
6.1. Cell-based ICC Therapy
6.2. Scaffold-Based ICC Therapy
7. Establishing a Source of Candidate Human ICC for Molecular Characterisation
8. Conclusions and Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GI | Gastrointestinal |
| ICC | Interstitial cells of Cajal |
| ER | Endoplasmic reticulum |
| PDGRFα+ | Platelet-derived growth factor receptor alpha-positive |
| ICC-MY | Myenteric ICC |
| ICC-IM | Intramuscular ICC |
| ICC-DMP | ICC of the deep myenteric plexus |
| ICC-SM | Submucosal ICC |
| ICC-SS | Subserosal ICC |
| ICC-SEP | Septal ICC |
| MACS | Magnetic-activated cell sorting |
| FACS | Fluorescence-activated cell sorting |
| RT-PCR | Real-time polymerase chain reaction |
| RNA-seq | RNA-sequencing |
| HSCR | Hirschsprung’s disease |
| GIST | Gastrointestinal stromal tumour |
References
- Lacy, B.E.; Weiser, K. Gastrointestinal Motility Disorders: An Update. Dig. Dis. 2006, 24, 228–242. [Google Scholar] [CrossRef] [PubMed]
- Huizinga, J.D.; Lammers, W.J. Gut peristalsis is governed by a multitude of cooperating mechanisms. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1–G8. [Google Scholar] [CrossRef] [PubMed]
- Huizinga, J.D. Gastrointestinal peristalsis: Joint action of enteric nerves, smooth muscle, and interstitial cells of Cajal. Microsc. Res. Tech. 1999, 47, 239–247. [Google Scholar] [CrossRef]
- Blair, P.J.; Rhee, P.L.; Sanders, K.M.; Ward, S.M. The significance of interstitial cells in neurogastroenterology. J. Neurogastroenterol. Motil. 2014, 20, 294–317. [Google Scholar] [CrossRef]
- Sanders, K.M.; Ward, S.M.; Koh, S.D. Interstitial cells: Regulators of smooth muscle function. Physiol. Rev. 2014, 94, 859–907. [Google Scholar] [CrossRef]
- Farrugia, G. Interstitial cells of Cajal in health and disease. Neurogastroenterol. Motil. 2008, 20 (Suppl. S1), 54–63. [Google Scholar] [CrossRef]
- Streutker, C.J.; Huizinga, J.D.; Driman, D.K.; Riddell, R.H. Interstitial cells of Cajal in health and disease. Part I: Normal ICC structure and function with associated motility disorders. Histopathology 2007, 50, 176–189. [Google Scholar] [CrossRef]
- Cajal, S.R. Histologie du syste me nerveux de I’Homme et des verte be s. Maloine (Paris) 1911, 2, 891–942. [Google Scholar]
- Komuro, T. Structure and organization of interstitial cells of Cajal in the gastrointestinal tract. J. Physiol. 2006, 576, 653–658. [Google Scholar] [CrossRef]
- Faussone Pellegrini, M.S.; Cortesini, C.; Romagnoli, P. Ultrastructure of the tunica muscularis of the cardial portion of the human esophagus and stomach, with special reference to the so-called Cajal’s interstitial cells. Arch. Ital. Anat. Embriol. 1977, 82, 157–177. [Google Scholar]
- Torihashi, S.; Kobayashi, S.; Gerthoffer, W.T.; Sanders, K.M. Interstitial cells in deep muscular plexus of canine small intestine may be specialized smooth muscle cells. Am. J. Physiol. 1993, 265, G638–G645. [Google Scholar] [CrossRef] [PubMed]
- Rumessen, J.J.; Mikkelsen, H.B.; Thuneberg, L. Ultrastructure of interstitial cells of Cajal associated with deep muscular plexus of human small intestine. Gastroenterology 1992, 102, 56–68. [Google Scholar] [CrossRef]
- Rumessen, J.J.; Thuneberg, L. Interstitial Cells of Cajal in Human Small Intestine: Ultrastructural Identification and Organization Between the Main Smooth Muscle Layers. Gastroenterology 1991, 100, 1417–1431. [Google Scholar] [CrossRef]
- Ward, S.M.; Ordog, T.; Koh, S.D.; Baker, S.A.; Jun, J.Y.; Amberg, G.; Monaghan, K.; Sanders, K.M. Pacemaking in interstitial cells of Cajal depends upon calcium handling by endoplasmic reticulum and mitochondria. J. Physiol. 2000, 525 Pt 2, 355–361. [Google Scholar] [CrossRef]
- Beckett, E.A.; Takeda, Y.; Yanase, H.; Sanders, K.M.; Ward, S.M. Synaptic specializations exist between enteric motor nerves and interstitial cells of Cajal in the murine stomach. J. Comp. Neurol. 2005, 493, 193–206. [Google Scholar] [CrossRef]
- Ward, S.M.; Beckett, E.A.; Wang, X.; Baker, F.; Khoyi, M.; Sanders, K.M. Interstitial cells of Cajal mediate cholinergic neurotransmission from enteric motor neurons. J. Neurosci. 2000, 20, 1393–1403. [Google Scholar] [CrossRef]
- Yamamoto, M. Electron microscopic studies on the innervation of the smooth muscle and the interstitial cell of Cajal in the small intestine of the mouse and bat. Arch. Histol. Jpn. 1977, 40, 171–201. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Y.; Paterson, C.; Huizinga, J.D. Cholinergic and nitrergic innervation of ICC-DMP and ICC-IM in the human small intestine. Neurogastroenterol. Motil. 2003, 15, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Ibba Manneschi, L.; Pacini, S.; Corsani, L.; Bechi, P.; Faussone-Pellegrini, M.S. Interstitital cells of Cajal in the human stomach: Distribution and relationship with enteric innervation. Histol. Histopathol. 2004, 19, 1153–1164. [Google Scholar] [CrossRef]
- Burns, A.J.; Lomax, A.E.; Torihashi, S.; Sanders, K.M.; Ward, S.M. Interstitial cells of Cajal mediate inhibitory neurotransmission in the stomach. Proc. Natl. Acad. Sci. USA 1996, 93, 12008–12013. [Google Scholar] [CrossRef] [Green Version]
- Maria Giuliana, V.; Chiara, T. Interstitial cells of Cajal and telocytes in the gut: Twins, related or simply neighbor cells? Biomol. Concepts 2016, 7, 93–102. [Google Scholar] [CrossRef]
- Popescu, L.M.; Faussone-Pellegrini, M.-S. TELOCYTES—A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to TELOCYTES. J. Cell. Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulzinga, J.D.; Thuneberg, L.; Klüppel, M.; Malysz, J.; Mikkelsen, H.B.; Bernstein, A. W/kit gene required for interstitial cells of Cajal and for intestinal pacemaker activity. Nature 1995, 373, 347–349. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ordog, T.; Chen, J.; Young, D.L.; Bardsley, M.R.; Redelman, D.; Ward, S.M.; Sanders, K.M. Differential gene expression in functional classes of interstitial cells of Cajal in murine small intestine. Physiol. Genom. 2007, 31, 492–509. [Google Scholar] [CrossRef] [Green Version]
- Ordog, T.; Redelman, D.; Horowitz, N.N.; Sanders, K.M. Immunomagnetic enrichment of interstitial cells of Cajal. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G351–G360. [Google Scholar] [CrossRef] [Green Version]
- Epperson, A.; Hatton, W.J.; Callaghan, B.; Doherty, P.; Walker, R.L.; Sanders, K.M.; Ward, S.M.; Horowitz, B. Molecular markers expressed in cultured and freshly isolated interstitial cells of Cajal. Am. J. Physiol. Cell Physiol. 2000, 279, C529–C539. [Google Scholar] [CrossRef]
- Ordog, T.; Redelman, D.; Miller, L.J.; Horvath, V.J.; Zhong, Q.; Almeida-Porada, G.; Zanjani, E.D.; Horowitz, B.; Sanders, K.M. Purification of interstitial cells of Cajal by fluorescence-activated cell sorting. Am. J. Physiol. Cell Physiol. 2004, 286, C448–C456. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Redelman, D.; Ro, S.; Ward, S.M.; Ordog, T.; Sanders, K.M. Selective labeling and isolation of functional classes of interstitial cells of Cajal of human and murine small intestine. Am. J. Physiol. Cell Physiol. 2007, 292, C497–C507. [Google Scholar] [CrossRef]
- Torihashi, S.; Nishi, K.; Tokutomi, Y.; Nishi, T.; Ward, S.; Sanders, K.M. Blockade of kit signaling induces transdifferentiation of interstitial cells of cajal to a smooth muscle phenotype. Gastroenterology 1999, 117, 140–148. [Google Scholar] [CrossRef]
- Torihashi, S.; Ward, S.M.; Nishikawa, S.; Nishi, K.; Kobayashi, S.; Sanders, K.M. c-kit-dependent development of interstitial cells and electrical activity in the murine gastrointestinal tract. Cell Tissue Res. 1995, 280, 97–111. [Google Scholar]
- Ward, S.M.; Harney, S.C.; Bayguinov, J.R.; McLaren, G.J.; Sanders, K.M. Development of electrical rhythmicity in the murine gastrointestinal tract is specifically encoded in the tunica muscularis. J. Physiol. 1997, 505 Pt 1, 241–258. [Google Scholar] [CrossRef]
- Gomez-Pinilla, P.J.; Gibbons, S.J.; Bardsley, M.R.; Lorincz, A.; Pozo, M.J.; Pasricha, P.J.; Van de Rijn, M.; West, R.B.; Sarr, M.G.; Kendrick, M.L.; et al. Ano1 is a selective marker of interstitial cells of Cajal in the human and mouse gastrointestinal tract. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1370–G1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.J.; Blair, P.J.; Britton, F.C.; O’Driscoll, K.E.; Hennig, G.; Bayguinov, Y.R.; Rock, J.R.; Harfe, B.D.; Sanders, K.M.; Ward, S.M. Expression of anoctamin 1/TMEM16A by interstitial cells of Cajal is fundamental for slow wave activity in gastrointestinal muscles. J. Physiol. 2009, 587, 4887–4904. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.H.; Kim, T.W.; Ro, S.; Yan, W.; Ward, S.M.; Koh, S.D.; Sanders, K.M. A Ca2+-activated Cl– conductance in interstitial cells of Cajal linked to slow wave currents and pacemaker activity. J. Physiol. 2009, 587, 4905–4918. [Google Scholar] [CrossRef] [PubMed]
- Komuro, T. Comparative morphology of interstitial cells of Cajal: Ultrastructural characterization. Microsc. Res. Tech. 1999, 47, 267–285. [Google Scholar] [CrossRef]
- Iino, S.; Horiguchi, K. Interstitial cells of cajal are involved in neurotransmission in the gastrointestinal tract. Acta Histochem. Cytochem. 2006, 39, 145–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ördög, T.; Redelman, D.; Horváth, V.J.; Miller, L.J.; Horowitz, B.; Sanders, K.M. Quantitative analysis by flow cytometry of interstitial cells of Cajal, pacemakers, and mediators of neurotransmission in the gastrointestinal tract. Cytom. Part A 2004, 62, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Lőrincz, A.; Redelman, D.; Horváth, V.J.; Bardsley, M.R.; Chen, H.; Ördög, T. Progenitors of Interstitial Cells of Cajal in the Postnatal Murine Stomach. Gastroenterology 2008, 134, 1083–1093. [Google Scholar] [CrossRef] [Green Version]
- Ro, S.; Park, C.; Jin, J.; Zheng, H.; Blair, P.J.; Redelman, D.; Ward, S.M.; Yan, W.; Sanders, K.M. A model to study the phenotypic changes of interstitial cells of Cajal in gastrointestinal diseases. Gastroenterology 2010, 138, 1068–1078. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.Y.; Ha, S.E.; Park, C.; Park, P.J.; Fuchs, R.; Wei, L.; Jorgensen, B.G.; Redelman, D.; Ward, S.M.; Sanders, K.M.; et al. Transcriptome of interstitial cells of Cajal reveals unique and selective gene signatures. PLoS ONE 2017, 12, e0176031. [Google Scholar] [CrossRef] [Green Version]
- Koh, S.D.; Sanders, K.M.; Ward, S.M. Spontaneous electrical rhythmicity in cultured interstitial cells of cajal from the murine small intestine. J. Physiol. 1998, 513 Pt 1, 203–213. [Google Scholar] [CrossRef]
- Thomsen, L.; Robinson, T.L.; Lee, J.C.; Farraway, L.A.; Hughes, M.J.; Andrews, D.W.; Huizinga, J.D. Interstitial cells of Cajal generate a rhythmic pacemaker current. Nat. Med. 1998, 4, 848–851. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Matsuoka, S.; Noma, A. Two types of spontaneous depolarizations in the interstitial cells freshly prepared from the murine small intestine. J. Physiol. 2004, 559, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.H.; Sung, T.S.; O’Driscoll, K.; Koh, S.D.; Sanders, K.M. Intracellular Ca2+ release from endoplasmic reticulum regulates slow wave currents and pacemaker activity of interstitial cells of Cajal. Am. J. Physiol. Cell Physiol. 2015, 308, C608–C620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.T.; Hennig, G.W.; Fleming, N.W.; Keef, K.D.; Spencer, N.J.; Ward, S.M.; Sanders, K.M.; Smith, T.K. Septal interstitial cells of Cajal conduct pacemaker activity to excite muscle bundles in human jejunum. Gastroenterology 2007, 133, 907–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Shamu, T.; Chen, H.; Besmer, P.; Sawyers, C.L.; Chi, P. Visualization of the interstitial cells of cajal (ICC) network in mice. J. Vis. Exp. 2011, e2802. [Google Scholar] [CrossRef]
- Ward, S.M.; Burns, A.J.; Torihashi, S.; Sanders, K.M. Mutation of the proto-oncogene c-kit blocks development of interstitial cells and electrical rhythmicity in murine intestine. J. Physiol. 1994, 480 Pt 1, 91–97. [Google Scholar] [CrossRef]
- Iino, S.; Horiguchi, S.; Horiguchi, K. Interstitial cells of Cajal in the gastrointestinal musculature of W(jic) c-kit mutant mice. J. Smooth Muscle Res. 2011, 47, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Malysz, J.; Gibbons, S.J.; Saravanaperumal, S.A.; Du, P.; Eisenman, S.T.; Cao, C.; Oh, U.; Saur, D.; Klein, S.; Ordog, T.; et al. Conditional genetic deletion of Ano1 in interstitial cells of Cajal impairs Ca2+ transients and slow waves in adult mouse small intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G228–G245. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.D.; Gibbons, S.J.; Saravanaperumal, S.A.; Du, P.; Hennig, G.W.; Eisenman, S.T.; Mazzone, A.; Hayashi, Y.; Cao, C.; Stoltz, G.J.; et al. Ano1, a Ca2+-activated Cl− channel, coordinates contractility in mouse intestine by Ca2+ transient coordination between interstitial cells of Cajal. J. Physiol. 2014, 592, 4051–4068. [Google Scholar] [CrossRef]
- Ordog, T.; Takayama, I.; Cheung, W.K.; Ward, S.M.; Sanders, K.M. Remodeling of networks of interstitial cells of Cajal in a murine model of diabetic gastroparesis. Diabetes 2000, 49, 1731–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagida, H.; Yanase, H.; Sanders, K.M.; Ward, S.M. Intestinal surgical resection disrupts electrical rhythmicity, neural responses, and interstitial cell networks. Gastroenterology 2004, 127, 1748–1759. [Google Scholar] [CrossRef] [PubMed]
- Chang, I.Y.; Glasgow, N.J.; Takayama, I.; Horiguchi, K.; Sanders, K.M.; Ward, S.M. Loss of interstitial cells of Cajal and development of electrical dysfunction in murine small bowel obstruction. J. Physiol. 2001, 536, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pinilla, P.J.; Gibbons, S.J.; Sarr, M.G.; Kendrick, M.L.; Shen, K.R.; Cima, R.R.; Dozois, E.J.; Larson, D.W.; Ordog, T.; Pozo, M.J.; et al. Changes in interstitial cells of cajal with age in the human stomach and colon. Neurogastroenterol. Motil. 2011, 23, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.M.; Lee, T.E.; Syed, S.A.; Burgoyne, A.M.; Leonard, S.Y.; Gao, F.; Chan, J.C.; Shi, E.; Chmielecki, J.; Morosini, D.; et al. Hedgehog pathway dysregulation contributes to the pathogenesis of human gastrointestinal stromal tumors via GLI-mediated activation of KIT expression. Oncotarget 2016, 7, 78226–78241. [Google Scholar] [CrossRef] [Green Version]
- Park, K.S.; Cho, K.B.; Hwang, I.S.; Park, J.H.; Jang, B.I.; Kim, K.O.; Jeon, S.W.; Kim, E.S.; Park, C.S.; Kwon, J.G. Characterization of smooth muscle, enteric nerve, interstitial cells of Cajal, and fibroblast-like cells in the gastric musculature of patients with diabetes mellitus. World J. Gastroenterol. 2016, 22, 10131–10139. [Google Scholar] [CrossRef]
- Yun, H.Y.; Sung, R.; Kim, Y.C.; Choi, W.; Kim, H.S.; Kim, H.; Lee, G.J.; You, R.Y.; Park, S.M.; Yun, S.J.; et al. Regional Distribution of Interstitial Cells of Cajal (ICC) in Human Stomach. Korean J. Physiol. Pharmacol. 2010, 14, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Rhee, P.-L.; Lee, J.Y.; Son, H.J.; Kim, J.J.; Rhee, J.C.; Kim, S.; Koh, S.D.; Hwang, S.J.; Sanders, K.M.; Ward, S.M. Analysis of pacemaker activity in the human stomach. J. Physiol. 2011, 589, 6105–6118. [Google Scholar] [CrossRef]
- Lee, H.T.; Hennig, G.W.; Fleming, N.W.; Keef, K.D.; Spencer, N.J.; Ward, S.M.; Sanders, K.M.; Smith, T.K. The Mechanism and Spread of Pacemaker Activity Through Myenteric Interstitial Cells of Cajal in Human Small Intestine. Gastroenterology 2007, 132, 1852–1865. [Google Scholar] [CrossRef]
- Strege, P.R.; Ou, Y.; Sha, L.; Rich, A.; Gibbons, S.J.; Szurszewski, J.H.; Sarr, M.G.; Farrugia, G. Sodium current in human intestinal interstitial cells of Cajal. Am. J. Physiol. 2003, 285, G1111–G1121. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.-H.; Zhang, Y.-C.; Jiang, W.-F.; Yang, C.; Zou, G.-M.; Kong, Y.; Cai, W. Characterization of Interstitial Cajal Progenitors Cells and Their Changes in Hirschsprung’s Disease. PLoS ONE 2014, 9, e86100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashyap, P.; Gomez-Pinilla, P.J.; Pozo, M.J.; Cima, R.R.; Dozois, E.J.; Larson, D.W.; Ordog, T.; Gibbons, S.J.; Farrugia, G. Immunoreactivity for Ano1 detects depletion of Kit-positive interstitial cells of Cajal in patients with slow transit constipation. Neurogastroenterol. Motil. 2011, 23, 760–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grover, M.; Farrugia, G.; Lurken, M.S.; Bernard, C.E.; Faussone-Pellegrini, M.S.; Smyrk, T.C.; Parkman, H.P.; Abell, T.L.; Snape, W.J.; Hasler, W.L.; et al. Cellular Changes in Diabetic and Idiopathic Gastroparesis. Gastroenterology 2011, 140, 1575–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herring, B.P.; Chen, M.; Mihaylov, P.; Hoggatt, A.M.; Gupta, A.; Nakeeb, A.; Choi, J.N.; Wo, J.M. Transcriptome profiling reveals significant changes in the gastric muscularis externa with obesity that partially overlap those that occur with idiopathic gastroparesis. BMC Med Genom. 2019, 12, 89. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wang, K.; Solorzano-Vargas, R.S.; Lin, P.Y.; Walthers, C.M.; Thomas, A.L.; Martín, M.G.; Dunn, J.C.Y. Bioengineered intestinal muscularis complexes with long-term spontaneous and periodic contractions. PLoS ONE 2018, 13, e0195315. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Khalil, H.A.; Lei, N.Y.; Wang, Q.; Wang, K.; Wu, B.M.; Dunn, J.C.Y. Bioengineering functional smooth muscle with spontaneous rhythmic contraction in vitro. Sci. Rep. 2018, 8, 13544. [Google Scholar] [CrossRef]
- Uchida, H.; Machida, M.; Miura, T.; Kawasaki, T.; Okazaki, T.; Sasaki, K.; Sakamoto, S.; Ohuchi, N.; Kasahara, M.; Umezawa, A.; et al. A xenogeneic-free system generating functional human gut organoids from pluripotent stem cells. JCI Insight 2017, 2, e86492. [Google Scholar] [CrossRef] [Green Version]
- Workman, M.J.; Mahe, M.M.; Trisno, S.; Poling, H.M.; Watson, C.L.; Sundaram, N.; Chang, C.-F.; Schiesser, J.; Aubert, P.; Stanley, E.G.; et al. Engineered human pluripotent-stem-cell-derived intestinal tissues with a functional enteric nervous system. Nat. Med. 2017, 23, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Meng, W.; Zhou, J.; Elliott, R.; Murphy, P.; Ho, V.; O’Connor, M. Is there a role for human pluripotent stem cells in modelling interstitial cells of Cajal and gut motility disorders? Curr. Stem Cell Res. Ther. 2015, 10, 251–257. [Google Scholar] [CrossRef]
- Zhou, J.; O’Connor, M.D.; Ho, V. The Potential for Gut Organoid Derived Interstitial Cells of Cajal in Replacement Therapy. Int. J. Mol. Sci. 2017, 18, 2059. [Google Scholar] [CrossRef] [Green Version]
- Breland, A.; Ha, S.E.; Jorgensen, B.G.; Jin, B.; Gardner, T.A.; Sanders, K.M.; Ro, S. Smooth Muscle Transcriptome Browser: Offering genome-wide references and expression profiles of transcripts expressed in intestinal SMC, ICC, and PDGFRα+ cells. Sci. Rep. 2019, 9, 387. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.M.; Ward, S.M. Nitric oxide and its role as a non-adrenergic, non-cholinergic inhibitory neurotransmitter in the gastrointestinal tract. Br. J. Pharm. 2019, 176, 212–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faussone-Pellegrini, M.S.; Grover, M.; Pasricha, P.J.; Bernard, C.E.; Lurken, M.S.; Smyrk, T.C.; Parkman, H.P.; Abell, T.L.; Snape, W.J.; Hasler, W.L.; et al. Ultrastructural differences between diabetic and idiopathic gastroparesis. J. Cell. Mol. Med. 2012, 16, 1573–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderwinden, J.M.; Rumessen, J.J.; Liu, H.; Descamps, D.; De Laet, M.H.; Vanderhaeghen, J.J. Interstitial cells of Cajal in human colon and in Hirschsprung’s disease. Gastroenterology 1996, 111, 901–910. [Google Scholar] [CrossRef]
- Rolle, U.; Piotrowska, A.P.; Nemeth, L.; Puri, P. Altered Distribution of Interstitial Cells of Cajal in Hirschsprung Disease. Arch. Pathol. Lab. Med. 2002, 126, 928–933. [Google Scholar] [CrossRef]
- Lyford, G.L.; He, C.L.; Soffer, E.; Hull, T.L.; Strong, S.A.; Senagore, A.J.; Burgart, L.J.; Young-Fadok, T.; Szurszewski, J.H.; Farrugia, G. Pan-colonic decrease in interstitial cells of Cajal in patients with slow transit constipation. Gut 2002, 51, 496. [Google Scholar] [CrossRef]
- Wedel, T.; Spiegler, J.; Soellner, S.; Roblick, U.J.; Schiedeck, T.H.K.; Bruch, H.-P.; Krammer, H.-J. Enteric nerves and interstitial cells of Cajal are altered in patients with slow-transit constipation and megacolon. Gastroenterology 2002, 123, 1459–1467. [Google Scholar] [CrossRef]
- Bhutto, A.; Morley, J.E. The clinical significance of gastrointestinal changes with aging. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 651–660. [Google Scholar] [CrossRef]
- Salles, N. Basic mechanisms of the aging gastrointestinal tract. Dig. Dis. 2007, 25, 112–117. [Google Scholar] [CrossRef]
- Soenen, S.; Rayner, C.K.; Jones, K.L.; Horowitz, M. The ageing gastrointestinal tract. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 12–18. [Google Scholar] [CrossRef]
- Sun, T.; Li, D.; Hu, S.; Huang, L.; Sun, H.; Yang, S.; Wu, B.; Ji, F.; Zhou, D. Aging-dependent decrease in the numbers of enteric neurons, interstitial cells of Cajal and expression of connexin43 in various regions of gastrointestinal tract. Aging 2018, 10, 3851–3865. [Google Scholar] [CrossRef] [PubMed]
- Gamage, P.P.K.M.; Patel, B.A.; Yeoman, M.S.; Ranson, R.N.; Saffrey, M.J. Interstitial cell network volume is reduced in the terminal bowel of ageing mice. J. Cell. Mol. Med. 2018, 22, 5160–5164. [Google Scholar] [CrossRef] [PubMed]
- Horvath, V.J.; Izbeki, F.; Lengyel, C.; Kempler, P.; Varkonyi, T. Diabetic gastroparesis: Functional/morphologic background, diagnosis, and treatment options. Curr. Diabetes Rep. 2014, 14, 527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharucha, A.E.; Camilleri, M.; Forstrom, L.A.; Zinsmeister, A.R. Relationship between clinical features and gastric emptying disturbances in diabetes mellitus. Clin. Endocrinol. 2009, 70, 415–420. [Google Scholar] [CrossRef]
- Bharucha, A.E.; Kudva, Y.; Basu, A.; Camilleri, M.; Low, P.A.; Vella, A.; Zinsmeister, A.R. Relationship between glycemic control and gastric emptying in poorly controlled type 2 diabetes. Clin. Gastroenterol. Hepatol. 2015, 13, 466–476. [Google Scholar] [CrossRef] [Green Version]
- Bashashati, M.; McCallum, R.W. Is Interstitial Cells of Cajal–opathy Present in Gastroparesis? J. Neurogastroenterol. Motil. 2015, 21, 486–493. [Google Scholar] [CrossRef]
- Bernard, C.E.; Gibbons, S.J.; Mann, I.S.; Froschauer, L.; Parkman, H.P.; Harbison, S.; Abell, T.L.; Snape, W.J.; Hasler, W.L.; McCallum, R.W.; et al. Association of low numbers of CD206-positive cells with loss of ICC in the gastric body of patients with diabetic gastroparesis. Neurogastroenterol. Motil. 2014, 26, 1275–1284. [Google Scholar] [CrossRef] [Green Version]
- Grover, M.; Gibbons, S.J.; Nair, A.A.; Bernard, C.E.; Zubair, A.S.; Eisenman, S.T.; Wilson, L.A.; Miriel, L.; Pasricha, P.J.; Parkman, H.P.; et al. Transcriptomic signatures reveal immune dysregulation in human diabetic and idiopathic gastroparesis. BMC Med. Genom. 2018, 11, 62. [Google Scholar] [CrossRef]
- Grover, M.; Dasari, S.; Bernard, C.E.; Chikkamenahalli, L.L.; Yates, K.P.; Pasricha, P.J.; Sarosiek, I.; McCallum, R.; Koch, K.L.; Abell, T.L.; et al. Proteomics in gastroparesis: Unique and overlapping protein signatures in diabetic and idiopathic gastroparesis. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G716–G726. [Google Scholar] [CrossRef]
- Yamamoto, T.; Watabe, K.; Nakahara, M.; Ogiyama, H.; Kiyohara, T.; Tsutsui, S.; Tamura, S.; Shinomura, Y.; Hayashi, N. Disturbed gastrointestinal motility and decreased interstitial cells of Cajal in diabetic db/db mice. J. Gastroenterol. Hepatol. 2008, 23, 660–667. [Google Scholar] [CrossRef]
- Choi, K.M.; Gibbons, S.J.; Nguyen, T.V.; Stoltz, G.J.; Lurken, M.S.; Ordog, T.; Szurszewski, J.H.; Farrugia, G. Heme oxygenase-1 protects interstitial cells of Cajal from oxidative stress and reverses diabetic gastroparesis. Gastroenterology 2008, 135, 2055–2064. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.M.; Gibbons, S.J.; Sha, L.; Beyder, A.; Verhulst, P.J.; Cipriani, G.; Phillips, J.E.; Bauer, A.J.; Ordog, T.; Camp, J.J.; et al. Interleukin 10 Restores Gastric Emptying, Electrical Activity, and Interstitial Cells of Cajal Networks in Diabetic Mice. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 454–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, V.J.; Vittal, H.; Lorincz, A.; Chen, H.; Almeida-Porada, G.; Redelman, D.; Ordog, T. Reduced stem cell factor links smooth myopathy and loss of interstitial cells of cajal in murine diabetic gastroparesis. Gastroenterology 2006, 130, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Toyomasu, Y.; Saravanaperumal, S.A.; Bardsley, M.R.; Smestad, J.A.; Lorincz, A.; Eisenman, S.T.; Cipriani, G.; Nelson Holte, M.H.; Al Khazal, F.J.; et al. Hyperglycemia Increases Interstitial Cells of Cajal via MAPK1 and MAPK3 Signaling to ETV1 and KIT, Leading to Rapid Gastric Emptying. Gastroenterology 2017, 153, 521–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coyle, D.; Kelly, D.A.; O’Donnell, A.M.; Gillick, J.; Puri, P. Use of anoctamin 1 (ANO1) to evaluate interstitial cells of Cajal in Hirschsprung’s disease. Pediatr. Surg. Int. 2016, 32, 125–133. [Google Scholar] [CrossRef]
- Corless, C.L.; Barnett, C.M.; Heinrich, M.C. Gastrointestinal stromal tumours: Origin and molecular oncology. Nat. Rev. Cancer 2011, 11, 865–878. [Google Scholar] [CrossRef]
- Robinson, T.L.; Sircar, K.; Hewlett, B.R.; Chorneyko, K.; Riddell, R.H.; Huizinga, J.D. Gastrointestinal stromal tumors may originate from a subset of CD34-positive interstitial cells of Cajal. Am. J. Pathol. 2000, 156, 1157–1163. [Google Scholar] [CrossRef] [Green Version]
- Sarlomo-Rikala, M.; Kovatich, A.J.; Barusevicius, A.; Miettinen, M. CD117: A sensitive marker for gastrointestinal stromal tumors that is more specific than CD34. Mod. Pathol. 1998, 11, 728–734. [Google Scholar]
- Vanderwinden, J.M.; Rumessen, J.J.; De Laet, M.H.; Vanderhaeghen, J.J.; Schiffmann, S.N. CD34+ cells in human intestine are fibroblasts adjacent to, but distinct from, interstitial cells of Cajal. Lab. Investig. J. Tech. Methods Pathol. 1999, 79, 59–65. [Google Scholar]
- Vanderwinden, J.M.; Rumessen, J.J.; De Laet, M.H.; Vanderhaeghen, J.J.; Schiffmann, S.N. CD34 immunoreactivity and interstitial cells of Cajal in the human and mouse gastrointestinal tract. Cell Tissue Res. 2000, 302, 145–153. [Google Scholar] [CrossRef]
- Nakai, N.; Ishikawa, T.; Nishitani, A.; Liu, N.N.; Shincho, M.; Hao, H.; Isozaki, K.; Kanda, T.; Nishida, T.; Fujimoto, J.; et al. A mouse model of a human multiple GIST family with KIT-Asp820Tyr mutation generated by a knock-in strategy. J. Pathol. 2008, 214, 302–311. [Google Scholar] [CrossRef]
- Rubin, B.P.; Antonescu, C.R.; Scott-Browne, J.P.; Comstock, M.L.; Gu, Y.; Tanas, M.R.; Ware, C.B.; Woodell, J. A knock-in mouse model of gastrointestinal stromal tumor harboring kit K641E. Cancer Res. 2005, 65, 6631–6639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardsley, M.R.; Horvath, V.J.; Asuzu, D.T.; Lorincz, A.; Redelman, D.; Hayashi, Y.; Popko, L.N.; Young, D.L.; Lomberk, G.A.; Urrutia, R.A.; et al. Kitlow stem cells cause resistance to Kit/platelet-derived growth factor alpha inhibitors in murine gastrointestinal stromal tumors. Gastroenterology 2010, 139, 942–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCann, C.J.; Hwang, S.J.; Bayguinov, Y.; Colletti, E.J.; Sanders, K.M.; Ward, S.M. Establishment of pacemaker activity in tissues allotransplanted with interstitial cells of Cajal. Neurogastroenterol. Motil. 2013, 25, e418–e428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Jahagirdar, B.N.; Reinhardt, R.L.; Schwartz, R.E.; Keene, C.D.; Ortiz-Gonzalez, X.R.; Reyes, M.; Lenvik, T.; Lund, T.; Blackstad, M.; et al. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 2002, 418, 41–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Wang, F.; Zou, Z.; Dong, S.; Shi, C.; Wang, J.; Ran, X.; Su, Y. Long-term repopulation effects of donor BMDCs on intestinal epithelium. Dig. Dis. Sci. 2010, 55, 2182–2193. [Google Scholar] [CrossRef]
- Liu, D.; Wang, F.; Zou, Z.; Dong, S.; Wang, J.; Ran, X.; Li, C.; Shi, C.; Su, Y. Bone marrow derivation of interstitial cells of cajal in small intestine following intestinal injury. J. Biomed. Biotechnol. 2010, 2010, 164986. [Google Scholar] [CrossRef] [Green Version]
- Ishii, S.; Tsuji, S.; Tsujii, M.; Nishida, T.; Watabe, K.; Iijima, H.; Takehara, T.; Kawano, S.; Hayashi, N. Restoration of gut motility in Kit-deficient mice by bone marrow transplantation. J. Gastroenterol. 2009, 44, 834–841. [Google Scholar] [CrossRef]
- McCann, C.J.; Hwang, S.J.; Hennig, G.W.; Ward, S.M.; Sanders, K.M. Bone Marrow Derived Kit-positive Cells Colonize the Gut but Fail to Restore Pacemaker Function in Intestines Lacking Interstitial Cells of Cajal. J. Neurogastroenterol. Motil. 2014, 20, 326–337. [Google Scholar] [CrossRef] [Green Version]
- Joddar, B.; Tasnim, N.; Thakur, V.; Kumar, A.; McCallum, R.W.; Chattopadhyay, M. Delivery of Mesenchymal Stem Cells from Gelatin-Alginate Hydrogels to Stomach Lumen for Treatment of Gastroparesis. Bioengineering 2018, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Rackham, O.J.L.; Firas, J.; Fang, H.; Oates, M.E.; Holmes, M.L.; Knaupp, A.S.; The, F.C.; Suzuki, H.; Nefzger, C.M.; Daub, C.O.; et al. A predictive computational framework for direct reprogramming between human cell types. Nat. Genet. 2016, 48, 331. [Google Scholar] [CrossRef] [Green Version]



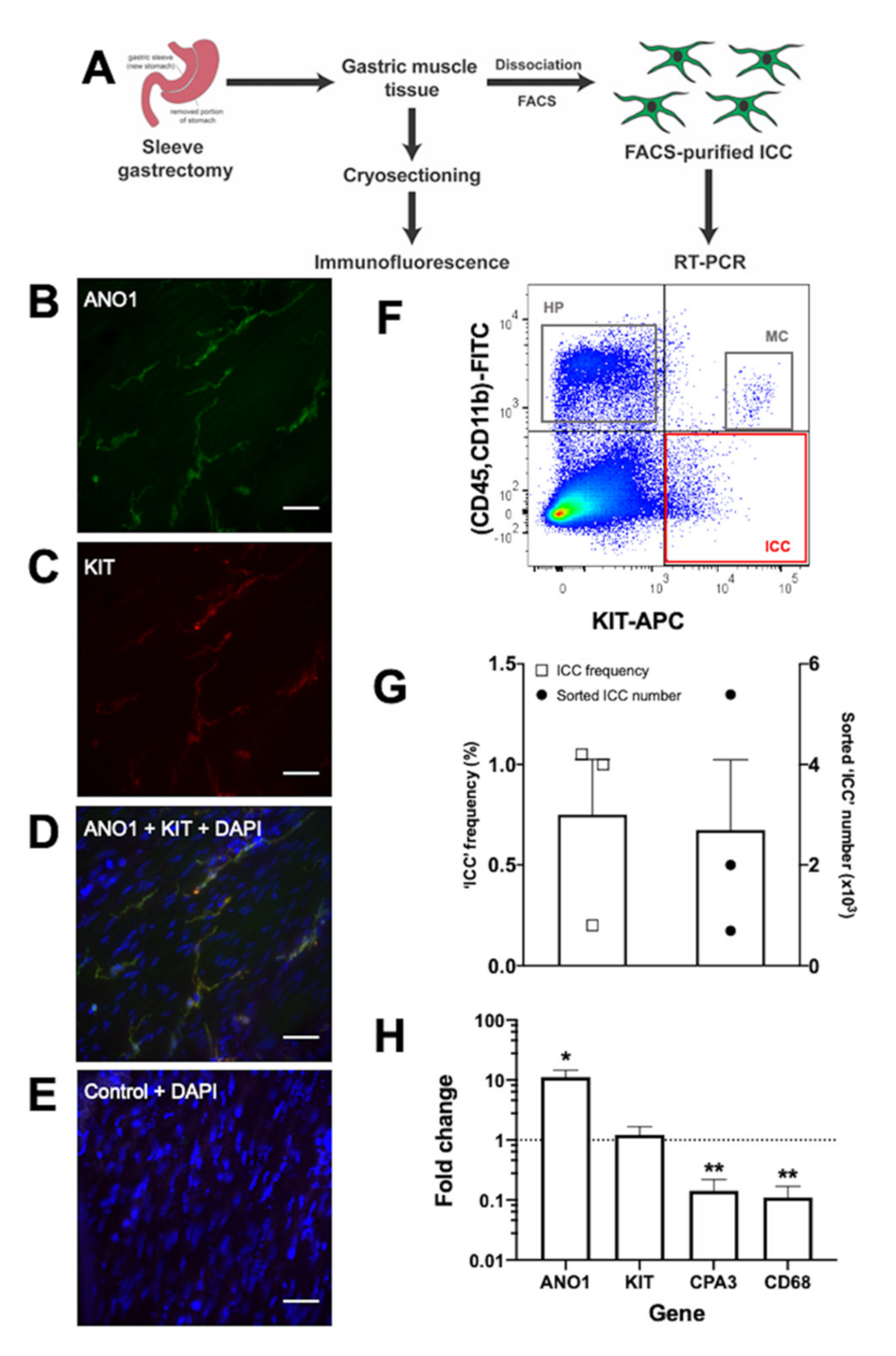{kind=link}
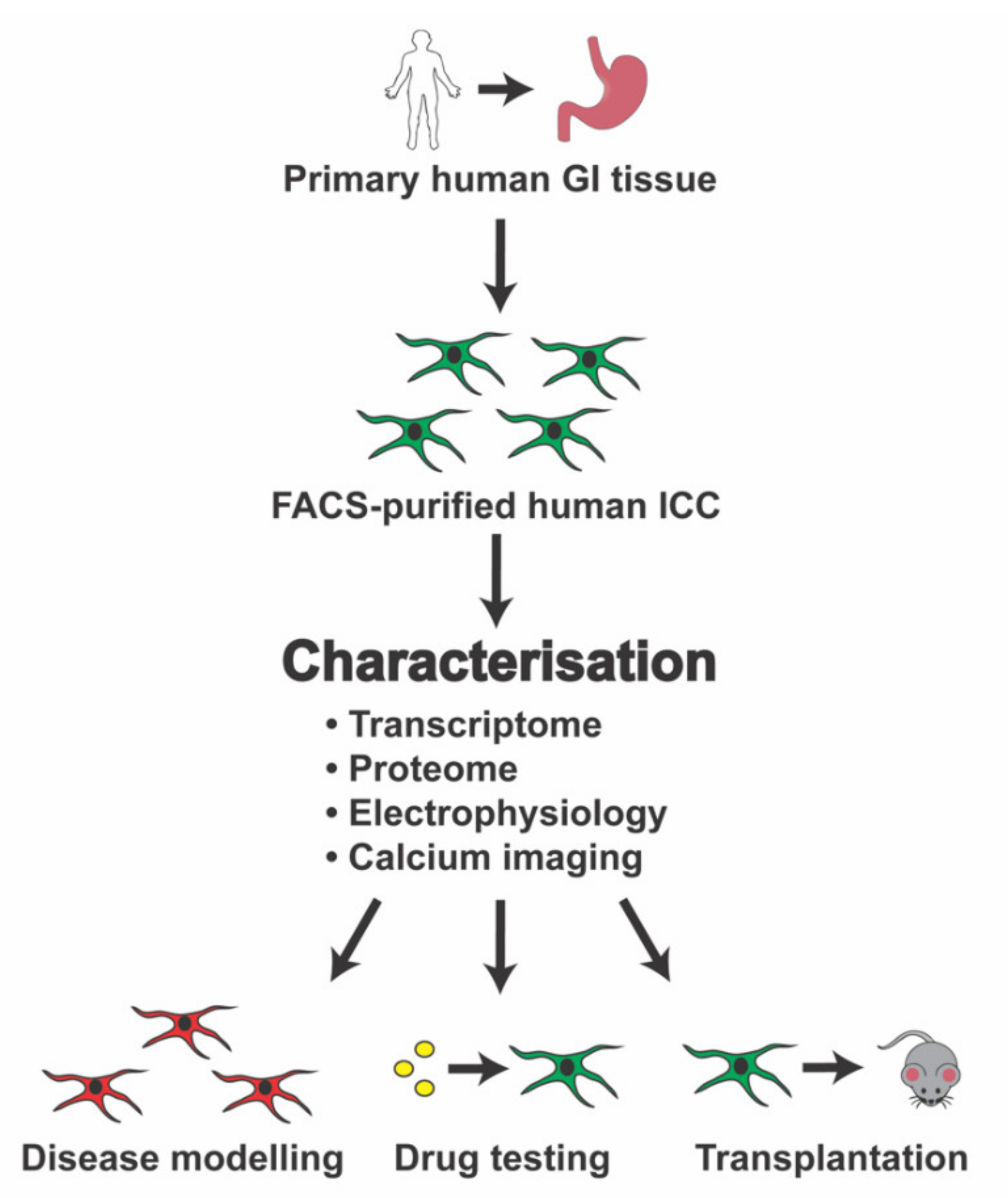{kind=link}
| Ref(s) 1 | Source | Purification Method | Downstream Technique(s) | Major Finding(s) |
|---|---|---|---|---|
| [26] | Small intestinal cultures and tissue | Collected Kit+ cells through micro-pipette | RT-PCR |
|
| [25] | Small intestinal cultures and tissue | MACS-purified Kit+ ICC | Flow cytometry Rhod-2 (calcium) and TMRM imaging |
|
| [27,37] | Small intestinal and gastric tissue | MACS- and/or FACS-purified Kit+ ICC | Flow cytometry RT-PCR |
|
| [28] | Small intestinal tissue | FACS-purified substance P+Kit+ ICC-DMP | Immunofluorescence RT-PCR |
|
| [24] | Small intestinal tissue | FACS-purified Kit+ ICC-MY and substance P+ Kit+ ICC-DMP | RT-PCR Microarray |
|
| [38] | Gastric tissue | FACS-purified Kit+ ICC | Flow cytometry |
|
| [39] | Small intestinal tissue (Kit+/copGFP) | FACS-purified GFP+ cells | RT-PCR |
|
| [40] | Small intestinal and colon tissue (Kit+/copGFP) | FACS-purified GFP+ ICC | RNA-sequencing (RNA-seq) |
|
| [41,42] | Small intestinal cultures | Identified single Kit+ ICC | Whole-cell patch clamping |
|
| [43] | Small intestinal tissue | Identified single Kit+ ICC | Whole-cell patch clamping |
|
| [34,44] | Small intestinal tissue (Kit+/copGFP) | Identified single GFP+ ICC | Whole-cell patch clamping |
|
| Ref(s) 1 | Source | Purification Method | Downstream Technique(s) | Major Finding(s) |
|---|---|---|---|---|
| [60] | Small intestinal tissue (gastric bypass) | Identified single KIT+ ICC | Single-cell RT-PCR Whole-cell patch clamping |
|
| [59] | Small intestinal tissue (gastric bypass) | Identified ICC-MY within muscle strips 2 | Electrophysiology Calcium imaging |
|
| [28] | Small intestinal tissue (gastric bypass) | Identified Substance P+ KIT+ ICC-DMP | Immunofluorescence |
|
| [55] | Gastric tissue (sleeve gastrectomy) | FACS-purified KIT+ ICC | Microarray |
|
| [61] | Colon tissue (colon cancer) | FACS-purified KIT+ ICC | Flow cytometry Immunofluorescence |
|
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foong, D.; Zhou, J.; Zarrouk, A.; Ho, V.; O’Connor, M.D. Understanding the Biology of Human Interstitial Cells of Cajal in Gastrointestinal Motility. Int. J. Mol. Sci. 2020, 21, 4540. https://doi.org/10.3390/ijms21124540
Foong D, Zhou J, Zarrouk A, Ho V, O’Connor MD. Understanding the Biology of Human Interstitial Cells of Cajal in Gastrointestinal Motility. International Journal of Molecular Sciences. 2020; 21(12):4540. https://doi.org/10.3390/ijms21124540
Chicago/Turabian StyleFoong, Daphne, Jerry Zhou, Ali Zarrouk, Vincent Ho, and Michael D. O’Connor. 2020. "Understanding the Biology of Human Interstitial Cells of Cajal in Gastrointestinal Motility" International Journal of Molecular Sciences 21, no. 12: 4540. https://doi.org/10.3390/ijms21124540
APA StyleFoong, D., Zhou, J., Zarrouk, A., Ho, V., & O’Connor, M. D. (2020). Understanding the Biology of Human Interstitial Cells of Cajal in Gastrointestinal Motility. International Journal of Molecular Sciences, 21(12), 4540. https://doi.org/10.3390/ijms21124540