Resolving the Ionotropic P2X4 Receptor Mystery Points towards a New Therapeutic Target for Cardiovascular Diseases



Abstract
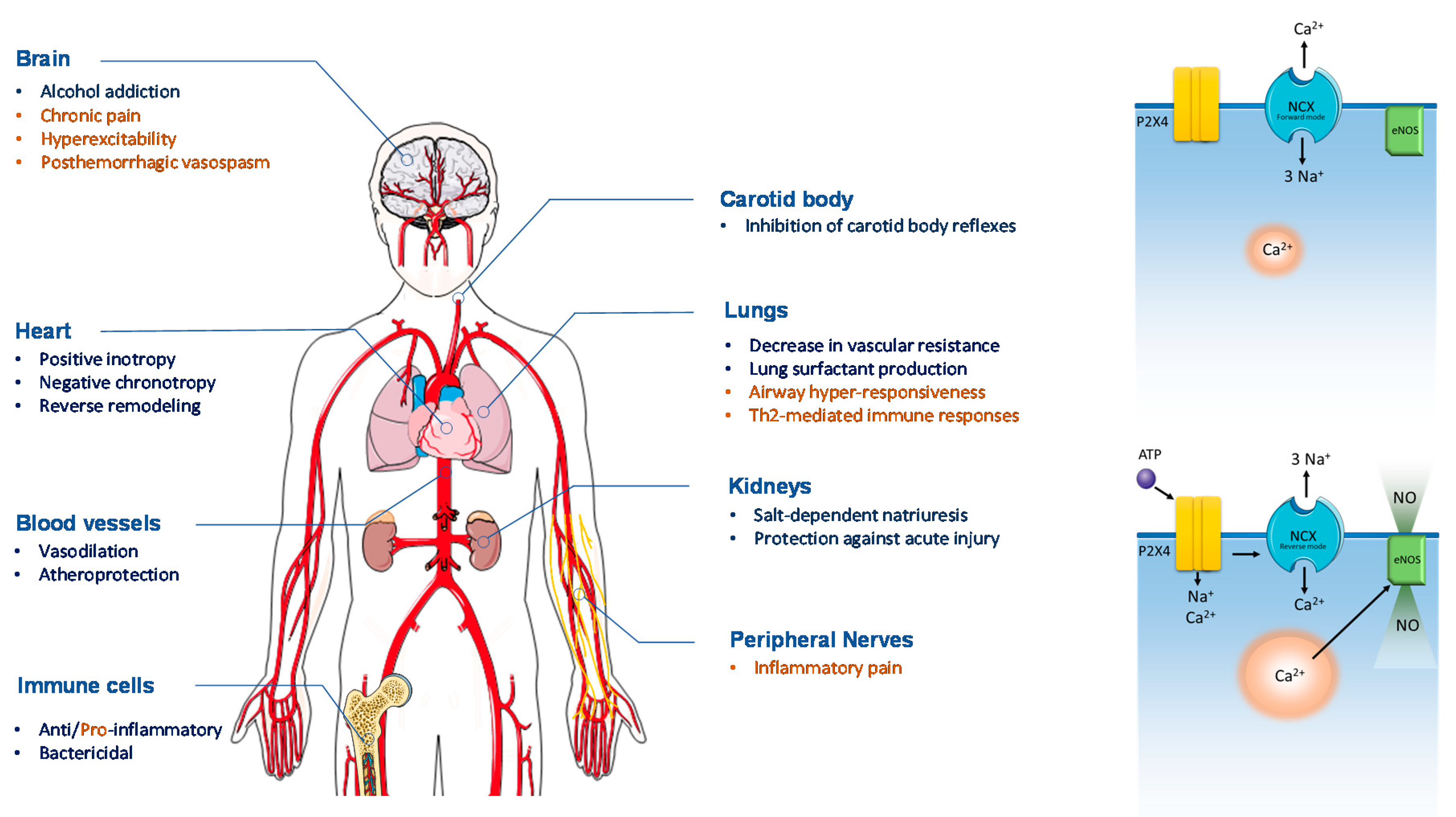
:1. The “Purinome”
2. The P2X4 Receptor: From Molecular Characteristics to Its Modes of Action
3. Benefits from the P2X4 Receptor Activation in the Heart
4. Cardiovascular Risk Factors: Is the P2X4 Receptor Friend or Foe?
5. Role Played by the P2X4 Receptor in Renal Function and Dysfunction
6. Is the P2X4 Receptor Activation Beneficial or Detrimental in the Lung?
7. Is there a Dark Side of P2X4 Receptor Activation Outside the Cardiovascular System?
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Burnstock, G. Purine and purinergic receptors. Brain Neurosci. Adv. 2018, 2, 2398212818817494. [Google Scholar] [CrossRef] [Green Version]
- Lazarowski, E.R. Vesicular and conductive mechanisms of nucleotide release. Purinergic Signal. 2012, 8, 359–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taruno, A. ATP Release Channels. Int. J. Mol. Sci. 2018, 19, 808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yegutkin, G.G. Enzymes involved in metabolism of extracellular nucleotides and nucleosides: Functional implications and measurement of activities. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 473–497. [Google Scholar] [CrossRef] [PubMed]
- Drury, A.N.; Szent-Gyorgyi, A. The physiological activity of adenine compounds with especial reference to their action upon the mammalian heart. J. Physiol. 1929, 68, 213–237. [Google Scholar] [CrossRef] [PubMed]
- Van der Meijden, P.E.J.; Heemskerk, J.W.M. Platelet biology and functions: New concepts and clinical perspectives. Nat. Rev. Cardiol. 2019, 16, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Roffi, M.; Patrono, C.; Collet, J.-P.; Mueller, C.; Valgimigli, M.; Andreotti, F.; Bax, J.J.; Borger, M.A.; Brotons, C.; Chew, D.P.; et al. 2015 ESC Guidelines for the management of acute coronary syndromes in patients presenting without persistent ST-segment elevation: Task Force for the Management of Acute Coronary Syndromes in Patients Presenting without Persistent ST-Segment Elevation of the European Society of Cardiology (ESC). Eur. Heart J. 2016, 37, 267–315. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic Signalling: Therapeutic Developments. Front. Pharmacol. 2017, 8, 661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suurvali, J.; Boudinot, P.; Kanellopoulos, J.; Ruutel Boudinot, S. P2X4: A fast and sensitive purinergic receptor. Biomed. J. 2017, 40, 245–256. [Google Scholar] [CrossRef]
- Kawate, T.; Michel, J.C.; Birdsong, W.T.; Gouaux, E. Crystal structure of the ATP-gated P2X(4) ion channel in the closed state. Nature 2009, 460, 592–598. [Google Scholar] [CrossRef]
- Sophocleous, R.A.; Berg, T.; Finol-Urdaneta, R.K.; Sluyter, V.; Keshiya, S.; Bell, L.; Curtis, S.J.; Curtis, B.L.; Seavers, A.; Bartlett, R.; et al. Pharmacological and genetic characterisation of the canine P2X4 receptor. Br. J. Pharmacol. 2020, 177, 2812–2829. [Google Scholar] [CrossRef] [PubMed]
- Stokes, L.; Scurrah, K.; Ellis, J.A.; Cromer, B.A.; Skarratt, K.K.; Gu, B.J.; Harrap, S.B.; Wiley, J.S. A loss-of-function polymorphism in the human P2X4 receptor is associated with increased pulse pressure. Hypertension 2011, 58, 1086–1092. [Google Scholar] [CrossRef] [Green Version]
- Saul, A.; Hausmann, R.; Kless, A.; Nicke, A. Heteromeric assembly of P2X subunits. Front. Cell Neurosci 2013, 7, 250. [Google Scholar] [CrossRef] [Green Version]
- Antonio, L.S.; Stewart, A.P.; Varanda, W.A.; Edwardson, J.M. Identification of P2X2/P2X4/P2X6 heterotrimeric receptors using atomic force microscopy (AFM) imaging. FEBS Lett. 2014, 588, 2125–2128. [Google Scholar] [CrossRef] [PubMed]
- Hattori, M.; Gouaux, E. Molecular mechanism of ATP binding and ion channel activation in P2X receptors. Nature 2012, 485, 207–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, K.A.; Jarvis, M.F.; Williams, M. Purine and pyrimidine (P2) receptors as drug targets. J. Med. Chem. 2002, 45, 4057–4093. [Google Scholar] [CrossRef] [PubMed]
- Kasuya, G.; Fujiwara, Y.; Tsukamoto, H.; Morinaga, S.; Ryu, S.; Touhara, K.; Ishitani, R.; Furutani, Y.; Hattori, M.; Nureki, O. Structural insights into the nucleotide base specificity of P2X receptors. Sci. Rep. 2017, 7, 45208. [Google Scholar] [CrossRef] [Green Version]
- Egan, T.M.; Samways, D.S.; Li, Z. Biophysics of P2X receptors. Pflugers Arch. 2006, 452, 501–512. [Google Scholar] [CrossRef]
- Samways, D.S.; Li, Z.; Egan, T.M. Principles and properties of ion flow in P2X receptors. Front. Cell Neurosci. 2014, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Coddou, C.; Yan, Z.; Obsil, T.; Huidobro-Toro, J.P.; Stojilkovic, S.S. Activation and regulation of purinergic P2X receptor channels. Pharmacol. Rev. 2011, 63, 641–683. [Google Scholar] [CrossRef] [Green Version]
- Soto, F.; Garcia-Guzman, M.; Gomez-Hernandez, J.M.; Hollmann, M.; Karschin, C.; Stuhmer, W. P2X4: An ATP-activated ionotropic receptor cloned from rat brain. Proc. Natl. Acad. Sci. USA 1996, 93, 3684–3688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khakh, B.S.; Bao, X.R.; Labarca, C.; Lester, H.A. Neuronal P2X transmitter-gated cation channels change their ion selectivity in seconds. Nat. Neurosci. 1999, 2, 322–330. [Google Scholar] [CrossRef]
- Bernier, L.P.; Ase, A.R.; Boue-Grabot, E.; Seguela, P. P2X4 receptor channels form large noncytolytic pores in resting and activated microglia. Glia 2012, 60, 728–737. [Google Scholar] [CrossRef]
- Shinozaki, Y.; Sumitomo, K.; Tsuda, M.; Koizumi, S.; Inoue, K.; Torimitsu, K. Direct observation of ATP-induced conformational changes in single P2X(4) receptors. PLoS Biol. 2009, 7, e1000103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bo, X.; Zhang, Y.; Nassar, M.; Burnstock, G.; Schoepfer, R. A P2X purinoceptor cDNA conferring a novel pharmacological profile. FEBS Lett. 1995, 375, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.A.; Chessell, I.P.; Simon, J.; Barnard, E.A.; Miller, K.J.; Michel, A.D.; Humphrey, P.P. Functional characterization of the P2X4 receptor orthologues. Br. J. Pharmacol. 2000, 129, 388–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelrahman, A.; Namasivayam, V.; Hinz, S.; Schiedel, A.C.; Köse, M.; Burton, M.; El-Tayeb, A.; Gillard, M.; Bajorath, J.; De Ryck, M.; et al. Characterization of P2X4 receptor agonists and antagonists by calcium influx and radioligand binding studies. Biochem. Pharmacol. 2017, 125, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, B.R.; Lynch, K.J.; Touma, E.; Niforatos, W.; Burgard, E.C.; Alexander, K.M.; Park, H.S.; Yu, H.; Metzger, R.; Kowaluk, E.; et al. Pharmacological characterization of recombinant human and rat P2X receptor subtypes. Eur. J. Pharmacol. 1999, 376, 127–138. [Google Scholar] [CrossRef]
- He, M.-L.; Gonzalez-Iglesias, A.E.; Stojilkovic, S.S. Role of Nucleotide P2 Receptors in Calcium Signaling and Prolactin Release in Pituitary Lactotrophs. J. Boil. Chem. 2003, 278, 46270–46277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusuf, S.; Bosch, J.; Dagenais, G.; Zhu, J.; Xavier, D.; Liu, L.; Pais, P.; Lopez-Jaramillo, P.; Leiter, L.A.; Dans, A.; et al. Cholesterol Lowering in Intermediate-Risk Persons without Cardiovascular Disease. N. Engl. J. Med. 2016, 374, 2021–2031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browne, L.E.; North, R.A. P2X receptor intermediate activation states have altered nucleotide selectivity. J. Neurosci. 2013, 33, 14801–14808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, J.A.; Evans, R.J. Atp Binding at Human P2x1 Receptors: Contribution of Aromatic and Basic Amino Acids Revealed Using Mutagenesis and Partial Agonists. J. Biol. Chem. 2004, 279, 9043–9055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.B.; Cronin, C.; Sonin, D.; Joshi, B.V.; Gongora Nieto, M.; Harrison, D.; Jacobson, K.A.; Liang, B.T. P2X purinergic receptor-mediated ionic current in cardiac myocytes of calsequestrin model of cardiomyopathy: Implications for the treatment of heart failure. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1077–H1084. [Google Scholar] [CrossRef]
- Kumar, T.S.; Yang, T.; Mishra, S.; Cronin, C.; Chakraborty, S.; Shen, J.B.; Liang, B.T.; Jacobson, K.A. 5′-Phosphate and 5′-phosphonate ester derivatives of (N)-methanocarba adenosine with in vivo cardioprotective activity. J. Med. Chem. 2013, 56, 902–914. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Olmos, V.; Abdelrahman, A.; El-Tayeb, A.; Freudendahl, D.; Weinhausen, S.; Müller, C.E. N-Substituted Phenoxazine and Acridone Derivatives: Structure–Activity Relationships of Potent P2X4 Receptor Antagonists. J. Med. Chem. 2012, 55, 9576–9588. [Google Scholar] [CrossRef]
- Balázs, B.; Danko, T.; Kovacs, G.; Köles, L.; Hediger, M.A. Zsembery, Ákos Investigation of the Inhibitory Effects of the Benzodiazepine Derivative, 5-BDBD on P2X4Purinergic Receptors by two Complementary Methods. Cell. Physiol. Biochem. 2013, 32, 11–24. [Google Scholar] [CrossRef]
- Bo, X.; Liu, M.; Schoepfer, R.; Burnstock, G. Characterization and expression of ATP P2X4 receptor from embryonic chick skeletal muscle. Drug Dev. Res. 2001, 53, 22–28. [Google Scholar] [CrossRef]
- Virginio, C.; Robertson, G.; Surprenant, A.; North, R.A. Trinitrophenyl-Substituted Nucleotides Are Potent Antagonists Selective for P2x1, P2x3, and Heteromeric P2x2/3 Receptors. Mol. Pharmacol. 1998, 53, 969–973. [Google Scholar] [PubMed]
- Tomioka, A.; Ueno, S.; Kohama, K.; Goto, F.; Inoue, K. Propofol potentiates ATP-activated currents of recombinant P2X4 receptor channels expressed in human embryonic kidney 293 cells. Neurosci. Lett. 2000, 284, 167–170. [Google Scholar] [CrossRef]
- Coddou, C.; Sandoval, R.; Hevia, M.J.; Stojilkovic, S.S. Characterization of the antagonist actions of 5-BDBD at the rat P2X4 receptor. Neurosci. Lett. 2019, 690, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Ase, A.R.; Honson, N.S.; Zaghdane, H.; Pfeifer, T.A.; Seguela, P. Identification and characterization of a selective allosteric antagonist of human P2X4 receptor channels. Mol. Pharmacol. 2015, 87, 606–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, K.; Tajima, M.; Nagano, T.; Obayashi, K.; Ito, M.; Yamamoto, K.; Matsuoka, I. Co-Stimulation of Purinergic P2X4 and Prostanoid EP3 Receptors Triggers Synergistic Degranulation in Murine Mast Cells. Int. J. Mol. Sci. 2019, 20, 5157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, M.; Abdelrahman, A.; Weinhausen, S.; Hinz, S.; Weyer, S.; Dosa, S.; El-Tayeb, A.; Müller, C.E. Carbamazepine derivatives with P2X4 receptor-blocking activity. Bioorganic Med. Chem. 2014, 22, 1077–1088. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Prudic, K.; Pippel, A.; Klapperstück, M.; Braam, U.; Müller, C.E.; Schmalzing, G.; Markwardt, F. Interaction of Purinergic P2x4 and P2x7 Receptor Subunits. Front. Pharmacol. 2017, 8, 860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, Y.; Yamashita, T.; Sasaki, A.; Nakata, E.; Kohno, K.; Masuda, T.; Tozaki-Saitoh, H.; Imai, T.; Kuraishi, Y.; Tsuda, M.; et al. A novel P2X4 receptor-selective antagonist produces anti-allodynic effect in a mouse model of herpetic pain. Sci. Rep. 2016, 6, 32461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, K.; Tsuda, M. Microglia in neuropathic pain: cellular and molecular mechanisms and therapeutic potential. Nat. Rev. Neurosci. 2018, 19, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Beswick, P.; Wahab, B.; Honey, M.A.; Paradowski, M.; Jiang, K.; Lochner, M.; Murrell-Lagnado, R.D.; Thompson, A.J. A challenge finding P2X1 and P2X4 ligands. Neuropharmacology 2019, 157, 107674. [Google Scholar] [CrossRef] [PubMed]
- Nagata, K.; Imai, T.; Yamashita, T.; Tsuda, M.; Tozaki-Saitoh, H.; Inoue, K. Antidepressants inhibit P2X4 receptor function: a possible involvement in neuropathic pain relief. Mol. Pain 2009, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T.; Iwamoto, S.; Yoshinaga, R.; Tozaki-Saitoh, H.; Nishiyama, A.; Mak, T.W.; Tamura, T.; Tsuda, M.; Inoue, K. Transcription factor IRF5 drives P2X4R+-reactive microglia gating neuropathic pain. Nat. Commun. 2014, 5, 3771. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, T.; Yamamoto, S.; Zhang, J.; Kometani, M.; Tomiyama, D.; Kohno, K.; Tozaki-Saitoh, H.; Inoue, K.; Tsuda, M. Duloxetine Inhibits Microglial P2X4 Receptor Function and Alleviates Neuropathic Pain after Peripheral Nerve Injury. PLoS ONE 2016, 11, e0165189. [Google Scholar] [CrossRef] [Green Version]
- Werner, S.; Mesch, S.; Hillig, R.C.; Laak, A.T.; Klint, J.; Neagoe, I.; Laux-Biehlmann, A.; Dahllof, H.; Brauer, N.; Puetter, V.; et al. Faculty Opinions recommendation of Discovery and Characterization of the Potent and Selective P2X4 Inhibitor N-[4-(3-Chlorophenoxy)-3-sulfamoylphenyl]-2-phenylacetamide (BAY-1797) and Structure-Guided Amelioration of Its CYP3A4 Induction Profile. Fac. Opin. Post Publ. Peer Rev. Biomed. Lit. 2019, 62, 11194–11217. [Google Scholar] [CrossRef]
- Williams, W.A.; Linley, J.E.; Jones, C.A.; Shibata, Y.; Snijder, A.; Button, J.; Hatcher, J.P.; Huang, L.; Taddese, B.; Thornton, P.; et al. Antibodies binding the head domain of P2X4 inhibit channel function and reverse neuropathic pain. Pain 2019, 160, 1989–2003. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.B.; Toti, K.S.; Chakraborty, S.; Kumar, T.S.; Cronin, C.; Liang, B.T.; Jacobson, K.A. Prevention and rescue of cardiac dysfunction by methanocarba adenosine monophosphonate derivatives. Purinergic Signal. 2020, 16, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Norenberg, W.; Sobottka, H.; Hempel, C.; Plötz, T.; Fischer, W.; Schmalzing, G.; Schaefer, M. Positive allosteric modulation by ivermectin of human but not murine P2X7 receptors. Br. J. Pharmacol. 2012, 167, 48–66. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.J.; Michel, A.D.; Chessell, I.; Humphrey, P. Cibacron blue allosterically modulates the rat P2X4 receptor. Neuropharmacol. 1998, 37, 1579–1586. [Google Scholar] [CrossRef]
- Dhuna, K.; Felgate, M.; Bidula, S.; Walpole, S.; Bibic, L.; Cromer, B.A.; Angulo, J.; Sanderson, J.; Stebbing, M.J.; Stokes, L. Ginsenosides Act As Positive Modulators of P2X4 Receptors. Mol. Pharmacol. 2018, 95, 210–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivcev, S.; Slavikova, B.; Rupert, M.; Ivetic, M.; Nekardova, M.; Kudova, E.; Zemkova, H. Synthetic testosterone derivatives modulate rat P2X2 and P2X4 receptor channel gating. J. Neurochem. 2019, 150, 28–43. [Google Scholar] [CrossRef] [PubMed]
- Coddou, C.; Lorca, R.; Acuña-Castillo, C.; Grauso, M.; Rassendren, F.; Huidobro-Toro, J. Heavy metals modulate the activity of the purinergic P2X4 receptor. Toxicol. Appl. Pharmacol. 2005, 202, 121–131. [Google Scholar] [CrossRef]
- Codocedo, J.F.; Rodríguez, F.E.; Huidobro-Toro, J.P. Neurosteroids differentially modulate P2X4ATP-gated channels through non-genomic interactions. J. Neurochem. 2009, 110, 734–744. [Google Scholar] [CrossRef]
- Popova, M.; Asatryan, L.; Ostrovskaya, O.; Wyatt, L.; Li, K.; Alkana, R.L.; Davies, D.L. A point mutation in the ectodomain-transmembrane 2 interface eliminates the inhibitory effects of ethanol in P2X4 receptors. J. Neurochem. 2009, 112, 307–317. [Google Scholar] [CrossRef] [Green Version]
- Davies, D.L.; Kochegarov, A.A.; Kuo, S.T.; Kulkarni, A.A.; Woodward, J.J.; King, B.F.; Alkana, R.L. Ethanol differentially affects ATP-gated P2X and P2X receptor subtypes expressed in oocytes. Neuropharmacol. 2005, 49, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Stoop, R.; Surprenant, A.; North, R.A. Different sensitivities to pH of ATP-induced currents at four cloned P2X receptors. J. Neurophysiol. 1997, 78, 1837–1840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Fountain, S.J. Fluvastatin suppresses native and recombinant human P2X4 receptor function. Purinergic Signal. 2012, 8, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilyaskin, A.V.; Sure, F.; Nesterov, V.; Haerteis, S.; Korbmacher, C. Bile acids inhibit human purinergic receptor P2X4 in a heterologous expression system. J. Gen. Physiol. 2019, 151, 820–833. [Google Scholar] [CrossRef] [Green Version]
- Schmid, R.; Evans, R.J. ATP-Gated P2X Receptor Channels: Molecular Insights into Functional Roles. Annu. Rev. Physiol. 2019, 81, 43–62. [Google Scholar] [CrossRef]
- Garcia-Guzman, M.; Soto, F.; Gomez-Hernandez, J.M.; Lund, P.E.; Stuhmer, W. Characterization of recombinant human P2X4 receptor reveals pharmacological differences to the rat homologue. Mol. Pharmacol. 1997, 51, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Coddou, C.; Stojilkovic, S.S.; Huidobro-Toro, J.P. Allosteric modulation of ATP-gated P2X receptor channels. Rev. Neurosci. 2011, 22, 335–354. [Google Scholar] [CrossRef] [Green Version]
- Kasuya, G.; Fujiwara, Y.; Takemoto, M.; Dohmae, N.; Nakada-Nakura, Y.; Ishitani, R.; Hattori, M.; Nureki, O. Structural Insights into Divalent Cation Modulations of ATP-Gated P2X Receptor Channels. Cell Rep. 2016, 14, 932–944. [Google Scholar] [CrossRef] [Green Version]
- Clarke, C.E.; Benham, C.D.; Bridges, A.; George, A.R.; Meadows, H.J. Mutation of histidine 286 of the human P2X4 purinoceptor removes extracellular pH sensitivity. J. Physiol. 2000, 523, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Khakh, B.S.; Proctor, W.R.; Dunwiddie, T.V.; Labarca, C.; Lester, H.A. Allosteric control of gating and kinetics at P2X(4) receptor channels. J. Neurosci. 1999, 19, 7289–7299. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.A.; Yule, D.I. Protein kinase A regulation of P2X(4) receptors: Requirement for a specific motif in the C-terminus. Biochim. Biophys. Acta 2010, 1803, 275–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernier, L.P.; Ase, A.R.; Chevallier, S.; Blais, D.; Zhao, Q.; Boue-Grabot, E.; Logothetis, D.; Seguela, P. Phosphoinositides regulate P2X4 ATP-gated channels through direct interactions. J. Neurosci. 2008, 28, 12938–12945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohata, Y.; Ogata, S.; Nakanishi, K.; Kanazawa, F.; Uenoyama, M.; Hiroi, S.; Tominaga, S.; Kawai, T. Expression of P2X4R mRNA and protein in rats with hypobaric hypoxia-induced pulmonary hypertension. Circ. J. 2011, 75, 945–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Wang, L.; Li, J.W.; Gong, M.; He, L.; Feng, R.; Dai, Z.; Li, S.Q. Hypoxia induced amoeboid microglial cell activation in postnatal rat brain is mediated by ATP receptor P2X4. BMC Neurosci. 2011, 12, 111. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Matsuoka, I.; Ono, T.; Inoue, K.; Kimura, J. Selective up-regulation of P2X4-receptor gene expression by interferon-gamma in vascular endothelial cells. J. Pharmacol. Sci. 2008, 107, 419–427. [Google Scholar] [CrossRef]
- Gu, B.J.; Sun, C.; Valova, V.A.; Skarratt, K.K.; Wiley, J.S. Identification of the promoter region of the P2RX4 gene. Mol. Biol Rep. 2010, 37, 3369–3376. [Google Scholar] [CrossRef]
- Fountain, S.J.; North, R.A. A C-terminal lysine that controls human P2X4 receptor desensitization. J. Biol. Chem. 2006, 281, 15044–15049. [Google Scholar] [CrossRef] [Green Version]
- Murrell-Lagnado, R.D. A role for P2X4 receptors in lysosome function. J. Gen. Physiol. 2018, 150, 185–187. [Google Scholar] [CrossRef]
- Xu, J.; Chai, H.; Ehinger, K.; Egan, T.M.; Srinivasan, R.; Frick, M.; Khakh, B.S. Imaging P2X4 receptor subcellular distribution, trafficking, and regulation using P2X4-pHluorin. J. Gen. Physiol. 2014, 144, 81–104. [Google Scholar] [CrossRef]
- Erlinge, D.; Burnstock, G. P2 receptors in cardiovascular regulation and disease. Purinergic Signal. 2008, 4, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G. Purinergic Signaling in the Cardiovascular System. Circ. Res. 2017, 120, 207–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnstock, G.; Pelleg, A. Cardiac purinergic signalling in health and disease. Purinergic Signal. 2015, 11, 1–46. [Google Scholar] [CrossRef]
- Musa, H.; Tellez, J.O.; Chandler, N.J.; Greener, I.D.; Maczewski, M.; Mackiewicz, U.; Beresewicz, A.; Molenaar, P.; Boyett, M.R.; Dobrzynski, H. P2 purinergic receptor mRNA in rat and human sinoatrial node and other heart regions. Naunyn Schmiedebergs Arch. Pharmacol. 2009, 379, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Bogdanov, Y.; Rubino, A.; Burnstock, G. Characterisation of subtypes of the P2X and P2Y families of ATP receptors in the foetal human heart. Life Sci. 1998, 62, 697–703. [Google Scholar] [CrossRef]
- Braganca, B.; Nogueira-Marques, S.; Ferreirinha, F.; Fontes-Sousa, A.P.; Correia-de-Sa, P. The Ionotropic P2X4 Receptor has Unique Properties in the Heart by Mediating the Negative Chronotropic Effect of ATP While Increasing the Ventricular Inotropy. Front. Pharmacol. 2019, 10, 1103. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Ruan, H.Z.; Burnstock, G.; Dunn, P.M. Differential expression of P2X receptors on neurons from different parasympathetic ganglia. Neuropharmacology 2005, 48, 766–777. [Google Scholar] [CrossRef]
- Chen, J.-B.; Liu, W.-J.; Che, H.; Liu, J.; Sun, H.-Y.; Li, G.-R. Adenosine-5′-triphosphate up-regulates proliferation of human cardiac fibroblasts. Br. J. Pharmacol. 2012, 166, 1140–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Mei, Q.B.; Yao, X.J.; Smith, E.; Barry, W.H.; Liang, B.T. A novel contractile phenotype with cardiac transgenic expression of the human P2X4 receptor. FASEB J. 2001, 15, 2739–2741. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.; Sonin, D.; Jones, L.; Barry, W.H.; Liang, B.T. A beneficial role of cardiac P2X4 receptors in heart failure: Rescue of the calsequestrin overexpression model of cardiomyopathy. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H1096–H1103. [Google Scholar] [CrossRef]
- Sonin, D.; Zhou, S.Y.; Cronin, C.; Sonina, T.; Wu, J.; Jacobson, K.A.; Pappano, A.; Liang, B.T. Role of P2X purinergic receptors in the rescue of ischemic heart failure. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1191–H1197. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.B.; Shutt, R.; Agosto, M.; Pappano, A.; Liang, B.T. Reversal of cardiac myocyte dysfunction as a unique mechanism of rescue by P2X4 receptors in cardiomyopathy. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1089–H1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Shen, J.B.; Yang, R.; Redden, J.; Dodge-Kafka, K.; Grady, J.; Jacobson, K.A.; Liang, B.T. Novel protective role of endogenous cardiac myocyte P2X4 receptors in heart failure. Circ. Heart Fail. 2014, 7, 510–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisner, D.A.; Caldwell, J.L.; Kistamas, K.; Trafford, A.W. Calcium and Excitation-Contraction Coupling in the Heart. Circ. Res. 2017, 121, 181–195. [Google Scholar] [CrossRef]
- Shen, J.B.; Yang, R.; Pappano, A.; Liang, B.T. Cardiac P2X purinergic receptors as a new pathway for increasing Na(+) entry in cardiac myocytes. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H1469–H1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakatta, E.G.; Maltsev, V.A.; Vinogradova, T.M. A coupled SYSTEM of intracellular Ca2+ clocks and surface membrane voltage clocks controls the timekeeping mechanism of the heart’s pacemaker. Circ. Res. 2010, 106, 659–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belz, G.G.; Breithaupt-Grogler, K.; Osowski, U. Treatment of congestive heart failure--current status of use of digitoxin. Eur. J. Clin. Investig. 2001, 31, 10–17. [Google Scholar] [CrossRef]
- Whayne, T.F., Jr. Clinical Use of Digitalis: A State of the Art Review. Am. J. Cardiovasc. Drugs 2018, 18, 427–440. [Google Scholar] [CrossRef]
- Kirchhof, P.; Benussi, S.; Kotecha, D.; Ahlsson, A.; Atar, D.; Casadei, B.; Castella, M.; Diener, H.C.; Heidbuchel, H.; Hendriks, J.; et al. 2016 ESC Guidelines for the management of atrial fibrillation developed in collaboration with EACTS. Eur. Heart J. 2016, 37, 2893–2962. [Google Scholar] [CrossRef] [Green Version]
- Ponikowski, P.; Voors, A.A.; Anker, S.D.; Bueno, H.; Cleland, J.G.F.; Coats, A.J.S.; Falk, V.; Gonzalez-Juanatey, J.R.; Harjola, V.P.; Jankowska, E.A.; et al. 2016 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure: The Task Force for the diagnosis and treatment of acute and chronic heart failure of the European Society of Cardiology (ESC)Developed with the special contribution of the Heart Failure Association (HFA) of the ESC. Eur. Heart J. 2016, 37, 2129–2200. [Google Scholar] [CrossRef]
- Bossu, A.; Kostense, A.; Beekman, H.D.M.; Houtman, M.J.C.; van der Heyden, M.A.G.; Vos, M.A. Istaroxime, a positive inotropic agent devoid of proarrhythmic properties in sensitive chronic atrioventricular block dogs. Pharmacol. Res. 2018, 133, 132–140. [Google Scholar] [CrossRef]
- Carubelli, V.; Zhang, Y.; Metra, M.; Lombardi, C.; Felker, G.M.; Filippatos, G.; O’Connor, C.M.; Teerlink, J.R.; Simmons, P.; Segal, R.; et al. Treatment with 24 hour istaroxime infusion in patients hospitalised for acute heart failure: A randomised, placebo-controlled trial. Eur. J. Heart Fail. 2020. [Google Scholar] [CrossRef] [PubMed]
- Chioncel, O.; Collins, S.P.; Butler, J. Istaroxime in acute heart failure: The holy grail is at HORIZON? Eur. J. Heart Fail. 2020. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.J.; Blair, J.E.; Filippatos, G.S.; Macarie, C.; Ruzyllo, W.; Korewicki, J.; Bubenek-Turconi, S.I.; Ceracchi, M.; Bianchetti, M.; Carminati, P.; et al. Effects of istaroxime on diastolic stiffness in acute heart failure syndromes: Results from the Hemodynamic, Echocardiographic, and Neurohormonal Effects of Istaroxime, a Novel Intravenous Inotropic and Lusitropic Agent: A Randomized Controlled Trial in Patients Hospitalized with Heart Failure (HORIZON-HF) trial. Am. Heart J. 2009, 157, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Eisner, D.A.; Caldwell, J.L.; Trafford, A.W.; Hutchings, D.C. The Control of Diastolic Calcium in the Heart: Basic Mechanisms and Functional Implications. Circ. Res. 2020, 126, 395–412. [Google Scholar] [CrossRef]
- Belhassen, B.; Pelleg, A.; Shoshani, D.; Laniado, S. Atrial fibrillation induced by adenosine triphosphate. Am. J. Cardiol. 1984, 53, 1405–1406. [Google Scholar] [CrossRef]
- Tao, S.; Yamauchi, Y.; Maeda, S.; Okada, H.; Yamaguchi, T.; Hara, N.; Konishi, Y.; Umemoto, T.; Miyamoto, T.; Obayashi, T.; et al. Adenosine triphosphate-induced atrial fibrillation: The clinical significance and relevance to spontaneous atrial fibrillation. J. Interv. Card. Electrophysiol. 2014, 39, 103–109. [Google Scholar] [CrossRef]
- Gilbert, G.; Demydenko, K.; Dries, E.; Puertas, R.D.; Jin, X.; Sipido, K.; Roderick, H.L. Calcium Signaling in Cardiomyocyte Function. Cold Spring Harb. Perspect. Biol. 2020, 12, 12. [Google Scholar] [CrossRef]
- Farah, C.; Michel, L.Y.M.; Balligand, J.L. Nitric oxide signalling in cardiovascular health and disease. Nat. Rev. Cardiol. 2018, 15, 292–316. [Google Scholar] [CrossRef]
- Vasan, R.S.; Sullivan, L.M.; Wilson, P.W.; Sempos, C.T.; Sundstrom, J.; Kannel, W.B.; Levy, D.; D’Agostino, R.B. Relative importance of borderline and elevated levels of coronary heart disease risk factors. Ann. Intern. Med. 2005, 142, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Group, S.R.; Wright, J.T., Jr.; Williamson, J.D.; Whelton, P.K.; Snyder, J.K.; Sink, K.M.; Rocco, M.V.; Reboussin, D.M.; Rahman, M.; Oparil, S.; et al. A Randomized Trial of Intensive versus Standard Blood-Pressure Control. N. Engl. J. Med. 2015, 373, 2103–2116. [Google Scholar] [CrossRef]
- Rawshani, A.; Rawshani, A.; Franzén, S.; Sattar, N.; Eliasson, B.; Svensson, A.-M.; Zethelius, B.; Miftaraj, M.; McGuire, D.K.; Rosengren, A.; et al. Risk Factors, Mortality, and Cardiovascular Outcomes in Patients with Type 2 Diabetes. N. Engl. J. Med. 2018, 379, 633–644. [Google Scholar] [CrossRef]
- Dzau, V.J.; Antman, E.M.; Black, H.R.; Hayes, D.L.; Manson, J.E.; Plutzky, J.; Popma, J.J.; Stevenson, W. The Cardiovascular Disease Continuum Validated: Clinical Evidence of Improved Patient Outcomes. Circulation 2006, 114, 2850–2870. [Google Scholar] [CrossRef]
- Burnstock, G.; Novak, I. Purinergic signalling and diabetes. Purinergic Signal. 2013, 9, 307–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparks, D.L.; Chatterjee, C. Purinergic signaling, dyslipidemia and inflammatory disease. Cell Physiol. Biochem. 2012, 30, 1333–1339. [Google Scholar] [CrossRef] [PubMed]
- Novak, I.; Solini, A. P2X receptor-ion channels in the inflammatory response in adipose tissue and pancreas-potential triggers in onset of type 2 diabetes? Curr. Opin. Immunol. 2018, 52, 1–7. [Google Scholar] [CrossRef]
- Ren, L.M.; Nakane, T.; Chiba, S. Muscarinic receptor subtypes mediating vasodilation and vasoconstriction in isolated, perfused simian coronary arteries. J. Cardiovasc. Pharmacol. 1993, 22, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Brock, J.A.; Van Helden, D.F. Enhanced excitatory junction potentials in mesenteric arteries from spontaneously hypertensive rats. Pflugers Arch. 1995, 430, 901–908. [Google Scholar] [CrossRef]
- Vial, C.; Evans, R.J. P2X(1) receptor-deficient mice establish the native P2X receptor and a P2Y6-like receptor in arteries. Mol. Pharmacol 2002, 62, 1438–1445. [Google Scholar] [CrossRef]
- Lewis, C.J.; Evans, R.J. P2X receptor immunoreactivity in different arteries from the femoral, pulmonary, cerebral, coronary and renal circulations. J. Vasc. Res. 2001, 38, 332–340. [Google Scholar] [CrossRef]
- Harhun, M.I.; Povstyan, O.V.; Albert, A.P.; Nichols, C.M. ATP-evoked sustained vasoconstrictions mediated by heteromeric P2X1/4 receptors in cerebral arteries. Stroke 2014, 45, 2444–2450. [Google Scholar] [CrossRef] [Green Version]
- Milner, P.; Bodin, P.; Loesch, A.; Burnstock, G. Rapid release of endothelin and ATP from isolated aortic endothelial cells exposed to increased flow. Biochem. Biophys. Res. Commun. 1990, 170, 649–656. [Google Scholar] [CrossRef]
- Yamamoto, K.; Sokabe, T.; Matsumoto, T.; Yoshimura, K.; Shibata, M.; Ohura, N.; Fukuda, T.; Sato, T.; Sekine, K.; Kato, S.; et al. Impaired flow-dependent control of vascular tone and remodeling in P2X4-deficient mice. Nat. Med. 2006, 12, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Piskuric, N.A.; Nurse, C.A. Expanding role of ATP as a versatile messenger at carotid and aortic body chemoreceptors. J. Physiol. 2013, 591, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Campanucci, V.A.; Zhang, M.; Vollmer, C.; Nurse, C.A. Expression of multiple P2X receptors by glossopharyngeal neurons projecting to rat carotid body O2-chemoreceptors: Role in nitric oxide-mediated efferent inhibition. J. Neurosci. 2006, 26, 9482–9493. [Google Scholar] [CrossRef] [PubMed]
- Pijacka, W.; Moraes, D.J.; Ratcliffe, L.E.; Nightingale, A.K.; Hart, E.C.; da Silva, M.P.; Machado, B.H.; McBryde, F.D.; Abdala, A.P.; Ford, A.P.; et al. Purinergic receptors in the carotid body as a new drug target for controlling hypertension. Nat. Med. 2016, 22, 1151–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conde, S.V.; Monteiro, E.C.; Sacramento, J.F. Purines and Carotid Body: New Roles in Pathological Conditions. Front. Pharmacol. 2017, 8, 913. [Google Scholar] [CrossRef] [Green Version]
- Libby, P.; Buring, J.E.; Badimon, L.; Hansson, G.K.; Deanfield, J.; Bittencourt, M.S.; Tokgozoglu, L.; Lewis, E.F. Atherosclerosis. Nat. Rev. Dis. Primers 2019, 5, 56. [Google Scholar] [CrossRef]
- Stachon, P.; Hergeth, S.H.; Anto Michel, N.M.; Dufner, B.D.; Rodriguez, A.R.; Hoppe, N.H.; Wolf, D.W.; Bode, C.B.; Idzko, M.I.; Zirlik, A.Z. Extracellular atp contributes to atherogenesis via purinergic receptors by inducing leukocyte recruitment in mice. Eur. Hear. J. 2013, 34. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.V.; Li, Y.; Liu, X.; Xia, S.; Shi, P.; Li, L.; Chen, Z.; Yin, C.; Eriguchi, M.; Chen, Y.; et al. ATP release drives heightened immune responses associated with hypertension. Sci. Immunol. 2019, 4, eaau6426. [Google Scholar] [CrossRef]
- Sathanoori, R.; Sward, K.; Olde, B.; Erlinge, D. The ATP Receptors P2X7 and P2X4 Modulate High Glucose and Palmitate-Induced Inflammatory Responses in Endothelial Cells. PLoS ONE 2015, 10, e0125111. [Google Scholar] [CrossRef] [Green Version]
- Pulvirenti, T.J.; Yin, J.L.; Chaufour, X.; McLachlan, C.; Hambly, B.D.; Bennett, M.R.; Barden, J.A. P2X (purinergic) receptor redistribution in rabbit aorta following injury to endothelial cells and cholesterol feeding. J. Neurocytol. 2000, 29, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Layhadi, J.A.; Turner, J.; Crossman, D.; Fountain, S.J. ATP Evokes Ca(2+) Responses and CXCL5 Secretion via P2X4 Receptor Activation in Human Monocyte-Derived Macrophages. J. Immunol. 2018, 200, 1159–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousselle, A.; Qadri, F.; Leukel, L.; Yilmaz, R.; Fontaine, J.F.; Sihn, G.; Bader, M.; Ahluwalia, A.; Duchene, J. CXCL5 limits macrophage foam cell formation in atherosclerosis. J. Clin. Investig. 2013, 123, 1343–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathanoori, R.; Rosi, F.; Gu, B.J.; Wiley, J.S.; Muller, C.E.; Olde, B.; Erlinge, D. Shear stress modulates endothelial KLF2 through activation of P2X4. Purinergic Signal. 2015, 11, 139–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozaki, T.; Muramatsu, R.; Sasai, M.; Yamamoto, M.; Kubota, Y.; Fujinaka, T.; Yoshimine, T.; Yamashita, T. The P2X4 receptor is required for neuroprotection via ischemic preconditioning. Sci. Rep. 2016, 6, 25893. [Google Scholar] [CrossRef] [Green Version]
- Anavekar, N.S.; McMurray, J.J.V.; Velazquez, E.J.; Solomon, S.D.; Kober, L.; Rouleau, J.-L.; White, H.D.; Nordlander, R.; Maggioni, A.; Dickstein, K.; et al. Relation between Renal Dysfunction and Cardiovascular Outcomes after Myocardial Infarction. N. Engl. J. Med. 2004, 351, 1285–1295. [Google Scholar] [CrossRef]
- McAlister, F.A.; Ezekowitz, J.; Tarantini, L.; Squire, I.; Komajda, M.; Bayes-Genis, A.; Gotsman, I.; Whalley, G.; Earle, N.; Poppe, K.K.; et al. Renal dysfunction in patients with heart failure with preserved versus reduced ejection fraction: Impact of the new Chronic Kidney Disease-Epidemiology Collaboration Group formula. Circ. Heart Fail. 2012, 5, 309–314. [Google Scholar] [CrossRef] [Green Version]
- Legrand, M.; Rossignol, P. Cardiovascular Consequences of Acute Kidney Injury. N. Engl. J. Med. 2020, 382, 2238–2247. [Google Scholar] [CrossRef]
- Rangaswami, J.; Bhalla, V.; Blair, J.E.A.; Chang, T.I.; Costa, S.; Lentine, K.L.; Lerma, E.V.; Mezue, K.; Molitch, M.; Mullens, W.; et al. Cardiorenal Syndrome: Classification, Pathophysiology, Diagnosis, and Treatment Strategies: A Scientific Statement From the American Heart Association. Circulation 2019, 139, e840–e878. [Google Scholar] [CrossRef]
- Burnstock, G.; Evans, L.C.; Bailey, M.A. Purinergic signalling in the kidney in health and disease. Purinergic Signal. 2014, 10, 71–101. [Google Scholar] [CrossRef] [Green Version]
- Carlstrom, M.; Wilcox, C.S.; Arendshorst, W.J. Renal autoregulation in health and disease. Physiol. Rev. 2015, 95, 405–511. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, O.; Wangensteen, R.; Osuna, A.; Vargas, F. Renal vascular reactivity to P(2)-purinoceptor activation in spontaneously hypertensive rats. Pharmacology 2000, 60, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Menzies, R.I.; Unwin, R.J.; Dash, R.K.; Beard, D.A.; Cowley, A.W., Jr.; Carlson, B.E.; Mullins, J.J.; Bailey, M.A. Effect of P2X4 and P2X7 receptor antagonism on the pressure diuresis relationship in rats. Front. Physiol. 2013, 4, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzies, R.I.; Unwin, R.J.; Bailey, M.A. Renal P2 receptors and hypertension. Acta Physiol. 2015, 213, 232–241. [Google Scholar] [CrossRef]
- Kowalski, R.; Kreft, E.; Kasztan, M.; Jankowski, M.; Szczepanska-Konkel, M. Chronic renal denervation increases renal tubular response to P2X receptor agonists in rats: Implication for renal sympathetic nerve ablation. Nephrol. Dial. Transplant. 2012, 27, 3443–3448. [Google Scholar] [CrossRef] [Green Version]
- Wildman, S.S.; Marks, J.; Churchill, L.J.; Peppiatt, C.M.; Chraibi, A.; Shirley, D.G.; Horisberger, J.D.; King, B.F.; Unwin, R.J. Regulatory interdependence of cloned epithelial Na+ channels and P2X receptors. J. Am. Soc. Nephrol. 2005, 16, 2586–2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wildman, S.S.; Marks, J.; Turner, C.M.; Yew-Booth, L.; Peppiatt-Wildman, C.M.; King, B.F.; Shirley, D.G.; Wang, W.; Unwin, R.J. Sodium-dependent regulation of renal amiloride-sensitive currents by apical P2 receptors. J. Am. Soc. Nephrol. 2008, 19, 731–742. [Google Scholar] [CrossRef] [Green Version]
- Craigie, E.; Menzies, R.I.; Larsen, C.K.; Jacquillet, G.; Carrel, M.; Wildman, S.S.; Loffing, J.; Leipziger, J.; Shirley, D.G.; Bailey, M.A.; et al. The renal and blood pressure response to low sodium diet in P2X4 receptor knockout mice. Physiol. Rep. 2018, 6, e13899. [Google Scholar] [CrossRef]
- Howarth, A.R.; Conway, B.R.; Bailey, M.A. Vascular and inflammatory actions of P2X receptors in renal injury. Auton. Neurosci. 2015, 191, 135–140. [Google Scholar] [CrossRef]
- Han, S.J.; Lovaszi, M.; Kim, M.; D’Agati, V.; Hasko, G.; Lee, H.T. P2X4 receptor exacerbates ischemic AKI and induces renal proximal tubular NLRP3 inflammasome signaling. FASEB J. 2020, 34, 5465–5482. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Turner, C.M.; Hewitt, R.; Smith, J.; Bhangal, G.; Pusey, C.D.; Unwin, R.J.; Tam, F.W. Exaggerated renal fibrosis in P2X4 receptor-deficient mice following unilateral ureteric obstruction. Nephrol. Dial. Transplant. 2014, 29, 1350–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohn, J.N.; Quyyumi, A.A.; Hollenberg, N.K.; Jamerson, K.A. Surrogate markers for cardiovascular disease: Functional markers. Circulation 2004, 109, IV31–IV46. [Google Scholar] [CrossRef] [Green Version]
- Forst, A.L.; Olteanu, V.S.; Mollet, G.; Wlodkowski, T.; Schaefer, F.; Dietrich, A.; Reiser, J.; Gudermann, T.; Mederos y Schnitzler, M.; Storch, U. Podocyte Purinergic P2X4 Channels Are Mechanotransducers That Mediate Cytoskeletal Disorganization. J. Am. Soc. Nephrol. 2016, 27, 848–862. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Zhang, J.; Zhang, W.; Zhang, J.; Yang, J.; Li, K.; He, Y. ATP-P2X4 signaling mediates NLRP3 inflammasome activation: A novel pathway of diabetic nephropathy. Int. J. Biochem. Cell Biol. 2013, 45, 932–943. [Google Scholar] [CrossRef]
- Forfia, P.R.; Vaidya, A.; Wiegers, S.E. Pulmonary heart disease: The heart-lung interaction and its impact on patient phenotypes. Pulm. Circ. 2013, 3, 5–19. [Google Scholar] [CrossRef] [Green Version]
- Galie, N.; Humbert, M.; Vachiery, J.L.; Gibbs, S.; Lang, I.; Torbicki, A.; Simonneau, G.; Peacock, A.; Vonk Noordegraaf, A.; Beghetti, M.; et al. 2015 ESC/ERS Guidelines for the diagnosis and treatment of pulmonary hypertension: The Joint Task Force for the Diagnosis and Treatment of Pulmonary Hypertension of the European Society of Cardiology (ESC) and the European Respiratory Society (ERS): Endorsed by: Association for European Paediatric and Congenital Cardiology (AEPC), International Society for Heart and Lung Transplantation (ISHLT). Eur. Heart J. 2016, 37, 67–119. [Google Scholar] [CrossRef]
- Guibert, C.; Pacaud, P.; Loirand, G.; Marthan, R.; Savineau, J.P. Effect of extracellular ATP on cytosolic Ca2+ concentration in rat pulmonary artery myocytes. Am. J. Physiol. 1996, 271, L450–L458. [Google Scholar] [CrossRef]
- Neely, C.F.; Matot, I.; Batra, V.K.; Bo, X.; Burnstock, G. P2X purinoceptors in the feline pulmonary vascular bed: Distribution and selective in vivo pharmacological probes. Am. J. Physiol. 1996, 270, L889–L897. [Google Scholar] [CrossRef]
- Visovatti, S.H.; Hyman, M.C.; Goonewardena, S.N.; Anyanwu, A.C.; Kanthi, Y.; Robichaud, P.; Wang, J.; Petrovic-Djergovic, D.; Rattan, R.; Burant, C.F.; et al. Purinergic dysregulation in pulmonary hypertension. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H286–H298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syed, N.I.; Tengah, A.; Paul, A.; Kennedy, C. Characterisation of P2X receptors expressed in rat pulmonary arteries. Eur. J. Pharmacol. 2010, 649, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Qasabian, R.A.; Schyvens, C.; Owe-Young, R.; Killen, J.P.; Macdonald, P.S.; Conigrave, A.D.; Williamson, D.J. Characterization of the P2 receptors in rabbit pulmonary artery. Br. J. Pharmacol. 1997, 120, 553–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, K.; Sokabe, T.; Ohura, N.; Nakatsuka, H.; Kamiya, A.; Ando, J. Endogenously released ATP mediates shear stress-induced Ca2+ influx into pulmonary artery endothelial cells. Am. J. Physiol Heart Circ. Physiol 2003, 285, H793–H803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, K.; Imamura, H.; Ando, J. Shear stress augments mitochondrial ATP generation that triggers ATP release and Ca(2+) signaling in vascular endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1477–H1485. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.G.; Jacobson, K.A. Purinergic Signaling in Mast Cell Degranulation and Asthma. Front. Pharmacol. 2017, 8, 947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnstock, G.; Brouns, I.; Adriaensen, D.; Timmermans, J.P. Purinergic signaling in the airways. Pharmacol Rev. 2012, 64, 834–868. [Google Scholar] [CrossRef] [Green Version]
- Antonioli, L.; Blandizzi, C.; Pacher, P.; Hasko, G. The Purinergic System as a Pharmacological Target for the Treatment of Immune-Mediated Inflammatory Diseases. Pharmacol. Rev. 2019, 71, 345–382. [Google Scholar] [CrossRef]
- Le, T.T.; Berg, N.K.; Harting, M.T.; Li, X.; Eltzschig, H.K.; Yuan, X. Purinergic Signaling in Pulmonary Inflammation. Front. Immunol. 2019, 10, 1633. [Google Scholar] [CrossRef]
- Celutkiene, J.; Balciunas, M.; Kablucko, D.; Vaitkeviciute, L.; Blasciuk, J.; Danila, E. Challenges of Treating Acute Heart Failure in Patients with Chronic Obstructive Pulmonary Disease. Card Fail. Rev. 2017, 3, 56–61. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Thompson, L.F.; Karhausen, J.; Cotta, R.J.; Ibla, J.C.; Robson, S.C.; Colgan, S.P. Endogenous adenosine produced during hypoxia attenuates neutrophil accumulation: Coordination by extracellular nucleotide metabolism. Blood 2004, 104, 3986–3992. [Google Scholar] [CrossRef] [PubMed]
- Winkelmann, V.E.; Thompson, K.E.; Neuland, K.; Jaramillo, A.M.; Fois, G.; Schmidt, H.; Wittekindt, O.H.; Han, W.; Tuvim, M.J.; Dickey, B.F.; et al. Inflammation-induced upregulation of P2X4 expression augments mucin secretion in airway epithelia. Am. J. Physiol. Lung Cell Mol. Physiol. 2019, 316, L58–L70. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G. P2X ion channel receptors and inflammation. Purinergic Signal. 2016, 12, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabala, A.; Vazquez-Villoldo, N.; Rissiek, B.; Gejo, J.; Martin, A.; Palomino, A.; Perez-Samartin, A.; Pulagam, K.R.; Lukowiak, M.; Capetillo-Zarate, E.; et al. P2X4 receptor controls microglia activation and favors remyelination in autoimmune encephalitis. EMBO Mol. Med. 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Zech, A.; Wiesler, B.; Ayata, C.K.; Schlaich, T.; Durk, T.; Hossfeld, M.; Ehrat, N.; Cicko, S.; Idzko, M. P2rx4 deficiency in mice alleviates allergen-induced airway inflammation. Oncotarget 2016, 7, 80288–80297. [Google Scholar] [CrossRef] [Green Version]
- Csoka, B.; Nemeth, Z.H.; Szabo, I.; Davies, D.L.; Varga, Z.V.; Paloczi, J.; Falzoni, S.; Di Virgilio, F.; Muramatsu, R.; Yamashita, T.; et al. Macrophage P2X4 receptors augment bacterial killing and protect against sepsis. JCI Insight 2018, 3, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Xia, Q.; Feng, X.; Cao, F.; Yu, H.; Song, Y.; Ni, X. Effect of P2X4R on airway inflammation and airway remodeling in allergic airway challenge in mice. Mol. Med. Rep. 2016, 13, 697–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledderose, C.; Liu, K.; Kondo, Y.; Slubowski, C.J.; Dertnig, T.; Denicolo, S.; Arbab, M.; Hubner, J.; Konrad, K.; Fakhari, M.; et al. Purinergic P2X4 receptors and mitochondrial ATP production regulate T cell migration. J. Clin. Investig. 2018, 128, 3583–3594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paalme, V.; Rump, A.; Mado, K.; Teras, M.; Truumees, B.; Aitai, H.; Ratas, K.; Bourge, M.; Chiang, C.S.; Ghalali, A.; et al. Human Peripheral Blood Eosinophils Express High Levels of the Purinergic Receptor P2X4. Front. Immunol. 2019, 10, 2074. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, M.; Nara, M.; Tamada, T.; Kume, H.; Oguma, T.; Kikuchi, T.; Zaini, J.; Moriya, T.; Ichinose, M.; Tamura, G.; et al. Regulation of adenosine 5’-triphosphate (ATP)-gated P2X(4) receptors on tracheal smooth muscle cells. Respir. Physiol. Neurobiol. 2009, 166, 61–67. [Google Scholar] [CrossRef]
- Mishra, A.; Chintagari, N.R.; Guo, Y.; Weng, T.; Su, L.; Liu, L. Purinergic P2X7 receptor regulates lung surfactant secretion in a paracrine manner. J. Cell Sci. 2011, 124, 657–668. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.S.; Reigada, D.; Mitchell, C.H.; Bates, S.R.; Margulies, S.S.; Koval, M. Paracrine stimulation of surfactant secretion by extracellular ATP in response to mechanical deformation. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 289, L489–L496. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Mallampalli, R.K. The Role of Surfactant in Lung Disease and Host Defense against Pulmonary Infections. Ann. Am. Thorac Soc. 2015, 12, 765–774. [Google Scholar] [CrossRef] [Green Version]
- Miklavc, P.; Mair, N.; Wittekindt, O.H.; Haller, T.; Dietl, P.; Felder, E.; Timmler, M.; Frick, M. Fusion-activated Ca2+ entry via vesicular P2X4 receptors promotes fusion pore opening and exocytotic content release in pneumocytes. Proc. Natl. Acad. Sci. USA 2011, 108, 14503–14508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, K.E.; Korbmacher, J.P.; Hecht, E.; Hobi, N.; Wittekindt, O.H.; Dietl, P.; Kranz, C.; Frick, M. Fusion-activated cation entry (FACE) via P2X(4) couples surfactant secretion and alveolar fluid transport. FASEB J. 2013, 27, 1772–1783. [Google Scholar] [CrossRef] [PubMed]
- Fois, G.; Winkelmann, V.E.; Bareis, L.; Staudenmaier, L.; Hecht, E.; Ziller, C.; Ehinger, K.; Schymeinsky, J.; Kranz, C.; Frick, M. ATP is stored in lamellar bodies to activate vesicular P2X4 in an autocrine fashion upon exocytosis. J. Gen. Physiol. 2018, 150, 277–291. [Google Scholar] [CrossRef] [Green Version]
- Hasan, D.; Satalin, J.; van der Zee, P.; Kollisch-Singule, M.; Blankman, P.; Shono, A.; Somhorst, P.; den Uil, C.; Meeder, H.; Kotani, T.; et al. Excessive Extracellular ATP Desensitizes P2Y2 and P2X4 ATP Receptors Provoking Surfactant Impairment Ending in Ventilation-Induced Lung Injury. Int. J. Mol. Sci. 2018, 19, 1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajagopal, K.; Keller, S.P.; Akkanti, B.; Bime, C.; Loyalka, P.; Cheema, F.H.; Zwischenberger, J.B.; El Banayosy, A.; Pappalardo, F.; Slaughter, M.S.; et al. Advanced Pulmonary and Cardiac Support of COVID-19 Patients: Emerging Recommendations From ASAIO-a Living Working Document. Circ. Heart Fail. 2020, 13, e007175. [Google Scholar] [CrossRef]
- Matthay, M.A.; Zemans, R.L.; Zimmerman, G.A.; Arabi, Y.M.; Beitler, J.R.; Mercat, A.; Herridge, M.; Randolph, A.G.; Calfee, C.S. Acute respiratory distress syndrome. Nat. Rev. Dis. Primers 2019, 5, 18. [Google Scholar] [CrossRef]
- Albert, R.K. The role of ventilation-induced surfactant dysfunction and atelectasis in causing acute respiratory distress syndrome. Am. J. Respir Crit Care Med. 2012, 185, 702–708. [Google Scholar] [CrossRef]
- Stokes, L.; Layhadi, J.A.; Bibic, L.; Dhuna, K.; Fountain, S.J. P2X4 Receptor Function in the Nervous System and Current Breakthroughs in Pharmacology. Front. Pharmacol. 2017, 8, 291. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.J.; Zhu, Z.M.; Liu, Z.X. The role of P2X4 receptor in neuropathic pain and its pharmacological properties. Pharmacol. Res. 2020, 158, 104875. [Google Scholar] [CrossRef]
- Tsuda, M.; Shigemoto-Mogami, Y.; Koizumi, S.; Mizokoshi, A.; Kohsaka, S.; Salter, M.W.; Inoue, K. P2X4 receptors induced in spinal microglia gate tactile allodynia after nerve injury. Nature 2003, 424, 778–783. [Google Scholar] [CrossRef] [PubMed]
- Ulmann, L.; Hatcher, J.P.; Hughes, J.P.; Chaumont, S.; Green, P.J.; Conquet, F.; Buell, G.N.; Reeve, A.J.; Chessell, I.P.; Rassendren, F. Up-regulation of P2X4 receptors in spinal microglia after peripheral nerve injury mediates BDNF release and neuropathic pain. J. Neurosci. 2008, 28, 11263–11268. [Google Scholar] [CrossRef] [PubMed]
- Ulmann, L.; Hirbec, H.; Rassendren, F. P2X4 receptors mediate PGE2 release by tissue-resident macrophages and initiate inflammatory pain. EMBO J. 2010, 29, 2290–2300. [Google Scholar] [CrossRef] [Green Version]
- Long, T.; He, W.; Pan, Q.; Zhang, S.; Zhang, D.; Qin, G.; Chen, L.; Zhou, J. Microglia P2X4R-BDNF signalling contributes to central sensitization in a recurrent nitroglycerin-induced chronic migraine model. J. Headache Pain 2020, 21, 4. [Google Scholar] [CrossRef] [Green Version]
- Beggs, S.; Trang, T.; Salter, M.W. P2X4R+ microglia drive neuropathic pain. Nat. Neurosci. 2012, 15, 1068–1073. [Google Scholar] [CrossRef]
- Sim, J.A.; Chaumont, S.; Jo, J.; Ulmann, L.; Young, M.T.; Cho, K.; Buell, G.; North, R.A.; Rassendren, F. Altered hippocampal synaptic potentiation in P2X4 knock-out mice. J. Neurosci. 2006, 26, 9006–9009. [Google Scholar] [CrossRef]
- Baxter, A.W.; Choi, S.J.; Sim, J.A.; North, R.A. Role of P2X4 receptors in synaptic strengthening in mouse CA1 hippocampal neurons. Eur. J. Neurosci. 2011, 34, 213–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Rivero Vaccari, J.P.; Bastien, D.; Yurcisin, G.; Pineau, I.; Dietrich, W.D.; De Koninck, Y.; Keane, R.W.; Lacroix, S. P2X4 receptors influence inflammasome activation after spinal cord injury. J. Neurosci. 2012, 32, 3058–3066. [Google Scholar] [CrossRef] [PubMed]
- Ulmann, L.; Levavasseur, F.; Avignone, E.; Peyroutou, R.; Hirbec, H.; Audinat, E.; Rassendren, F. Involvement of P2X4 receptors in hippocampal microglial activation after status epilepticus. Glia 2013, 61, 1306–1319. [Google Scholar] [CrossRef] [PubMed]
- Khoja, S.; Huynh, N.; Warnecke, A.M.P.; Asatryan, L.; Jakowec, M.W.; Davies, D.L. Preclinical evaluation of avermectins as novel therapeutic agents for alcohol use disorders. Psychopharmacology 2018, 235, 1697–1709. [Google Scholar] [CrossRef] [PubMed]
- Franklin, K.M.; Asatryan, L.; Jakowec, M.W.; Trudell, J.R.; Bell, R.L.; Davies, D.L. P2X4 receptors (P2X4Rs) represent a novel target for the development of drugs to prevent and/or treat alcohol use disorders. Front. Neurosci. 2014, 8, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Guo, N.; Ma, Y.; Ning, B.; Wang, Y.; Kou, L. Inhibition of P2X4 suppresses joint inflammation and damage in collagen-induced arthritis. Inflammation 2014, 37, 146–153. [Google Scholar] [CrossRef]


{kind=link}
| Compound | EC50 or IC50 (µM) * | P2X Cross-Reactivity/Fold Selectivity | References | |||
|---|---|---|---|---|---|---|
| Human | Rat | Mouse | ||||
| Agonist | ATP | 0.7–5.7 | 1.7–16.2 | 0.3–2.3 | [12,17,21,22,25,26,27] | |
| ATPγS | 10.9 | [28] | ||||
| 2-meSATP | 0.3–2.2 | ~ 1–10 | 1.4 | [21,27,28,29] | ||
| CTP | ~0.1–1 mM | P2X1,2,3,7 | [21,30,31,32] | |||
| BzATP | 0.5–9.4 | >100 | 2.9 | P2X7 (h) | [12,27,28] | |
| α,β-meATP | 0.8–19 | ≥100 | 7–>100 | P2X1,3 | [26,27,28] | |
| β,γ-meATP | >100 | >100 | >100 | [27] | ||
| AP4AƔ | 0.1-3 | 20–>100 | 2.6–>100 | [26,27,28] | ||
| AR-C67085MX | 2.5 | >100 | >100 | [27] | ||
| 8-Azido-ATP | >100 | >100 | >100 | [27] | ||
| β,γ-Imido-ATP | 6.5 | >100 | >100 | [27] | ||
| MRS-2339 | [33] | |||||
| MRS-2978 | [34] | |||||
| PSB-0412 | 2.1 | [27] | ||||
| Antagonist | PPADS | 9.6–>100 | >300 | >100 | [26,28] | |
| Suramin | >100 | >100 | >100 | [26,28] | ||
| KN-62 | >10 | >10 | >10 | [26] | ||
| TNP-ATP | 1.5 | 1.3–4.71 | 1.3–4.2 | P2X1,2,3,7 | [27,35,36,37,38] | |
| Brilliant Blue G | 3–100 | 1000-fold more potent at P2X7 (r) | [39] | |||
| 5-BDBD | 0.3–1.2 | 0.75–3.5 | 2.04 | 10-fold P2X1 (r), 3-fold P2X3 (r) | [27,36,40] | |
| BX-430 | 0.54–1 | 10 to 100-fold P2X1,3,5,7 | [41,42] | |||
| Carbamazepine der. † | 3.44 | 54.6 | 14.9 | 2 to 30-fold P2X1,2,3,7 (h) | [43] | |
| PSB-12054 | 0.19 | 2.10 | 1.8 | ≥50-fold P2X1,2,3,7 (h) | [35] | |
| PSB-12062 | 1.4 | 0.9 | 1.8 | ≥35-fold P2X1,2,3,7 (h) | [35] | |
| PSB-15417 | 10 ‡ | >5-fold P2X7 (h) | [44] | |||
| NP-1815-PX | 0.26 | P2X1,2,3,7 | [45] | |||
| NC-2600 Ʊ | [46] | |||||
| UoS14919 | 61 nM | [47] | ||||
| Paroxetine | 1.87–4.8 | 1.64–2.45 | 0.7 | [27,48] | ||
| Duloxetine | 1.59–17 | [49,50] | ||||
| BAY-1797 | 0.2 | [51] | ||||
| IgG#151-LO || | 0.7 | 10,000-fold more potent over other P2X | [52] | |||
| PAM | Ivermectin | 0.1 | P2X7 (h) | [53,54] | ||
| Cibacron blue | >300 | [55] | ||||
| Ginsenosides | 7.5–10.5 | P2X7 | [56] | |||
| Testosterone butyrate | 30 ‡ | P2X2 | [57] | |||
| Zn2+ | 1.8 | [58] | ||||
| Cd2+ | 7.3 | [58] | ||||
| Alfaxalone | 0.4–1.6 | [59] | ||||
| Allopregnanolone | 0.4 | [59] | ||||
| Propofol | 56 | [39] | ||||
| NM | Ethanol | 5–200 mM ‡ | PAM at rat P2X3 | [60,61] | ||
| H + | pKa ~6.8 | inhibits P2X1,3,4; stimulates P2X2 | [62] | |||
| Hg2+ | 9 | [58] | ||||
| Cu2+ | 8.6 | [58] | ||||
| Fluvastatin ¥ | 10 for 1 h | [63] | ||||
| Filipin III ¥ | 10 for 30 min | [63] | ||||
| Methyl-β-cyclodextrin ¥ | 10 mM, 1 h | [63] | ||||
| t-DCA § | 160 | [64] | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bragança, B.; Correia-de-Sá, P. Resolving the Ionotropic P2X4 Receptor Mystery Points towards a New Therapeutic Target for Cardiovascular Diseases. Int. J. Mol. Sci. 2020, 21, 5005. https://doi.org/10.3390/ijms21145005
Bragança B, Correia-de-Sá P. Resolving the Ionotropic P2X4 Receptor Mystery Points towards a New Therapeutic Target for Cardiovascular Diseases. International Journal of Molecular Sciences. 2020; 21(14):5005. https://doi.org/10.3390/ijms21145005
Chicago/Turabian StyleBragança, Bruno, and Paulo Correia-de-Sá. 2020. "Resolving the Ionotropic P2X4 Receptor Mystery Points towards a New Therapeutic Target for Cardiovascular Diseases" International Journal of Molecular Sciences 21, no. 14: 5005. https://doi.org/10.3390/ijms21145005
APA StyleBragança, B., & Correia-de-Sá, P. (2020). Resolving the Ionotropic P2X4 Receptor Mystery Points towards a New Therapeutic Target for Cardiovascular Diseases. International Journal of Molecular Sciences, 21(14), 5005. https://doi.org/10.3390/ijms21145005