The Abuse Potential of Novel Synthetic Phencyclidine Derivative 1-(1-(4-Fluorophenyl)Cyclohexyl)Piperidine (4′-F-PCP) in Rodents

 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. 4′-F-PCP Increased Locomotor and Rotational Activities in Mice
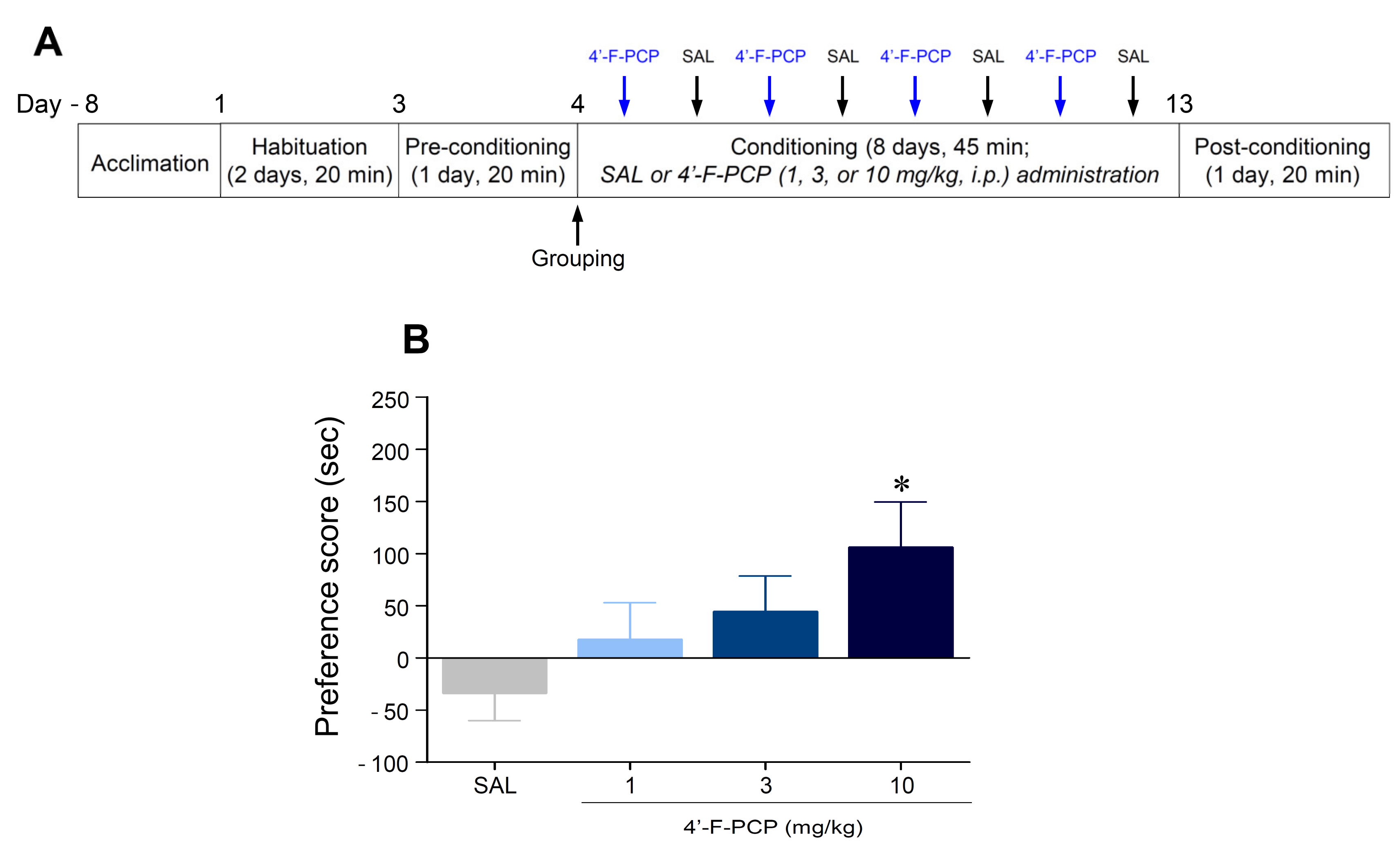
2.2. 4′-F-PCP Induced Conditioned Place Preference in Mice
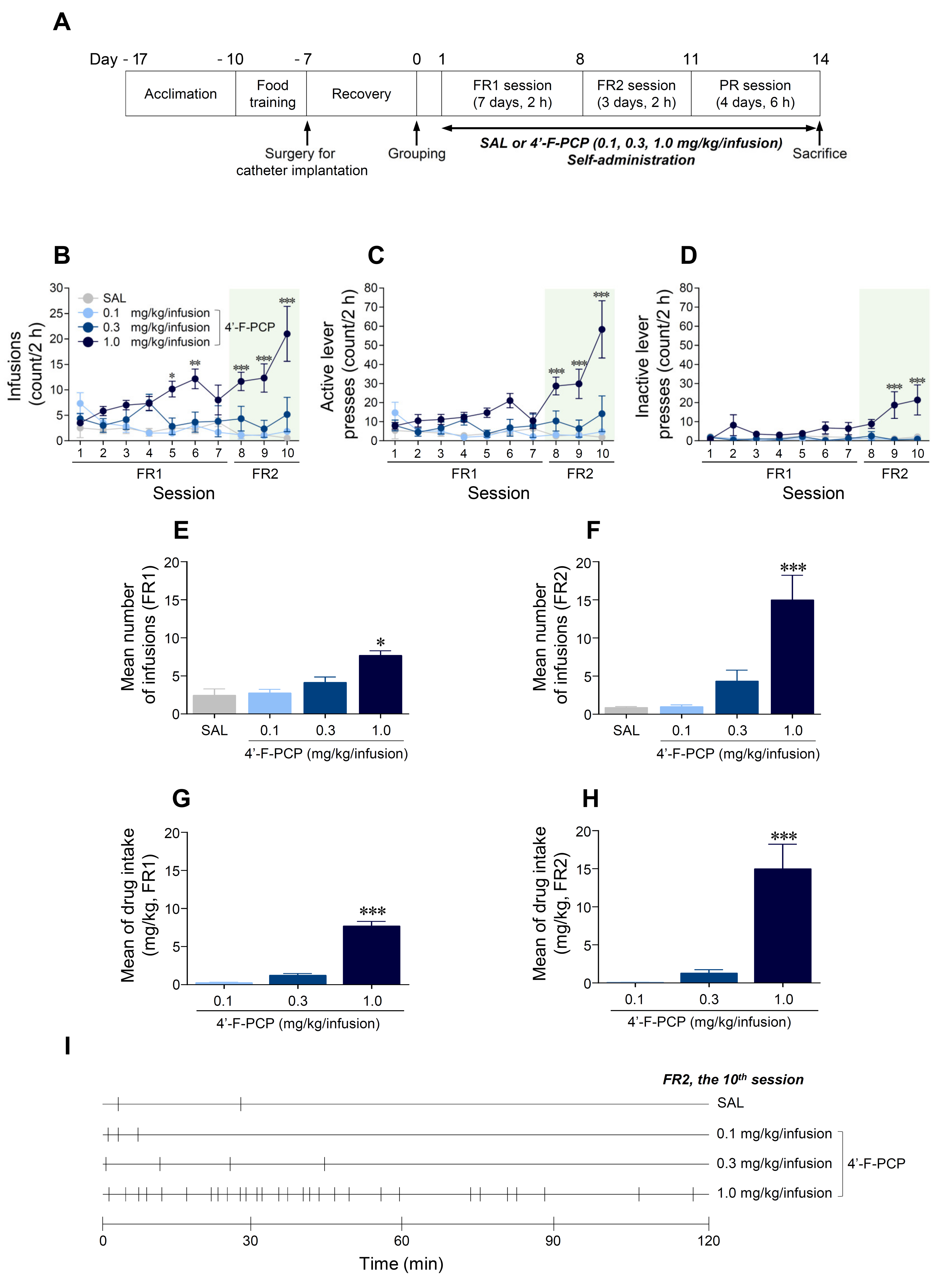
2.3. 4′-F-PCP Induced Self-Administration under Fixed Ratio Schedule in Rats
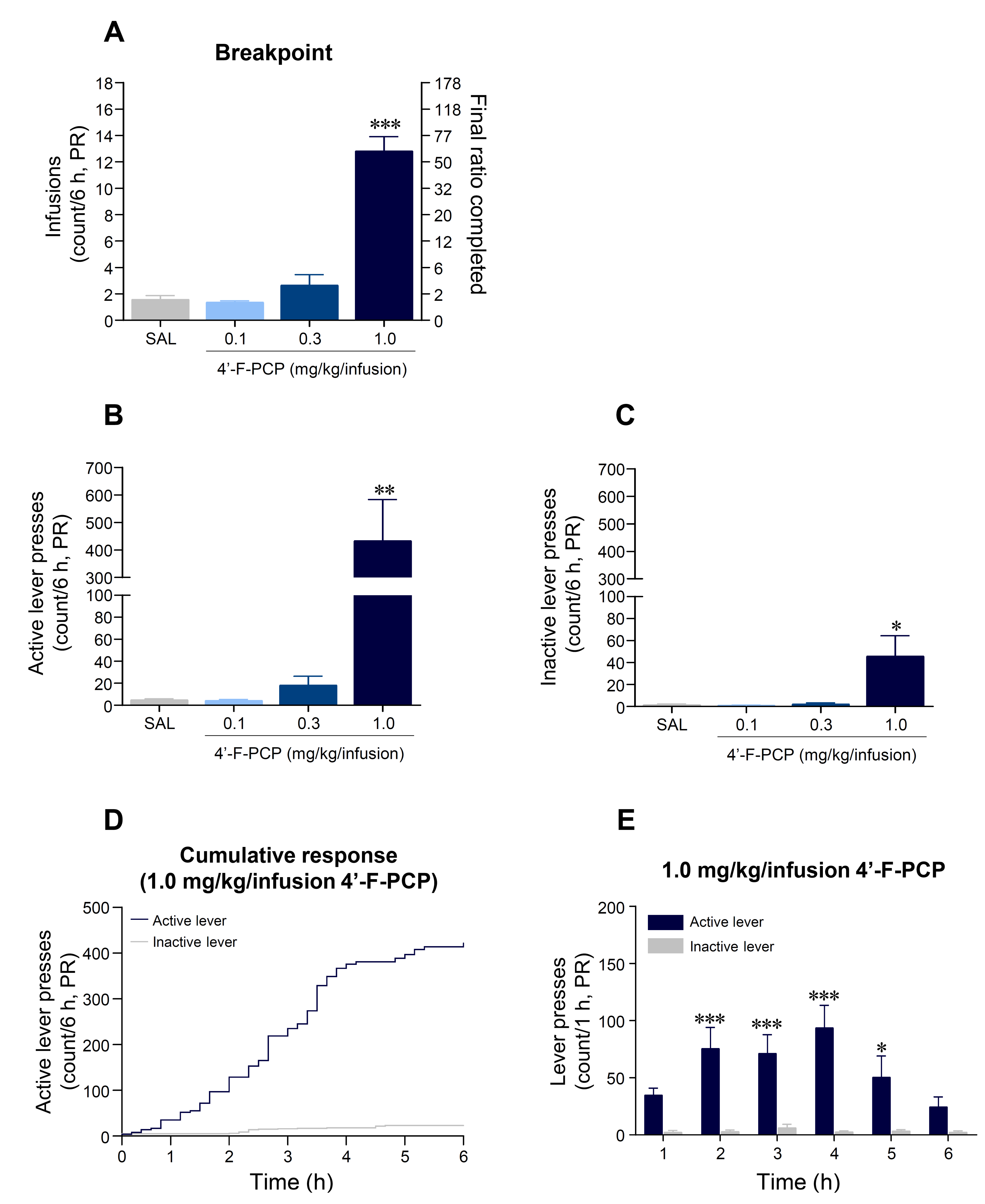
2.4. 4′-F-PCP Produced Higher Breakpoint in Self-Administration under Progressive Ratio Schedule in Rats
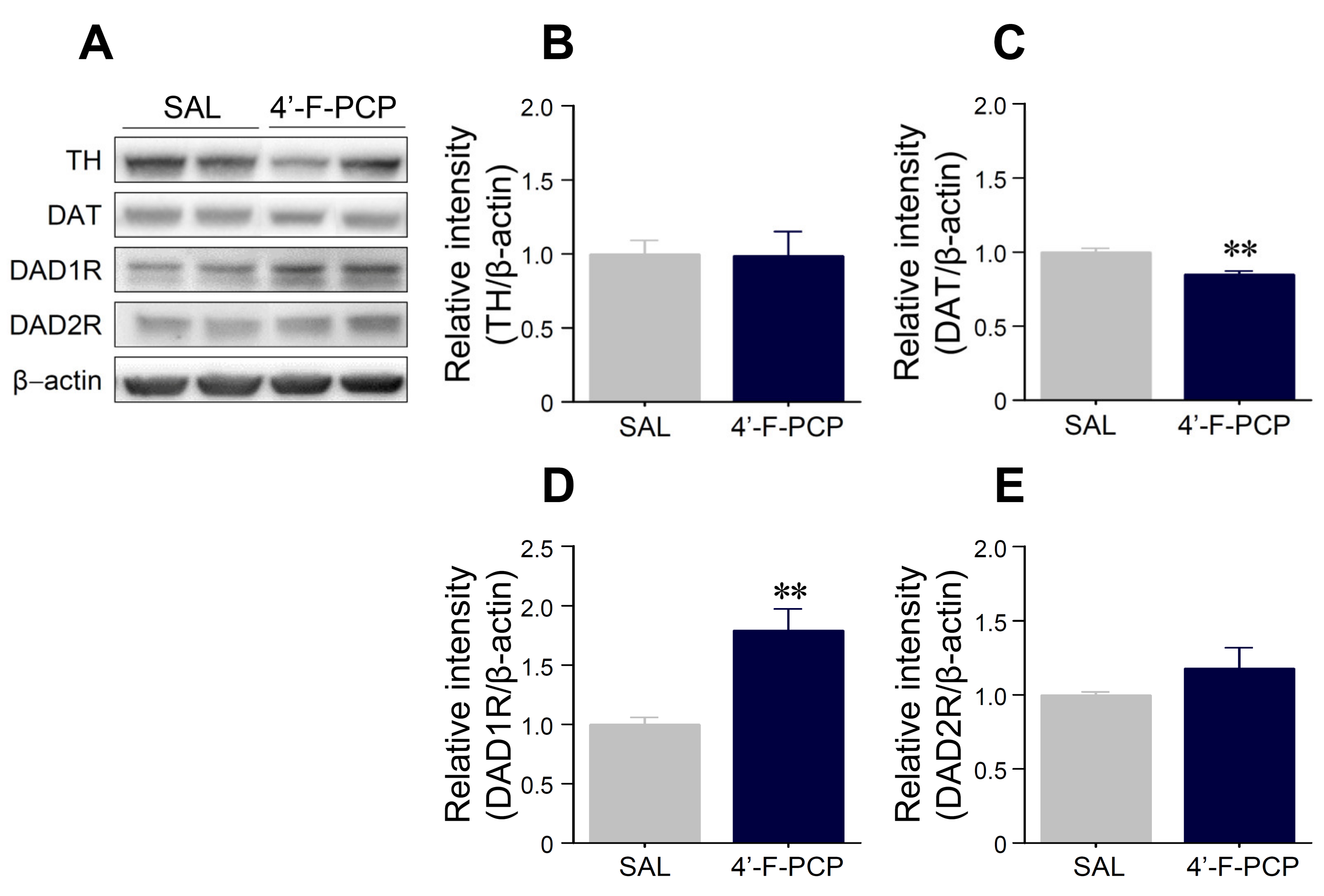
2.5. 4′-F-PCP Self-Administration Altered Expression of Dopamine-Related Proteins in the Nucleus Accumbens
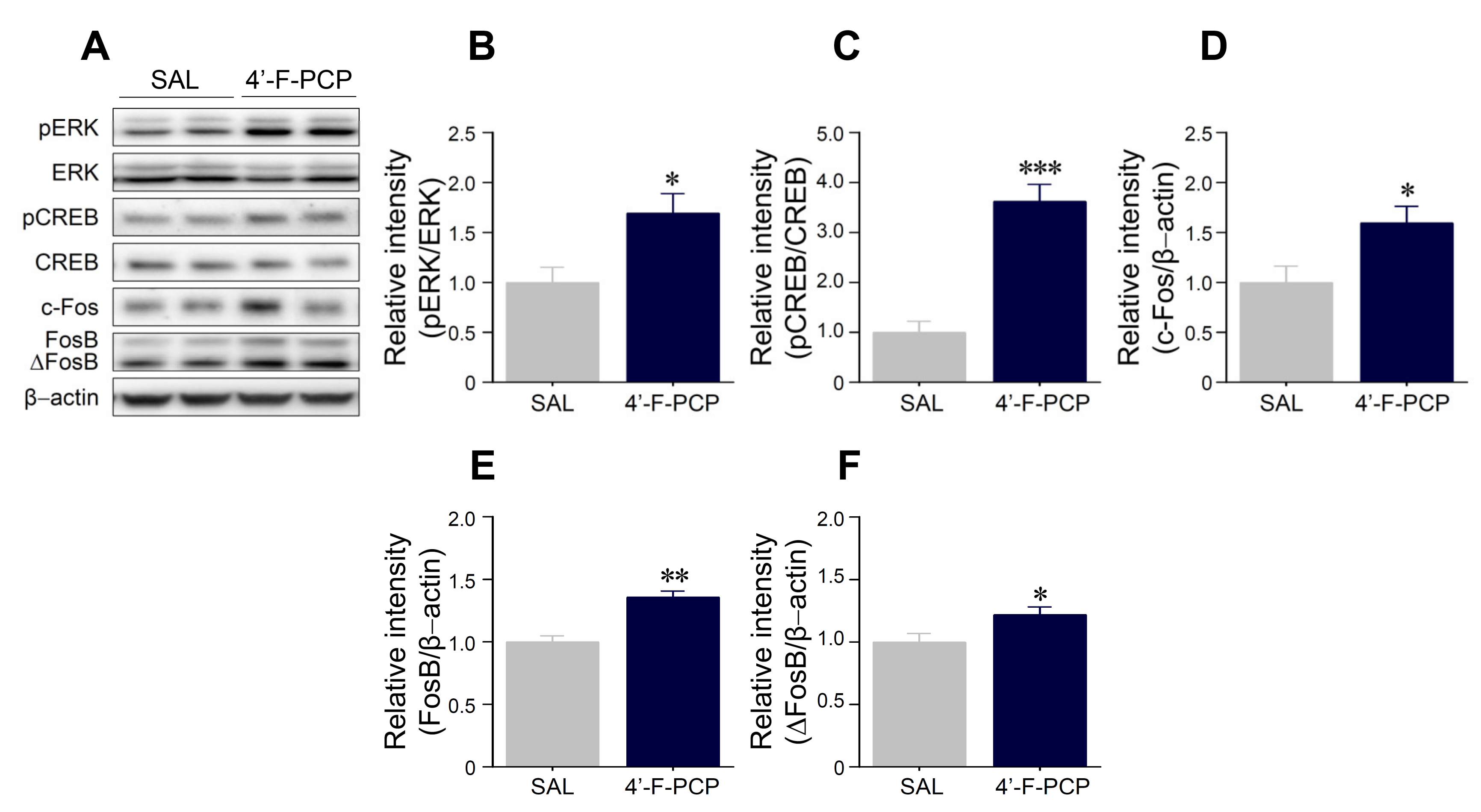
2.6. 4′-F-PCP Self-Administration Altered Expression of pERK, pCREB, c-Fos, and FosB/ΔFosB in the Nucleus Accumbens
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Open-Field Test
4.4. Conditioned Place Preference Paradigm
4.5. Food Training and Catheter Implantation Surgery for Self-Administration
4.6. Drug Self-Administration
4.7. Western Immunoblotting Procedures
4.8. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3-MeO-PCP | 3-Methoxyphencyclidine |
| 4-MeO-PCP | 4-Methoxyphencyclidine |
| 4′-F-PCP | 1-(1-(4-fluorophenyl)cyclohexyl)piperidine |
| CPP | Conditioned Place Preference |
| CREB | cyclic AMP Response-Element Binding Protein |
| DA | Dopamine |
| DAD1R | Dopamine D1 Receptor |
| DAD2R | Dopamine D2 Receptor |
| DAergic | Dopaminergic |
| DAT | Dopamine Transporter |
| ERK | Extracellular Signal-regulated Kinase |
| FR | Fixed Ratio |
| I.V. | intravenous |
| MAPK | Mitogen-activated Protein Kinase |
| NAc | Nucleus Accumbens |
| NMDAR | N-methyl-D-aspartate Glutamate Receptor |
| PCP | Phencyclidine |
| PR | Progressive Ratio |
| TH | Tyrosine Hydroxylase |
| VTA | Ventral Tegmental Area |
References
- Thornton, S.; Lisbon, D.; Lin, T.; Gerona, R. Beyond Ketamine and Phencyclidine: Analytically Confirmed Use of Multiple Novel Arylcyclohexylamines. J. Psychoact. Drugs 2017, 49, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Lodge, D.; Mercier, M.S. Ketamine and phencyclidine: The good, the bad and the unexpected. Br. J. Pharm. 2015, 172, 4254–4276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohmori, T.; Koyama, T.; Nakamura, F.; Wang, P.; Yamashita, I. Effect of phencyclidine on spontaneous and N-methyl-D-aspartate (NMDA)-induced efflux of dopamine from superfused slices of rat striatum. Neuropharmacology 1992, 31, 461–467. [Google Scholar] [CrossRef]
- Balster, R.L.; Mansbach, R.S.; Shelton, K.L.; Nicholson, K.L.; Grech, D.M.; Wiley, J.L.; Li, H.; Weber, E. Behavioral pharmacology of two novel substituted quinoxalinedione glutamate antagonists. Behav. Pharm. 1995, 6, 577–589. [Google Scholar] [CrossRef]
- Smith, R.C.; Meltzer, H.Y.; Arora, R.C.; Davis, J.M. Effects of phencyclidine on [3H]catecholamine and [3H]serotonin uptake in synaptosomal preparations from rat brain. Biochem. Pharm. 1977, 26, 1435–1439. [Google Scholar] [CrossRef]
- Nishijima, K.; Kashiwa, A.; Hashimoto, A.; Iwama, H.; Umino, A.; Nishikawa, T. Differential effects of phencyclidine and methamphetamine on dopamine metabolism in rat frontal cortex and striatum as revealed by in vivo dialysis. Synapse 1996, 22, 304–312. [Google Scholar] [CrossRef]
- Morris, H.; Wallach, J. From PCP to MXE: A comprehensive review of the non-medical use of dissociative drugs. Drug Test. Anal. 2014, 6, 614–632. [Google Scholar] [CrossRef]
- Wallach, J.; Brandt, S.D. Phencyclidine-Based New Psychoactive Substances. Handb. Exp. Pharm. 2018, 252, 261–303. [Google Scholar] [CrossRef]
- Pang, K.; Johnson, S.W.; Maayani, S.; Freedman, R. Structure-activity relationships of phencyclidine derivatives in rat cerebellum. Pharm. Biochem. Behav. 1986, 24, 127–134. [Google Scholar] [CrossRef]
- Cone, E.J.; McQuinn, R.L.; Shannon, H.E. Structure-activity relationship studies of phencyclidine derivatives in rats. J. Pharm. Exp. 1984, 228, 147–153. [Google Scholar]
- Blair, J.B.; Kurrasch-Orbaugh, D.; Marona-Lewicka, D.; Cumbay, M.G.; Watts, V.J.; Barker, E.L.; Nichols, D.E. Effect of ring fluorination on the pharmacology of hallucinogenic tryptamines. J. Med. Chem. 2000, 43, 4701–4710. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Wallach, J.; Adejare, A. Syntheses and N-methyl-D-aspartate receptor antagonist pharmacology of fluorinated arylcycloheptylamines. Med. Chem. 2014, 10, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Ogunbadeniyi, A.M.; Adejare, A. Syntheses of fluorinated phencyclidine analogs. J. Fluor. Chem. 2002, 114, 39–42. [Google Scholar] [CrossRef]
- Bardo, M.T.; Bevins, R.A. Conditioned place preference: What does it add to our preclinical understanding of drug reward? Psychopharmacology 2000, 153, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Huskinson, S.L.; Naylor, J.E.; Rowlett, J.K.; Freeman, K.B. Predicting abuse potential of stimulants and other dopaminergic drugs: Overview and recommendations. Neuropharmacology 2014, 87, 66–80. [Google Scholar] [CrossRef] [Green Version]
- Ryu, I.S.; Yoon, S.S.; Choi, M.J.; Lee, Y.E.; Kim, J.S.; Kim, W.H.; Cheong, J.H.; Kim, H.J.; Jang, C.G.; Lee, Y.S.; et al. The potent psychomotor, rewarding and reinforcing properties of 3-fluoromethamphetamine in rodents. Addict. Biol. 2019, e12846. [Google Scholar] [CrossRef]
- Phillips, M.; Wang, C.; Johnson, K.M. Pharmacological characterization of locomotor sensitization induced by chronic phencyclidine administration. J. Pharm. Exp. 2001, 296, 905–913. [Google Scholar]
- Abiero, A.; Botanas, C.J.; Custodio, R.J.; Sayson, L.V.; Kim, M.; Lee, H.J.; Kim, H.J.; Lee, K.W.; Jeong, Y.; Seo, J.W.; et al. 4-MeO-PCP and 3-MeO-PCMo, new dissociative drugs, produce rewarding and reinforcing effects through activation of mesolimbic dopamine pathway and alteration of accumbal CREB, deltaFosB, and BDNF levels. Psychopharmacology 2020, 237, 757–772. [Google Scholar] [CrossRef]
- Noda, Y.; Miyamoto, Y.; Mamiya, T.; Kamei, H.; Furukawa, H.; Nabeshima, T. Involvement of dopaminergic system in phencyclidine-induced place preference in mice pretreated with phencyclidine repeatedly. J. Pharm. Exp. 1998, 286, 44–51. [Google Scholar]
- Botanas, C.J.; de la Pena, J.B.; Dela Pena, I.J.; Tampus, R.; Yoon, R.; Kim, H.J.; Lee, Y.S.; Jang, C.G.; Cheong, J.H. Methoxetamine, a ketamine derivative, produced conditioned place preference and was self-administered by rats: Evidence of its abuse potential. Pharm. Biochem. Behav. 2015, 133, 31–36. [Google Scholar] [CrossRef]
- Campbell, U.C.; Thompson, S.S.; Carroll, M.E. Acquisition of oral phencyclidine (PCP) self-administration in rhesus monkeys: Effects of dose and an alternative non-drug reinforcer. Psychopharmacology 1998, 137, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Marquis, K.L.; Webb, M.G.; Moreton, J.E. Effects of fixed ratio size and dose on phencyclidine self-administration by rats. Psychopharmacology 1989, 97, 179–182. [Google Scholar] [CrossRef]
- Adinoff, B. Neurobiologic processes in drug reward and addiction. Harv. Rev. Psychiatry 2004, 12, 305–320. [Google Scholar] [CrossRef]
- Arias-Carrion, O.; Stamelou, M.; Murillo-Rodriguez, E.; Menendez-Gonzalez, M.; Poppel, E. Dopaminergic reward system: A short integrative review. Int. Arch. Med. 2010, 3, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, K.; Morishima, M.; Sakai, K.; Kawaguchi, Y. Dopaminergic control of motivation and reinforcement learning: A closed-circuit account for reward-oriented behavior. J. Neurosci. 2013, 33, 8866–8890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNamara, R.K.; Logue, A.; Stanford, K.; Xu, M.; Zhang, J.; Richtand, N.M. Dose-response analysis of locomotor activity and stereotypy in dopamine D3 receptor mutant mice following acute amphetamine. Synapse 2006, 60, 399–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fessler, R.G.; Sturgeon, R.D.; Meltzer, H.Y. Phencyclidine-induced ipsilateral rotation in rats with unilateral 6-hydroxydopamine-induced lesions of the substantia nigra. Life Sci. 1979, 24, 1281–1288. [Google Scholar] [CrossRef]
- French, E.D.; Vantini, G. Phencyclidine-induced locomotor activity in the rat is blocked by 6-hydroxydopamine lesion of the nucleus accumbens: Comparisons to other psychomotor stimulants. Psychopharmacology 1984, 82, 83–88. [Google Scholar] [CrossRef]
- Imre, G.; Fokkema, D.S.; Den Boer, J.A.; Ter Horst, G.J. Dose-response characteristics of ketamine effect on locomotion, cognitive function and central neuronal activity. Brain Res. Bull. 2006, 69, 338–345. [Google Scholar] [CrossRef]
- Halberstadt, A.L.; Slepak, N.; Hyun, J.; Buell, M.R.; Powell, S.B. The novel ketamine analog methoxetamine produces dissociative-like behavioral effects in rodents. Psychopharmacol 2016, 233, 1215–1225. [Google Scholar] [CrossRef]
- Marcus, M.M.; Mathe, J.M.; Nomikos, G.G.; Svensson, T.H. Effects of competitive and non-competitive NMDA receptor antagonists on dopamine output in the shell and core subdivisions of the nucleus accumbens. Neuropharmacology 2001, 40, 482–490. [Google Scholar] [CrossRef]
- Richardson, N.R.; Roberts, D.C. Progressive ratio schedules in drug self-administration studies in rats: A method to evaluate reinforcing efficacy. J. Neurosci. Methods 1996, 66, 1–11. [Google Scholar] [CrossRef]
- Roberts, D.C.; Loh, E.A.; Vickers, G. Self-administration of cocaine on a progressive ratio schedule in rats: Dose-response relationship and effect of haloperidol pretreatment. Psychopharmacology 1989, 97, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Rodefer, J.S.; Carroll, M.E. Concurrent progressive-ratio schedules to compare reinforcing effectiveness of different phencyclidine (PCP) concentrations in rhesus monkeys. Psychopharmacology 1999, 144, 163–174. [Google Scholar] [CrossRef]
- French, E.D.; Lopez, M.; Peper, S.; Kamenka, J.M.; Roberts, D.C. A comparison of the reinforcing efficacy of PCP, the PCP derivatives TCP and BTCP, and cocaine using a progressive ratio schedule in the rat. Behav. Pharm. 1995, 6, 223–228. [Google Scholar] [CrossRef]
- Kirk, K.L.; Cantacuzene, D.; Nimitkitpaisan, Y.; McCulloh, D.; Padgett, W.L.; Daly, J.W.; Creveling, C.R. Synthesis and biological properties of 2-, 5-, and 6-fluoronorepinephrines. J. Med. Chem. 1979, 22, 1493–1497. [Google Scholar] [CrossRef]
- Levine, M.S.; Howard-Butcher, S. Behavioral effects of phencyclidine and some of its metabolites in developing cats. Pharm. Biochem. Behav. 1986, 25, 359–363. [Google Scholar] [CrossRef]
- Vaughan, R.A.; Foster, J.D. Mechanisms of dopamine transporter regulation in normal and disease states. Trends Pharm. Sci. 2013, 34, 489–496. [Google Scholar] [CrossRef] [Green Version]
- White, R.B.; Thomas, M.G. Moving beyond tyrosine hydroxylase to define dopaminergic neurons for use in cell replacement therapies for Parkinson’s disease. CNS Neurol. Disord. Drug Targets 2012, 11, 340–349. [Google Scholar] [CrossRef]
- Sun, W.L.; Quizon, P.M.; Zhu, J. Molecular Mechanism: ERK Signaling, Drug Addiction, and Behavioral Effects. Prog. Mol. Biol. Transl. Sci. 2016, 137, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Robison, A.J.; Nestler, E.J. Transcriptional and epigenetic mechanisms of addiction. Nat. Rev. Neurosci. 2011, 12, 623–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M. c-Fos is an intracellular regulator of cocaine-induced long-term changes. Ann. N. Y. Acad. Sci. 2008, 1139, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrotti, L.I.; Weaver, R.R.; Robison, B.; Renthal, W.; Maze, I.; Yazdani, S.; Elmore, R.G.; Knapp, D.J.; Selley, D.E.; Martin, B.R.; et al. Distinct patterns of DeltaFosB induction in brain by drugs of abuse. Synapse 2008, 62, 358–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yager, L.M.; Garcia, A.F.; Wunsch, A.M.; Ferguson, S.M. The ins and outs of the striatum: Role in drug addiction. Neuroscience 2015, 301, 529–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, G.H. Complete Latin Squares and Related Experimental Designs. J. R. Stat. Soc. Ser. B Methodol. 1979, 41, 253–262. [Google Scholar] [CrossRef]
- Yang, J.H.; Seo, S.Y.; Oh, J.H.; Ryu, I.S.; Kim, J.; Lee, D.K.; Ryu, Y.; Choe, E.S. Activation of Protein Kinase G After Repeated Cocaine Administration Is Necessary for the Phosphorylation of alpha-Amino-3-Hydroxy-5-Methyl-4-Isoxazolepropionic Acid Receptor GluA1 at Serine 831 in the Rat Nucleus Accumbens. Front. Mol. Neurosci. 2018, 11, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryu, I.S.; Kim, O.-H.; Lee, Y.E.; Kim, J.S.; Li, Z.-H.; Kim, T.W.; Lim, R.-N.; Lee, Y.J.; Cheong, J.H.; Kim, H.J.; et al. The Abuse Potential of Novel Synthetic Phencyclidine Derivative 1-(1-(4-Fluorophenyl)Cyclohexyl)Piperidine (4′-F-PCP) in Rodents. Int. J. Mol. Sci. 2020, 21, 4631. https://doi.org/10.3390/ijms21134631
Ryu IS, Kim O-H, Lee YE, Kim JS, Li Z-H, Kim TW, Lim R-N, Lee YJ, Cheong JH, Kim HJ, et al. The Abuse Potential of Novel Synthetic Phencyclidine Derivative 1-(1-(4-Fluorophenyl)Cyclohexyl)Piperidine (4′-F-PCP) in Rodents. International Journal of Molecular Sciences. 2020; 21(13):4631. https://doi.org/10.3390/ijms21134631
Chicago/Turabian StyleRyu, In Soo, Oc-Hee Kim, Young Eun Lee, Ji Sun Kim, Zhan-Hui Li, Tae Wan Kim, Ri-Na Lim, Young Ju Lee, Jae Hoon Cheong, Hee Jin Kim, and et al. 2020. "The Abuse Potential of Novel Synthetic Phencyclidine Derivative 1-(1-(4-Fluorophenyl)Cyclohexyl)Piperidine (4′-F-PCP) in Rodents" International Journal of Molecular Sciences 21, no. 13: 4631. https://doi.org/10.3390/ijms21134631
APA StyleRyu, I. S., Kim, O. -H., Lee, Y. E., Kim, J. S., Li, Z. -H., Kim, T. W., Lim, R. -N., Lee, Y. J., Cheong, J. H., Kim, H. J., Lee, Y. S., Steffensen, S. C., Lee, B. H., Seo, J. -W., & Jang, E. Y. (2020). The Abuse Potential of Novel Synthetic Phencyclidine Derivative 1-(1-(4-Fluorophenyl)Cyclohexyl)Piperidine (4′-F-PCP) in Rodents. International Journal of Molecular Sciences, 21(13), 4631. https://doi.org/10.3390/ijms21134631