Oxidation of Erythrocytes Enhance the Production of Reactive Species in the Presence of Artemisinins



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
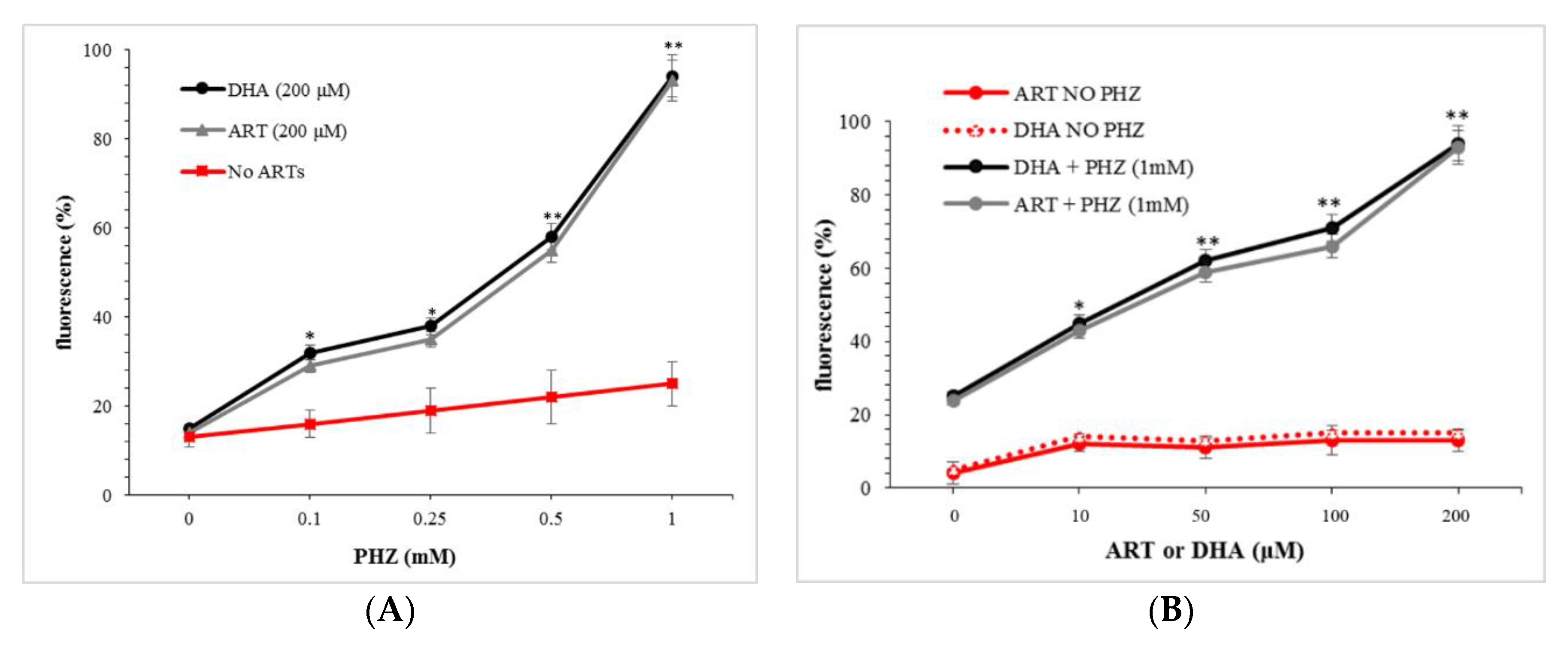
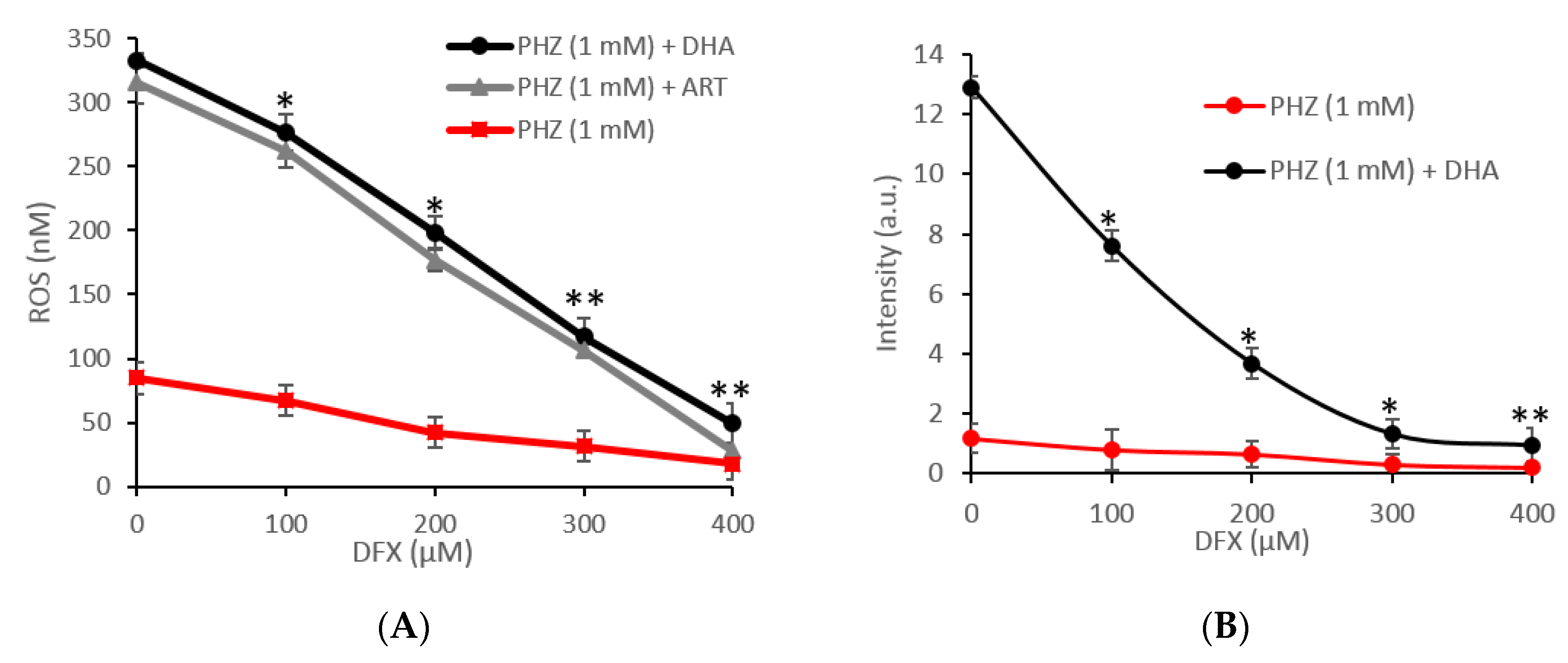
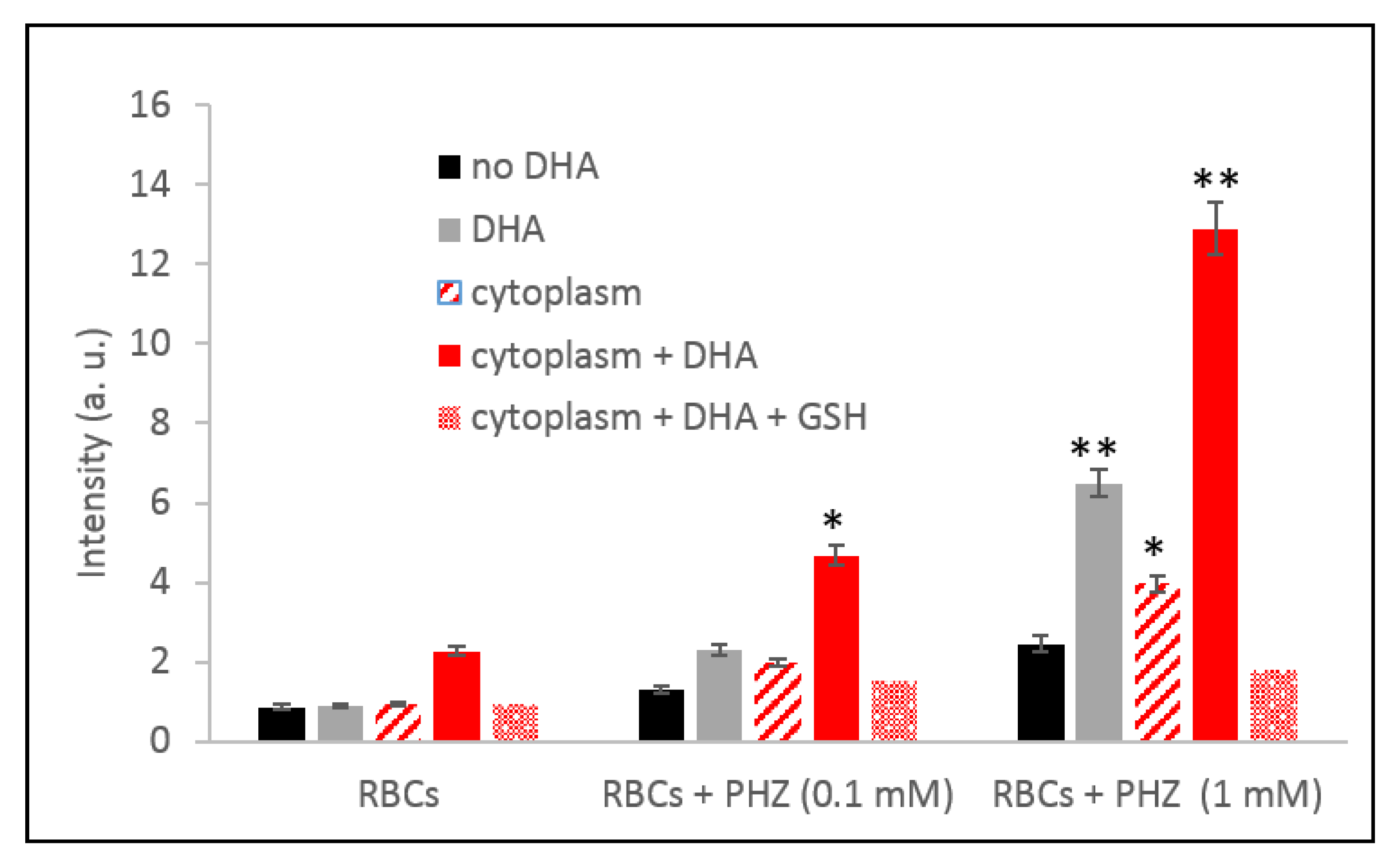
2.1. Global Evaluation of ROS Production for Oxidized RBCs in the Presence of Artemisinins
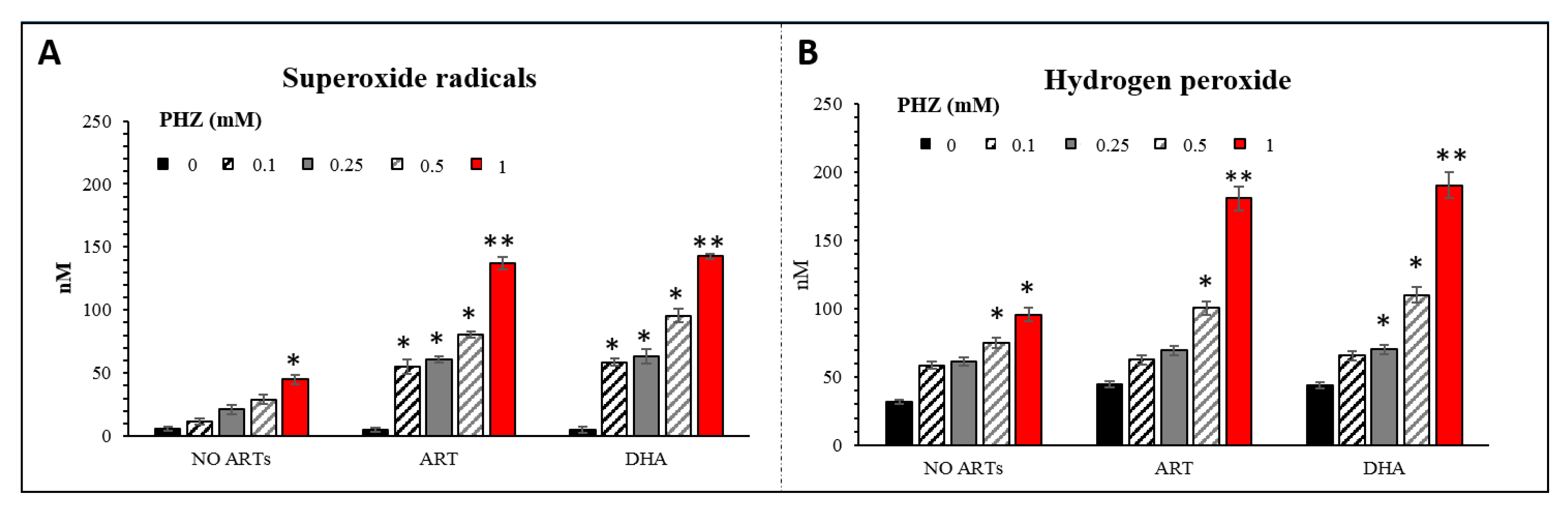
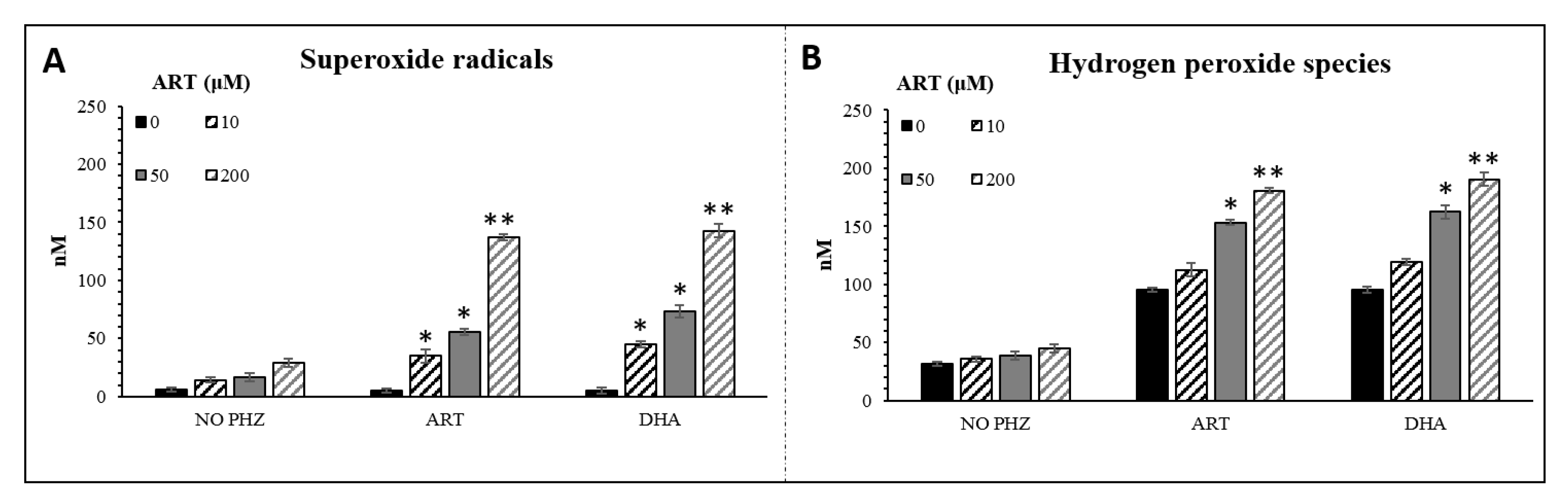
2.2. Specific Evaluation of Superoxide Radicals and Hydrogen Peroxide in Oxidized RBCs in the Presence of Artemisinins
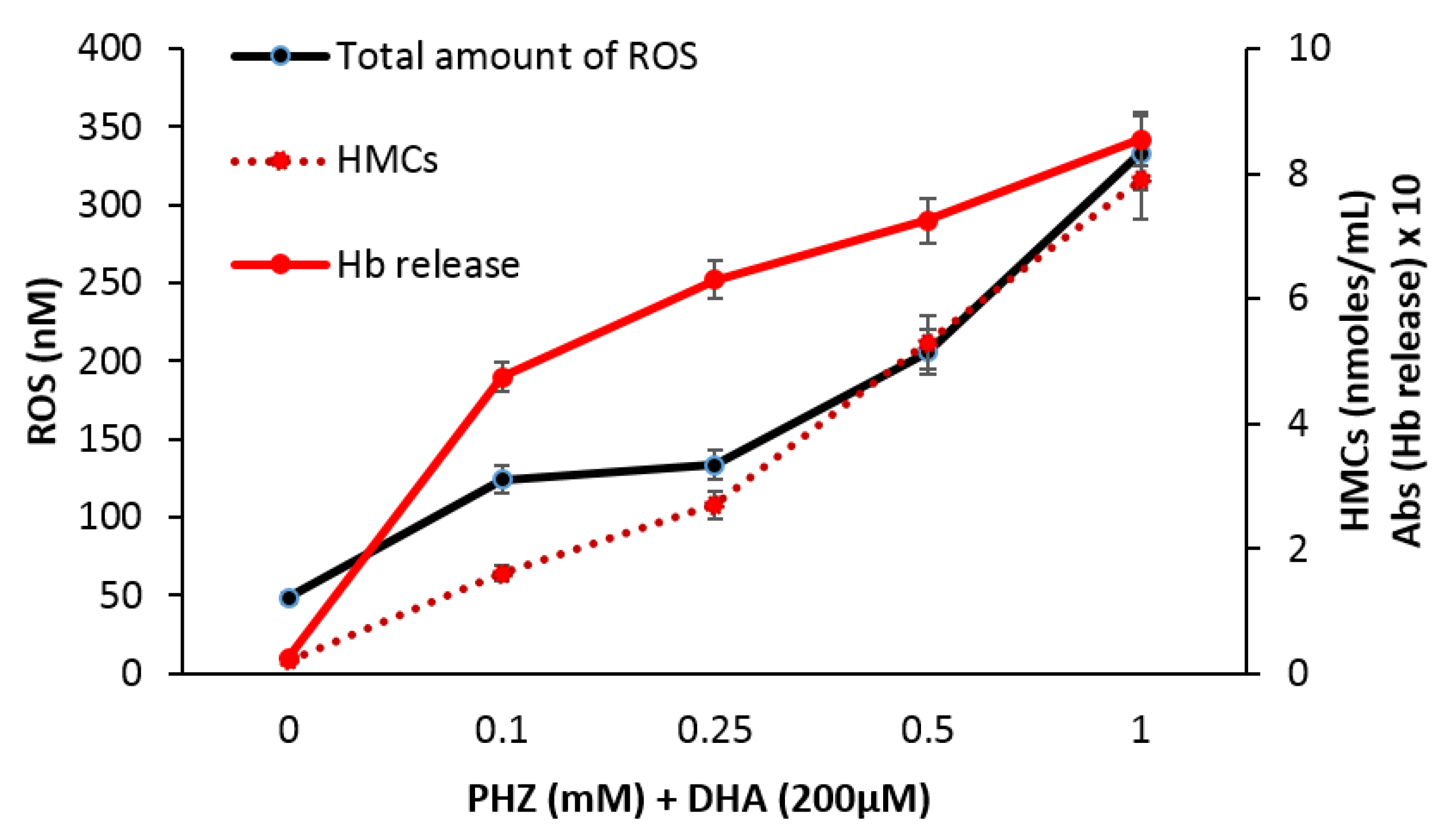
2.3. Correlation of Total Reactive Species with Hemoglobin Byproducts
3. Materials and Methods
3.1. Blood Sample Collection
3.2. Ethics Statement
3.3. Treatment of Red Blood Cells
3.4. Fluorescence Assay
3.5. Liquid Chromatography Coupled to Mass Spectrometry Analysis
3.6. EPR Assay
3.7. Hemoglobin Release Analysis
3.8. Hemichromes Analysis
3.9. Data Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Origa, R. β-Thalassemia. Genet. Med. 2017, 19, 609–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, J.A.; Raghupathy, R.; Manwani, D. Iron overload in sickle cell disease. Adv. Hematol. 2010. [Google Scholar] [CrossRef]
- Dos Santos, J.L.; Pavan, A.R. Advances in Sickle Cell Disease Treatments. Curr. Med. Chem. 2020. [Google Scholar] [CrossRef]
- Thomas, C.; Mackey, M.M.; Diaz, A.A.; Cox, D.P. Hydroxyl radical is produced via the Fenton reaction in submitochondrial particles under oxidative stress: Implications for diseases associated with iron accumulation. Redox Rep. 2009, 14, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.J.; Rose, A.L.; Waite, T.D. Importance of iron complexation for fenton-mediated hydroxyl radical production at circumneutral pH. Front. Mar. Sci. 2016, 3, 134. [Google Scholar] [CrossRef]
- Muhammad, A.; Waziri, A.D.; Forcados, G.E.; Sanusi, B.; Sani, H.; Malami, I.; Abubakar, I.B.; Muhammad, A.; Muhammad, R.A.; Mohammed, H.A. Sickling-suppressive effects of chrysin may be associated with sequestration of deoxy-haemoglobin, 2,3-bisphosphoglycerate mutase, alteration of redox homeostasis and functional chemistry of sickle erythrocytes. Hum. Exp. Toxicol. 2020, 39, 537–546. [Google Scholar] [CrossRef]
- Muhammad, A.; Waziri, A.D.; Forcados, G.E.; Sanusi, B.; Sani, H.; Malami, I.; Abubakar, I.B.; Oluwatoyin, H.Y.; Adinoyi, O.A.; Mohammed, H.A. Sickling-preventive effects of rutin is associated with modulation of deoxygenated haemoglobin, 2,3-bisphosphoglycerate mutase, redox status and alteration of functional chemistry in sickle erythrocytes. Heliyon 2019, 5, e01905. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, A.; Waziri, A.D.; Forcados, G.E.; Sanusi, B.; Sani, H.; Malami, I.; Abubakar, I.B.; Abbah, M.F.; Nelson, A.T.; Musa, B.; et al. Antisickling Effects of Quercetin may be Associated with Modulation of Deoxyhaemoglobin, 2, 3-bisphosphoglycerate mutase, Redox Homeostasis and Alteration of Functional Chemistry in Human Sickle Erythrocytes. Ann. Sci. Technol. 2019, 4, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Kavishe, R.A.; Koenderink, J.B.; Alifrangis, M. Oxidative stress in malaria and artemisinin combination therapy: Pros and Cons. FEBS J. 2017, 284, 2579–2591. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Li, Q.; Wu, J.; Wang, M.; Yu, J. Artemisinin and its derivatives as a repurposing anticancer agent: What else do we need to do? Molecules 2016, 21, 1331. [Google Scholar] [CrossRef]
- Zhang, J. The osteoprotective effects of artemisinin compounds and the possible mechanisms associated with intracellular iron: A review of in vivo and in vitro studies. Environ. Toxicol. Pharmacol. 2020, 103358. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.R.; Fujioka, H.; Williams, P.S.; Chalmers, J.J.; Grimberg, B.; Zimmerman, M.; Zborowski, P.A. Hemoglobin degradation in malaria-infected erythrocytes determined from live cell magnetophoresis. FASEB J. 2006, 20, 747–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coronado, L.M.; Nadovich, C.T.; Spadafora, C. Malarial hemozoin: From target to tool. Biochim. Biophys. Acta-Gen. Subj. 2014, 1840, 2032–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoine, T.; Fisher, N.; Amewu, R.; O’Neill, P.M.; Ward, S.A.; Biagini, G.A. Rapid kill of malaria parasites by artemisinin and semi-synthetic endoperoxides involves ROS-dependent depolarization of the membrane potential. J. Antimicrob. Chemother. 2014, 69, 1005–1016. [Google Scholar] [CrossRef]
- O’Neill, P.M.; Barton, V.E.; Ward, S.A. The molecular mechanism of action of artemisinin—The debate continues. Molecules 2010, 15, 1705–1721. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, H.; Gerhard, G.S. Heme synthesis increases artemisinin-induced radical formation and cytotoxicity that can be suppressed by superoxide scavengers. Chem. Biol. Interact. 2010, 186, 30–35. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, C.J.; Chia, W.N.; Loh, C.C.Y.; Li, Z.; Lee, Y.M.; He, Y.; Yuan, L.X.; Lim, T.K.; Liu, M.; et al. Haem-activated promiscuous targeting of artemisinin in Plasmodium falciparum. Nat. Commun. 2015, 6, 10111. [Google Scholar] [CrossRef]
- Haynes, R.K.; Ho, W.Y.; Chan, H.W.; Fugmann, B.; Stetter, J.; Croft, S.L.; Vivas, L.; Peters, W.; Robinson, B.L. Highly antimalaria-active artemisinin derivatives: Biological activity does not correlate with chemical reactivity. Angew. Chem. Int. Ed. 2004, 43, 1381–1385. [Google Scholar] [CrossRef]
- Haynes, R.K.; Krishna, S. Artemisinins: Activities and actions. Microbes Infect. 2004, 6, 1339–1346. [Google Scholar] [CrossRef]
- Budhraja, A.; Turnis, M.E.; Churchman, M.L.; Kothari, A.; Yang, X.; Xu, H.; Kaminska, E.; Panetta, J.C.; Finkelstein, D.; Mullighan, C.G.; et al. Modulation of navitoclax sensitivity by dihydroartemisinin-mediated MCL-1 repression in BCR-ABL+ B-lineage acute lymphoblastic leukemia. Clin. Cancer Res. 2017, 23, 7558–7568. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.C.; Deng, T.; Fan, M.L.; Lv, W.B.; Liu, J.H.; Yu, B.Y. Synthesis and in vitro antitumor evaluation of dihydroartemisinin-cinnamic acid ester derivatives. Eur. J. Med. Chem. 2016, 107, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, G.; Zhang, S.; Wang, D.; Prabha, P.S.; Zuo, Z. Antitumor Research on Artemisinin and Its Bioactive Derivatives. Nat. Prod. Bioprospect. 2018, 8, 303–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, C.; Feng, H.; Shi, X.; Wang, Y.; Feng, Z.; Liu, J.; Han, Z.; Fu, J.; Fu, Z.; Tong, H. Artesunate down-regulates immunosuppression from colorectal cancer Colon26 and RKO cells in vitro by decreasing transforming growth factor β1 and interleukin-10. Int. Immunopharmacol. 2015, 27, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.C.; Singh, N.P.; Sasaki, T. Development of artemisinin compounds for cancer treatment. Investig. New Drugs 2013, 31, 230–246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.X.; Liu, Z.N.; Ye, J.; Sha, M.; Qian, H.; Bu, X.H.; Luan, Z.Y.; Xu, X.L.; Huang, A.H.; Yuan, D.L.; et al. Artesunate exerts an anti-immunosuppressive effect on cervical cancer by inhibiting PGE2 production and Foxp3 expression. Cell Biol. Int. 2014, 38, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Dadgar, N.; Alavi, S.E.; Esfahani, M.K.M.; Akbarzadeh, A. Study of toxicity effect of pegylated nanoliposomal artemisinin on breast cancer cell line. Indian J. Clin. Biochem. 2013, 28, 410–412. [Google Scholar] [CrossRef] [Green Version]
- Leto, I.; Coronnello, M.; Righeschi, C.; Bergonzi, M.C.; Mini, E.; Bilia, A.R. Enhanced Efficacy of Artemisinin Loaded in Transferrin-Conjugated Liposomes versus Stealth Liposomes against HCT-8 Colon Cancer Cells. ChemMedChem 2016, 11, 1745–1751. [Google Scholar] [CrossRef]
- Creek, D.J.; Ryan, E.; Charman, W.N.; Chiu, F.C.K.; Prankerd, R.J.; Vennerstrom, J.L.; Charman, S.A. Stability of peroxide antimalarials in the presence of human hemoglobin. Antimicrob. Agents Chemother. 2009, 53, 3496–3500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, Y.; Shigemasa, Y.; Hayashi, Y.; Abe, Y.; Ohgushi, I.; Ueno, E. New mode (molecular-sensing) of heinz body formation mechanisms inherent in human erythrocytes: Basis for understanding of clinical aspects of drug-induced hemolytic anemia and the like. J. Bioanal. Biomed. 2013. [Google Scholar] [CrossRef] [Green Version]
- Pantaleo, A.; Ferru, E.; Pau, M.C.; Khadjavi, A.; Mandili, G.; Mattè, A.; Spano, A.; De Franceschi, L.; Pippia, P.; Turrini, F. Band 3 erythrocyte membrane protein acts as redox stress sensor leading to its phosphorylation by p72 Syk. Oxidative Med. Cell. Longev. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, H.P.; Fridovich, I. The Oxidation of Phenylhydrazine: Superoxide and Mechanism. Biochemistry 1976, 15, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Berger, J. Phenylhydrazine haematotoxicity. J. Appl. Biomed. 2007, 5, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Chevion, M.Z.B.; Berenshtein, E. The Role of Transition Metal Ions in Free Radical-Mediated Damage. React. Oxyg. Species Biol. Syst. 2002, 103–131. [Google Scholar] [CrossRef]
- Cohen, G.; Hochstein, P. Generation of Hydrogen Peroxide in Erythrocytes by Hemolytic Agents. Biochemistry 1964, 3, 895–900. [Google Scholar] [CrossRef]
- Schrier, S.L.; Mohandas, N. Globin-chain specificity of oxidation-induced changes in red blood cell membrane properties. Blood 1992, 79, 1586–1592. [Google Scholar] [CrossRef] [Green Version]
- Winterbourn, C.C. Free-radical production and oxidative reactions of hemoglobin. Environ. Health Perspect. 1985, 64, 321–330. [Google Scholar] [CrossRef]
- Rachmilewitz, E.A.; Peisach, J.; Bradley, T.B.; Blumberg, W.E. Role of haemichromes in the formation of inclusion bodies in haemoglobin H disease. Nature 1969, 222, 248–250. [Google Scholar] [CrossRef]
- Rachmilewitz, E.A.; Peisach, J.; Blumberg, W.E. Studies on the stability of oxyhemoglobin A and its constituent chains and their derivatives. J. Biol. Chem. 1971, 246, 3356–3366. [Google Scholar]
- Rachmilewitz, E.A. Denaturation of the normal and abnormal hemoglobin molecule. Semin. Hematol. 1974, 11, 441–462. [Google Scholar]
- Macdonald, V.W. Measuring relative rates of hemoglobin oxidation and denaturation. Methods Enzymol. 1994, 231, 480–490. [Google Scholar] [CrossRef]
- Rifkind, J.M.; Abugo, O.; Levy, A.; Heim, J. Detection, formation, and relebance of hemochromes and hemochromes. Methods Enzymol. 1994, 231, 449–480. [Google Scholar] [CrossRef] [PubMed]
- Henry, E.R.; Cellmer, T.; Dunkelberger, E.B.; Metaferia, B.; Hofrichter, J.; Li, Q.; Ostrowski, D.; Ghirlando, R.; Louis, J.M.; Moutereau, S.; et al. Allosteric control of hemoglobin S fiber formation by oxygen and its relation to the pathophysiology of sickle cell disease. Proc. Natl. Acad. Sci. USA 2020. [Google Scholar] [CrossRef]
- Haynes, R.K.; Chan, W.C.; Lung, C.M.; Uhlemann, A.C.; Eckstein, U.; Taramelli, D.; Parapini, S.; Monti, D.; Krishna, S. The Fe2+-mediated decomposition, PfATP6 binding, and antimalarial activities of artemisone and other artemisinins: The unlikelihood of C-centered radicals as bioactive intermediates. ChemMedChem 2007, 2, 1480–1497. [Google Scholar] [CrossRef] [PubMed]
- Kapishnikov, S.; Grolimund, D.; Schneider, G.; Pereiro, E.; McNally, J.G.; Als-Nielsen, J.; Leiserowitz, L. Unraveling heme detoxification in the malaria parasite by in situ correlative X-ray fluorescence microscopy and soft X-ray tomography. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapishnikov, S.; Leiserowitz, L.; Yang, Y.; Cloetens, P.; Pereiro, E.; Ndonglack, F.A.; Matuschewski, K.; Als-Nielsen, J. Biochemistry of malaria parasite infected red blood cells by X-ray microscopy. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Tsamesidis, K.R.I.; Egwu, C.O.; Perio, P.; Augerau, J.; Benoit-Vical, F. An LC–MS Assay to Measure Superoxide Radicals and Hydrogen Peroxide in the Blood System. Metabolites 2020, 10, 175. [Google Scholar] [CrossRef]
- Zielonka, J.; Sikora, A.; Hardy, M.; Joseph, J.; Dranka, B.P.; Kalyanaraman, B. Boronate probes as diagnostic tools for real time monitoring of peroxynitrite and hydroperoxides. Chem. Res. Toxicol. 2012, 25, 1793–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielonka, J.; Vasquez-Vivar, J.; Kalyanaraman, B. Detection of 2-hydroxyethidium in cellular systems: A unique marker product of superoxide and hydroethidine. Nat. Protoc. 2008, 3, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Pabón, A.; Carmona, J.; Burgos, L.C.; Blair, S. Oxidative stress in patients with non-complicated malaria. Clin. Biochem. 2003, 36, 71–78. [Google Scholar] [CrossRef]
- Seixas, E.; Gozzelino, R.; Chora, Â.; Ferreira, A.; Silva, G.; Larsen, R.; Rebelo, S.; Penido, C.; Smith, N.R.; Coutinho, A.; et al. Heme oxygenase-1 affords protection against noncerebral forms of severe malaria. Proc. Natl. Acad. Sci. USA 2009, 106, 15837–15842. [Google Scholar] [CrossRef] [Green Version]
- Mannu, F.; Arese, P.; Cappellini, M.D.; Fiorelli, G.; Cappadoro, M.; Giribaldi, G.; Turrini, F. Role of hemichrome binding to erythrocyte membrane in the generation of band-3 alterations in beta-thalassemia intermedia erythrocytes. Blood 1995, 86, 2014–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Guo, Z.; Wang, H.B.; Zhou, J.J.; Zhang, W.J.; Chen, Q.W. Multifunctional mesoporous nanoparticles as pH-responsive Fe2+ reservoirs and artemisinin vehicles for synergistic inhibition of tumor growth. Biomaterials 2014, 35, 6498–6507. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Ortiz, M.P.; Wei, M.Q. Antitumor activity of artemisinin and its derivatives: From a well-known antimalarial agent to a potential anticancer drug. J. Biomed. Biotechnol. 2012, 2012, 247597. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.M.; Liu, Q.; Song, F.M.; Lin, X.X.; Su, Y.J.; Xu, J.; Huang, L.; Zong, S.H.; Zhao, J.M. Artesunate inhibits RANKL-induced osteoclastogenesis and bone resorption in vitro and prevents LPS-induced bone loss in vivo. J. Cell. Physiol. 2018, 233, 476–485. [Google Scholar] [CrossRef]
- Richardson, D.; Ponka, P.; Baker, E. The Effect of the Iron(III) Chelator, Desferoxamine, on Iron and Transferrin Uptake by the Human Malignant Melanoma Cell. Cancer Res. 1994, 54, 685–689. [Google Scholar]
- Wojtala, A.; Bonora, M.; Malinska, D.; Pinton, P.; Duszynski, J.; Wieckowski, M.R. Methods to monitor ROS production by fluorescence microscopy and fluorometry. Methods Enzymol. 2014, 542, 243–262. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Li, Y.; Chang, W.Q.; Zheng, J.Y.; Li, P.; Liu, L.F.; Xin, G.Z. Development and validation of a LC/MS-based method for the measurement of intracellular superoxide anion. Anal. Chim. Acta 2018. [Google Scholar] [CrossRef]
- Alvarez-Llamas, G.; De la Cuesta, F.; Barderas, M.G.; Zubiri, I.; Posada-Ayala, M.; Fernando, V. Characterization of membrane and cytosolic proteins of erythrocytes. Methods Mol. Biol. 2013. [Google Scholar] [CrossRef]
- Antonini, E.E.; Brunori, M. Hemoglobin and methemoglobin. In the Red Blood; Surgenor, D.M., Ed.; Academic Press: New York, NY, USA, 1975; pp. 753–797. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsamesidis, I.; Pério, P.; Pantaleo, A.; Reybier, K. Oxidation of Erythrocytes Enhance the Production of Reactive Species in the Presence of Artemisinins. Int. J. Mol. Sci. 2020, 21, 4799. https://doi.org/10.3390/ijms21134799
Tsamesidis I, Pério P, Pantaleo A, Reybier K. Oxidation of Erythrocytes Enhance the Production of Reactive Species in the Presence of Artemisinins. International Journal of Molecular Sciences. 2020; 21(13):4799. https://doi.org/10.3390/ijms21134799
Chicago/Turabian StyleTsamesidis, Ioannis, Pierre Pério, Antonella Pantaleo, and Karine Reybier. 2020. "Oxidation of Erythrocytes Enhance the Production of Reactive Species in the Presence of Artemisinins" International Journal of Molecular Sciences 21, no. 13: 4799. https://doi.org/10.3390/ijms21134799
APA StyleTsamesidis, I., Pério, P., Pantaleo, A., & Reybier, K. (2020). Oxidation of Erythrocytes Enhance the Production of Reactive Species in the Presence of Artemisinins. International Journal of Molecular Sciences, 21(13), 4799. https://doi.org/10.3390/ijms21134799