Arming Oncolytic Adenoviruses: Effect of Insertion Site and Splice Acceptor on Transgene Expression and Viral Fitness

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
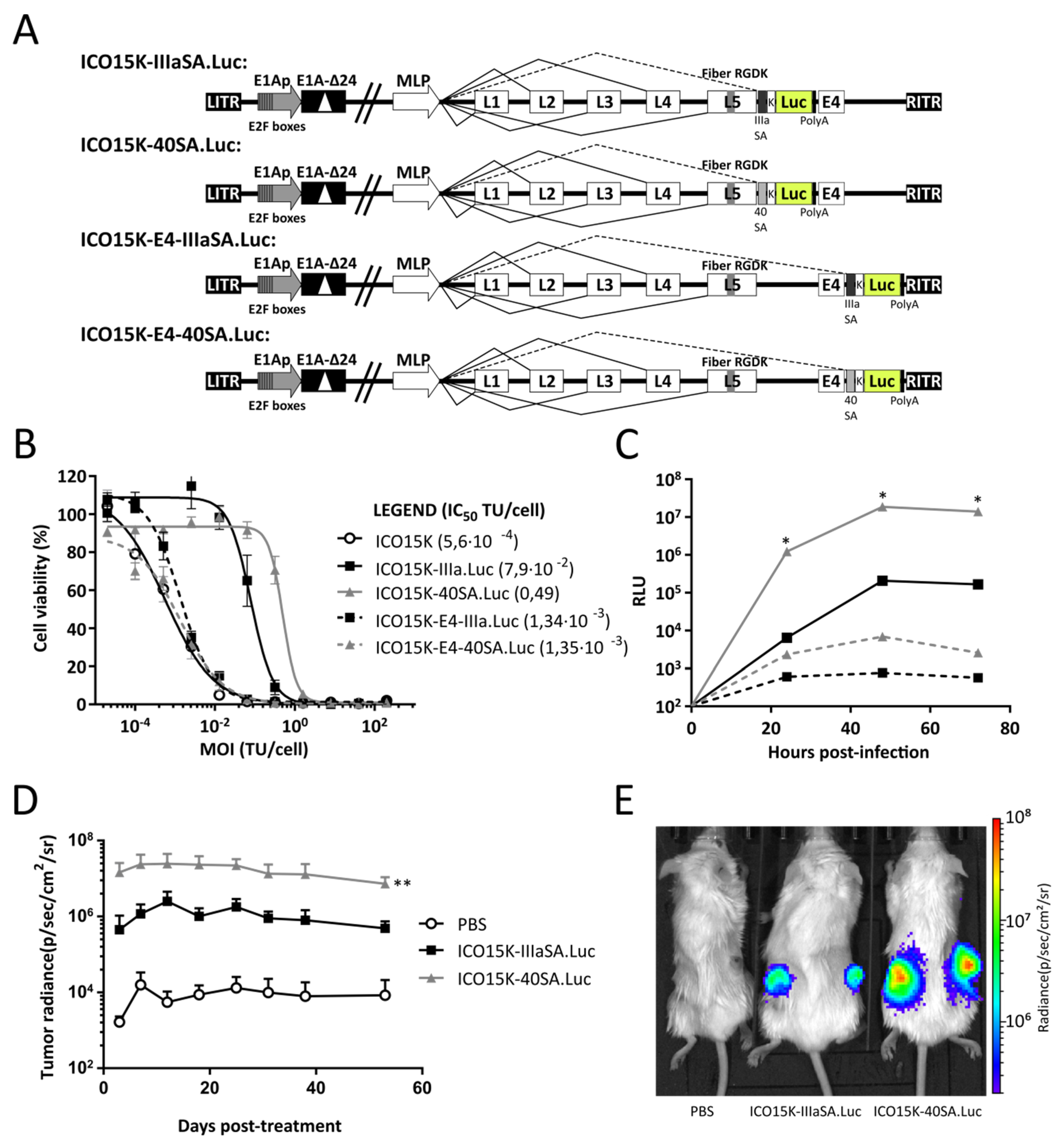
2.1. Generation and Characterization of Luciferase-Armed Oncolytic Adenoviruses
2.2. After-Fiber-Armed Virus with 40SA Produced the Highest Luminescence Levels In Vitro and In Vivo
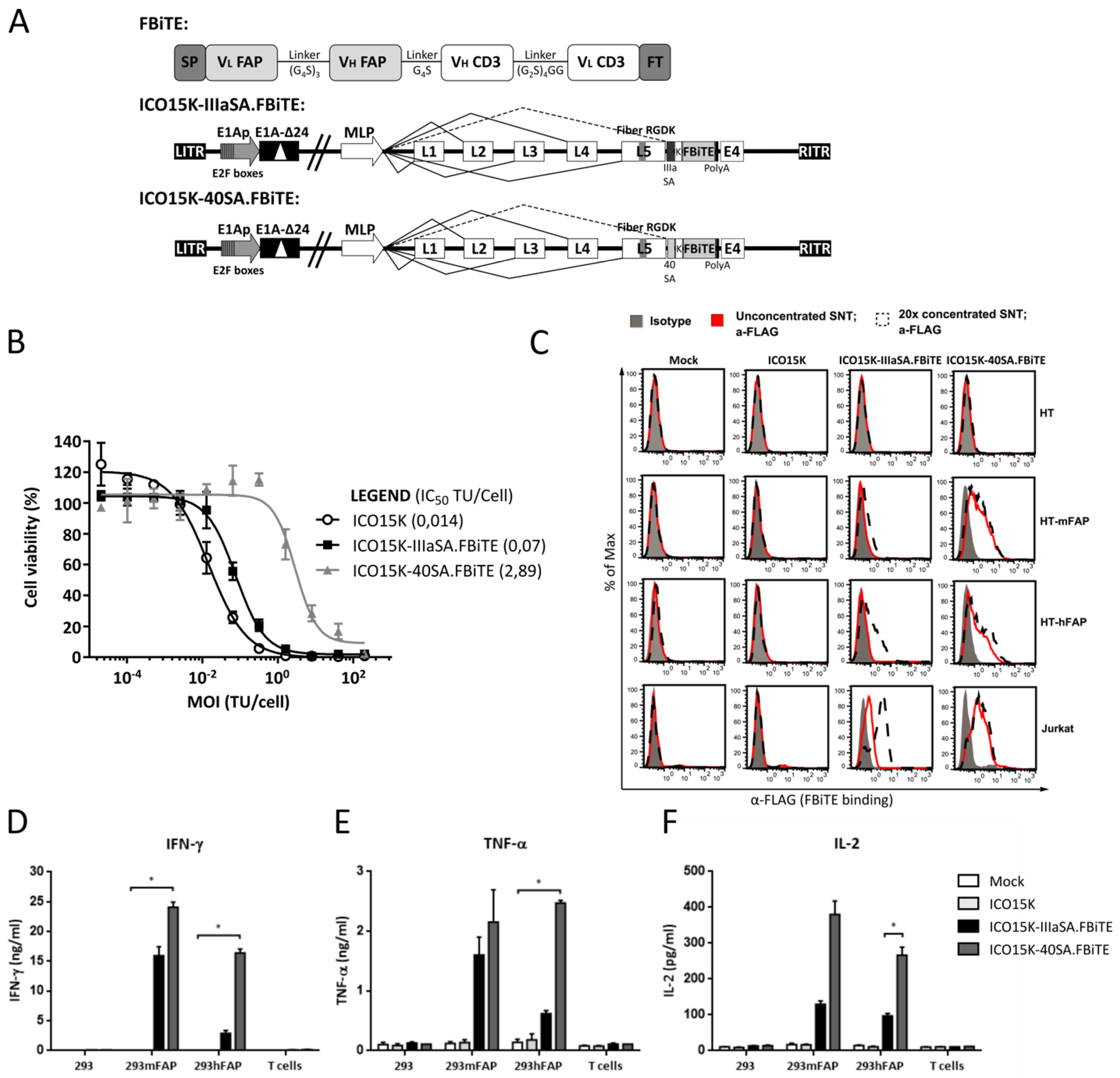
2.3. Generation and Characterization of FBiTE-Expressing Oncolytic Adenoviruses
2.4. The After-Fiber-Armed Virus with 40SA Expressed Higher FBiTE Levels than IIIaSA
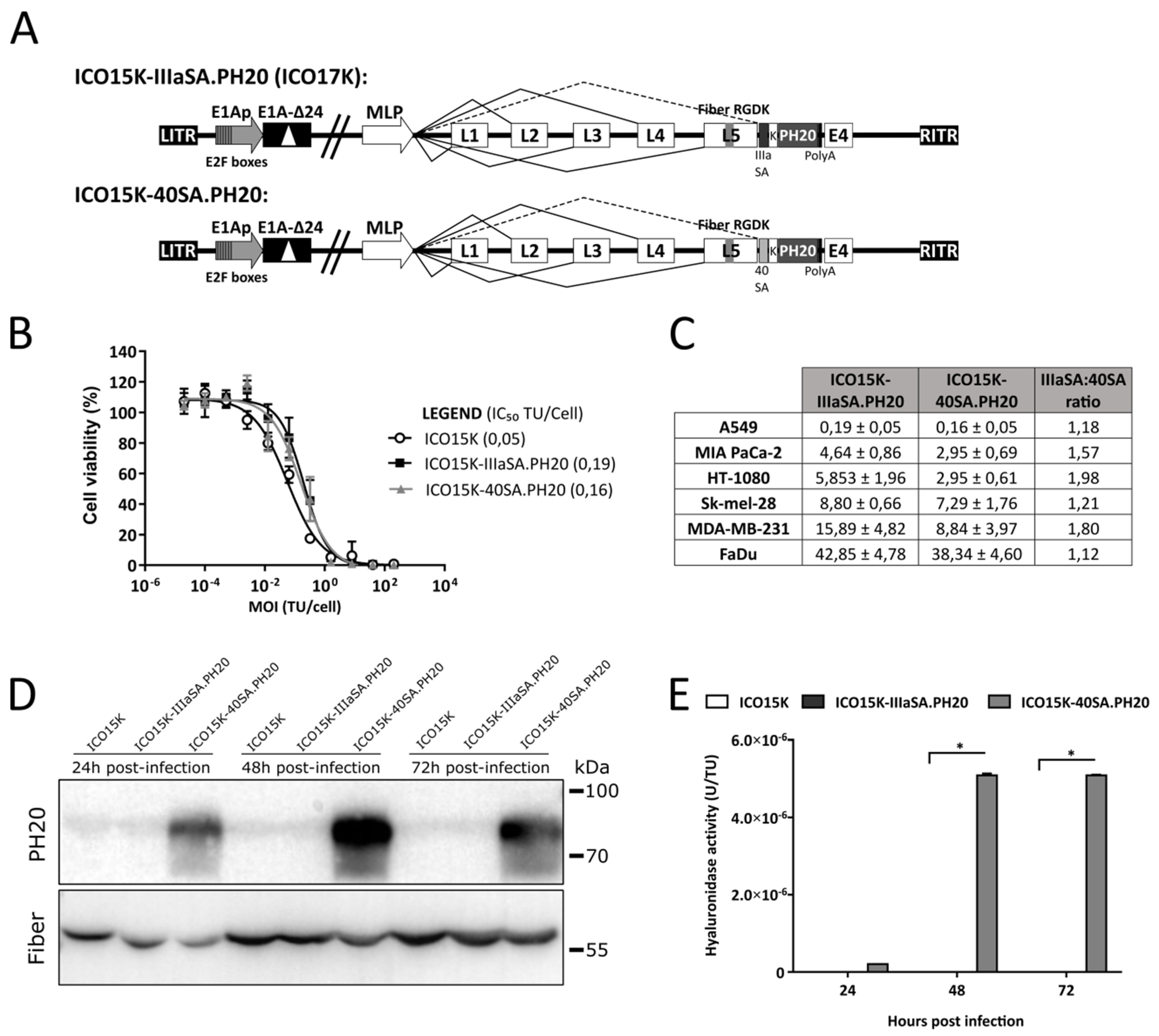
2.5. Generation and Characterization of Hyaluronidase-Expressing Oncolytic Adenoviruses
2.6. The 40SA Generated Higher Hyaluronidase Levels than IIIaSA
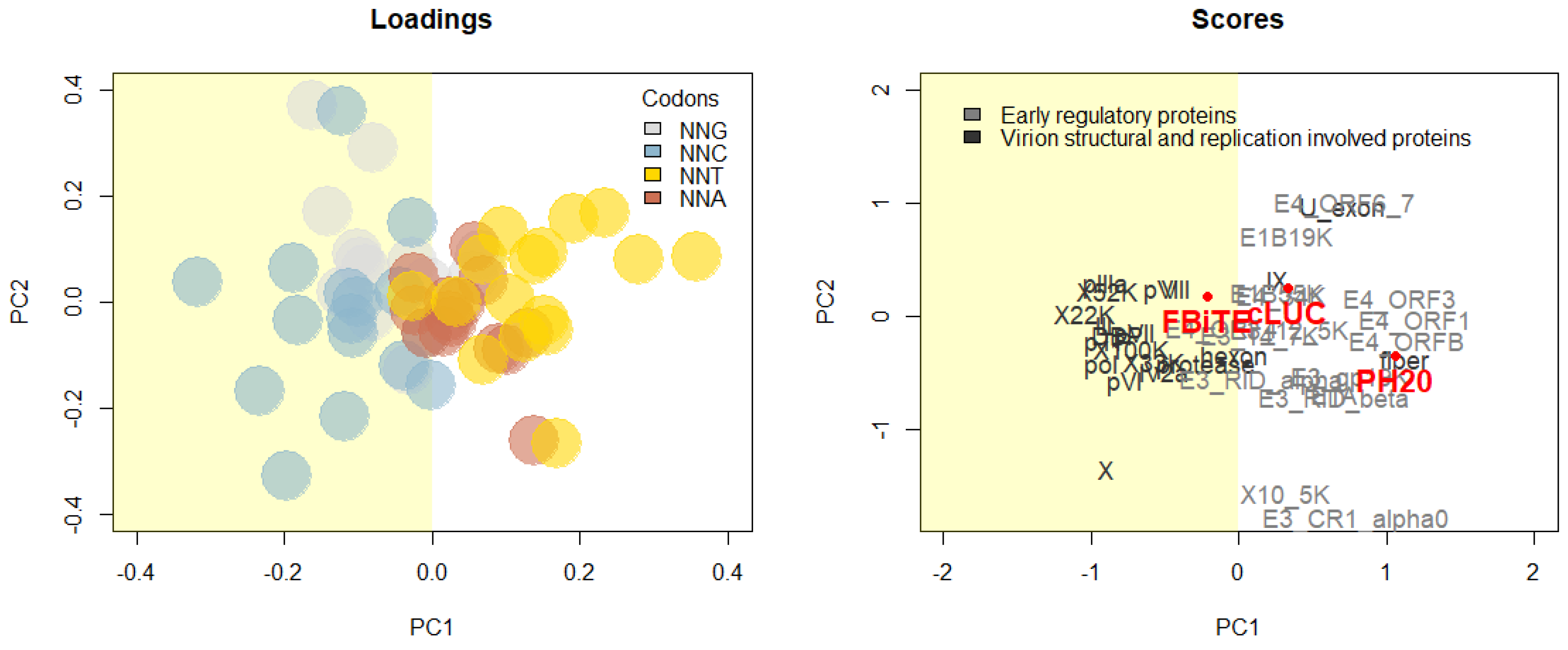
2.7. SA drives Oncolytic Acctivity in GC3-Rich Transgenes
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Isolation and Expansion of T Cells
4.3. Construction of Recombinant Adenoviruses
4.4. Virus Cytotoxicity Assays
4.5. Luciferase Reporter Assays
4.6. Production of Supernatants
4.7. Binding Assays
4.8. T-cell Activation Assay
4.9. Western Blot
4.10. Assay for Hyaluronidase Activity—Turbidimetric Assay
4.11. Codon Usage Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lang, F.F.; Conrad, C.; Gomez-Manzano, C.; Yung, W.A.; Sawaya, R.; Weinberg, J.S.; Prabhu, S.S.; Rao, G.; Fuller, G.N.; Aldape, K.D.; et al. Phase I study of DNX-2401 (Delta-24-RGD) oncolytic adenovirus: Replication and immunotherapeutic effects in recurrent malignant glioma. J. Clin. Oncol. 2018, 36, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.; Ruano, D.; Moreno, L.; Lassaletta, A.; Sirvent, F.J.B.; Andión, M.; Hernández, C.; Gonzalez, A.; Melen, G.; Alemany, R.; et al. First-in-child trial of celyvir (autologous mesenchymal stem cells carrying the oncolytic virus ICOVIR-5) in patients with relapsed and refractory pediatric solid tumors. J. Clin. Oncol. 2018, 36, 10543. [Google Scholar] [CrossRef]
- Hermiston, T.W.; Kuhn, I. Armed therapeutic viruses: Strategies and challenges to arming oncolytic viruses with therapeutic genes. Cancer Gene Ther. 2002, 9, 1022–1035. [Google Scholar] [CrossRef] [Green Version]
- Larsson, S.; Svensson, C.; Akusjärvi, G. Control of adenovirus major late gene expression at multiple levels. J. Mol. Boil. 1992, 225, 287–298. [Google Scholar] [CrossRef]
- Akusjärvi, G. Temporal regulation of adenovirus major late alternative RNA splicing. Front. Biosci. 2008, 13, 5006. [Google Scholar] [CrossRef]
- Bett, A.J.; Prevec, L.; Graham, F.L. Packaging capacity and stability of human adenovirus type 5 vectors. J. Virol. 1993, 67, 5911–5921. [Google Scholar] [CrossRef] [Green Version]
- Kelly, T.J.; Lewis, A.M. Use of nondefective adenovirus-simian virus 40 hybrids for mapping the simian virus 40 genome. J. Virol. 1973, 12, 643–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flint, S.J.; Wewerka-Lutz, Y.; Levine, A.S.; Sambrook, J.; Sharp, P.A., II. RNA sequences complementary to simian virus 40 and adenovirus 2 DNA in Ad2+NDr- and Ad2+ND3-Infected Cells. J. Virol. 1975, 16, 12. [Google Scholar] [CrossRef] [Green Version]
- Berkner, K.L.; Sharp, P.A. Generation of adenovirus by transfection of plasmids. Nucleic Acids Res. 1983, 11, 6003–6020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Halldén, G.; Hill, R.; Anand, A.; Liu, T.-C.; Francis, J.; Brooks, G.; Lemoine, N.; Kirn, D. E3 gene manipulations affect oncolytic adenovirus activity in immunocompetent tumor models. Nat. Biotechnol. 2003, 21, 1328–1335. [Google Scholar] [CrossRef]
- Bortolanza, S.; Buñuales, M.; Alzuguren, P.; Lamas, O.; Aldabe, R.; Prieto, J.; Hernandez-Alcoceba, R. Deletion of the E3-6.7K/gp19K region reduces the persistence of wild-type adenovirus in a permissive tumor model in Syrian hamsters. Cancer Gene Ther. 2009, 16, 703–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Alemany, R.; Yamamoto, M.; Curiel, D.T. The presence of the adenovirus E3 region improves the oncolytic potency of conditionally replicative adenoviruses. Clin. Cancer Res. 2002, 8, 3348–3359. [Google Scholar] [PubMed]
- Quirin, C.; Rohmer, S.; Fernández-Ulibarri, I.; Behr, M.; Hesse, A.; Engelhardt, S.; Erbs, P.; Enk, A.H.; Nettelbeck, D.M. Selectivity and efficiency of late transgene expression by transcriptionally targeted oncolytic adenoviruses are dependent on the transgene insertion strategy. Hum. Gene Ther. 2011, 22, 389–404. [Google Scholar] [CrossRef] [Green Version]
- Fuerer, C.; Iggo, R. 5-Fluorocytosine increases the toxicity of Wnt-targeting replicating adenoviruses that express cytosine deaminase as a late gene. Gene Ther. 2004, 11, 142–151. [Google Scholar] [CrossRef]
- Mühlemann, O.; Yue, B.-G.; Petersen-Mahrt, S.; Akusjärvi, G. A novel type of splicing enhancer regulating adenovirus Pre-mRNA splicing. Mol. Cell. Boil. 2000, 20, 2317–2325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Castro, J.; Martínez-Palacio, J.; Lillo, R.; García-Sánchez, F.; Alemany, R.; Madero, L.; Bueren, J.A.; Ramírez, M. Tumor cells as cellular vehicles to deliver gene therapies to metastatic tumors. Cancer Gene Ther. 2005, 12, 341–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carette, J.E.; Graat, H.C.A.; Schagen, F.H.E.; El Hassan, M.A.I.A.; Gerritsen, W.R.; Van Beusechem, V.W. Replication-dependent transgene expression from a conditionally replicating adenovirus via alternative splicing to a heterologous splice-acceptor site. J. Gene Med. 2005, 7, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Kretschmer, P.J.; Hermiston, T.W. Identification of novel insertion sites in the Ad5 genome that utilize the Ad splicing machinery for therapeutic gene expression. Mol. Ther. 2005, 12, 1052–1063. [Google Scholar] [CrossRef]
- Fernández-Ulibarri, I.; Hammer, K.; Arndt, M.A.; Kaufmann, J.K.; Dorer, D.; Engelhardt, S.; Kontermann, R.E.; Hess, J.; Allgayer, H.; Krauss, J.; et al. Genetic delivery of an immunoRNase by an oncolytic adenovirus enhances anticancer activity. Int. J. Cancer 2014, 136, 2228–2240. [Google Scholar] [CrossRef]
- Robinson, M.; Ge, Y.; Ko, D.; Yendluri, S.; Laflamme, G.; Hawkins, L.; Jooss, K. Comparison of the E3 and L3 regions for arming oncolytic adenoviruses to achieve a high level of tumor-specific transgene expression. Cancer Gene Ther. 2007, 15, 9–17. [Google Scholar] [CrossRef] [Green Version]
- De Sostoa, J.; Fajardo, C.A.; Moreno, R.; Ramos, M.D.; Farrera-Sal, M.; Alemany, R. Targeting the tumor stroma with an oncolytic adenovirus secreting a fibroblast activation protein-targeted bispecific T-cell engager. J. Immunother. Cancer 2019, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-García, A.; Gimenez-Alejandre, M.; Rojas, J.J.; Moreno, R.; Bazan-Peregrino, M.; Cascallo, M.; Alemany, R. Safety and efficacy of VCN-01, an oncolytic adenovirus combining fiber HSG-binding domain replacement with RGD and hyaluronidase expression. Clin. Cancer Res. 2014, 21, 1406–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, J.J.; Gimenez-Alejandre, M.; Gil-Hoyos, R.; Cascallo, M.; Alemany, R. Improved systemic antitumor therapy with oncolytic adenoviruses by replacing the fiber shaft HSG-binding domain with RGD. Gene Ther. 2011, 19, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Ostermann, E.; Garin-Chesa, P.; Heider, K.H.; Kalat, M.; Lamche, H.; Puri, C.; Kerjaschki, D.; Rettig, W.J.; Adolf, G.R. Effective immunoconjugate therapy in cancer models targeting a serine protease of tumor fibroblasts. Clin. Cancer Res. 2008, 14, 4584–4592. [Google Scholar] [CrossRef] [Green Version]
- Núñez-Manchón, E.; Farrera-Sal, M.; Castellano, G.; Medel, D.; Alemany, R.; Villanueva, E.; Fillat, C. Transgene codon usage drives viral fitness and therapeutic efficacy in oncolytic adenoviruses. bioRxiv 2020. [Google Scholar]
- Hawkins, L.K.; Johnson, L.; Bauzon, M.; Nye, J.A.; Castro, D.; Kitzes, G.A.; Young, M.D.; Holt, J.K.; Trown, P.; Hermiston, T.W. Gene delivery from the E3 region of replicating human adenovirus: Evaluation of the 6.7 K/gp19 K region. Gene Ther. 2001, 8, 1123–1131. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, L.; Hermiston, T. Gene delivery from the E3 region of replicating human adenovirus: Evaluation of the E3B region. Gene Ther. 2001, 8, 1142–1148. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, L.; Hermiston, T. Gene delivery from the E3 region of replicating human adenovirus: Evaluation of the ADP region. Gene Ther. 2001, 8, 1132–1141. [Google Scholar] [CrossRef] [Green Version]
- Kretschmer, P.J.; Jin, F.; Chartier, C.; Hermiston, T.W. Development of a transposon-based approach for identifying novel transgene insertion sites within the replicating adenovirus. Mol. Ther. 2005, 12, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Funston, G.M.; Kallioinen, S.E.; De Felipe, P.; Ryan, M.D.; Iggo, R.D. Expression of heterologous genes in oncolytic adenoviruses using picornaviral 2A sequences that trigger ribosome skipping. J. Gen. Virol. 2008, 89, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, E.; Marti, M.; Fillat, C. Codon optimization of the adenoviral fiber negatively impacts structural protein expression and viral fitness. Sci. Rep. 2016, 6, 27546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanton, R.J.; McSharry, B.; Armstrong, M.; Tomasec, P.; Wilkinson, G.W.G. Re-engineering adenovirus vector systems to enable high-throughput analyses of gene function. BioTechniques 2008, 45, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Puig-Saus, C.; Rojas, L.A.; Laborda, E.; Figueras, A.; Alba, R.; Fillat, C.; Alemany, R. iRGD tumor-penetrating peptide-modified oncolytic adenovirus shows enhanced tumor transduction, intratumoral dissemination and antitumor efficacy. Gene Ther. 2014, 21, 767–774. [Google Scholar] [CrossRef] [Green Version]
- Gros, A.; Martinez-Quintanilla, J.; Puig, C.; Guedan, S.; Mollevi, D.G.; Alemany, R.; Cascallo, M. Bioselection of a gain of function mutation that enhances adenovirus 5 release and improves its antitumoral potency. Cancer Res. 2008, 68, 8928–8937. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cells Per Well | Initial MOI (Serial Dilution) | |
|---|---|---|
| A549 | 30,000 | 200 (1/5) |
| Sk-mel-28 | 20,000 | 200 (1/5) |
| MIA PaCa-2 | 30,000 | 200 (1/3) |
| FaDU | 20,000 | 600 (1/3) |
| HT-1080 | 20,000 | 600 (1/3) |
| MDA-MB-231 | 15,000 | 400 (1/3) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farrera-Sal, M.; de Sostoa, J.; Nuñez-Manchón, E.; Moreno, R.; Fillat, C.; Bazan-Peregrino, M.; Alemany, R. Arming Oncolytic Adenoviruses: Effect of Insertion Site and Splice Acceptor on Transgene Expression and Viral Fitness. Int. J. Mol. Sci. 2020, 21, 5158. https://doi.org/10.3390/ijms21145158
Farrera-Sal M, de Sostoa J, Nuñez-Manchón E, Moreno R, Fillat C, Bazan-Peregrino M, Alemany R. Arming Oncolytic Adenoviruses: Effect of Insertion Site and Splice Acceptor on Transgene Expression and Viral Fitness. International Journal of Molecular Sciences. 2020; 21(14):5158. https://doi.org/10.3390/ijms21145158
Chicago/Turabian StyleFarrera-Sal, Martí, Jana de Sostoa, Estela Nuñez-Manchón, Rafael Moreno, Cristina Fillat, Miriam Bazan-Peregrino, and Ramon Alemany. 2020. "Arming Oncolytic Adenoviruses: Effect of Insertion Site and Splice Acceptor on Transgene Expression and Viral Fitness" International Journal of Molecular Sciences 21, no. 14: 5158. https://doi.org/10.3390/ijms21145158
APA StyleFarrera-Sal, M., de Sostoa, J., Nuñez-Manchón, E., Moreno, R., Fillat, C., Bazan-Peregrino, M., & Alemany, R. (2020). Arming Oncolytic Adenoviruses: Effect of Insertion Site and Splice Acceptor on Transgene Expression and Viral Fitness. International Journal of Molecular Sciences, 21(14), 5158. https://doi.org/10.3390/ijms21145158