Angiotensin-(1-7) Improves Integrated Cardiometabolic Function in Aged Mice



Abstract
:1. Introduction
2. Results
2.1. Ang-(1-7) Does Not Affect Body Weight or Composition in Aged Mice
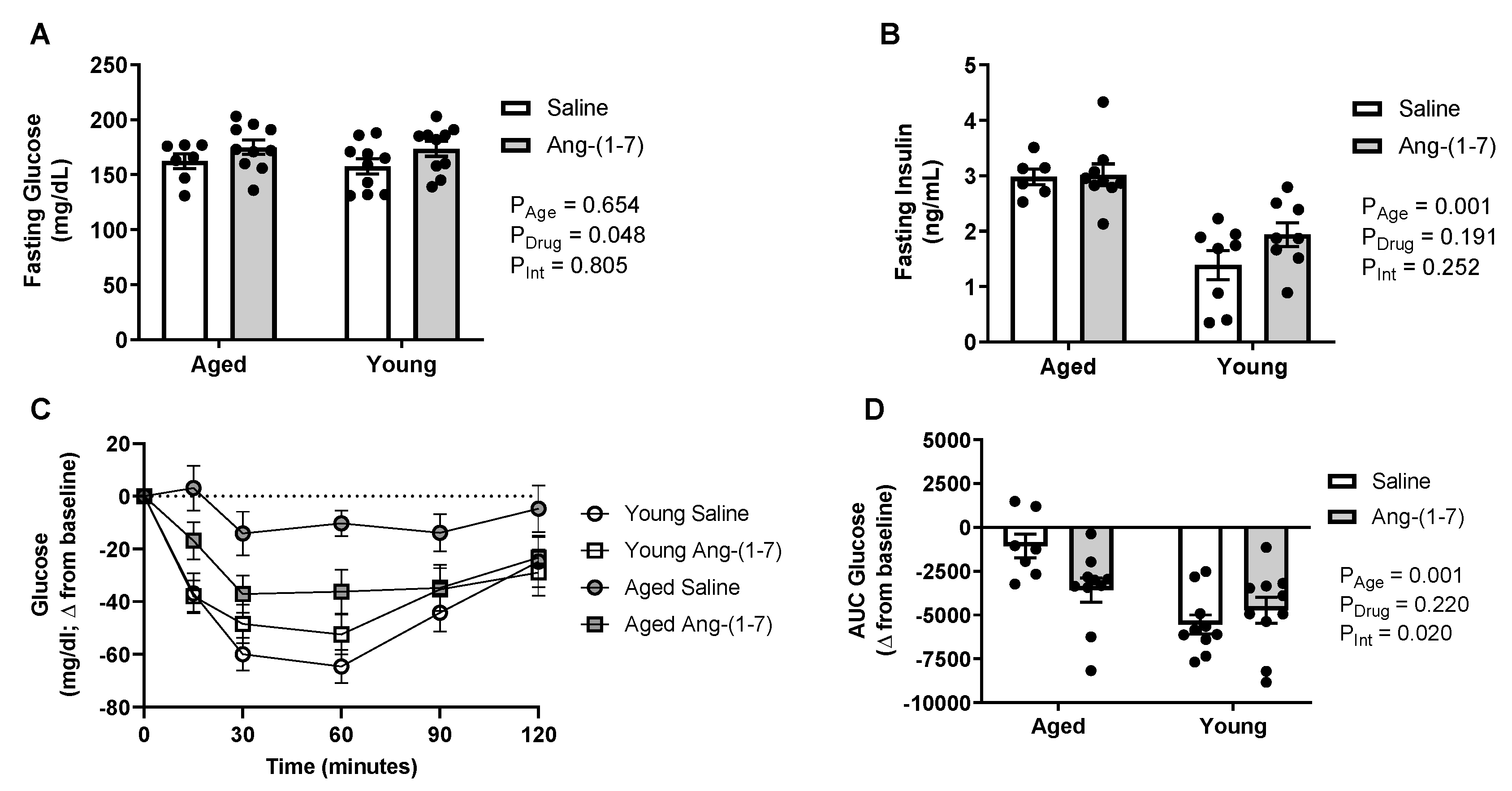
2.2. Ang-(1-7) Improves Insulin Sensitivity in Aged Mice
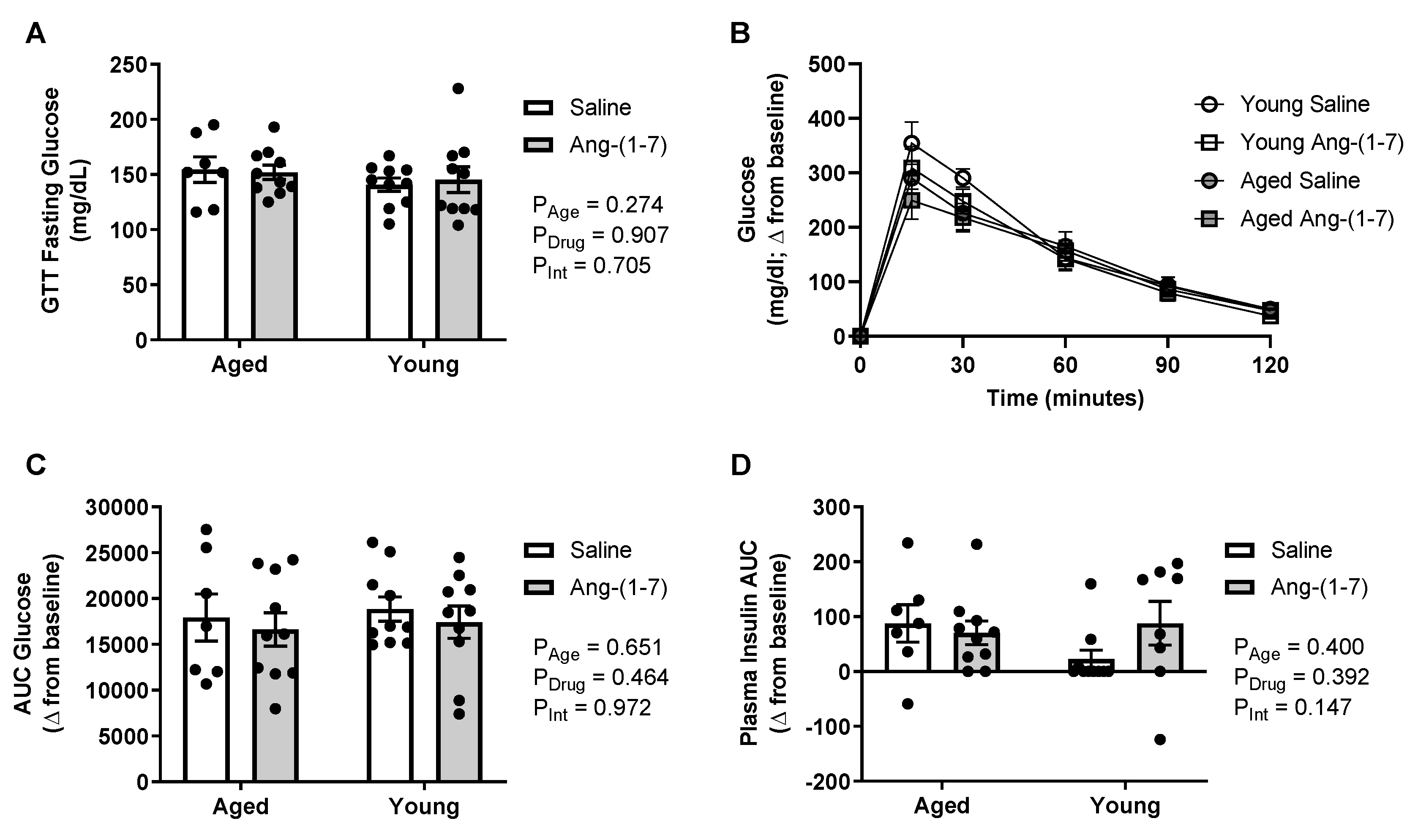
2.3. Glucose Tolerance Is Not Affected by Either Aging or Ang-(1-7) Treatment
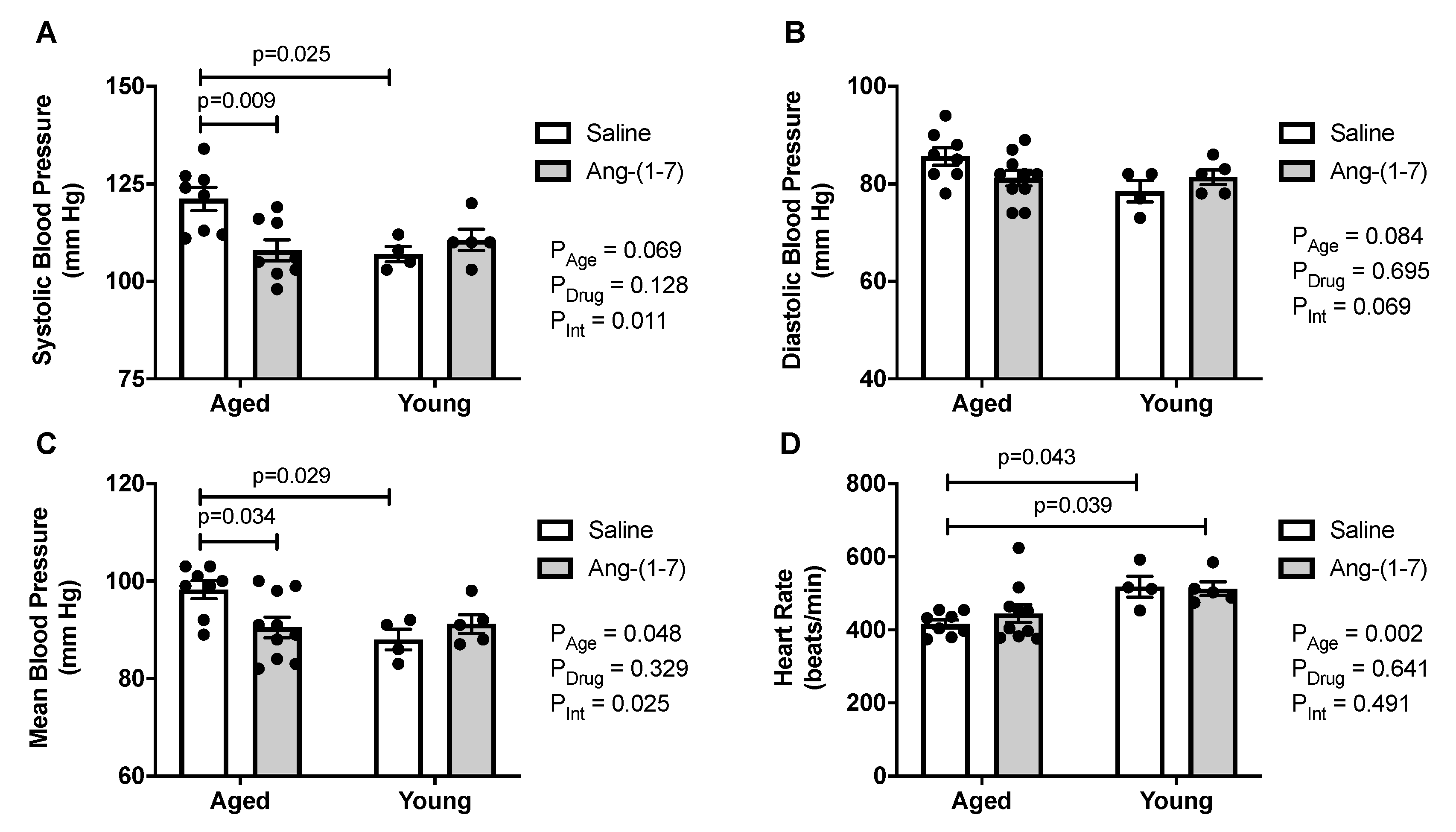
2.4. Ang-(1-7) Decreases Systolic and Mean Blood Pressure in Aged Mice
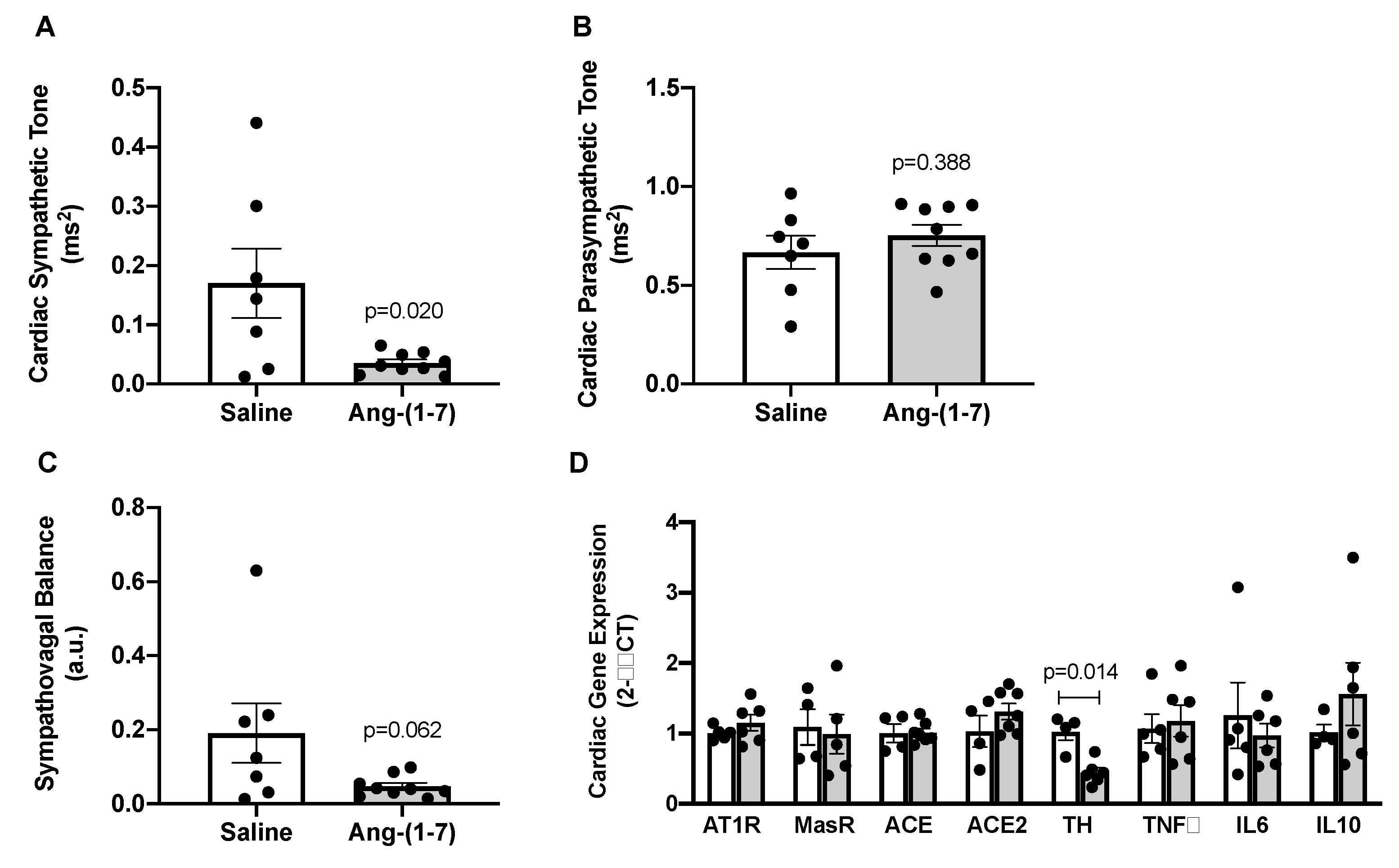
2.5. Ang-(1-7) Decreases Measures of Cardiac Sympathetic Tone in Aged Mice
3. Discussion
4. Materials and Methods
4.1. Approvals
4.2. General Protocol
4.3. Insulin and Glucose Tolerance Testing
4.4. Body Composition
4.5. Blood Pressure and Heart Rate Measurements
4.6. Euthanasia and Blood and Tissue Collection
4.7. Cardiac Gene Expression of RAS, Inflammatory and Sympathetic Markers
4.8. Statistical Considerations
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACE | angiotensin-converting enzyme |
| ACE2 | angiotensin-converting enzyme 2 |
| Ang | angiotensin |
| ARBs | angiotensin receptor blockers |
| AT1R | angiotensin II type 1 receptor |
| AUC | area under the curve |
| BAT | brown adipose tissue |
| EPF | epididymal visceral white adipose tissue |
| GTT | glucose tolerance test |
| IL6 | interleukin 6 |
| IL10 | interleukin 10 |
| ITT | insulin tolerance test |
| SCF | inguinal subcutaneous white adipose tissue |
| Th | tyrosine hydroxylase |
| TNFα | tumor necrosis factor α |
References
- North, B.J.; Sinclair, D.A. The intersection between aging and cardiovascular disease. Circ. Res. 2012, 110, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Tune, J.D.; Goodwill, A.G.; Sassoon, D.J.; Mather, K.J. Cardiovascular consequences of metabolic syndrome. Transl. Res. 2017, 183, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Unger, T. The role of the renin-angiotensin system in the development of cardiovascular disease. Am. J. Cardiol. 2002, 89, 3–9. [Google Scholar] [CrossRef]
- Miller, A.J.; Arnold, A.C. The renin-angiotensin system in cardiovascular autonomic control: Recent developments and clinical implications. Clin. Auton. Res. 2019, 29, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Luther, J.M.; Brown, N.J. The renin-angiotensin-aldosterone system and glucose homeostasis. Trends Pharmacol. Sci. 2011, 32, 734–739. [Google Scholar] [CrossRef] [Green Version]
- White, M.C.; Fleeman, R.; Arnold, A.C. Sex differences in the metabolic effects of the renin-angiotensin system. Biol. Sex. Differ. 2019, 10, 31. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.C.; Gallagher, P.E.; Diz, D.I. Brain renin-angiotensin system in the nexus of hypertension and aging. Hypertens. Res. 2013, 36, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S. Ageing and the renin-angiotensin system. Nephrol. Dial. Transplant. 1997, 12, 1093–1094. [Google Scholar] [CrossRef]
- Thomas, G.N.; Chan, P.; Tomlinson, B. The role of angiotensin II type 1 receptor antagonists in elderly patients with hypertension. Drugs Aging 2006, 23, 131–155. [Google Scholar] [CrossRef]
- Mohamed, M.A.; Weir, M.R. Renin angiotensin system inhibition in the older person: A review. Clin. Geriatr. Med. 2009, 25, 245–257. [Google Scholar] [CrossRef]
- Lindholm, L.H.; Ibsen, H.; Borch-Johnsen, K.; Olsen, M.H.; Wachtell, K.; Dahlof, B.; Devereux, R.B.; Beevers, G.; de Faire, U.; Fyhrquist, F.; et al. Risk of new-onset diabetes in the Losartan Intervention for Endpoint reduction in hypertension study. J. Hypertens. 2002, 20, 1879–1886. [Google Scholar] [CrossRef]
- Loloi, J.; Miller, A.J.; Bingaman, S.S.; Silberman, Y.; Arnold, A.C. Angiotensin-(1-7) contributes to insulin-sensitizing effects of angiotensin-converting enzyme inhibition in obese mice. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E1204–E1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousif, M.H.; Dhaunsi, G.S.; Makki, B.M.; Qabazard, B.A.; Akhtar, S.; Benter, I.F. Characterization of Angiotensin-(1-7) effects on the cardiovascular system in an experimental model of type-1 diabetes. Pharmacol. Res. 2012, 66, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Kucharewicz, I.; Pawlak, R.; Matys, T.; Pawlak, D.; Buczko, W. Antithrombotic effect of captopril and losartan is mediated by angiotensin-(1-7). Hypertension 2002, 40, 774–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benter, I.F.; Yousif, M.H.; Al-Saleh, F.M.; Raghupathy, R.; Chappell, M.C.; Diz, D.I. Angiotensin-(1-7) blockade attenuates captopril- or hydralazine-induced cardiovascular protection in spontaneously hypertensive rats treated with NG-nitro-L-arginine methyl ester. J. Cardiovasc. Pharmacol. 2011, 57, 559–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangalore, S.; Kumar, S.; Messerli, F.H. Angiotensin-converting enzyme inhibitor associated cough: Deceptive information from the Physicians’ Desk Reference. Am. J. Med. 2010, 123, 1016–1030. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.A.S.; Sampaio, W.O.; Alzamora, A.C.; Motta-Santos, D.; Alenina, N.; Bader, M.; Campagnole-Santos, M.J. The ACE2/Angiotensin-(1-7)/MAS Axis of the Renin-Angiotensin System: Focus on Angiotensin-(1-7). Physiol. Rev. 2018, 98, 505–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, D.; Arnold, A.C. Angiotensin-(1-7): Translational Avenues in Cardiovascular Control. Am. J. Hypertens. 2019, 32, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Grobe, J.L.; Mecca, A.P.; Lingis, M.; Shenoy, V.; Bolton, T.A.; Machado, J.M.; Speth, R.C.; Raizada, M.K.; Katovich, M.J. Prevention of angiotensin II-induced cardiac remodeling by angiotensin-(1-7). Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H736–H742. [Google Scholar] [CrossRef] [Green Version]
- Giani, J.F.; Mayer, M.A.; Munoz, M.C.; Silberman, E.A.; Hocht, C.; Taira, C.A.; Gironacci, M.M.; Turyn, D.; Dominici, F.P. Chronic infusion of angiotensin-(1-7) improves insulin resistance and hypertension induced by a high-fructose diet in rats. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E262–E271. [Google Scholar] [CrossRef] [Green Version]
- Williams, I.M.; Otero, Y.F.; Bracy, D.P.; Wasserman, D.H.; Biaggioni, I.; Arnold, A.C. Chronic Angiotensin-(1-7) Improves Insulin Sensitivity in High-Fat Fed Mice Independent of Blood Pressure. Hypertension 2016, 67, 983–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, S.H.; Andrade, J.M. Angiotensin 1-7: A peptide for preventing and treating metabolic syndrome. Peptides 2014, 59, 34–41. [Google Scholar] [CrossRef]
- Sampaio, W.O.; Nascimento, A.A.; Santos, R.A. Systemic and regional hemodynamic effects of angiotensin-(1-7) in rats. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H1985–H1994. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yin, A.; Zhang, Q.; Zhong, T.; O’Rourke, S.T.; Sun, C. Angiotensin-(1-7) attenuates angiotensin II-induced cardiac hypertrophy via a Sirt3-dependent mechanism. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H980–H991. [Google Scholar] [CrossRef] [PubMed]
- Costa-Fraga, F.P.; Goncalves, G.K.; Souza-Neto, F.P.; Reis, A.M.; Capettini, L.A.; Santos, R.A.; Fraga-Silva, R.A.; Stergiopulos, N.; da Silva, R.F. Age-related changes in vascular responses to angiotensin-(1-7) in female mice. J. Renin Angiotensin Aldosterone Syst. 2018, 19, 1470320318789332. [Google Scholar] [CrossRef] [Green Version]
- Yoon, H.E.; Kim, E.N.; Kim, M.Y.; Lim, J.H.; Jang, I.A.; Ban, T.H.; Shin, S.J.; Park, C.W.; Chang, Y.S.; Choi, B.S. Age-Associated Changes in the Vascular Renin-Angiotensin System in Mice. Oxid. Med. Cell. Longev. 2016, 2016, 6731093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Castillo, A.; Tobon-Cornejo, S.; Del Valle-Mondragon, L.; Torre-Villalvazo, I.; Schcolnik-Cabrera, A.; Guevara-Cruz, M.; Pichardo-Ontiveros, E.; Fuentes-Romero, R.; Bader, M.; Alenina, N.; et al. Angiotensin-(1-7) induces beige fat thermogenesis through the Mas receptor. Metabolism 2020, 103, 154048. [Google Scholar] [CrossRef]
- Carey, R.M. Angiotensin receptors and aging. Hypertension 2007, 50, 33–34. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Takagi, G.; Asai, K.; Resuello, R.G.; Natividad, F.F.; Vatner, D.E.; Vatner, S.F.; Lakatta, E.G. Aging increases aortic MMP-2 activity and angiotensin II in nonhuman primates. Hypertension 2003, 41, 1308–1316. [Google Scholar] [CrossRef] [Green Version]
- Conti, S.; Cassis, P.; Benigni, A. Aging and the renin-angiotensin system. Hypertension 2012, 60, 878–883. [Google Scholar] [CrossRef]
- Seals, D.R.; Esler, M.D. Human ageing and the sympathoadrenal system. J. Physiol. 2000, 528, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Franklin, S.S. Ageing and hypertension: The assessment of blood pressure indices in predicting coronary heart disease. J. Hypertens. Suppl. 1999, 17, S29–S36. [Google Scholar] [PubMed]
- Ferrari, A.U.; Radaelli, A.; Centola, M. Invited review: Aging and the cardiovascular system. J. Appl. Physiol. 2003, 95, 2591–2597. [Google Scholar] [CrossRef]
- Barsha, G.; Denton, K.M.; Mirabito Colafella, K.M. Sex- and age-related differences in arterial pressure and albuminuria in mice. Biol. Sex. Differ. 2016, 7, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosnyak, S.; Widdop, R.E.; Denton, K.M.; Jones, E.S. Differential mechanisms of ang (1-7)-mediated vasodepressor effect in adult and aged candesartan-treated rats. Int. J. Hypertens. 2012, 2012, 192567. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, P.S.; Oliveira, M.F.; Braga, J.F.; Nadu, A.P.; Schreihofer, A.; Santos, R.A.; Campagnole-Santos, M.J. Increasing angiotensin-(1-7) levels in the brain attenuates metabolic syndrome-related risks in fructose-fed rats. Hypertension 2014, 63, 1078–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giani, J.F.; Munoz, M.C.; Mayer, M.A.; Veiras, L.C.; Arranz, C.; Taira, C.A.; Turyn, D.; Toblli, J.E.; Dominici, F.P. Angiotensin-(1-7) improves cardiac remodeling and inhibits growth-promoting pathways in the heart of fructose-fed rats. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H1003–H1013. [Google Scholar] [CrossRef] [Green Version]
- Santiago, N.M.; Guimaraes, P.S.; Sirvente, R.A.; Oliveira, L.A.; Irigoyen, M.C.; Santos, R.A.; Campagnole-Santos, M.J. Lifetime overproduction of circulating Angiotensin-(1-7) attenuates deoxycorticosterone acetate-salt hypertension-induced cardiac dysfunction and remodeling. Hypertension 2010, 55, 889–896. [Google Scholar] [CrossRef] [Green Version]
- Ito, N.; Ohishi, M.; Yamamoto, K.; Tatara, Y.; Shiota, A.; Hayashi, N.; Komai, N.; Yanagitani, Y.; Rakugi, H.; Ogihara, T. Renin-angiotensin inhibition reverses advanced cardiac remodeling in aging spontaneously hypertensive rats. Am. J. Hypertens. 2007, 20, 792–799. [Google Scholar] [CrossRef]
- Chia, C.W.; Egan, J.M.; Ferrucci, L. Age-Related Changes in Glucose Metabolism, Hyperglycemia, and Cardiovascular Risk. Circ. Res. 2018, 123, 886–904. [Google Scholar] [CrossRef]
- Shou, J.; Chen, P.J.; Xiao, W.H. Mechanism of increased risk of insulin resistance in aging skeletal muscle. Diabetol. Metab. Syndr. 2020, 12, 14. [Google Scholar] [CrossRef] [Green Version]
- Santos, S.H.; Braga, J.F.; Mario, E.G.; Porto, L.C.; Rodrigues-Machado Mda, G.; Murari, A.; Botion, L.M.; Alenina, N.; Bader, M.; Santos, R.A. Improved lipid and glucose metabolism in transgenic rats with increased circulating angiotensin-(1-7). Arterioscler. Thromb. Vasc. Biol. 2010, 30, 953–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, M.C.; Miller, A.J.; Loloi, J.; Bingaman, S.S.; Shen, B.; Wang, M.; Silberman, Y.; Lindsey, S.H.; Arnold, A.C. Sex differences in metabolic effects of angiotensin-(1-7) treatment in obese mice. Biol. Sex. Differ. 2019, 10, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcus, Y.; Shefer, G.; Sasson, K.; Kohen, F.; Limor, R.; Pappo, O.; Nevo, N.; Biton, I.; Bach, M.; Berkutzki, T.; et al. Angiotensin 1-7 as means to prevent the metabolic syndrome: Lessons from the fructose-fed rat model. Diabetes 2013, 62, 1121–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nozato, S.; Yamamoto, K.; Takeshita, H.; Nozato, Y.; Imaizumi, Y.; Fujimoto, T.; Yokoyama, S.; Nagasawa, M.; Takeda, M.; Hongyo, K.; et al. Angiotensin 1-7 alleviates aging-associated muscle weakness and bone loss, but is not associated with accelerated aging in ACE2-knockout mice. Clin. Sci. 2019, 133, 2005–2018. [Google Scholar] [CrossRef] [PubMed]
- Sahr, A.; Wolke, C.; Maczewsky, J.; Krippeit-Drews, P.; Tetzner, A.; Drews, G.; Venz, S.; Gurtler, S.; van den Brandt, J.; Berg, S.; et al. The Angiotensin-(1-7)/Mas Axis Improves Pancreatic beta-Cell Function in Vitro and in Vivo. Endocrinology 2016, 157, 4677–4690. [Google Scholar] [CrossRef]
- Andrade, J.M.; Lemos Fde, O.; da Fonseca Pires, S.; Millan, R.D.; de Sousa, F.B.; Guimaraes, A.L.; Qureshi, M.; Feltenberger, J.D.; de Paula, A.M.; Neto, J.T.; et al. Proteomic white adipose tissue analysis of obese mice fed with a high-fat diet and treated with oral angiotensin-(1-7). Peptides 2014, 60, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Muoio, D.M.; Newgard, C.B. Obesity-related derangements in metabolic regulation. Annu. Rev. Biochem. 2006, 75, 367–401. [Google Scholar] [CrossRef]
- Esler, M.; Straznicky, N.; Eikelis, N.; Masuo, K.; Lambert, G.; Lambert, E. Mechanisms of sympathetic activation in obesity-related hypertension. Hypertension 2006, 48, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, H.; Mori, J.; Nakajima, H.; Kawabe, Y.; Tsuma, Y.; Fukuhara, S.; Kodo, K.; Ikoma, K.; Matoba, S.; Oudit, G.Y.; et al. Angiotensin 1-7 stimulates brown adipose tissue and reduces diet-induced obesity. Am. J. Physiol Endocrinol Metab 2018, 314, E131–E138. [Google Scholar] [CrossRef]
- Roks, A.J.; van Geel, P.P.; Pinto, Y.M.; Buikema, H.; Henning, R.H.; de Zeeuw, D.; van Gilst, W.H. Angiotensin-(1-7) is a modulator of the human renin-angiotensin system. Hypertension 1999, 34, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Chappell, M.C.; Ferrario, C.M.; Brosnihan, K.B. Angiotensin-(1-7) augments bradykinin-induced vasodilation by competing with ACE and releasing nitric oxide. Hypertension 1997, 29, 394–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter, Unit | Young Saline | Young Ang-(1-7) | Aged Saline | Aged Ang-(1-7) | PAge | PDrug | PInt |
|---|---|---|---|---|---|---|---|
| n | 10 | 10 | 8 | 10 | |||
| Body composition | |||||||
| Body mass, g | 30.0 ± 0.8 | 30.9 ± 0.6 | 46.1 ± 1.7 *† | 44.4 ± 1.6 *† | 0.001 | 0.740 | 0.302 |
| Adiposity, % | 7.0 ± 0.5 | 7.2 ± 0.7 | 14.6 ± 1.7 *† | 16.7 ± 1.5 *† | 0.001 | 0.328 | 0.451 |
| Lean mass, % | 69.7 ± 0.8 | 69.3 ± 0.9 | 60.7 ± 1.9 *† | 58.2 ± 1.3 *† | 0.001 | 0.251 | 0.403 |
| Fluid mass, % | 6.9 ± 0.1 | 6.7 ± 0.1 | 8.6 ± 0.2 *† | 8.4 ± 0.1 *† | 0.001 | 0.207 | 0.969 |
| Adipose and heart tissue weights | |||||||
| EPF, % | 1.3 ± 0.2 | 1.6 ± 0.3 | 2.8 ± 0.5 | 3.7 ± 0.5 *† | 0.001 | 0.152 | 0.457 |
| SCF, % | 1.0 ± 0.1 | 1.1 ± 0.2 | 2.3 ± 0.5 | 3.2 ± 0.4 *† | 0.001 | 0.178 | 0.278 |
| BAT, % | 0.29 ± 0.05 | 0.29 ± 0.03 | 0.44 ± 0.06 † | 0.45 ± 0.02 *† | 0.001 | 0.809 | 0.877 |
| Heart, % | 0.36 ± 0.06 | 0.44 ± 0.01 | 0.48 ± 0.02 | 0.45 ± 0.03 | 0.038 | 0.362 | 0.085 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miller, A.J.; Bingaman, S.S.; Mehay, D.; Medina, D.; Arnold, A.C. Angiotensin-(1-7) Improves Integrated Cardiometabolic Function in Aged Mice. Int. J. Mol. Sci. 2020, 21, 5131. https://doi.org/10.3390/ijms21145131
Miller AJ, Bingaman SS, Mehay D, Medina D, Arnold AC. Angiotensin-(1-7) Improves Integrated Cardiometabolic Function in Aged Mice. International Journal of Molecular Sciences. 2020; 21(14):5131. https://doi.org/10.3390/ijms21145131
Chicago/Turabian StyleMiller, Amanda J., Sarah S. Bingaman, Darren Mehay, Daniela Medina, and Amy C. Arnold. 2020. "Angiotensin-(1-7) Improves Integrated Cardiometabolic Function in Aged Mice" International Journal of Molecular Sciences 21, no. 14: 5131. https://doi.org/10.3390/ijms21145131
APA StyleMiller, A. J., Bingaman, S. S., Mehay, D., Medina, D., & Arnold, A. C. (2020). Angiotensin-(1-7) Improves Integrated Cardiometabolic Function in Aged Mice. International Journal of Molecular Sciences, 21(14), 5131. https://doi.org/10.3390/ijms21145131