Arginase as a Potential Biomarker of Disease Progression: A Molecular Imaging Perspective

 , ,
, ,
Abstract
:1. Introduction
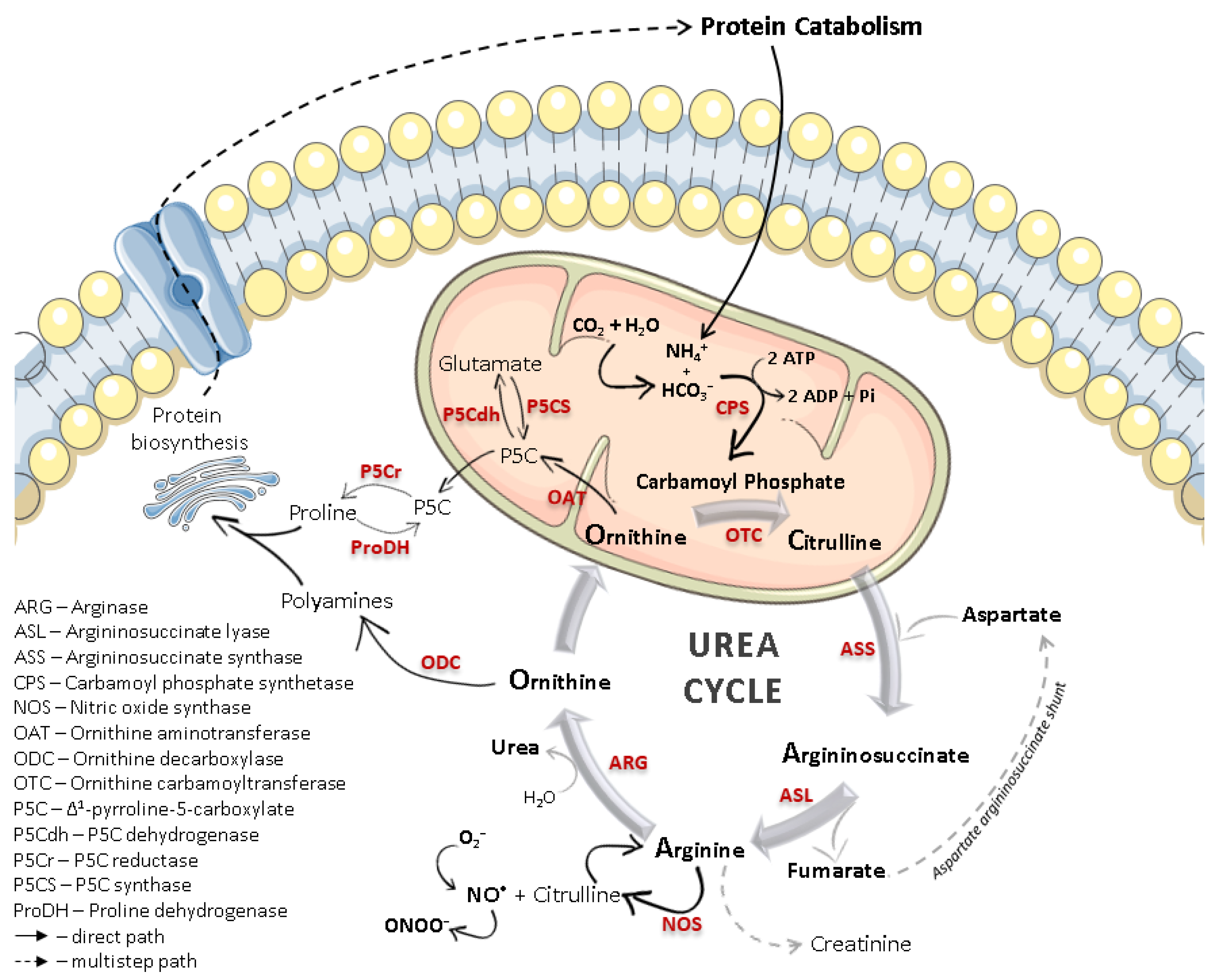
1.1. Arginase Isoforms
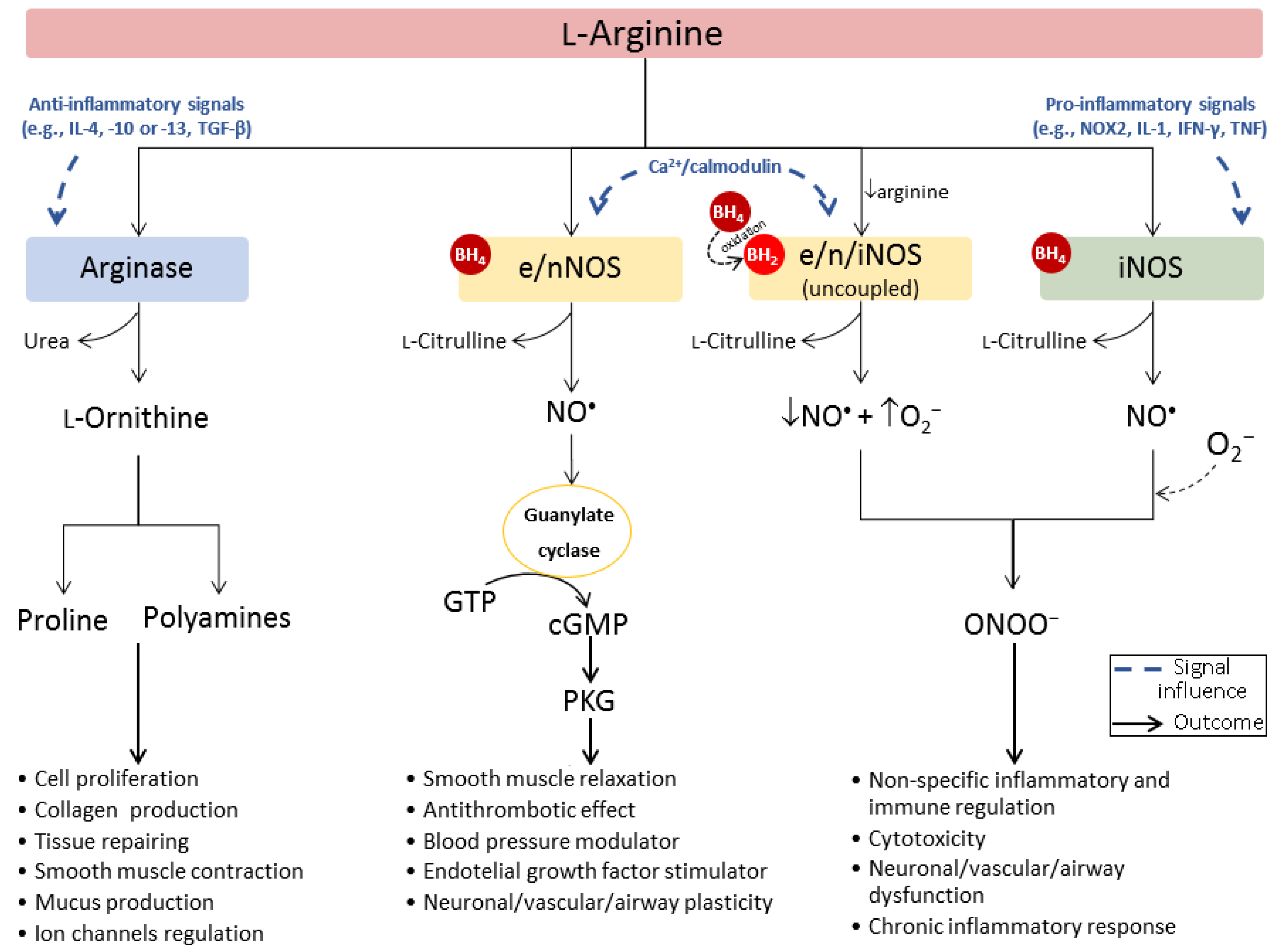
1.2. Arginase/Nitric Oxide Synthase (Patho)Physiological Interplay
2. The Pathophysiological Role of Arginase
2.1. Immune System Cells
2.2. Cardiovascular Endothelium
2.3. Neuronal Cells
2.4. Overview of the Pathologies Related to Arginase Deregulation
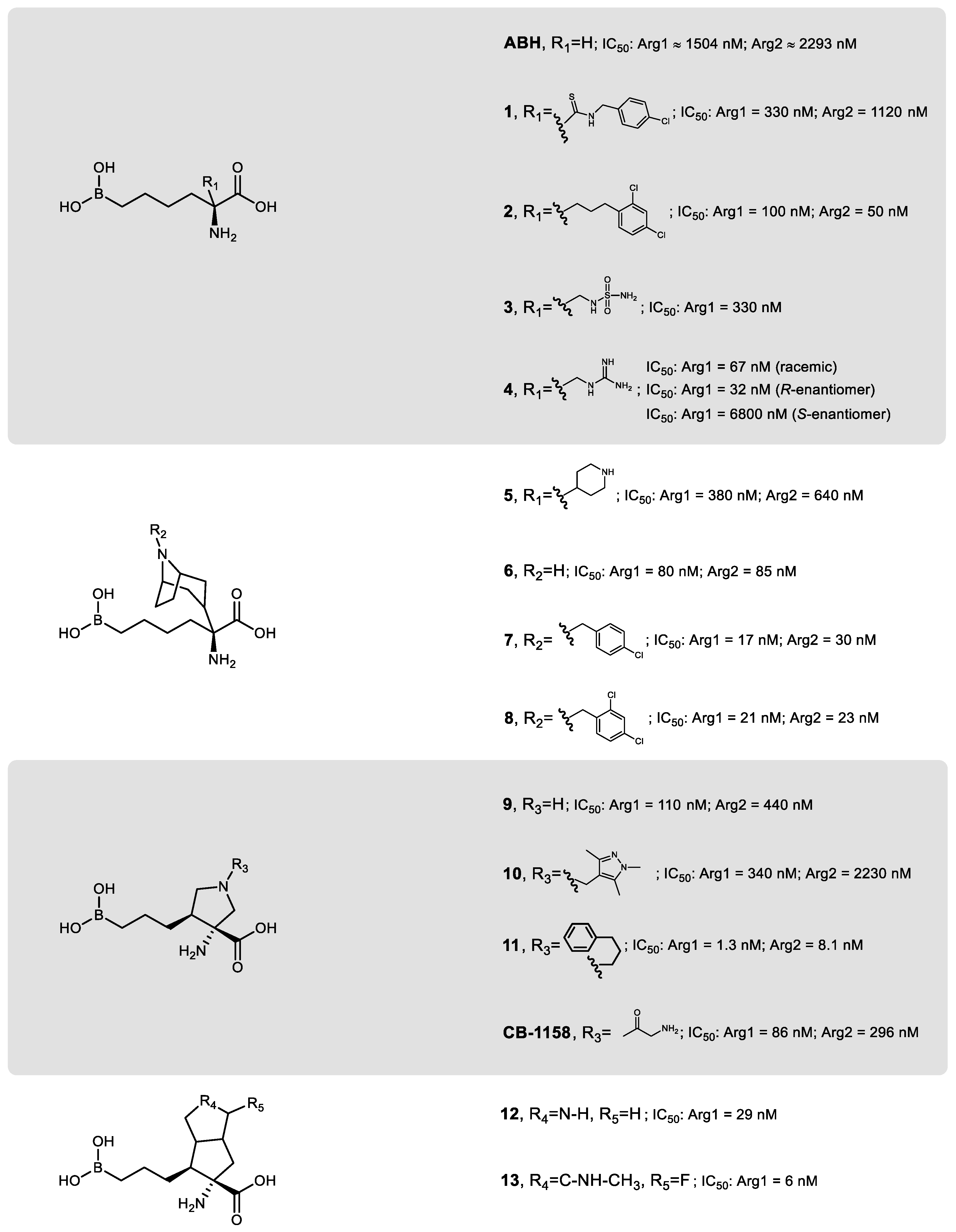
3. Development of Arginase Inhibitors
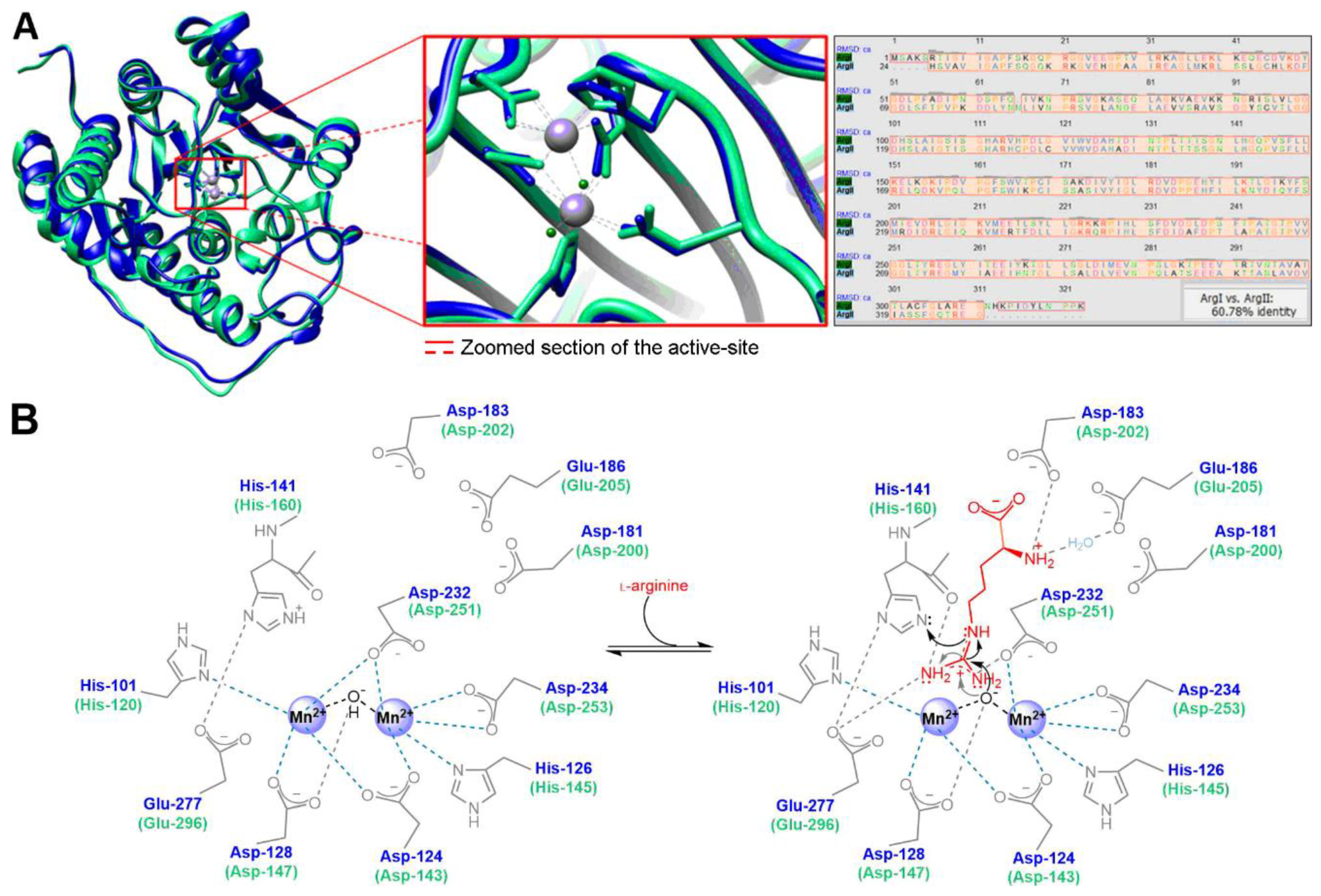
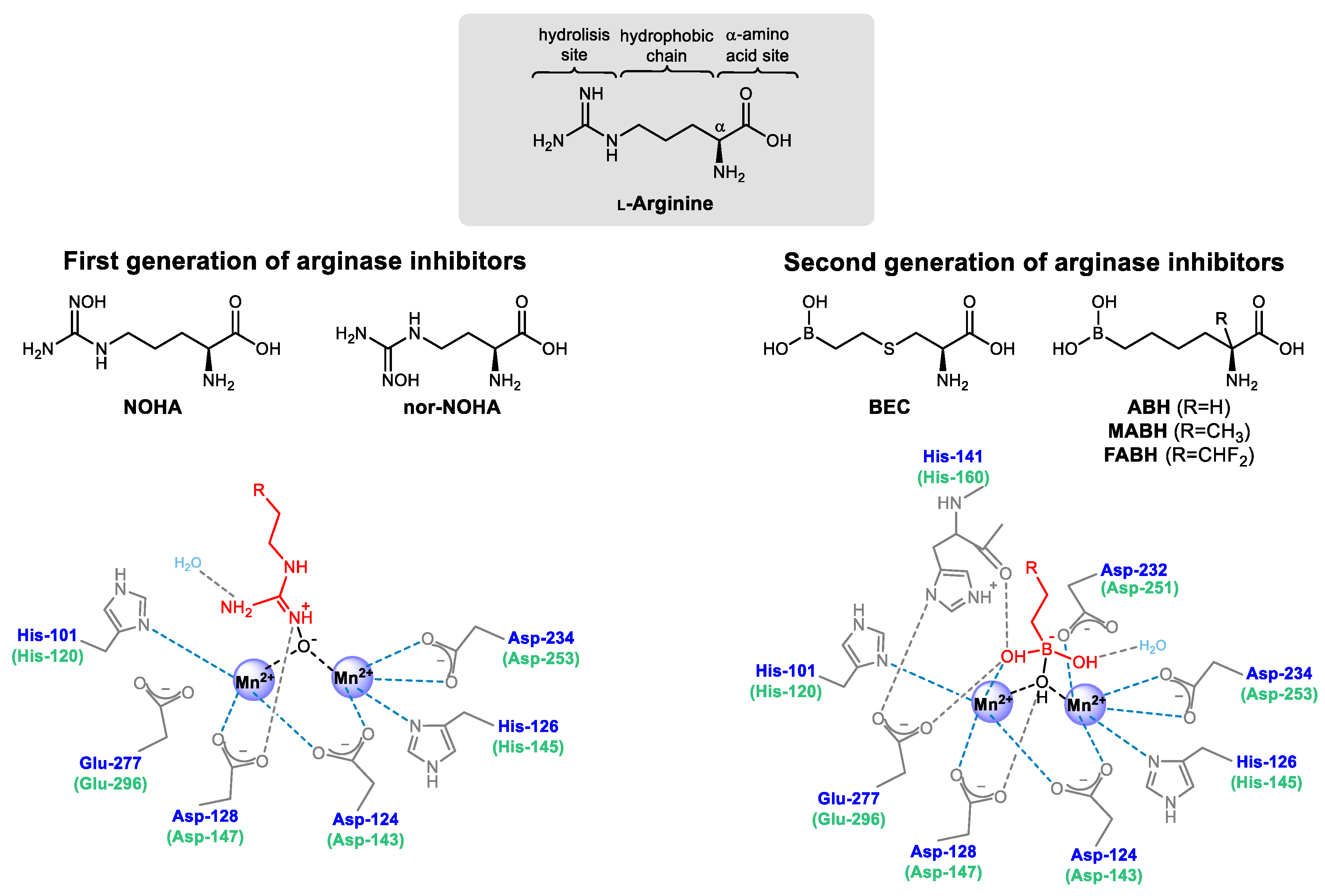
3.1. Arginase Inhibitors from the First and Second Generation
3.2. Third Generation of Arginase Inhibitors
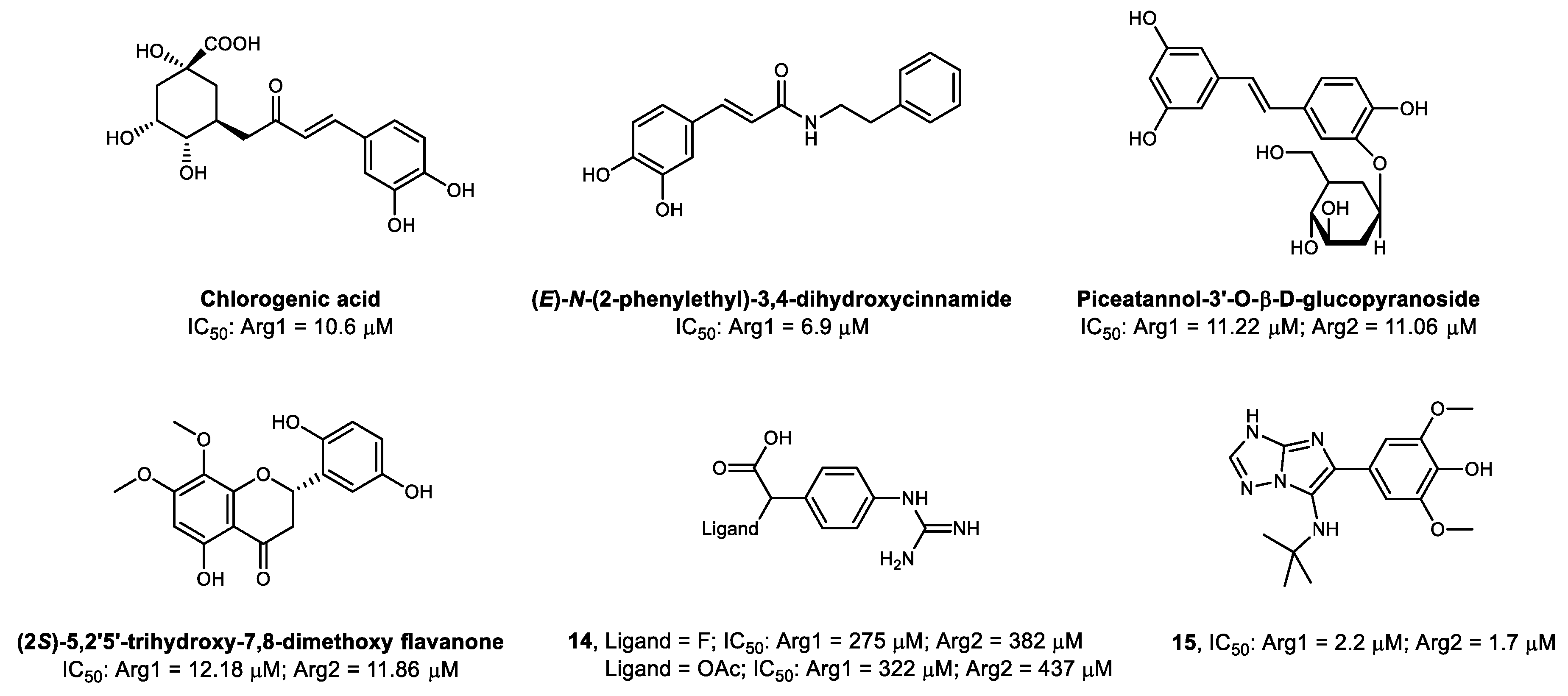
3.3. Non-Amino Acid-Based Arginase Inhibitors
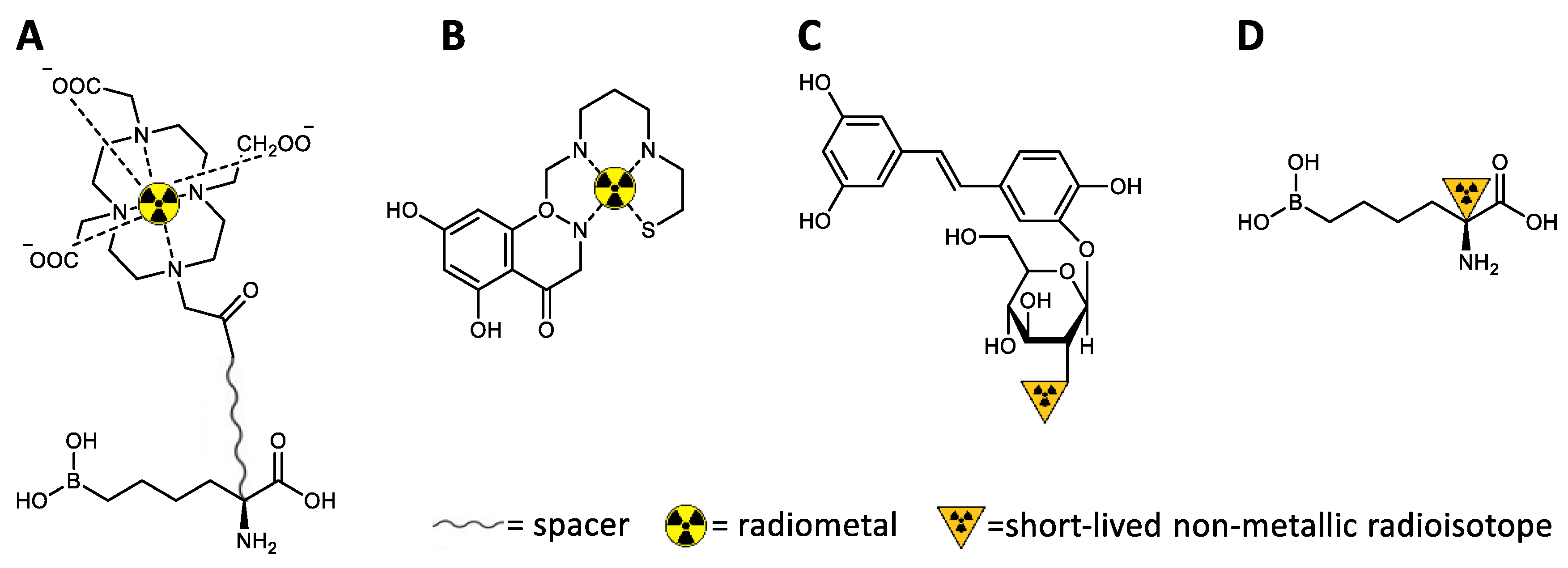
4. Molecular Imaging of Arginase
4.1. Molecular Imaging Modalities
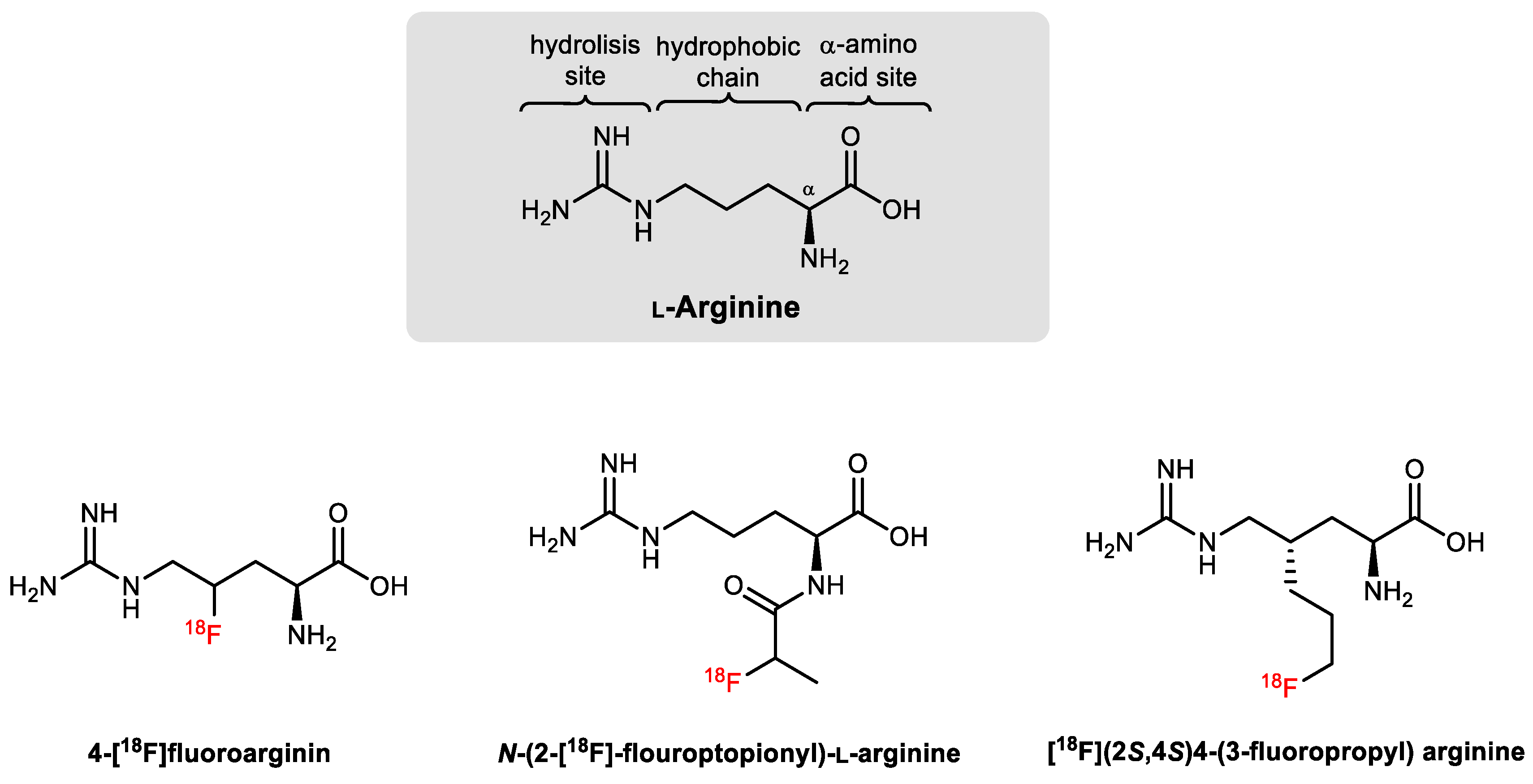
4.2. Development of Arginase-Targeted Radiotracers for Nuclear Imaging
4.3. Future Perspectives for Arginase-Directed Radiotracers
5. Conclusions
Funding
Conflicts of Interest
Abbreviations
| ABH | 2-(S)-Amino-6-boronohexanoic acid |
| Arg1 | Arginase type I |
| Arg2 | Arginase type II |
| ASS | Argininosuccinate synthase |
| BEC | S-(2-Boronoethyl)-L-cysteine |
| cAMP | Cyclic adenosine monophosphates |
| COX | Cyclooxygenase |
| CT | Computed tomography |
| FABH | 2-Amino-6-borono-2-(difluoromethyl)hexanoic acid |
| IC50 | Half-maximal inhibitory concentration |
| IFN | Interferon |
| IgE | Immunoglobulin E |
| IL | Interleukin |
| ki | Inhibitory constant |
| Km | Michaelis-Menten kinetics |
| MABH | 2-Amino-6-borono-2-methylhexanoic acid |
| MRI | Magnetic resonance imaging |
| NOHA | Nω-hydroxy-L-arginine |
| NOS | Nitric oxide synthase |
| NOX | Nicotinamide adenine dinucleotide phosphate oxidase |
| OTC | Ornithine carbamoyltransferase |
| PET | Positron emission tomography |
| SPECT | Single-photon emission computed tomography |
| STAT3 | Signal transducer and activator of transcription 3 |
| TGF | Transforming growth factor |
| TNF | Tumor necrosis factor |
| Vmax | Maximum rate of reaction |
References
- Hunter, A.; Dauphinee, J.A.; Dale, H.H. Quantitative studies concerning the distribution of arginase in fishes and other animals. Proc. R. Soc. B 1924, 97, 227. [Google Scholar]
- Ratner, S.; Petrack, B. The mechanism of arginine synthesis from citrulline in kidney. J. Biol. Chem. 1953, 200, 175. [Google Scholar] [PubMed]
- Mezl, V.A.; Knox, W.E. Metabolism of arginine in lactating rat mammary gland. Biochem. J. 1977, 166, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Scott, E.J. Arginase Activity in Human Skin. Science 1951, 113, 601. [Google Scholar] [CrossRef] [PubMed]
- Emmanuel, B. Urea cycle enzymes in tissues (liver, rumen epithelium, heart, kidney, lung and spleen) of sheep (Ovis aries). Comp. Biochem. Physiol. B 1980, 65, 693. [Google Scholar] [CrossRef]
- Spector, E.B.; Rice, S.C.H.; Cederbaum, S.D. Immunologic Studies of Arginase in Tissues of Normal Human Adult and Arginase-Deficient Patients. Pediatr. Res. 1983, 17, 941. [Google Scholar] [CrossRef] [Green Version]
- Human Protein Atlas. Available online: http://www.proteinatlas.org (accessed on 29 April 2020).
- Munder, M. Arginase: An emerging key player in the mammalian immune system. Br. J. Pharmacol. 2009, 158, 638. [Google Scholar] [CrossRef] [Green Version]
- Cama, E.; Colleluori, D.M.; Emig, F.A.; Shin, H.; Kim, S.W.; Kim, N.N.; Traish, A.M.; Ash, D.E.; Christianson, D.W. Human Arginase II: Crystal Structure and Physiological Role in Male and Female Sexual Arousal. Biochemistry 2003, 42, 8445. [Google Scholar] [CrossRef]
- Di Costanzo, L.; Sabio, G.; Mora, A.; Rodriguez, P.C.; Ochoa, A.C.; Centeno, F.; Christianson, D.W. Crystal structure of human arginase I at 1.29-Å resolution and exploration of inhibition in the immune response. Proc. Natl. Acad. Sci. USA 2005, 102, 13058. [Google Scholar] [CrossRef] [Green Version]
- Kanyo, Z.F.; Scolnick, L.R.; Ash, D.E.; Christianson, D.W. Structure of a unique binuclear manganese cluster in arginase. Nature 1996, 383, 554. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera---A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Costanzo, L.; Pique, M.E.; Christianson, D.W. Crystal Structure of Human Arginase I Complexed with Thiosemicarbazide Reveals an Unusual Thiocarbonyl μ-Sulfide Ligand in the Binuclear Manganese Cluster. J. Am. Chem. Soc. 2007, 129, 6388. [Google Scholar] [CrossRef] [Green Version]
- Nagagarajan, S.; Xue, F.; MacKerell, A.D. Impact of Substrate Protonation and Tautomerization States on Interactions with the Active Site of Arginase I. J. Chem. Inf. Model. 2013, 53, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, J.D.; Cama, E.; Colleluori, D.M.; Pethe, S.; Boucher, J.-L.; Mansuy, D.; Ash, D.E.; Christianson, D.W. Mechanistic and Metabolic Inferences from the Binding of Substrate Analogues and Products to Arginase. Biochemistry 2001, 40, 2689. [Google Scholar] [CrossRef] [PubMed]
- Tommasi, S.; Elliot, D.J.; Da Boit, M.; Gray, S.R.; Lewis, B.C.; Mangoni, A.A. Homoarginine and inhibition of human arginase activity: Kinetic characterization and biological relevance. Sci. Rep. 2018, 8, 3697. [Google Scholar] [CrossRef] [PubMed]
- Di Costanzo, L.; Moulin, M.; Haertlein, M.; Meilleur, F.; Christianson, D.W. Expression, purification, assay, and crystal structure of perdeuterated human arginase I. Arch. Biochem. Biophys. 2007, 465, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowling, D.P.; Di Costanzo, L.; Gennadios, H.A.; Christianson, D.W. Evolution of the arginase fold and functional diversity. Cell Mol. Life Sci. 2008, 65, 2039. [Google Scholar] [CrossRef] [Green Version]
- Minemura, M.; Tajiri, K.; Shimizu, Y. Systemic abnormalities in liver disease. World J. Gastroenterol. 2009, 15, 2960. [Google Scholar] [CrossRef]
- Bredt, D.S.; Snyder, S.H. Isolation of nitric oxide synthetase, a calmodulin-requiring enzyme. Proc. Natl. Acad. Sci. USA 1990, 87, 682. [Google Scholar] [CrossRef] [Green Version]
- Pollock, J.S.; Förstermann, U.; Mitchell, J.A.; Warner, T.D.; Schmidt, H.H.; Nakane, M.; Murad, F. Purification and characterization of particulate endothelium-derived relaxing factor synthase from cultured and native bovine aortic endothelial cells. Proc. Natl. Acad. Sci. USA 1991, 88, 10480. [Google Scholar] [CrossRef] [Green Version]
- Hevel, J.M.; White, K.A.; Marletta, M.A. Purification of the inducible murine macrophage nitric oxide synthase. Identification as a flavoprotein. J. Biol. Chem. 1991, 266, 22789. [Google Scholar] [PubMed]
- Mori, M. Regulation of Nitric Oxide Synthesis and Apoptosis by Arginase and Arginine Recycling. J. Nutr. 2007, 137, 1616S. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Morris, S.M., Jr. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336, 1. [Google Scholar] [CrossRef] [PubMed]
- Berkowitz, D.E.; White, R.; Li, D.; Minhas, K.M.; Cernetich, A.; Kim, S.; Burke, S.; Shoukas, A.A.; Nyhan, D.; Champion, H.C.; et al. Arginase Reciprocally Regulates Nitric Oxide Synthase Activity and Contributes to Endothelial Dysfunction in Aging Blood Vessels. Circulation 2003, 108, 2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roe, N.D.; Ren, J. Nitric oxide synthase uncoupling: A therapeutic target in cardiovascular diseases. Vascul. Pharmacol. 2012, 57, 168. [Google Scholar] [CrossRef] [PubMed]
- Moncada, S.; Higgs, E.A. The discovery of nitric oxide and its role in vascular biology. Br. J. Pharmacol. 2006, 147, S193. [Google Scholar] [CrossRef] [Green Version]
- Kuwabara, M.; Kakinuma, Y.; Ando, M.; Katare, R.G.; Yamasaki, F.; Doi, Y.; Sato, T. Nitric Oxide Stimulates Vascular Endothelial Growth Factor Production in Cardiomyocytes Involved in Angiogenesis. J. Physiol. Sci. 2006, 56, 95. [Google Scholar] [CrossRef] [Green Version]
- Liu, V.W.T.; Huang, P.L. Cardiovascular roles of nitric oxide: A review of insights from nitric oxide synthase gene disrupted mice. Cardiovasc. Res. 2008, 77, 19. [Google Scholar] [CrossRef] [Green Version]
- Esplugues, J.V. NO as a signalling molecule in the nervous system. Br. J. Pharmacol. 2002, 135, 1079. [Google Scholar] [CrossRef] [Green Version]
- Casado, M.; MJ, D.i.-G.; Rodrigo, J.; Fernández, A.P.; Boscá, L.; Martín-Sanz, P. Expression of the calcium-independent cytokine-inducible (iNOS) isoform of nitric oxide synthase in rat placenta. Biochem. J. 1997, 324, 201. [Google Scholar] [CrossRef] [Green Version]
- Satoh, M.; Nakamura, M.; Tamura, G.; Makita, S.; Segawa, I.; Tashiro, A.; Satodate, R.; Hiramori, K. Inducible nitric oxide synthase and tumor necrosis factor-alpha in myocardium in human dilated cardiomyopathy. J. Am. Coll. Cardiol. 1997, 29, 716. [Google Scholar] [CrossRef] [Green Version]
- Teng, X.; Zhang, H.; Snead, C.; Catravas, J.D. Molecular mechanisms of iNOS induction by IL-1 beta and IFN-gamma in rat aortic smooth muscle cells. Am. J. Physiol. Cell Physiol. 2002, 282, C144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Tyml, K.; Wilson, J.X. iNOS expression requires NADPH oxidase-dependent redox signaling in microvascular endothelial cells. J. Cell Physiol. 2008, 217, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wink, D.A.; Hines, H.B.; Cheng, R.Y.S.; Switzer, C.H.; Flores-Santana, W.; Vitek, M.P.; Ridnour, L.A.; Colton, C.A. Nitric oxide and redox mechanisms in the immune response. J. Leukoc. Biol. 2011, 89, 873. [Google Scholar] [CrossRef] [Green Version]
- Bronte, V.; Zanovello, P. Regulation of immune responses by L-arginine metabolism. Nat. Rev. Immunol. 2005, 5, 641. [Google Scholar] [CrossRef]
- Bauer, P.M.; Buga, G.M.; Fukuto, J.M.; Pegg, A.E.; Ignarro, L.J. Nitric Oxide Inhibits Ornithine Decarboxylase viaS-Nitrosylation of Cysteine 360 in the Active Site of the Enzyme. J. Biol. Chem. 2001, 276, 34458. [Google Scholar] [CrossRef] [Green Version]
- Bussière, F.I.; Chaturvedi, R.; Cheng, Y.; Gobert, A.P.; Asim, M.; Blumberg, D.R.; Xu, H.; Kim, P.Y.; Hacker, A.; Casero, R.A.; et al. Spermine Causes Loss of Innate Immune Response to Helicobacter pylori by Inhibition of Inducible Nitric-oxide Synthase Translation. J. Biol. Chem. 2005, 280, 2409. [Google Scholar] [CrossRef] [Green Version]
- Boucher, J.L.; Custot, J.; Vadon, S.; Delaforge, M.; Lepoivre, M.; Tenu, J.P.; Yapo, A.; Mansuy, D. Nω-Hydroxy-L-Arginine, an Intermediate in the L-Arginine to Nitric Oxide Pathway, Is a Strong Inhibitor of Liver and Macrophage Arginase. Biochem. Biophys. Res. Commun. 1994, 203, 1614. [Google Scholar] [CrossRef]
- Durante, W. Role of arginase in vessel wall remodeling. Front. Immunol. 2013, 4, 111. [Google Scholar] [CrossRef] [Green Version]
- Wiesinger, H. Arginine metabolism and the synthesis of nitric oxide in the nervous system. Prog. Neurobiol. 2001, 64, 365. [Google Scholar] [CrossRef]
- Chang, C.-I.; Liao, J.C.; Kuo, L. Arginase modulates nitric oxide production in activated macrophages. Am. J. Physiol. Heart Circ. Physiol. 1998, 274, H342. [Google Scholar] [CrossRef]
- Tötemeyer, S.; Sheppard, M.; Lloyd, A.; Roper, D.; Dowson, C.; Underhill, D.; Murray, P.; Maskell, D.; Bryant, C. IFN-γ Enhances Production of Nitric Oxide from Macrophages via a Mechanism That Depends on Nucleotide Oligomerization Domain-2. J. Immunol. 2006, 176, 4804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, E.; Christopoulos, P.F.; Halder, S.; Lunde, A.; Beraki, K.; Speth, M.; Øynebråten, I.; Corthay, A. Toll-Like Receptor Ligands and Interferon-γ Synergize for Induction of Antitumor M1 Macrophages. Front. Immunol. 2017, 8, 1383. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.M., Jr. Arginine Metabolism Revisited. J. Nutr. 2016, 146, 2579S. [Google Scholar] [CrossRef] [PubMed]
- Louis, C.A.; Mody, V.; William, L.; Henry, J.; Reichner, J.S.; Albina, J.E. Regulation of arginase isoforms I and II by IL-4 in cultured murine peritoneal macrophages. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1999, 276, R237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, P.C.; Hernandez, C.P.; Quiceno, D.; Dubinett, S.M.; Zabaleta, J.; Ochoa, J.B.; Gilbert, J.; Ochoa, A.C. Arginase I in myeloid suppressor cells is induced by COX-2 in lung carcinoma. J. Exp. Med. 2005, 202, 931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Wang, H.; Wang, X.; Jiang, G.; Liu, H.; Zhang, G.; Wang, H.; Fang, R.; Bu, X.; Cai, S.; et al. TGF-β induces M2-like macrophage polarization via SNAIL-mediated suppression of a pro-inflammatory phenotype. Oncotarget 2016, 7, 52294. [Google Scholar] [CrossRef] [Green Version]
- Gotoh, T.; Mori, M. Arginase II downregulates nitric oxide (NO) production and prevents NO-mediated apoptosis in murine macrophage-derived RAW 264.7 cells. J. Cell Biol. 1999, 144, 427. [Google Scholar] [CrossRef] [Green Version]
- Ley, K. M1 Means Kill; M2 Means Heal. J. Immunol. 2017, 199, 2191. [Google Scholar] [CrossRef]
- Mieulet, V.; Yan, L.; Choisy, C.; Sully, K.; Procter, J.; Kouroumalis, A.; Krywawych, S.; Pende, M.; Ley, S.C.; Moinard, C.; et al. TPL-2–Mediated Activation of MAPK Downstream of TLR4 Signaling Is Coupled to Arginine Availability. Sci. Signal. 2010, 3, ra61. [Google Scholar] [CrossRef]
- Erdely, A.; Kepka-Lenhart, D.; Salmen-Muniz, R.; Chapman, R.; Hulderman, T.; Kashon, M.; Simeonova, P.P.; Morris, S.M., Jr. Arginase activities and global arginine bioavailability in wild-type and ApoE-deficient mice: Responses to high fat and high cholesterol diets. PLoS ONE 2010, 5, e15253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Higgins, C.B.; Fortune, H.M.; Chen, P.; Stothard, A.I.; Mayer, A.L.; Swarts, B.M.; DeBosch, B.J. Hepatic arginase 2 (Arg2) is sufficient to convey the therapeutic metabolic effects of fasting. Nat. Commun. 2019, 10, 1587. [Google Scholar] [CrossRef] [PubMed]
- Lange, P.S.; Langley, B.; Lu, P.; Ratan, R.R. Novel Roles for Arginase in Cell Survival, Regeneration, and Translation in the Central Nervous System. J. Nutr. 2004, 134, 2812S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Fleete, M.S.; Jing, Y.; Collie, N.D.; Curtis, M.A.; Waldvogel, H.J.; Faull, R.L.M.; Abraham, W.C.; Zhang, H. Altered arginine metabolism in Alzheimer’s disease brains. Neurobiol. Aging 2014, 35, 1992. [Google Scholar] [CrossRef]
- Caldwell, R.B.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.W. Arginase: An old enzyme with new tricks. Trends Pharmacol. Sci. 2015, 36, 395. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, R.W.; Rodriguez, P.C.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.B. Arginase: A Multifaceted Enzyme Important in Health and Disease. Physiol. Rev. 2018, 98, 641. [Google Scholar] [CrossRef] [Green Version]
- Chandra, S.; Fulton, D.J.R.; Caldwell, R.B.; Caldwell, R.W.; Toque, H.A. Hyperglycemia-impaired aortic vasorelaxation mediated through arginase elevation: Role of stress kinase pathways. Eur. J. Pharmacol. 2019, 844, 26. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, J.; Qu, D.; Wang, L.; Wong, C.M.; Lau, C.-W.; Huang, Y.; Wang, Y.F.; Huang, H.; Xia, Y.; et al. Serum exosomes mediate delivery of arginase 1 as a novel mechanism for endothelial dysfunction in diabetes. Proc. Natl. Acad. Sci. USA 2018, 115, E6927. [Google Scholar] [CrossRef] [Green Version]
- Rojas, M.; Lemtalsi, T.; Toque, H.A.; Xu, Z.; Fulton, D.; Caldwell, R.W.; Caldwell, R.B. NOX2-Induced Activation of Arginase and Diabetes-Induced Retinal Endothelial Cell Senescence. Antioxidants 2017, 6, 43. [Google Scholar] [CrossRef] [Green Version]
- Bhatta, A.; Yao, L.; Xu, Z.; Toque, H.A.; Chen, J.; Atawia, R.T.; Fouda, A.Y.; Bagi, Z.; Lucas, R.; Caldwell, R.B.; et al. Obesity-induced vascular dysfunction and arterial stiffening requires endothelial cell arginase 1. Cardiovasc. Res. 2017, 113, 1664. [Google Scholar] [CrossRef]
- Bhatta, A.; Yao, L.; Toque, H.A.; Shatanawi, A.; Xu, Z.; Caldwell, R.B.; Caldwell, R.W. Angiotensin II-Induced Arterial Thickening, Fibrosis and Stiffening Involves Elevated Arginase Function. PLoS ONE 2015, 10, e0121727. [Google Scholar]
- Peyton, K.J.; Liu, X.-M.; Shebib, A.R.; Johnson, F.K.; Johnson, R.A.; Durante, W. Arginase inhibition prevents the development of hypertension and improves insulin resistance in obese rats. Amino Acids 2018, 50, 747. [Google Scholar] [CrossRef] [PubMed]
- Lasch, M.; Caballero-Martinez, A.; Troidl, K.; Schloegl, I.; Lautz, T.; Deindl, E. Arginase inhibition attenuates arteriogenesis and interferes with M2 macrophage accumulation. Lab. Investig. 2016, 96, 830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.; Goetsch, S.C.; Wang, Z.; Luo, R.; Hill, J.A.; Schneider, J.; Morris, S.M., Jr.; Liu, Z.-P. FoxO4 promotes early inflammatory response upon myocardial infarction via endothelial Arg1. Circ. Res. 2015, 117, 967. [Google Scholar] [CrossRef]
- Lacchini, R.; Muniz, J.J.; Nobre, Y.T.D.A.; Cologna, A.J.; Martins, A.C.P.; Tanus-Santos, J.E. Relationship between Arginase 1 and Arginase 2 levels and genetic polymorphisms with erectile dysfunction. Nitric Oxide Biol. Chem. 2015, 51, 36. [Google Scholar] [CrossRef]
- Henno, P.; Maurey, C.; Le Pimpec-Barthes, F.; Devillier, P.; Delclaux, C.; Israël-Biet, D. Is arginase a potential drug target in tobacco-induced pulmonary endothelial dysfunction? Respir. Res. 2015, 16, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, J.; Nelin, L.D.; Chen, B. Hypoxia induces arginase II expression and increases viable human pulmonary artery smooth muscle cell numbers via AMPKα(1) signaling. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 312, L568. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.; XiangLi, X.; Niu, H.; Wang, H.; Jia, P.; Gong, W.; Wu, D.; Qin, W.; Xing, C. Arginase inhibitor attenuates pulmonary artery hypertension induced by hypoxia. Mol. Cell Biochem. 2016, 412, 91. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Zhang, J.-H.; Chen, X.-X.; Ren, H.-L.; Feng, X.-L.; Wang, J.-L.; Xiao, J.-H. Combination of L-Arginine and L-Norvaline protects against pulmonary fibrosis progression induced by bleomycin in mice. Biomed. Pharm. 2019, 113, 108768. [Google Scholar] [CrossRef]
- Wu, Y.-S.; Jiang, J.; Ahmadi, S.; Lew, A.; Laselva, O.; Xia, S.; Bartlett, C.; Ip, W.; Wellhauser, L.; Ouyang, H.; et al. ORKAMBI-Mediated Rescue of Mucociliary Clearance in Cystic Fibrosis Primary Respiratory Cultures Is Enhanced by Arginine Uptake, Arginase Inhibition, and Promotion of Nitric Oxide Signaling to the Cystic Fibrosis Transmembrane Conductance Regulator Channel. Mol. Pharm. 2019, 96, 515. [Google Scholar] [CrossRef]
- Lucca, F.; Da Dalt, L.; Ros, M.; Gucciardi, A.; Pirillo, P.; Naturale, M.; Perilongo, G.; Giordano, G.; Baraldi, E. Asymmetric dimethylarginine and related metabolites in exhaled breath condensate of children with cystic fibrosis. Clin. Respir. J. 2018, 12, 140. [Google Scholar] [CrossRef] [PubMed]
- Ingersoll, S.A.; Laval, J.; Forrest, O.A.; Preininger, M.; Brown, M.R.; Arafat, D.; Gibson, G.; Tangpricha, V.; Tirouvanziam, R. Mature Cystic Fibrosis Airway Neutrophils Suppress T Cell Function: Evidence for a Role of Arginase 1 but Not Programmed Death-Ligand 1. J. Immunol. 2015, 194, 5520. [Google Scholar] [CrossRef] [PubMed]
- Amin Said, A.M.; Al-Naqshbandi, A.A.; Amin, Y.K.; Ali, R.J. Association of the arginase Ι with bronchial asthma. JUBPAS 2019, 27, 238. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Comhair, S.A.A.; Janocha, A.J.; Lara, A.; Mavrakis, L.A.; Bennett, C.D.; Kalhan, S.C.; Erzurum, S.C. Arginine metabolic endotypes related to asthma severity. PLoS ONE 2017, 12, e0183066. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Ghosh, S.; Comhair, S.A.A.; Asosingh, K.; Janocha, A.J.; Mavrakis, D.A.; Bennett, C.D.; Gruca, L.L.; Graham, B.B.; Queisser, K.A.; et al. Increased mitochondrial arginine metabolism supports bioenergetics in asthma. J. Clin. Investig. 2016, 126, 2465. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Kubo, M.; Murakami, I.; Setiawan, H.; Takemoto, K.; Inoue, K.; Fujikura, Y.; Ogino, K. l-Arginine administration attenuates airway inflammation by altering l-arginine metabolism in an NC/Nga mouse model of asthma. J. Clin. Biochem. Nutr. 2015, 56, 201. [Google Scholar] [CrossRef] [Green Version]
- Vlad, D.; Albu, S. Arginase Isoform Expression in Chronic Rhinosinusitis. J. Clin. Med. 2019, 8, 1809. [Google Scholar] [CrossRef] [Green Version]
- Monin, L.; Griffiths, K.L.; Lam, W.Y.; Gopal, R.; Kang, D.D.; Ahmed, M.; Rajamanickam, A.; Cruz-Lagunas, A.; Zúñiga, J.; Babu, S.; et al. Helminth-induced arginase-1 exacerbates lung inflammation and disease severity in tuberculosis. J. Clin. Investig. 2015, 125, 4699. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, X.; Luo, Z.; Ma, L.; Zhu, S.; Wang, Z.; Wen, J.; Cheng, S.; Gu, W.; Lian, Q.; et al. ECM1 is an essential factor for the determination of M1 macrophage polarization in IBD in response to LPS stimulation. Proc. Natl. Acad. Sci. USA 2020, 117, 3083. [Google Scholar] [CrossRef]
- Hernandez, L.F.; Buchwald, P.; Abdulreda, M.H. Effect of Arginase-1 Inhibition on the Incidence of Autoimmune Diabetes in NOD Mice. Curr. Res. Diabetes Obes. J. 2018, 5, 555661. [Google Scholar]
- Hannemann, N.; Cao, S.; Eriksson, D.; Schnelzer, A.; Jordan, J.; Eberhardt, M.; Schleicher, U.; Rech, J.; Ramming, A.; Uebe, S.; et al. Transcription factor Fra-1 targets arginase-1 to enhance macrophage-mediated inflammation in arthritis. J. Clin. Investig. 2019, 129, 2669. [Google Scholar] [CrossRef] [Green Version]
- Choudry, M.; Tang, X.; Santorian, T.; Wasnik, S.; Xiao, J.; Xing, W.; Lau, K.-H.W.; Mohan, S.; Baylink, D.J.; Qin, X. Deficient arginase II expression without alteration in arginase I expression attenuated experimental autoimmune encephalomyelitis in mice. Immunology 2018, 155, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-K.; Lu, Q.-B.; Chen, W.-W.; Xu, W.; Liu, R.; Zhang, S.-F.; Du, J.; Li, H.; Yao, K.; Zhai, D.; et al. Arginine deficiency is involved in thrombocytopenia and immunosuppression in severe fever with thrombocytopenia syndrome. Sci. Transl. Med. 2018, 10, eaat4162. [Google Scholar] [CrossRef] [Green Version]
- Felizardo, A.A.; Caldas, I.S.; Mendonça, A.A.S.; Gonçalves, R.V.; Tana, F.L.; Almeida, L.A.; Novaes, R.D. Impact of Trypanosoma cruzi infection on nitric oxide synthase and arginase expression and activity in young and elderly mice. Free Radic. Biol. Med. 2018, 129, 227. [Google Scholar] [CrossRef]
- Zhang, N.; Deng, J.; Wu, F.; Lu, X.; Huang, L.; Zhao, M. Expression of arginase I and inducible nitric oxide synthase in the peripheral blood and lymph nodes of HIV-positive patients. Mol. Med. Rep. 2016, 13, 731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, L.-X.; Tang, X.; Zhu, J.-Y.; Zhang, W.; Tang, W.-Q.; Yan, J.; Xu, X.; Liang, H.-P. Cytochrome P450 1A1 enhances Arginase-1 expression, which reduces LPS-induced mouse peritonitis by targeting JAK1/STAT6. Cell Immunol. 2020, 349, 104047. [Google Scholar] [CrossRef]
- Mussai, F.; Wheat, R.; Sarrou, E.; Booth, S.; Stavrou, V.; Fultang, L.; Perry, T.; Kearns, P.; Cheng, P.; Keeshan, K.; et al. Targeting the arginine metabolic brake enhances immunotherapy for leukaemia. Int. J. Cancer 2019, 145, 2201. [Google Scholar] [CrossRef] [PubMed]
- Cull, A.H.; Mahendru, D.; Snetsinger, B.; Good, D.; Tyryshkin, K.; Chesney, A.; Ghorab, Z.; Reis, M.; Buckstein, R.; Wells, R.A.; et al. Overexpression of Arginase 1 is linked to DNMT3A and TET2 mutations in lower-grade myelodysplastic syndromes and chronic myelomonocytic leukemia. Leuk. Res. 2018, 65, 5. [Google Scholar] [CrossRef] [PubMed]
- Roci, I.; Watrous, J.D.; Lagerborg, K.A.; Lafranchi, L.; Lindqvist, A.; Jain, M.; Nilsson, R. Mapping Metabolic Events in the Cancer Cell Cycle Reveals Arginine Catabolism in the Committed SG(2)M Phase. Cell Rep. 2019, 26, 1691. [Google Scholar] [CrossRef] [Green Version]
- Fultang, L.; Gamble, L.D.; Gneo, L.; Berry, A.M.; Egan, S.A.; De Bie, F.; Yogev, O.; Eden, G.L.; Booth, S.; Brownhill, S.; et al. Macrophage-Derived IL1β and TNFα Regulate Arginine Metabolism in Neuroblastoma. Cancer Res. 2019, 79, 611. [Google Scholar] [CrossRef] [Green Version]
- Zaytouni, T.; Tsai, P.-Y.; Hitchcock, D.S.; DuBois, C.D.; Freinkman, E.; Lin, L.; Morales-Oyarvide, V.; Lenehan, P.J.; Wolpin, B.M.; Mino-Kenudson, M.; et al. Critical role for arginase 2 in obesity-associated pancreatic cancer. Nat. Commun. 2017, 8, 242. [Google Scholar] [CrossRef] [PubMed]
- De Santo, C.; Cheng, P.; Beggs, A.; Egan, S.; Bessudo, A.; Mussai, F. Metabolic therapy with PEG-arginase induces a sustained complete remission in immunotherapy-resistant melanoma. J. Hematol. Oncol. 2018, 11, 68. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ladeiras, D.; Xiong, Y.; Boligan, K.F.; Liang, X.; von Gunten, S.; Hunger, R.E.; Ming, X.-F.; Yang, Z. Arginase-II promotes melanoma migration and adhesion through enhancing hydrogen peroxide production and STAT3 signaling. J. Cell Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Czystowska-Kuzmicz, M.; Sosnowska, A.; Nowis, D.; Ramji, K.; Szajnik, M.; Chlebowska-Tuz, J.; Wolinska, E.; Gaj, P.; Grazul, M.; Pilch, Z.; et al. Small extracellular vesicles containing arginase-1 suppress T-cell responses and promote tumor growth in ovarian carcinoma. Nat. Commun. 2019, 10, 3000. [Google Scholar] [CrossRef]
- Setty, B.A.; Jin, Y.; Houghton, P.J.; Yeager, N.D.; Gross, T.G.; Nelin, L.D. Hypoxic Proliferation of Osteosarcoma Cells Depends on Arginase II. Cell Physiol. Biochem. 2016, 39, 802. [Google Scholar] [CrossRef]
- Zhang, I.; Alizadeh, D.; Liang, J.; Zhang, L.; Gao, H.; Song, Y.; Ren, H.; Ouyang, M.; Wu, X.; D’Apuzzo, M.; et al. Characterization of Arginase Expression in Glioma-Associated Microglia and Macrophages. PLoS ONE 2016, 11, e0165118. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Chen, W.; Chen, J.; Zheng, Q.; Dong, J.; Zhu, Y. The Oncogenic Role of ARG1 in Progression and Metastasis of Hepatocellular Carcinoma. BioMed. Res. Int. 2018, 2018, 2109865. [Google Scholar] [CrossRef]
- Yau, T.; Cheng, P.N.; Chan, P.; Chen, L.; Yuen, J.; Pang, R.; Fan, S.T.; Wheatley, D.N.; Poon, R.T. Preliminary efficacy, safety, pharmacokinetics, pharmacodynamics and quality of life study of pegylated recombinant human arginase 1 in patients with advanced hepatocellular carcinoma. Investig. New Drugs 2015, 33, 496. [Google Scholar] [CrossRef]
- Bedoya, A.M.; Tate, D.J.; Baena, A.; Córdoba, C.M.; Borrero, M.; Pareja, R.; Rojas, F.; Patterson, J.R.; Herrero, R.; Zea, A.H.; et al. Immunosuppression in cervical cancer with special reference to arginase activity. Gynecol. Oncol. 2014, 135, 74. [Google Scholar] [CrossRef]
- Polis, B.; Srikanth, K.D.; Gurevich, V.; Bloch, N.; Gil-Henn, H.; Samson, A.O. Arginase Inhibition Supports Survival and Differentiation of Neuronal Precursors in Adult Alzheimer’s Disease Mice. Int. J. Mol. Sci. 2020, 8, 1133. [Google Scholar] [CrossRef] [Green Version]
- Villalba, N.; Sackheim, A.M.; Nunez, I.A.; Hill-Eubanks, D.C.; Nelson, M.T.; Wellman, G.C.; Freeman, K. Traumatic Brain Injury Causes Endothelial Dysfunction in the Systemic Microcirculation through Arginase-1-Dependent Uncoupling of Endothelial Nitric Oxide Synthase. J. Neurotrauma 2017, 34, 192. [Google Scholar] [CrossRef] [PubMed]
- Vemula, P.; Jing, Y.; Zhang, H.; Hunt, J.B.; Sandusky-Beltran, L.A.; Lee, D.C.; Liu, P. Altered brain arginine metabolism in a mouse model of tauopathy. Amino Acids 2019, 51, 513. [Google Scholar] [CrossRef] [PubMed]
- Nath, N.; Prasad, H.K.; Kumar, M. Cerebroprotective effects of hydrogen sulfide in homocysteine-induced neurovascular permeability: Involvement of oxidative stress, arginase, and matrix metalloproteinase-9. J. Cell Physiol. 2019, 234, 3007. [Google Scholar] [CrossRef] [PubMed]
- Patassini, S.; Begley, P.; Reid, S.J.; Xu, J.; Church, S.J.; Curtis, M.; Dragunow, M.; Waldvogel, H.J.; Unwin, R.D.; Snell, R.G.; et al. Identification of elevated urea as a severe, ubiquitous metabolic defect in the brain of patients with Huntington’s disease. Biochem. Biophys. Res. Commun. 2015, 468, 161. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.; Kim, J.; Lee, A.-R.; Lee, J.-M.; Kim, O.-J.; Kim, J.-K.; Oh, S.-H. Alteration of microRNA 340-5p and Arginase-1 Expression in Peripheral Blood Cells during Acute Ischemic Stroke. Mol. Neurobiol. 2019, 56, 3211. [Google Scholar] [CrossRef]
- Ahmad, A.S.; Shah, Z.A.; Doré, S. Protective Role of Arginase II in Cerebral Ischemia and Excitotoxicity. J. Neurol. Neurosci. 2016, 7, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pudlo, M.; Demougeot, C.; Girard-Thernier, C. Arginase Inhibitors: A Rational Approach Over One Century. Med. Res. Rev. 2017, 37, 475. [Google Scholar] [CrossRef]
- Hunter, A.; Downs, C.E. The inhibition of arginase by amino acids. J. Biol. Chem. 1945, 157, 427. [Google Scholar]
- Lättig-Tünnemann, G.; Prinz, M.; Hoffmann, D.; Behlke, J.; Palm-Apergi, C.; Morano, I.; Herce, H.D.; Cardoso, M.C. Backbone rigidity and static presentation of guanidinium groups increases cellular uptake of arginine-rich cell-penetrating peptides. Nat. Commun. 2011, 2, 453. [Google Scholar] [CrossRef]
- Rosenfeld, J.L.; Dutta, S.P.; Chheda, G.B.; Tritsch, G.L. Purine and pyrimidine inhibitors of arginase. Biochim. Biophys. Acta 1975, 410, 164. [Google Scholar] [CrossRef]
- Méndez, J.D.; Yáñez, R.; Wong, C.; Hicks, J.J. Uterine arginase inhibition affect the rat embryonic development. Contraception 1986, 33, 597. [Google Scholar] [CrossRef]
- Trujillo, J.G.; Ceballos, G.; Yañez, R.; Joseph-Nathan, P. Regioselective Synthesis of (+) -S-2-Amino-5-iodoacetamidopentanoic and (+) -S-2-amino-6-iodoacetamidohexanoic Acids. Synth. Commun. 1991, 21, 683. [Google Scholar] [CrossRef]
- Trujillo-Ferrara, J.; Koizumi, G.; Mun~noz, O.; Joseph-Nathan, P.; Yañez, R. Antitumor effect and toxicity of two new active-site-directed irreversible ornithine decarboxylase and extrahepatic arginase inhibitors. Cancer Lett. 1992, 67, 193. [Google Scholar] [CrossRef]
- Baydoun, A.R.; Morgan, D.M. Inhibition of ornithine decarboxylase potentiates nitric oxide production in LPS-activated J774 cells. Br. J. Pharmacol. 1998, 125, 1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LoGiudice, N.; Le, L.; Abuan, I.; Leizorek, Y.; Roberts, S.C. Alpha-Difluoromethylornithine, an Irreversible Inhibitor of Polyamine Biosynthesis, as a Therapeutic Strategy against Hyperproliferative and Infectious Diseases. Med. Sci 2018, 6, 12. [Google Scholar]
- Selamnia, M.; Mayeur, C.; Robert, V.; Blachier, F. Alpha-difluoromethylornithine (DFMO) as a potent arginase activity inhibitor in human colon carcinoma cells. Biochem. Pharmacol. 1998, 55, 1241. [Google Scholar] [CrossRef]
- Daghigh, F.; Fukuto, J.M.; Ash, D.E. Inhibition of Rat Liver Arginase by an Intermediate in NO Biosynthesis, NG-Hydroxy-L-arginine: Implications for the Regulation of Nitric Oxide Biosynthesis by Arginase. Biochem. Biophys. Res. Commun. 1994, 202, 174. [Google Scholar] [CrossRef]
- Custot, J.; Boucher, J.-L.; Vadon, S.; Guedes, C.; Dijols, S.; Delaforge, M.; Mansuy, D. Nω-Hydroxyamino-α-amino acids as a new class of very strong inhibitors of arginases. J. Biol. Inorg. Chem. 1996, 1, 73. [Google Scholar] [CrossRef]
- Baggio, R.; Emig, F.A.; Christianson, D.W.; Ash, D.E.; Chakder, S.; Rattan, S. Biochemical and functional profile of a newly developed potent and isozyme-selective arginase inhibitor. J. Pharmacol. Exp. Ther. 1999, 290, 1409. [Google Scholar]
- Tenu, J.-P.; Lepoivre, M.; Moali, C.; Brollo, M.; Mansuy, D.; Boucher, J.-L. Effects of the New Arginase Inhibitor Nω-Hydroxy-nor-l-Arginine on NO Synthase Activity in Murine Macrophages. Nitric Oxide Biol. Chem. 1999, 3, 427. [Google Scholar] [CrossRef]
- Xian, M.; Fujiwara, N.; Wen, Z.; Cai, T.; Kazuma, S.; Janczuk, A.J.; Tang, X.; Telyatnikov, V.V.; Zhang, Y.; Chen, X.; et al. Novel substrates for nitric oxide synthases. Bioorg. Med. Chem. 2002, 10, 3049. [Google Scholar] [CrossRef]
- Custot, J.; Moali, C.; Brollo, M.; Boucher, J.L.; Delaforge, M.; Mansuy, D.; Tenu, J.P.; Zimmermann, J.L. The New α-Amino Acid Nω-Hydroxy-nor-l-arginine: a High-Affinity Inhibitor of Arginase Well Adapted To Bind to Its Manganese Cluster. J. Am. Chem. Soc. 1997, 119, 4086. [Google Scholar] [CrossRef]
- Vadon, S.; Custot, J.; Boucher, J.-L.; Mansuy, D. Synthesis and effects on arginase and nitric oxide synthase of two novel analogues of Nω-hydroxyarginine, Nω-hydroxyindospicine and p-hydroxyamidinophenylalanine. J. Chem. Soc. Perkin 1 1996, 7, 645. [Google Scholar] [CrossRef]
- Havlínová, Z.; Hroch, M.; Nagy, A.; Sišpera, L.; Holeček, M.; Chládek, J. Single- and multiple-dose pharmacokinetics of arginase inhibitor Nω-hydroxy-nor-L-arginine, and its effect on plasma amino acids concentrations in Wistar rats. Gen. Physiol. Biophys. 2014, 33, 189. [Google Scholar] [CrossRef] [Green Version]
- Havlinova, Z.; Babicova, A.; Hroch, M.; Chladek, J. Comparative pharmacokinetics of N(ω)-hydroxy-nor-L-arginine, an arginase inhibitor, after single-dose intravenous, intraperitoneal and intratracheal administration to brown Norway rats. Xenobiotica 2013, 43, 886. [Google Scholar] [CrossRef]
- Avtandilyan, N.; Javrushyan, H.; Mamikonyan, A.; Grigoryan, A.; Trchounian, A. The potential therapeutic effect of NG-hydroxy-nor-L-arginine in 7,12-dimethylbenz(a)anthracene-induced breast cancer in rats. Exp. Mol. Pathol. 2019, 111, 104316. [Google Scholar] [CrossRef]
- Ng, K.P.; Manjeri, A.; Lee, L.M.; Chan, Z.E.; Tan, C.Y.; Tan, Q.D.; Majeed, A.Q.; Lee, K.L.; Chuah, C.; Suda, T.; et al. The arginase inhibitor Nω-hydroxy-nor-arginine (nor-NOHA) induces apoptosis in leukemic cells specifically under hypoxic conditions but CRISPR/Cas9 excludes arginase 2 (ARG2) as the functional target. PLoS ONE 2018, 13, e0205254. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, N.; Ogino, K.; Takemoto, K.; Hamanishi, S.; Wang, D.-H.; Takigawa, T.; Shibamori, M.; Ishiyama, H.; Fujikura, Y. Direct inhibition of arginase attenuated airway allergic reactions and inflammation in a Dermatophagoides farinae-induced NC/Nga mouse model. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 299, L17. [Google Scholar] [CrossRef] [Green Version]
- Arıkan-Ayyıldız, Z.; Karaman, M.; Tuncel, T.; Kiray, M.; Bağrıyanık, A.; Yilmaz, O.; Uzuner, N.; Karaman, O. Beneficial effects of arginase inhibition and inhaled L-arginine administration on airway histology in a murine model of chronic asthma. Allergol. Immunopathol. 2014, 42, 316. [Google Scholar] [CrossRef]
- Bagnost, T.; Berthelot, A.; Bouhaddi, M.; Laurant, P.; André, C.; Guillaume, Y.; Demougeot, C. Treatment with the arginase inhibitor N(omega)-hydroxy-nor-L-arginine improves vascular function and lowers blood pressure in adult spontaneously hypertensive rat. J. Hypertens. 2008, 26, 1110. [Google Scholar] [CrossRef]
- Jung, C.; Gonon, A.T.; Sjöquist, P.-O.; Lundberg, J.O.; Pernow, J. Arginase inhibition mediates cardioprotection during ischaemia–reperfusion. Cardiovasc. Res. 2009, 85, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prati, C.; Berthelot, A.; Kantelip, B.; Wendling, D.; Demougeot, C. Treatment with the arginase inhibitor Nw-hydroxy-nor-L-arginine restores endothelial function in rat adjuvant-induced arthritis. Arthritis Res. Ther. 2012, 14, R130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivon, V.C.; Fraga-Silva, R.A.; Segers, D.; Demougeot, C.; de Oliveira, A.M.; Savergnini, S.S.; Berthelot, A.; de Crom, R.; Krams, R.; Stergiopulos, N.; et al. Arginase inhibition prevents the low shear stress-induced development of vulnerable atherosclerotic plaques in ApoE -/- mice. Atherosclerosis 2013, 227, 236. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.-L.; Aoyama, S.; Nagai, R.; Miyoshi, N.; Ohshima, H. Inhibition of L-arginine metabolizing enzymes by L-arginine-derived advanced glycation end products. J. Clin. Biochem. Nutr. 2010, 46, 177. [Google Scholar] [CrossRef] [Green Version]
- Reczkowski, R.S.; Ash, D.E. Rat Liver Arginase: Kinetic Mechanism, Alternate Substrates, and Inhibitors. Arch. Biochem. Biophys. 1994, 312, 31. [Google Scholar] [CrossRef]
- Baggio, R.; Elbaum, D.; Kanyo, Z.F.; Carroll, P.J.; Cavalli, R.C.; Ash, D.E.; Christianson, D.W. Inhibition of Mn2+2-Arginase by Borate Leads to the Design of a Transition State Analogue Inhibitor, 2(S)-Amino-6-boronohexanoic Acid. J. Am. Chem. Soc. 1997, 119, 8107. [Google Scholar] [CrossRef]
- Colleluori, D.M.; Ash, D.E. Classical and Slow-Binding Inhibitors of Human Type II Arginase. Biochemistry 2001, 40, 9356. [Google Scholar] [CrossRef]
- Di Costanzo, L.; Ilies, M.; Thorn, K.J.; Christianson, D.W. Inhibition of human arginase I by substrate and product analogues. Arch. Biochem. Biophys. 2010, 496, 101. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.D.; Kim, N.N.; Traish, A.M.; Christianson, D.W. Arginase-boronic acid complex highlights a physiological role in erectile function. Nat. Struct. Biol. 1999, 6, 1043. [Google Scholar]
- Cama, E.; Pethe, S.; Boucher, J.-L.; Han, S.; Emig, F.A.; Ash, D.E.; Viola, R.E.; Mansuy, D.; Christianson, D.W. Inhibitor Coordination Interactions in the Binuclear Manganese Cluster of Arginase. Biochemistry 2004, 43, 8987. [Google Scholar] [CrossRef]
- Christianson, D.W. Arginase: Structure, Mechanism, and Physiological Role in Male and Female Sexual Arousal. Acc. Chem. Res. 2005, 38, 191. [Google Scholar] [CrossRef] [PubMed]
- Maarsingh, H.; Zuidhof, A.B.; Bos, I.S.T.; Duin, M.v.; Boucher, J.-L.; Zaagsma, J.; Meurs, H. Arginase Inhibition Protects against Allergen-induced Airway Obstruction, Hyperresponsiveness, and Inflammation. Am. J. Respir. Crit. Care Med. 2008, 178, 565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Bhatta, A.; Toque, H.A.; Rojas, M.; Yao, L.; Xu, Z.; Patel, C.; Caldwell, R.B.; Caldwell, R.W. Arginase inhibition enhances angiogenesis in endothelial cells exposed to hypoxia. Microvasc. Res. 2015, 98, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, H.M.; Lee, J.H.; Min, B.S.; Jeon, B.H.; Hoe, K.L.; Kim, Y.M.; Ryoo, S. A Novel Arginase Inhibitor Derived from Scutellavia indica Restored Endothelial Function in ApoE-Null Mice Fed a High-Cholesterol Diet. J. Pharmacol. Exp. Ther. 2015, 355, 57. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.C.; Park, J.T.; Jeon, Y.G.; Jeon, B.H.; Hoe, K.L.; Kim, Y.M.; Lim, H.K.; Ryoo, S. Arginase Inhibition Restores Peroxynitrite-Induced Endothelial Dysfunction via L-Arginine-Dependent Endothelial Nitric Oxide Synthase Phosphorylation. Yonsei Med. J. 2016, 57, 1329. [Google Scholar] [CrossRef] [Green Version]
- Steppan, J.; Tran, H.T.; Bead, V.R.; Oh, Y.J.; Sikka, G.; Bivalacqua, T.J.; Burnett, A.L.; Berkowitz, D.E.; Santhanam, L. Arginase Inhibition Reverses Endothelial Dysfunction, Pulmonary Hypertension, and Vascular Stiffness in Transgenic Sickle Cell Mice. Anesth. Analg. 2016, 123, 652. [Google Scholar] [CrossRef] [Green Version]
- Mehl, A.; Ghorbani, P.; Douda, D.; Huang, H.; Palaniyar, N.; Ratjen, F.; Grasemann, H. Effect of Arginase Inhibition on Pulmonary L-Arginine Metabolism in Murine Pseudomonas Pneumonia. PLoS ONE 2014, 9, e90232. [Google Scholar] [CrossRef]
- Pera, T.; Zuidhof, A.B.; Smit, M.; Menzen, M.H.; Klein, T.; Flik, G.; Zaagsma, J.; Meurs, H.; Maarsingh, H. Arginase inhibition prevents inflammation and remodeling in a guinea pig model of chronic obstructive pulmonary disease. J. Pharmacol. Exp. Ther. 2014, 349, 229. [Google Scholar] [CrossRef]
- Cho, J.S.; Oh, Y.J.; Kim, O.S.; Na, S. The effects of arginase inhibitor on lung oxidative stress and inflammation caused by pneumoperitoneum in rats. BMC Anesthesiol. 2015, 15, 129. [Google Scholar] [CrossRef] [Green Version]
- Grasemann, H.; Dhaliwal, R.; Ivanovska, J.; Kantores, C.; McNamara, P.J.; Scott, J.A.; Belik, J.; Jankov, R.P. Arginase inhibition prevents bleomycin-induced pulmonary hypertension, vascular remodeling, and collagen deposition in neonatal rat lungs. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 308, L503. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Hilliard, B.; Carmody, R.J.; Tsabary, G.; Shin, H.; Christianson, D.W.; Chen, Y.H. Arginase and autoimmune inflammation in the central nervous system. Immunology 2003, 110, 141. [Google Scholar] [CrossRef] [PubMed]
- Collet, S.; Carreaux, F.; Boucher, J.-L.; Pethe, S.; Lepoivre, M.; Danion-Bougot, R.; Danion, D. Synthesis and evaluation of ω-borono-α-amino acids as active-site probes of arginase and nitric oxide synthases. J. Chem. Soc. Perkin 1 2000, 2, 177. [Google Scholar] [CrossRef]
- Cama, E.; Shin, H.; Christianson, D.W. Design of amino acid sulfonamides as transition-state analogue inhibitors of arginase. J. Am. Chem. Soc. 2003, 125, 13052. [Google Scholar] [CrossRef]
- Shin, H.; Cama, E.; Christianson, D.W. Design of amino acid aldehydes as transition-state analogue inhibitors of arginase. J. Am. Chem. Soc. 2004, 126, 10278. [Google Scholar] [CrossRef] [PubMed]
- Ilies, M.; Di Costanzo, L.; North, M.L.; Scott, J.A.; Christianson, D.W. 2-Aminoimidazole Amino Acids as Inhibitors of the Binuclear Manganese Metalloenzyme Human Arginase I. J. Med. Chem. 2010, 53, 4266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.K.; Sieburth, S.M. Synthesis and Properties of a Sterically Unencumbered δ-Silanediol Amino Acid. J. Org. Chem. 2012, 77, 2901. [Google Scholar] [CrossRef] [PubMed]
- Zakharian, T.Y.; Di Costanzo, L.; Christianson, D.W. (S)-2-Amino-6-nitrohexanoic Acid Binds to Human Arginase I through Multiple Nitro−Metal Coordination Interactions in the Binuclear Manganese Cluster. J. Am. Chem. Soc. 2008, 130, 17254. [Google Scholar] [CrossRef]
- Kim, N.N.; Cox, J.D.; Baggio, R.F.; Emig, F.A.; Mistry, S.K.; Harper, S.L.; Speicher, D.W.; Morris, S.M.; Ash, D.E.; Traish, A.; et al. Probing Erectile Function: S-(2-Boronoethyl)-l-Cysteine Binds to Arginase as a Transition State Analogue and Enhances Smooth Muscle Relaxation in Human Penile Corpus Cavernosum. Biochemistry 2001, 40, 2678. [Google Scholar] [CrossRef]
- Johnson, F.K.; Johnson, R.A.; Peyton, K.J.; Durante, W. Arginase inhibition restores arteriolar endothelial function in Dahl rats with salt-induced hypertension. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1057. [Google Scholar] [CrossRef]
- Ryoo, S.; Gupta, G.; Benjo, A.; Lim, H.K.; Camara, A.; Sikka, G.; Lim, H.K.; Sohi, J.; Santhanam, L.; Soucy, K.; et al. Endothelial arginase II: A novel target for the treatment of atherosclerosis. Circ. Res. 2008, 102, 923. [Google Scholar] [CrossRef] [Green Version]
- Lewis, C.; Zhu, W.; Pavkov, M.L.; Kinney, C.M.; Dicorleto, P.E.; Kashyap, V.S. Arginase blockade lessens endothelial dysfunction after thrombosis. J. Vasc. Surg. 2008, 48, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ckless, K.; Lampert, A.; Reiss, J.; Kasahara, D.; Poynter, M.E.; Irvin, C.G.; Lundblad, L.K.A.; Norton, R.; van der Vliet, A.; Janssen-Heininger, Y.M.W. Inhibition of arginase activity enhances inflammation in mice with allergic airway disease, in association with increases in protein S-nitrosylation and tyrosine nitration. J. Immunol. 2008, 181, 4255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krotova, K.; Patel, J.M.; Block, E.R.; Zharikov, S. Endothelial arginase II responds to pharmacological inhibition by elevation in protein level. Mol. Cell Biochem. 2010, 343, 211. [Google Scholar] [CrossRef] [PubMed]
- Busnel, O.; Carreaux, F.; Carboni, B.; Pethe, S.; Goff, S.V.-L.; Mansuy, D.; Boucher, J.-L. Synthesis and evaluation of new ω-borono-α-amino acids as rat liver arginase inhibitors. Bioorg. Med. Chem. 2005, 13, 2373. [Google Scholar] [CrossRef]
- Ilies, M.; Di Costanzo, L.; Dowling, D.P.; Thorn, K.J.; Christianson, D.W. Binding of α,α-Disubstituted Amino Acids to Arginase Suggests New Avenues for Inhibitor Design. J. Med. Chem. 2011, 54, 5432. [Google Scholar] [CrossRef] [Green Version]
- Mortier, J.; Prévost, J.R.C.; Sydow, D.; Teuchert, S.; Omieczynski, C.; Bermudez, M.; Frédérick, R.; Wolber, G. Arginase Structure and Inhibition: Catalytic Site Plasticity Reveals New Modulation Possibilities. Sci. Rep. 2017, 7, 13616. [Google Scholar] [CrossRef] [Green Version]
- Burlingham, B.T.; Widlanski, T.S. An Intuitive Look at the Relationship of Ki and IC50: A More General Use for the Dixon Plot. J. Chem. Educ. 2003, 80, 214. [Google Scholar] [CrossRef]
- Golebiowski, A.; Paul Beckett, R.; Van Zandt, M.; Ji, M.K.; Whitehouse, D.; Ryder, T.R.; Jagdmann, E.; Andreoli, M.; Mazur, A.; Padmanilayam, M.; et al. 2-Substituted-2-amino-6-boronohexanoic acids as arginase inhibitors. Bioorganic Med. Chem. Lett. 2013, 23, 2027. [Google Scholar] [CrossRef]
- Van Zandt, M.C.; Whitehouse, D.L.; Golebiowski, A.; Ji, M.K.; Zhang, M.; Beckett, R.P.; Jagdmann, G.E.; Ryder, T.R.; Sheeler, R.; Andreoli, M.; et al. Discovery of (R)-2-Amino-6-borono-2-(2-(piperidin-1-yl)ethyl)hexanoic Acid and Congeners As Highly Potent Inhibitors of Human Arginases I and II for Treatment of Myocardial Reperfusion Injury. J. Med. Chem. 2013, 56, 2568. [Google Scholar] [CrossRef]
- Golebiowski, A.; Whitehouse, D.; Beckett, R.P.; Van Zandt, M.; Ji, M.K.; Ryder, T.R.; Jagdmann, E.; Andreoli, M.; Lee, Y.; Sheeler, R.; et al. Synthesis of quaternary α-amino acid-based arginase inhibitors via the Ugi reaction. Bioorganic Med. Chem. Lett. 2013, 23, 4837. [Google Scholar] [CrossRef]
- Blaszczyk, R.; Brzezinska, J.; Dymek, B.; Stanczak, P.S.; Mazurkiewicz, M.; Olczak, J.; Nowicka, J.; Dzwonek, K.; Zagozdzon, A.; Golab, J.; et al. Discovery and Pharmacokinetics of Sulfamides and Guanidines as Potent Human Arginase 1 Inhibitors. ACS Med. Chem. Lett. 2020, 11, 433. [Google Scholar] [CrossRef] [PubMed]
- Van Zandt, M.C.; Jagdmann, G.E.; Whitehouse, D.L.; Ji, M.; Savoy, J.; Potapova, O.; Cousido-Siah, A.; Mitschler, A.; Howard, E.I.; Pyle, A.M.; et al. Discovery of N-Substituted 3-Amino-4-(3-boronopropyl)pyrrolidine-3-carboxylic Acids as Highly Potent Third-Generation Inhibitors of Human Arginase I and II. J. Med. Chem. 2019, 62, 8164. [Google Scholar] [PubMed]
- Mitcheltree, M.J.; Li, D.; Achab, A.; Beard, A.; Chakravarthy, K.; Cheng, M.; Cho, H.; Eangoor, P.; Fan, P.; Gathiaka, S.; et al. Discovery and Optimization of Rationally Designed Bicyclic Inhibitors of Human Arginase to Enhance Cancer Immunotherapy. ACS Med. Chem. Lett. 2020, 11, 582. [Google Scholar] [CrossRef] [PubMed]
- Ivanenkov, Y.A.; Chufarova, N.V. Small-molecule arginase inhibitors. Pharm. Pat. Anal. 2014, 3, 65. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.; Jackson, A.; Patel, P. The Characterisation of Arginase-1 Expressing Myeloid-Derived Suppressor Cells in Advanced Melanoma. Ann. Oncol. 2014, 25, iv386. [Google Scholar] [CrossRef] [Green Version]
- Schupp, J.; Krebs, F.K.; Zimmer, N.; Trzeciak, E.; Schuppan, D.; Tuettenberg, A. Targeting myeloid cells in the tumor sustaining microenvironment. Cell Immunol. 2019, 343, 103713. [Google Scholar] [CrossRef]
- Grobben, Y.; Uitdehaag, J.C.M.; Willemsen-Seegers, N.; Tabak, W.W.A.; de Man, J.; Buijsman, R.C.; Zaman, G.J.R. Structural insights into human Arginase-1 pH dependence and its inhibition by the small molecule inhibitor CB-1158. J. Struct. Biol. 2020, 4, 100014. [Google Scholar] [CrossRef]
- Steggerda, S.M.; Bennett, M.K.; Chen, J.; Emberley, E.; Huang, T.; Janes, J.R.; Li, W.; MacKinnon, A.L.; Makkouk, A.; Marguier, G.; et al. Inhibition of arginase by CB-1158 blocks myeloid cell-mediated immune suppression in the tumor microenvironment. J. Immunother. Cancer 2017, 5, 101. [Google Scholar] [CrossRef]
- Naing, A.; Bauer, T.; Papadopoulos, K.P.; Rahma, O.; Tsai, F.; Garralda, E.; Naidoo, J.; Pai, S.; Gibson, M.K.; Rybkin, I.; et al. Phase I study of the arginase inhibitor INCB001158 (1158) alone and in combination with pembrolizumab (PEM) in patients (Pts) with advanced/metastatic (adv/met) solid tumours. Ann. Oncol. 2019, 30, v160. [Google Scholar] [CrossRef]
- Arginase Inhibitor CB-280. Available online: https://www.calithera.com/arginase-inhibitor-cb-280/ (accessed on 11 June 2020).
- Grzybowski, M.M.; Stańczak, P.S.; Pęczkowicz-Szyszka, J.; Wolska, P.; Zdziarska, A.M.; Mazurkiewicz, M.; Brzezińska, J.; Blaszczyk, R.; Gołębiowski, A.; Dobrzański, P.; et al. Novel dual arginase 1/2 inhibitor OATD-02 (OAT-1746) improves the efficacy of immune checkpoint inhibitors. Ann. Oncol. 2017, 28, xi20. [Google Scholar] [CrossRef]
- Pilanc-kudlek, P.; Cyranowski, S.; Wojnicki, K.; Ochocka, N.; Grzybowski, M.; Stańczak, P.; Pomper, P.; Błaszczyk, R.; Gołębiowski, A.; Dobrzański, P.; et al. Novel arginase inhibitor alone and in combination with an immune check point inhibitor reduces tumour growth in murine experimental gliomas. Ann. Oncol. 2019, 30, xi56. [Google Scholar] [CrossRef]
- Xiong, H.; Mittman, S.; Rodriguez, R.; Moskalenko, M.; Pacheco-Sanchez, P.; Yang, Y.; Nickles, D.; Cubas, R. Anti–PD-L1 Treatment Results in Functional Remodeling of the Macrophage Compartment. Cancer Res. 2019, 79, 1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miret, J.J.; Kirschmeier, P.; Koyama, S.; Zhu, M.; Li, Y.Y.; Naito, Y.; Wu, M.; Malladi, V.S.; Huang, W.; Walker, W.; et al. Suppression of Myeloid Cell Arginase Activity leads to Therapeutic Response in a NSCLC Mouse Model by Activating Anti-Tumor Immunity. J. Immunother. Cancer 2019, 7, 32. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, M.P.M.; Kurhade, S.H.; Maarsingh, H.; Erceg, S.; Hulsbeek, I.R.; Boekema, P.H.; Kistemaker, L.E.M.; van Faassen, M.; Kema, I.P.; Elsinga, P.H.; et al. Pharmacological screening identifies SHK242 and SHK277 as novel arginase inhibitors with efficacy against allergen-induced airway narrowing in vitro and in vivo. J. Pharmacol. Exp. Ther. 2020, 374, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelkawy, K.S.; Lack, K.; Elbarbry, F. Pharmacokinetics and Pharmacodynamics of Promising Arginase Inhibitors. Eur J. Drug Metab. Pharm. 2017, 42, 355. [Google Scholar] [CrossRef] [PubMed]
- Achilli, C.; Ciana, A.; Fagnoni, M.; Balduini, C.; Minetti, G. Susceptibility to hydrolysis of phenylboronic pinacol esters at physiological pH. Cent. Eur. J. Chem. 2013, 11, 137. [Google Scholar] [CrossRef]
- Girard-Thernier, C.; Pham, T.N.; Demougeot, C. The Promise of Plant-Derived Substances as Inhibitors of Arginase. Mini Rev. Med. Chem. 2015, 15, 798. [Google Scholar] [CrossRef]
- Rahman, N.N.; Khan, M.; Hasan, R. Bioactive components from Ficus glomerata. Pure Appl. Chem. 1994, 66, 2287. [Google Scholar] [CrossRef] [Green Version]
- Duffy, C.F.; Killeen, G.F.; Connolly, C.D.; Power, R.F. Effects of Dietary Supplementation with Yucca schidigera Roezl ex Ortgies and Its Saponin and Non-saponin Fractions on Rat Metabolism. J. Agric. Food Chem. 2001, 49, 3408. [Google Scholar] [CrossRef]
- Schnorr, O.; Brossette, T.; Momma, T.Y.; Kleinbongard, P.; Keen, C.L.; Schroeter, H.; Sies, H. Cocoa flavanols lower vascular arginase activity in human endothelial cells in vitro and in erythrocytes in vivo. Arch. Biochem. Biophys. 2008, 476, 211. [Google Scholar] [CrossRef]
- Shin, W.; Cuong, T.D.; Lee, J.H.; Min, B.; Jeon, B.H.; Lim, H.K.; Ryoo, S. Arginase Inhibition by Ethylacetate Extract of Caesalpinia sappan Lignum Contributes to Activation of Endothelial Nitric Oxide Synthase. Korean J. Physiol. Pharmacol. 2011, 15, 123. [Google Scholar] [CrossRef] [PubMed]
- Iwalokun, B.A.; Hodonu, S.A.; Nwoke, S.; Ojo, O.; Agomo, P.U. Evaluation of the Possible Mechanisms of Antihypertensive Activity of Loranthus micranthus: An African Mistletoe. Biochem. Res. Int. 2011, 2011, 159439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, W.; Yoon, J.; Oh, G.T.; Ryoo, S. Korean red ginseng inhibits arginase and contributes to endotheliumdependent vasorelaxation through endothelial nitric oxide synthase coupling. J. Ginseng Res. 2013, 37, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- André, C.; Herlem, G.; Gharbi, T.; Guillaume, Y.C. A new arginase enzymatic reactor: Development and application for the research of plant-derived inhibitors. J. Pharm. Biomed. Anal. 2011, 55, 48. [Google Scholar] [CrossRef] [PubMed]
- André, C.; Kapustikova, I.; Lethier, L.; Guillaume, Y.C. A Particulate Biochromatographic Support for the Research of Arginase Inhibitors Doped with Nanomaterials: Differences Observed Between Carbon and Boron Nitride Nanotubes. Application to Three Plant Extracts. Chromatographia 2014, 77, 1521. [Google Scholar] [CrossRef]
- Anadozie, S.O.; Akinyemi, J.A.; Agunbiade, S.; Ajiboye, B.O.; Adewale, O.B. Bryophyllum pinnatum inhibits arginase II activity and prevents oxidative damage occasioned by carbon tetrachloride (CCl4) in rats. Biomed. Pharm. 2018, 101, 8. [Google Scholar] [CrossRef]
- Attia, R.; Messaoud, C.; Arraki, K.; Zedet, A.; Demougeot, C.; Boussaïd, M.; Girard, C. Phytochemical screening and arginase inhibitory activity of extracts from several Tunisian medicinal plants. S. Afr. J. Bot. 2019, 120, 313. [Google Scholar] [CrossRef]
- Bujor, A.; Miron, A.; Luca, S.V.; Skalicka-Wozniak, K.; Silion, M.; Trifan, A.; Girard, C.; Demougeot, C.; Totoson, P. Vasorelaxant effects of Crataegus pentagyna: Links with arginase inhibition and phenolic profile. J. Ethnopharmacol. 2020, 252, 112559. [Google Scholar] [CrossRef]
- Bordage, S.; Pham, T.-N.; Zedet, A.; Gugglielmetti, A.-S.; Nappey, M.; Demougeot, C.; Girard-Thernier, C. Investigation of Mammal Arginase Inhibitory Properties of Natural Ubiquitous Polyphenols by Using an Optimized Colorimetric Microplate Assay. Planta Medica 2017, 83, 647. [Google Scholar] [CrossRef] [Green Version]
- Muller, J.; Cardey, B.; Zedet, A.; Desingle, C.; Grzybowski, M.; Pomper, P.; Foley, S.; Harakat, D.; Ramseyer, C.; Girard, C.; et al. Synthesis, evaluation and molecular modelling of piceatannol analogues as arginase inhibitors. RSC Med. Chem. 2020. [Google Scholar] [CrossRef]
- Pham, T.-N.; Bordage, S.; Pudlo, M.; Demougeot, C.; Thai, K.-M.; Girard-Thernier, C. Cinnamide Derivatives as Mammalian Arginase Inhibitors: Synthesis, Biological Evaluation and Molecular Docking. Int. J. Mol. Sci. 2016, 17, 1656. [Google Scholar] [CrossRef] [PubMed]
- Bourjot, M.; Zedet, A.; Demange, B.; Pudlo, M.; Girard-Thernier, C. In Vitro Mammalian Arginase Inhibitory and Antioxidant Effects of Amide Derivatives Isolated from the Hempseed Cakes (Cannabis sativa). Planta Med. Int. Open 2016, 3, e64. [Google Scholar] [CrossRef] [Green Version]
- Woo, A.; Min, B.; Ryoo, S. Piceatannol-3′-O-β-D-glucopyranoside as an active component of rhubarb activates endothelial nitric oxide synthase through inhibition of arginase activity. Exp. Mol. Med. 2010, 42, 524. [Google Scholar] [CrossRef] [PubMed]
- Yi, B.; Nguyen, M.C.; Won, M.-H.; Kim, Y.M.; Ryoo, S. Arginase Inhibitor 2,3,5,4′-Tetrahydroxystilbene-2-O-β-D-Glucoside Activates Endothelial Nitric Oxide Synthase and Improves Vascular Function. Planta Med. 2017, 83, 210. [Google Scholar] [CrossRef] [PubMed]
- Woo, A.; Shin, W.; Cuong, T.D.; Min, B.; Lee, J.H.; Jeon, B.H.; Ryoo, S. Arginase inhibition by piceatannol-3′-O-β-D-glucopyranoside improves endothelial dysfunction via activation of endothelial nitric oxide synthase in ApoE-null mice fed a high-cholesterol diet. Int. J. Mol. Med. 2013, 31, 803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.W.; Cuong, T.D.; Hung, T.M.; Ryoo, S.; Lee, J.H.; Min, B.S. Arginase II inhibitory activity of flavonoid compounds from Scutellaria indica. Arch. Pharm. Res. 2013, 36, 922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arraki, K.; Totoson, P.; Attia, R.; Zedet, A.; Pudlo, M.; Messaoud, C.; Demougeot, C.; Girard, C. Arginase inhibitory properties of flavonoid compounds from the leaves of Mulberry (Morus alba, Moraceae). J. Pharm. Pharm. 2020. [Google Scholar] [CrossRef] [PubMed]
- Mukai, R.; Shirai, Y.; Saito, N.; Yoshida, K.-I.; Ashida, H. Subcellular localization of flavonol aglycone in hepatocytes visualized by confocal laser scanning fluorescence microscope. Cytotechnology 2009, 59, 177. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, B.T.; Thompson, E.P. A method for visualizing fluorescence of flavonoid therapeutics in vivo in the model eukaryote Dictyostelium discoideum. BioTechniques 2019, 66, 65. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Chen, Y.; Seto, C.T. Rational design of novel irreversible inhibitors for human arginase. Bioorg. Med. Chem. 2018, 26, 3939. [Google Scholar] [CrossRef]
- Hochstedler, C.M.; Leidinger, M.R.; Maher-Sturm, M.T.; Gibson-Corley, K.N.; Meyerholz, D.K. Immunohistochemical detection of arginase-I expression in formalin-fixed lung and other tissues. J. Histotechnol. 2013, 36, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berninger, T.; Bliem, C.; Piccinini, E.; Azzaroni, O.; Knoll, W. Cascading reaction of arginase and urease on a graphene-based FET for ultrasensitive, real-time detection of arginine. Biosens. Bioelectron. 2018, 115, 104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, J.; Li, P.; Wang, X.; Bi, S.; Zhang, J.; Zhang, W.; Wang, H.; Tang, B. In situ and real-time imaging of superoxide anion and peroxynitrite elucidating arginase 1 nitration aggravating hepatic ischemia-reperfusion injury. Biomaterials 2019, 225, 119499. [Google Scholar] [CrossRef] [PubMed]
- Weissleder, R. Molecular Imaging: Principles and Practice; People’s Medical Publishing House: Shelton, CT, USA, 2010. [Google Scholar]
- Wang, Y.; Lu, M.; Tang, D. Novel photoluminescence enzyme immunoassay based on supramolecular host-guest recognition using L-arginine/6-aza-2-thiothymine-stabilized gold nanocluster. Biosens. Bioelectron. 2018, 109, 70. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.-H.; Shi, X.-Q.; Peng, H.-P.; Zhuang, Q.-Q.; Yang, Y.; Liu, A.-L.; Xia, X.-H.; Chen, W. Gold Nanoparticle-Based Photoluminescent Nanoswitch Controlled by Host–Guest Recognition and Enzymatic Hydrolysis for Arginase Activity Assay. ACS Appl. Mater. Interfaces 2018, 10, 5358. [Google Scholar] [CrossRef]
- Aalipour, A.; Chuang, H.-Y.; Murty, S.; D’Souza, A.L.; Park, S.-m.; Gulati, G.S.; Patel, C.B.; Beinat, C.; Simonetta, F.; Martinić, I.; et al. Engineered immune cells as highly sensitive cancer diagnostics. Nat. Biotechnol. 2019, 37, 531. [Google Scholar] [CrossRef]
- Fu, Q.; Zhu, R.; Song, J.; Yang, H.; Chen, X. Photoacoustic Imaging: Contrast Agents and Their Biomedical Applications. Adv. Mater. 2019, 31, 1805875. [Google Scholar] [CrossRef]
- Razgulin, A.; Ma, N.; Rao, J. Strategies for in vivo imaging of enzyme activity: An overview and recent advances. Chem. Soc. Rev. 2011, 40, 4186. [Google Scholar] [CrossRef]
- Yan, R.; Ye, D. Molecular imaging of enzyme activity in vivo using activatable probes. Sci. Bull. 2016, 61, 1672. [Google Scholar] [CrossRef]
- Oldham, M.S.; VanMeter, J.W.; Shattuck, K.F.; Cederbaum, S.D.; Gropman, A.L. Diffusion tensor imaging in arginase deficiency reveals damage to corticospinal tracts. Pediatr. Neurol. 2010, 42, 49. [Google Scholar] [CrossRef] [Green Version]
- Bitner, B.R.; Brink, D.C.; Mathew, L.C.; Pautler, R.G.; Robertson, C.S. Impact of arginase II on CBF in experimental cortical impact injury in mice using MRI. J. Cereb. Blood Flow Metab. 2010, 30, 1105. [Google Scholar] [CrossRef] [Green Version]
- Hingorani, D.V.; Yoo, B.; Bernstein, A.S.; Pagel, M.D. Detecting enzyme activities with exogenous MRI contrast agents. Chem. Eur. J. 2014, 20, 9840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roeda, D.; Crouzel, C.; Brouillet, E.; Valette, H. Synthesis and in vivo distribution of no-carrier-added N(omega)-Nitro-L-arginine [11C]methyl ester, a nitric oxide synthase inhibitor. Nucl. Med. Biol. 1996, 23, 509. [Google Scholar] [CrossRef]
- Brouillet, E.; Roeda, D.; Valette, H.; Fuseau, C.; Guyot, M.C.; Crouzel, C. In vivo metabolites of N omega-nitro-L-arginine methyl ester: Methanol and N omega-nitro-L-arginine. Eur. J Pharmacol. 1995, 293, 487. [Google Scholar] [CrossRef]
- Pomper, M.G.; Musachio, J.L.; Scheffel, U.; Macdonald, J.E.; McCarthy, D.J.; Reif, D.W.; Villemagne, V.L.; Yokoi, F.; Dannals, R.F.; Wong, D.F. Radiolabeled neuronal nitric oxide synthase inhibitors: Synthesis, in vivo evaluation, and primate PET studies. J. Nucl. Med. 2000, 41, 1417. [Google Scholar] [PubMed]
- de Vries, E.F.J.; Vroegh, J.; Dijkstra, G.; Moshage, H.; Elsinga, P.H.; Jansen, P.L.M.; Vaalburg, W. Synthesis and evaluation of a fluorine-18 labeled antisense oligonucleotide as a potential PET tracer for iNOS mRNA expression. Nucl. Med. Biol. 2004, 31, 605. [Google Scholar] [CrossRef] [PubMed]
- Haibin, T.; Zhenghong, L. Radiosynthesis of 8-Fluoro-3-(4-[18F]Fluorophenyl)-3,4-Dihydro-1-Isoquinolinamine ([18F]FFDI), a Potential PET Radiotracer for the Inducible Nitric Oxide Synthase. Curr. Radiopharm. 2008, 1, 49. [Google Scholar]
- Herrero, P.; Laforest, R.; Shoghi, K.; Zhou, D.; Ewald, G.; Pfeifer, J.; Duncavage, E.; Krupp, K.; Mach, R.; Gropler, R. Feasibility and dosimetry studies for 18F-NOS as a potential PET radiopharmaceutical for inducible nitric oxide synthase in humans. J. Nucl. Med. 2012, 53, 994. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.J.; Isakow, W.; Byers, D.E.; Engle, J.T.; Griffin, E.A.; Kemp, D.; Brody, S.L.; Gropler, R.J.; Miller, J.P.; Chu, W.; et al. Imaging pulmonary inducible nitric oxide synthase expression with PET. J. Nucl. Med. 2015, 56, 76. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, B.L.; Morais, M.; Mendes, F.; Moreira, I.S.; Cordeiro, C.; Fernandes, P.A.; Ramos, M.J.; Alberto, R.; Santos, I.; Correia, J.D. Re(I) and Tc(I) complexes for targeting nitric oxide synthase: Influence of the chelator in the affinity for the enzyme. Chem. Biol. Drug Des. 2015, 86, 1072. [Google Scholar] [CrossRef]
- Mori, S. Primary assimilation process of triply (15N, 14C and 3H) labeled arginine in the roots of arginine-fed barley. J. Soil Sci. Plant. Nutr. 1981, 27, 29. [Google Scholar] [CrossRef]
- Castillo, L.; DeRojas-Walker, T.; Yu, Y.M.; Sanchez, M.; Chapman, T.E.; Shannon, D.; Tannenbaum, S.; Burke, J.F.; Young, V.R. Whole body arginine metabolism and nitric oxide synthesis in newborns with persistent pulmonary hypertension. Pediatr. Res. 1995, 38, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubec, B.; Ya-hua, Z.; Pertti, S.; Pentti, T.; Kitzmüller, E.; Lubec, G. Distribution and disappearance of the radiolabeled carbon derived from L-arginine and taurine in the mouse. Life Sci. 1997, 60, 2373. [Google Scholar] [CrossRef]
- Knowles, R.G.; Salter, M. Measurement of NOS activity by conversion of radiolabeled arginine to citrulline using ion-exchange separation. Methods Mol. Biol. 1998, 100, 67. [Google Scholar]
- Ong, S.-E.; Kratchmarova, I.; Mann, M. Properties of 13C-Substituted Arginine in Stable Isotope Labeling by Amino Acids in Cell Culture (SILAC). J. Proteome. Res. 2003, 2, 173. [Google Scholar] [CrossRef]
- Borek, W.E.; Zou, J.; Rappsilber, J.; Sawin, K.E. Deletion of Genes Encoding Arginase Improves Use of “Heavy” Isotope-Labeled Arginine for Mass Spectrometry in Fission Yeast. PLoS ONE 2015, 10, e0129548. [Google Scholar] [CrossRef]
- Huang, C.; McConathy, J. Radiolabeled amino acids for oncologic imaging. J. Nucl. Med. 2013, 54, 1007. [Google Scholar] [CrossRef] [Green Version]
- Rempel, B.P.; Price, E.W.; Phenix, C.P. Molecular Imaging of Hydrolytic Enzymes Using PET and SPECT. Mol. Imaging 2017, 16, 1536012117717852. [Google Scholar] [CrossRef] [Green Version]
- Mei, H.; Han, J.; Klika, K.D.; Izawa, K.; Sato, T.; Meanwell, N.A.; Soloshonok, V.A. Applications of fluorine-containing amino acids for drug design. Eur. J. Med. Chem. 2020, 186, 111826. [Google Scholar] [CrossRef]
- Dittmar, C. Zur trägerarmen Synthese von 4-[18F]Fluorarginin als Leitstruktur potentieller Radiopharmaka der NO-Synthasen-Aktivität. Ph.D. Thesis, Köln University, Köln, Germany, 2000. Available online: http://juser.fz-juelich.de/record/23590/files/Jl_3766.pdf (accessed on 26 May 2020).
- Gao, S.; Tang, G.; Zhu, S.; Hu, K.; Yao, S.; Tang, C.; Yang, C.; Wang, Y.; Li, J.; Pan, X.; et al. Semi-automated radiosynthesis of 18F-labeled l-arginine derivative as a potential PET tracer for lung cancer imaging. J. Radioanal. Nucl. Chem. 2016, 309, 1257. [Google Scholar] [CrossRef]
- Wu, R.; Liu, S.; Liu, Y.; Sun, Y.; Cheng, X.; Huang, Y.; Yang, Z.; Wu, Z. Synthesis and biological evaluation of [18F](2S,4S)4-(3-fluoropropyl) arginine as a tumor imaging agent. Eur. J. Med. Chem. 2019, 183, 111730. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.W.; Long, N.J.; Vilar, R.; Gee, A.D. Synthesis of 11C, 18F, 15O, and 13N radiolabels for positron emission tomography. Angew Chem. Int. Ed. 2008, 47, 8998. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Rong, J.; Wang, L.; Vasdev, N.; Zhang, L.; Josephson, L.; Liang, S.H. Chemistry for Positron Emission Tomography: Recent Advances in 11C-, 18F-, 13N-, and 15O-Labeling Reactions. Angew Chem. Int. Ed. 2019, 58, 2580. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.S.; Windhorst, A.D.; Zeglis, B.M. Radiopharmaceutical Chemistry; Springer International Publishing: Cham, Switzerland, 2019. [Google Scholar]
- Abram, U.; Alberto, R. Technetium and rhenium: Coordination chemistry and nuclear medical applications. J. Braz. Chem. Soc. 2006, 17, 1486. [Google Scholar] [CrossRef]
- Wuest, F.; Berndt, M.; Bergmann, R.; van den Hoff, J.; Pietzsch, J. Synthesis and Application of [18F]FDG-Maleimidehexyloxime ([18F]FDG-MHO): A [18F]FDG-Based Prosthetic Group for the Chemoselective 18F-Labeling of Peptides and Proteins. Bioconjug. Chem. 2008, 19, 1202. [Google Scholar] [CrossRef]
- Gómez-Vallejo, V.; Gaja, V.; Koziorowski, J.; Llop, J. Specific Activity of 11C-Labelled Radiotracers: A Big Challenge for PET Chemists. In Positron Emission Tomography-Current Clinical and Research Aspects; Hsieh, C.-H., Ed.; In Tech: Taichung, Taiwan, 2012; p. 183. [Google Scholar]
- Choi, S.; Park, C.; Ahn, M.; Lee, J.H.; Shin, T. Immunohistochemical study of arginase 1 and 2 in various tissues of rats. Acta Histochem. 2012, 114, 487. [Google Scholar] [CrossRef]
- Clark, J.; O’Hagan, D. Strategies for radiolabelling antibody, antibody fragments and affibodies with fluorine-18 as tracers for positron emission tomography (PET). J. Fluor. Chem. 2017, 203, 31. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathology | Animal/Cell Line Model | Arginase Levels | Proposed Trigger Signal | Proposed Disease Mechanism | Ref. |
|---|---|---|---|---|---|
| Diabetes-induced vasculo-pathy | Bovine aortic endothelial cells exposed to glucose or activated for Arg1 upregulation by adenoviral delivery; Arg1-deficient mouse model | ↑ Arg1 | Glucose treatment activates Rho-associated protein kinases, which induce macrophages to upregulate Arg1 | Substrate depletion by Arg1 reduces NO• and leads to impaired vascular relaxation, increased blood flow, and upsurge of reactive oxygen species, which causes premature endothelial cell senescence and defective vascular repair | [58] |
| Diabetic mouse model; blood samples from diabetic patients | Increased plasma glucose levels induce the release of Arg1 via serum exosomes | [59] | |||
| Mice induced to diabetes by streptozotocin; bovine retinal endothelial cells | High glucose levels activate NOX2 leading to upregulated Arg1 | [60] | |||
| Obesity-induced vasculo-pathy | Diet-induced obesity and metabolic syndrome mouse model | ↑ Arg1 | High-fat, high-sucrose treatment activates Rho-associated protein kinases, which increases Arg1 expression | Upregulated synthesis of polyamines by Arg1 promotes cell proliferation and fibrosis; increased levels of reactive oxygen species contribute to dysfunction | [61] |
| Arterial thickening, fibrosis, and stiffening | Arg1-deficient mouse model; rat aortic smooth muscle cells | ↑ Arg1 | Angiotensin II acts upon the renin–angiotensin system and induces arginase upregulation | Enhanced synthesis of polyamines/proline leads to vascular cell proliferation and collagen formation, which changes smooth muscle tone | [62] |
| Hyper-tension | Obese and lean male rat models | ↑ Arginase1 | Obesity-induced arginase upregulation | L-Arginine depletion reduces NO•-mediated arterial vasodilation | [63] |
| Arterio-genesis | Male mice submitted to peripheral arteriogenesis; mouse primary artery endothelial cells and smooth muscle cells | ↑ Arg1 | Shear stress induces monocytes maturation to macrophages, which impairs M1/M2 to favor Arg1 expression | Enhanced Arg1 activity promotes perivascular M2 macrophage accumulation, which contributes to cell proliferation | [64] |
| Myocardial infarction | Male mouse submitted to surgical ligation of the left anterior descending coronary artery to induce myocardial infarction | ↑ Arg1 | Neutrophils are recruited and infiltrate into the infarcted area, activating the macrophages to favor Arg1 expression | Increased Arg1 activity results in enhanced proline and collagen synthesis, leading to fibrosis, ventricular remodeling, and eventual heart failure | [65] |
| Erectile dysfunction | Patients with a medical diagnosis of erectile dysfunction | ↑ Arg1 and Arg2 | Genetic polymorphisms induce Arg1 and Arg2 expression and activity | L-Arginine depletion leads to endothelial dysfunction and impaired smooth muscle relaxation; erectile dysfunction is an early sign of cardiovascular diseases | [66] |
| Chronic obstructive pulmonary disease | Ex vivo pulmonary vascular tissue from smokers | ↑ Arg1 | Tobacco smoking upregulates the arginase pathway | Imbalance of polyamines/NO• causes vascular remodeling, airway dysfunction, and oxidative stress | [67] |
| Pulmonary hypertension | Human pulmonary artery smooth muscle cell | ↑ Arg2 | Induced hypoxia activates protein kinases and transcription factors leading to the upregulation of Arg2 expression | Increased synthesis of polyamines leads to vascular smooth muscle cell proliferation and remodeling; decreased NO• synthesis impairs vasodilation, which contributes to dysfunction and pulmonary hypertension | [68] |
| Human pulmonary artery smooth muscle cell; male mice exposed to hypoxia | [69] | ||||
| Pulmonary fibrosis | Male mice with bleomycin-induced pulmonary fibrosis | ↑ Arg2 | Pro-inflammatory T helper cells change M1/M2 polarization and increase Arg2 expression | Increased biosynthesis of polyamines and collagen activates lung fibroblast proliferation and differentiation | [70] |
| Primary bronchial cultures from cystic fibrosis patients | ↑ Arginase1 | F508del gene mutation leads to excessive arginase activity in the pulmonary tissue | Increased arginase expression results in a build-up of fibrotic mass; a decrease of NO• levels induces the deregulation of epithelial fluid transport in the lungs and reduce cilia motility | [71] | |
| Cystic fibrosis pediatric patients | High levels of arginase promote collagen deposition and NOS uncoupling, causing oxidative stress and tissue damage | [72] | |||
| Cystic fibrosis patients | ↑ Arg1 | Recessive gene mutation leads to an excessive arginase activity in pulmonary tissue | Reduced NO• impairs smooth muscle relaxation, bronchodilation, and bacterial killing mechanisms | [73] | |
| Asthma | Asthmatic patients | ↑ Arg1 | Allergen activation of IgE leads to neutrophil infiltration in lungs and activation of M2 arginase-expressing macrophages | Upregulation of Arg1 increases mucus production and smooth muscle contraction. Arg1 seems to correlate to bronchial asthma | [74] |
| ↑ Arg2 | Chronic airway inflammations have high co-expression of Arg2 and iNOS | Arg2 delivers L-ornithine into mitochondria, providing nitrogen to an autonomous L-arginine-NO•-citrulline cycle and sustaining high NO• levels, which seems related to more severe and reactive conditions | [75] | ||
| Human bronchial epithelial cell line (BET1A); Arg2-deficient mice with allergen-induced asthma | ↑ Arg2 | Allergens enhance hypoxia-induced factors, which activate IL-13 to upregulate Arg2 | Increased Arg2 is suggested to be a counter-regulatory mechanism to reduce signal transduction and suppress airway inflammation | [76] | |
| Mite-challenged NC/Nga mouse model of asthma | ↑ Arg1 | Allergen activation induces the expression of arginase-upregulating mechanisms | Arginase decreases NO• levels, suppressing anti-inflammatory, bronchodilatory, and vascular modulating effects | [77] | |
| Chronic rhino-sinusitis | Fragments of mucosa collected from the ethmoid sinus of chronic rhinosinusitis patients | ↑ Arg2 | Several cytokines found in the sinus mucosa lead to enhanced arginase expression | Increased Arg2 leads to cell and collagen proliferation and decreases NO• levels, which suppresses bronchodilatory and anti-inflammatory effects | [78] |
| Tuberculosis | Tissue samples from active tuberculosis patients; mouse model infected with Mycobacterium tuberculosis | ↑ Arg1 | Intracellular parasites circumvent NO• toxicity through the induction of Arg1-expressing macrophages in lungs | High Arg1 expression leads to collagen deposition and lung damage, which drives to inflammation by inhibiting type 1 helper T cells | [79] |
| Inflammatory bowel disease | Mouse model of inflammatory bowel disease by dextran sulfate sodium induction | ↓ Arg1 | Extracellular matrix protein 1 (ECM1) in macrophages impairs M1/M2 polarization decreasing the expression of Arg1 | Reduction of Arg1 suppresses tissue repair mechanisms and, together with upregulated expression of inflammatory cytokines, increases chronic inflammatory response | [80] |
| Autoimmune (type 1) diabetes | Diabetic female mouse model induced by hyperglycemia | ↑ Arg1 | Increased plasma glucose levels impair M1/M2 polarization | Decreased NO• levels lead to a pro-inflammatory effect, weakening innate immunity | [81] |
| Arthritis | Synovial tissue samples from rheumatoid arthritis patients; arthritis mouse model (K/BxN) | ↓ Arg1 | Transcription factor Fos-related antigen 1 downregulate Arg1 expression by binding to the promoter region | Reduction of Arg1 suppresses polyamines synthesis and subsequently downregulates tissue repair mechanisms and counter-regulates pro-inflammatory cytokines | [82] |
| Multiple sclerosis | Arg2-knockout mice with induced autoimmune encephalomyelitis | ↑ Arg2 | Impaired M1/M2 macrophage polarization | Upregulated Arg2 stimulates the production of T helper 17 cells-differentiating cytokines, which induces inflammation | [83] |
| Viral infection | Patients with severe fever and thrombocytopenia syndrome | ↑ Arg1 | Viral-induced impairment of M1/M2 polarization favors the upregulation of Arg1 | Arg1 causes L-arginine deficiency, which is associated with decreased NO• and suppresses antiviral immunity | [84] |
| Mice infected with Trypanosoma cruzi and Schistosoma mansoni | [85] | ||||
| Peripheral lymph node cells from HIV patients | [86] | ||||
| Peritonitis | Murine macrophage-like cell line (RAW264.7) and human monocyte cell line (THP-1) | ↑ Arg1 | IL-4-stimulated inflammation upregulates cytochrome P450 1A1, which impairs M1/M2 polarization | Increased Arg1 expression is associated with compensatory response mechanisms against an uncontrolled inflammation | [87] |
| Acute myeloid leukemia | Human acute myeloid leukemia cell lines (THP-1, U937, MOLM16, K562) | ↑ Arg2 | Increased acute myeloid leukemia blast cells overexpressing Arg2 | Arg2 activity reduces IFN-γ and inhibits T cell immune-suppressive response | [88] |
| Chronic myelo-monocytic leukemia | Human bone marrow mononuclear cells | ↑ Arg1 | Mutations in epigenetic regulators upregulate Arg1 | L-Arginine depletion by Arg1 suppress T-cells and contributes to immune evasion | [89] |
| Basal-like breast cancer | Human mammary epithelial cells (HeLa, HMEC, HMEC-ras, MDA-MB-231, MDA-MB-468) | ↑ Arg2 | Oncogene transformations trigger Arg2 expression | Arg2 upregulated between DNA synthesis and mitotic phases of cancer cells cycle promotes cell proliferation | [90] |
| Neuro-blastoma | Neural crest cell line (R1113T); neuroblastoma cell lines (SKNAS, KELLY, LAN-1, IMR-32,); Ewing’s sarcoma cell line (SKNMC); sympathetic ganglion-derived stem cells (SZ16) | ↑ Arg2 | IL-1β and TNF-α established a feedback loop to upregulate Arg2 expression via p38 and extracellular regulated kinases signaling | Arg2 induces cell proliferation and an immunosuppressive microenvironment due to inhibition of T cell cytotoxicity | [91] |
| Pancreatic ductal adeno-carcinoma | Human pancreatic ductal adenocarcinoma cell lines (AsPC-1, HPAC, MIA PaCa-2, PANC-1, SUIT-2, PA-TU-8988T); Arg2-deficient mouse pancreatic ductal adenocarcinoma cell lines | ↑ Arg2 | Arg2 is increased upon obesity and as a result of activating oncogenic mutations | Tumors (but not cultured cancer cells lacking the in vivo tumor microenvironment) need arginase to dispose of the excess of nitrogen accumulated to enhance tumorigenicity | [92] |
| Melanoma | Patient with metastatic L-arginine auxotrophic melanoma | ↑ Arg2 | Defects in the expression of OTC and ASS enzymes result in a dependence of extracellular L-arginine; counter-regulatory mechanisms lead to the upregulation of Arg2 | Tumor cells were shown to be auxotrophic and avid for L-arginine to keep cell proliferation; high expression of Arg2 is induced to increase catalytic efficiency | [93] |
| Human melanoma cell lines from patients with melanoma metastasis adhered to confluent human umbilical vein endothelial cells layers | Pro-inflammatory T helper cells change M1/M2 polarization and increase Arg2 expression | Arg2 enhances melanoma cell proliferation through polyamine production and promotes metastasis through enhancing H2O2 production and STAT3 signaling | [94] | ||
| Ovarian carcinoma | Human ovarian cancer cell lines (OVP-10, AD-10, A2780, Skov3, CaOv-3, MDAH2774, OvCa-14) | ↑ Arg1 | Tumor-derived exosomes containing Arg1 are released into circulation | Increased Arg1 expression inhibits antigen-specific T-cell proliferation and is related to a worse prognosis | [95] |
| Osteosarcoma | Human osteosarcoma cell lines (SaOS-2 and OS-17) | ↑ Arg2 | Hypoxic environment upregulates Arg2 | Arg2 induces immunosuppression by inhibition of T-cells function | [96] |
| Glioma | Mouse glioma cell lines (GL261, KR158B) | ↑ Arg1 | Myeloid-derived suppressor cells overexpressing Arg1 infiltrate into the tumor | Increased Arg1 expression suppresses the efficacy of the immune system | [97] |
| Hepato-cellular carcinoma | Human hepatocellular carcinoma cell line (Huh7) | ↑ Arg1 | Impaired M1/M2 polarization induces Arg1 upregulation | Overexpression of Arg1 promotes cell proliferation, migration, and invasion, being a critical process in cancer metastasis and progression | [98] |
| Patients with advanced hepatocellular carcinoma | Deprivation of L-arginine recycling enzymes OTC and ASS at the transcription or translational level | Tumor auxotrophic for L-arginine to enable cell proliferation and viability; L-arginine deprivation therapy can be a therapeutic approach | [99] | ||
| Cervical cancer | Human squamous cell carcinoma cells from patients | ↑ Arginase1 | Increased levels of circulating IL-10 and decreased levels of IFN-γ enhance arginase activity | Upregulated arginase levels contribute to the tumor immunosuppressive microenvironment | [100] |
| Alzheimer | Alzheimer’s disease mouse models | ↑ Arg1 and Arg2 | Microglial activation results in cytokines production, which induces the expression of arginase in brain | Arginase overexpression at β-amyloid deposition sites leads to NOS uncoupling, O2•− generation, and neuro-degenerative oxidative stress | [101] |
| Acute traumatic brain injury | Male rats submitted to traumatic brain injury surgery | ↑ Arg1 | Elevation of pro-inflammatory cytokines induces Arg1 expression | Increased Arg1 leads to eNOS uncoupling and enhances oxidative stress, inflammation, and vascular dysfunction | [102] |
| Fronto-temporal dementia | Male transgenic mice expressing a mutant form of human microtubule-associated protein tau | ↑ Arginase1 | Mutations in microtubule-associated protein tau | Functional significance of arginase remains uncertain as the production of polyamines enhances microtubule stability, which should reduce inflammation and tau proteins | [103] |
| Neuro-degeneration and neuro-vascular permeability | Male mice treated with homocysteine to induce vascular dysfunction and stroke-like symptoms | ↓ Arginase1 | Elevated levels of homocysteine, produced from methionine, lead to hyperhomocysteinemia, impairing NOS pathway | Upregulated NO• levels lead to nitrosative stress, extracellular matrix degradation, blood–brain barrier permeability, and neurodegeneration | [104] |
| Huntington’s disease | Post-mortem brain sections from patients with Huntington’s disease | ↑ Arg1 | Metabolic impairment of the urea cycle in the brain | Increased urea in the brain induces neurodegeneration by impaired osmoregulation | [105] |
| Acute ischemic stroke | Peripheral blood samples from patients with a first-ever acute ischemic stroke | ↑ Arg1 | Stroke induces the downregulation of a microRNA, which upregulates the Arg1 expression | Increased Arg1 acts against the activation of pro-inflammatory signals after stroke but may also be implicated in stroke-induced immunosuppression | [106] |
| Cerebral ischemia and excitotoxicity | Arg2-knockout mice with permanent distal middle cerebral artery occlusion or induced excitotoxicity | ↓ Arg2 | Arg2 deficiency worsens brain injury after an ischemic event | Arg2 may play a substantial protective role by regulating NO• levels and controlling reactive species | [107] |
| Technique | Imaging Agent | Spatial Resolution | Detection Sensitivity | Penetration Depth | Quantification Efficiency |
|---|---|---|---|---|---|
| Optical | Fluorophores or lanthanides | 2–3 mm | 10−11 mol·L−1 | <20 mm | Medium |
| Photoacoustic | Light absorbing agents | 0.1–1 mm | 10−11 mol·L−1 | <70 mm | Medium |
| Ultrasound | Gas microbubbles | 0.5–1 mm | 10−8 mol·L−1 | <200 mm | Low |
| MRI | (Super)para-magnetic agents | 0.03–1 mm | 10−5 mol·L−1 | >300 mm | Medium |
| CT | I or Ba agents | 0.03–1 mm | 0.1 mol·L−1 | >300 mm | − |
| PET | β+-emitters | 1–10 mm | 10−12 mol·L−1 | >300 mm | High |
| SPECT | γ-emitters | 0.5–15 mm | 10−11 mol·L−1 | >300 mm | High |
| Modality | Radio-nuclide | Physical Half-Life | Production | Target /Parent Isotope | Primary Precursor | Main Emissions |
|---|---|---|---|---|---|---|
| SPECT | 99mTc | 6.01 h | Generator (99Mo/99mTc) | 99Mo, parent isotope | [99mTc]TcO4Na | γ, 141 keV |
| 111In | 2. 81 d | Cyclotron 111Cd(p,n)111In 112Cd(p,2n)111In | 111/112Cd-enriched sample | [111In]InCl | γ, 245, 171 keV | |
| 67Ga | 78.3 h | Cyclotron (68Zn(p, 2n)67Ga) | 68Zn-enriched sample | [67Ga]GaCl3 | γ, 300, 181, 93 keV | |
| 123I | 13.2 h | Cyclotron (124Te(p, 2n)123I) | 124Te-enriched sample | [123I]I2 | γ, 159 keV | |
| PET | 13N | 9.97 min | Cyclotron 16O(p, α)13N | H2O H2O+ethanol | [13N]NO2/3 [13N]NH3 | 100% β+, 1200 keV |
| 11C | 20.4 min | Cyclotron 14N(p,α)11C | N2(+O2) N2(+H2) | [11C]CO2 [11C]CH4 | 99% β+, 960 keV | |
| 18F | 109.8 min | Cyclotron 20Ne(d,α)18F 18O(p,n)18F | Ne(+F2) [18O]H2O | [18F]F2 [18F]F− aq. | 97% β+, 630 keV; 3% electron capture | |
| 68Ga | 68 min | Generator (68Ge/68Ga) | 68Ge, parent isotope | [68Ga]GaCl3 | 90% β+, 1830 keV; 10% electron capture | |
| 64Cu | 12.7 h | Cyclotron 64Ni(p,n)64Cu | 64Ni-enriched sample | [64Cu]CuCl2 | 18% β+, 650 keV; 39% β−, 579 keV; 43% electron capture | |
| 89Zr | 78.4 h | Cyclotron 89Y(p,n)89Zr | 89Y-enriched sample | [89Zr]Zr(C2O4)2 | 77% electron capture; 23% β+, 902 keV |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
S. Clemente, G.; van Waarde, A.; F. Antunes, I.; Dömling, A.; H. Elsinga, P. Arginase as a Potential Biomarker of Disease Progression: A Molecular Imaging Perspective. Int. J. Mol. Sci. 2020, 21, 5291. https://doi.org/10.3390/ijms21155291
S. Clemente G, van Waarde A, F. Antunes I, Dömling A, H. Elsinga P. Arginase as a Potential Biomarker of Disease Progression: A Molecular Imaging Perspective. International Journal of Molecular Sciences. 2020; 21(15):5291. https://doi.org/10.3390/ijms21155291
Chicago/Turabian StyleS. Clemente, Gonçalo, Aren van Waarde, Inês F. Antunes, Alexander Dömling, and Philip H. Elsinga. 2020. "Arginase as a Potential Biomarker of Disease Progression: A Molecular Imaging Perspective" International Journal of Molecular Sciences 21, no. 15: 5291. https://doi.org/10.3390/ijms21155291
APA StyleS. Clemente, G., van Waarde, A., F. Antunes, I., Dömling, A., & H. Elsinga, P. (2020). Arginase as a Potential Biomarker of Disease Progression: A Molecular Imaging Perspective. International Journal of Molecular Sciences, 21(15), 5291. https://doi.org/10.3390/ijms21155291