Heat Shock Protein 70 as a Double Agent Acting Inside and Outside the Cell: Insights into Autoimmunity


{kind=link}
Abstract
:1. Introduction
2. Hsp70: Structure, Expression, and Mechanism of Action
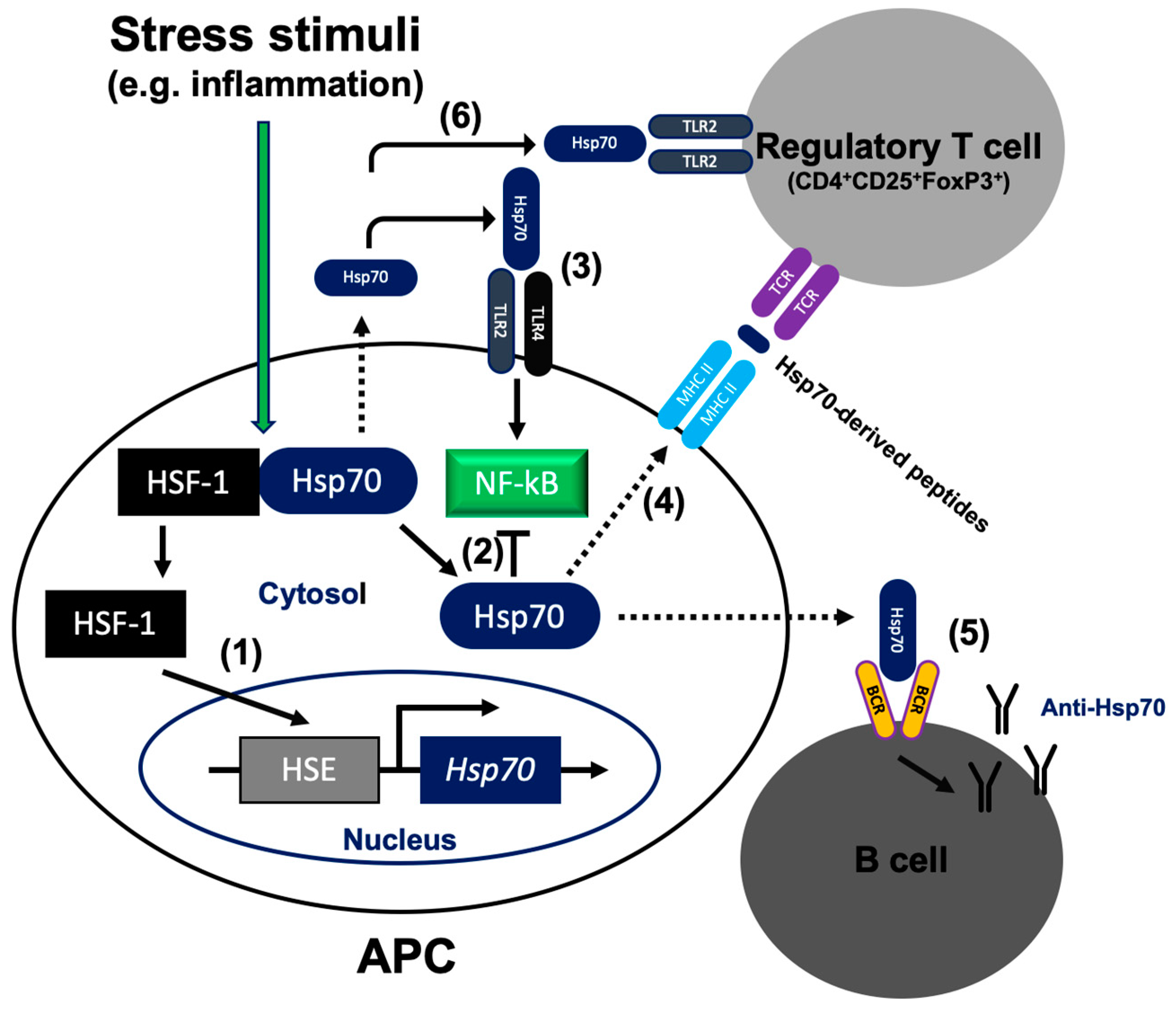
3. Hsp70 Modulates NF-κB Activation
4. Hsp70 and Cancer
5. Hsp70 Is Present Outside the Cell
6. Extracellular Hsp70 Activates the Humoral Autoimmune Response
7. Hsp70 Promotes Regulatory T Cells
8. Clinical Perspectives
9. Conclusions
Funding
Conflicts of Interest
Abbreviations
| 17-DMAG | 17-Dimethylaminoethylamino-17-demethoxygeldanamycin |
| 17AAG | 17-N-allylamino-17-demethoxygeldanamycin |
| ADP | Adenosine diphosphate |
| AA | Adjuvant-induced arthritis |
| AKT | Protein Kinase B |
| APC | Antigen-presenting cell |
| ATP | Adenosine triphosphate |
| BCR | B cell receptor |
| BiP | Binding immunoglobulin protein |
| CIA | Collagen-induced arthritis |
| CRP | C-reactive protein |
| DAMP | Damage-associated molecular patterns |
| DC | Dendritic cell |
| ER | Endoplasmic reticulum |
| GA | Geldanamycin |
| HSE | Heat shock element |
| HSF | Heat shock factor |
| Hsp | Heat shock protein |
| Ig | Immunoglobulin |
| IkB | Inhibitor of nuclear factor kappa B |
| IKK | I kappa B kinase |
| IL | Interleukin |
| JNK | c-Jun N-terminal kinases |
| kDa | Kilodalton |
| LPS | Lipopolysaccharides |
| mAb | Monoclonal antibodies |
| MCP-1 | Monocyte chemoattractant protein 1 |
| MHC | Major histocompatibility complex |
| NBD | Nucleotide binding domain |
| NEF | Nucleotide exchange factor |
| NF-κB | Nuclear Factor-kappa B |
| NK | Natural killer |
| p38 | Mitogen-activated protein kinase p38 |
| PAMP | Pathogen-associated molecular patterns |
| PBMC | Human peripheral blood mononuclear cell |
| PI3K | Phosphoinositide 3-kinases |
| PRR | Pattern recognition receptor |
| RA | Rheumatoid arthritis |
| SBD | Substrate binding domain |
| Tc | Cytotoxic T cell |
| TCR | T cell receptor |
| TGF-β | Transforming growth factor β |
| Th | T helper cell |
| TLR | Toll-like receptors |
| TNF-α | Tumor necrosis factor α |
| Treg | Regulatory T helper cell |
| VEGF | Vascular endothelial growth factor |
References
- Kampinga, H.H.; Hageman, J.; Vos, M.J.; Kubota, H.; Tanguay, R.M.; Bruford, E.A.; Cheetham, M.E.; Chen, B.; Hightower, L.E. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress Chaperones 2008, 14, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saibil, H. Chaperone machines for protein folding, unfolding and disaggregation. Nat. Rev. Mol. Cell Biol. 2013, 14, 630–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomasello, G.; Sciumé, C.; Rappa, F.; Rodolico, V.; Zerilli, M.; Martorana, A.; Cicero, G.; De Luca, R.; Damiani, P.; Accardo, F.M.; et al. Hsp10, Hsp70, and Hsp90 immunohistochemical levels change in ulcerative colitis after therapy. Eur. J. Histochem. 2011, 55, e38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tukaj, S.; Kleszczyński, K.; Vafia, K.; Groth, S.; Meyersburg, D.; Trzonkowski, P.; Ludwig, R.J.; Zillikens, D.; Schmidt, E.; Fischer, T.W.; et al. Aberrant expression and secretion of heat shock protein 90 in patients with bullous pemphigoid. PLoS ONE 2013, 8, e70496. [Google Scholar] [CrossRef] [PubMed]
- Kurzik-Dumke, U.; Schick, C.; Rzepka, R.; Melchers, I. Overexpression of human homologs of the bacterial DnaJ chaperone in the synovial tissue of patients with rheumatoid arthritis. Arthritis Rheum. 1999, 42, 210–220. [Google Scholar] [CrossRef]
- Tukaj, S.; Węgrzyn, G. Anti-Hsp90 therapy in autoimmune and inflammatory diseases: A review of preclinical studies. Cell Stress Chaperones 2016, 21, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Wieten, L.; Broere, F.; van der Zee, R.; Koerkamp, E.K.; Wagenaar, J.; van Eden, W. Cell stress induced HSP are targets of regulatory T cells: A role for HSP inducing compounds as anti-inflammatory immuno-modulators? FEBS Lett. 2007, 581, 3716–3722. [Google Scholar] [CrossRef] [Green Version]
- Wieten, L.; van der Zee, R.; Spiering, R.; Wagenaar-Hilbers, J.; van Kooten, P.J.; Broere, F.; van Eden, W. A novel heat-shock protein coinducer boosts stress protein Hsp70 to activate T cell regulation of inflammation in autoimmune arthritis. Arthritis Rheum. 2010, 62, 1026–1035. [Google Scholar] [CrossRef]
- Wieten, L.; van der Zee, R.; Goedemans, R.; Sijtsma, J.; Serafini, M.; Lubsen, N.H.; van Eden, W.; Broere, F. Hsp70 expression and induction as a readout for detection of immune modulatory components in food. Cell Stress Chaperones 2010, 15, 25–37. [Google Scholar] [CrossRef] [Green Version]
- De Maio, A. Extracellular Hsp70: Export and Function. Curr. Protein Pept. Sci. 2014, 15, 225–231. [Google Scholar] [CrossRef]
- Pockley, A.G.; Henderson, B. Extracellular cell stress (heat shock) proteins—Immune responses and disease: An overview. Philos. Trans. R. Soc. B 2017, 373, 20160522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, T.J.; Wieten, L.; van Herwijnen, M.J.C.; Broere, F.; van der Zee, R.; Bonorino, C.; van Eden, W. The anti-inflammatory mechanisms of Hsp70. Front. Immunol. 2012, 3, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radons, J. The human HSP70 family of chaperones: Where do we stand? Cell Stress Chaperones 2016, 21, 379–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tukaj, S.; Kaminski, M. Heat shock proteins in the therapy of autoimmune diseases: Too simple to be true? Cell Stress Chaperones 2019, 24, 475–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tukaj, S.; Mantej, J.; Sobala, M.; Potrykus, K.; Sitko, K. Autologous extracellular Hsp70 exerts a dual role in rheumatoid arthritis. Cell Stress Chaperones 2020. [Google Scholar] [CrossRef]
- Kasperkiewicz, M.; Tukaj, S.; Gembicki, A.-J.; Silló, P.; Görög, A.; Zillikens, D.; Kárpáti, S. Evidence for a role of autoantibodies to heat shock protein 60, 70, and 90 in patients with dermatitis herpetiformis. Cell Stress Chaperones 2014, 19, 837–843. [Google Scholar] [CrossRef] [Green Version]
- Tukaj, S.; Görög, A.; Kleszczyński, K.; Zillikens, D.; Kárpáti, S.; Kasperkiewicz, M. Autoimmunity to heat shock proteins and vitamin D status in patients with celiac disease without associated dermatitis herpetiformis. J. Steroid Biochem. Mol. Biol. 2017, 173, 23–27. [Google Scholar] [CrossRef]
- Mantej, J.; Polasik, K.; Piotrowska, E.; Tukaj, S. Autoantibodies to heat shock proteins 60, 70, and 90 in patients with rheumatoid arthritis. Cell Stress Chaperones 2018, 24, 283–287. [Google Scholar] [CrossRef]
- van Eden, W.; van der Zee, R.; Prakken, B. Heat-shock proteins induce T-cell regulation of chronic inflammation. Nat. Rev. Immunol. 2005, 5, 318–330. [Google Scholar] [CrossRef] [Green Version]
- Wieten, L.; Berlo, S.E.; Brink, C.B.T.; van Kooten, P.J.; Singh, M.; van der Zee, R.; Glant, T.T.; Broere, F.; van Eden, W. IL-10 Is Critically Involved in Mycobacterial HSP70 Induced Suppression of Proteoglycan-Induced Arthritis. PLoS ONE 2009, 4, e4186. [Google Scholar] [CrossRef] [Green Version]
- van Herwijnen, M.J.C.; Wieten, L.; van der Zee, R.; van Kooten, P.J.; Wagenaar-Hilbers, J.P.; Hoek, A.; den Braber, I.; Anderton, S.M.; Singh, M.; Meiring, H.D.; et al. Regulatory T cells that recognize a ubiquitous stress-inducible self-antigen are long-lived suppressors of autoimmune arthritis. Proc. Natl. Acad. Sci. USA 2012, 109, 14134–14139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Eden, W. Immune tolerance therapies for autoimmune diseases based on heat shock protein T-cell epitopes. Philos. Trans. R. Soc. 2018, 373, 20160531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, M.P.; Gierasch, L.M. Recent advances in the structural and mechanistic aspects of Hsp70 molecular chaperones. J. Biol. Chem. 2019, 294, 2085–2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Åkerfelt, M.; Morimoto, R.I.; Sistonen, L. Heat shock factors: Integrators of cell stress, development and lifespan. Nat. Rev. Mol. Cell Biol. 2010, 11, 545–555. [Google Scholar] [CrossRef]
- Abravaya, K.; Myers, M.P.; Murphy, S.P.; Morimoto, R.I. The human heat shock protein hsp70 interacts with HSF, the transcription factor that regulates heat shock gene expression. Genes Dev. 1992, 6, 1153–1164. [Google Scholar] [CrossRef] [Green Version]
- Vihervaara, A.; Sistonen, L. HSF1 at a glance. J. Cell. Sci. 2014, 127, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Peffer, S.; Gonçalves, D.; Morano, K.A. Regulation of the Hsf1-dependent transcriptome via conserved bipartite contacts with Hsp70 promotes survival in yeast. J. Biol. Chem. 2019, 294, 12191–12202. [Google Scholar] [CrossRef]
- Naidu, S.D.; Dinkova-Kostova, A.T. Regulation of the mammalian heat shock factor 1. FEBS J. 2017, 284, 1606–1627. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Velasco; Dublang; Moro; Muga The Complex Phosphorylation Patterns that Regulate the Activity of Hsp70 and Its Cochaperones. Int. J. Mol. Sci. 2019, 20, 4122. [CrossRef] [Green Version]
- Assimon, V.A.; Gillies, A.T.; Rauch, J.N.; Gestwicki, J.E. Hsp70 Protein Complexes as Drug Targets. Curr. Pharm. Des. 2013, 19, 404–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, R.; Baltimore, D. Inducibility of κ immunoglobulin enhancer-binding protein NF-κB by a posttranslational mechanism. Cell 1986, 47, 921–928. [Google Scholar] [CrossRef]
- May, M.J.; Ghosh, S. Signal transduction through NF-κB. Immunol. Today 1998, 19, 80–88. [Google Scholar] [CrossRef]
- Christian, F.; Smith, E.; Carmody, R. The Regulation of NF-κB Subunits by Phosphorylation. Cells 2016, 5, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayden, M.S.; West, A.P.; Ghosh, S. NF-κB and the immune response. Oncogene 2006, 25, 6758–6780. [Google Scholar] [CrossRef] [Green Version]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Hayden, M.S. Signaling to NF-κB. Genes Dev. 2004, 18, 2195–2224. [Google Scholar] [CrossRef] [Green Version]
- Ran, R. Hsp70 promotes TNF-mediated apoptosis by binding IKK and impairing NF-κB survival signaling. Genes Dev. 2004, 18, 1466–1481. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Wu, Y.; Zhang, Y.; Jin, L.; Luo, L.; Xue, B.; Lu, C.; Zhang, X.; Yin, Z. Hsp70 inhibits lipopolysaccharide-induced NF-κB activation by interacting with TRAF6 and inhibiting its ubiquitination. FEBS Lett. 2006, 580, 3145–3152. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.H.; Chou, P.C.; Chung, F.T.; Lin, H.C.; Huang, K.H.; Kuo, H.P. Heat shock protein70 is implicated in modulating NF-κB activation in alveolar macrophages of patients with active pulmonary tuberculosis. Sci. Rep. 2017, 7, 1214. [Google Scholar] [CrossRef] [PubMed]
- Feinstein, D.L.; Galea, E.; Reis, D.J. Suppression of Glial Nitric Oxide Synthase Induction by Heat Shock: Effects on Proteolytic Degradation of IκB-α. Nitric Oxide 1997, 1, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Guzhova, I.V.; Darieva, Z.A.; Melo, A.R.; Margulis, B.A. Major stress protein Hsp70 interacts with NF-κB regulatory complex in human T-lymphoma cells. Cell Stress Chaperones 1997, 2, 132. [Google Scholar] [CrossRef] [Green Version]
- Curry, H.A.; Clemens, R.A.; Shah, S.; Bradbury, C.M.; Botero, A.; Goswami, P.; Gius, D. Heat Shock Inhibits Radiation-induced Activation of NF-κB via Inhibition of I-κB Kinase. J. Biol. Chem. 1999, 274, 23061–23067. [Google Scholar] [CrossRef] [Green Version]
- Andrés, D.; Díez-Fernández, C.; Castrillo, A.; Cascales, M. Relationship between the activation of heat shock factor and the suppression of nuclear factor-κB activity in rat hepatocyte cultures treated with cyclosporine A. Biochem. Pharmacol. 2002, 64, 247–256. [Google Scholar] [CrossRef]
- Malhotra, V.; Wong, H.R. Interactions between the heat shock response and the nuclear factor-κB signaling pathway. Crit. Care Med. 2002, 30, S89–S95. [Google Scholar] [CrossRef] [PubMed]
- Hulina, A.; Rajković, M.G.; Despot, D.J.; Jelić, D.; Dojder, A.; Čepelak, I.; Rumora, L. Extracellular Hsp70 induces inflammation and modulates LPS/LTA-stimulated inflammatory response in THP-1 cells. Cell Stress Chaperones 2017, 23, 373–384. [Google Scholar] [CrossRef]
- Asea, A.; Kraeft, S.K.; Kurt-Jones, E.A.; Stevenson, M.A.; Chen, L.B.; Finberg, R.W.; Koo, G.C.; Calderwood, S.K. HSP70 stimulates cytokine production through a CD14-dependant pathway, demonstrating its dual role as a chaperone and cytokine. Nat. Med. 2000, 6, 435–442. [Google Scholar] [CrossRef]
- Fang, H.; Wu, Y.; Huang, X.; Wang, W.; Ang, B.; Cao, X.; Wan, T. Toll-like Receptor 4 (TLR4) Is Essential for Hsp70-like Protein 1 (HSP70L1) to Activate Dendritic Cells and Induce Th1 Response. J. Biol. Chem. 2011, 286, 30393–30400. [Google Scholar] [CrossRef] [Green Version]
- Somensi, N.; Brum, P.O.; De Miranda Ramos, V.; Gasparotto, J.; Zanotto-Filho, A.; Rostirolla, D.C.; Da Silva Morrone, M.; Moreira, J.C.F.; Gelain, D.P. Extracellular HSP70 Activates ERK1/2, NF-κB and Pro-Inflammatory Gene Transcription Through Binding with RAGE in A549 Human Lung Cancer Cells. Cell. Physiol. Biochem. 2017, 42, 2507–2522. [Google Scholar] [CrossRef]
- Luo, X.; Zuo, X.; Zhou, Y.; Zhang, B.; Shi, Y.; Liu, M.; Wang, K.; Mcmillian, D.R.; Xiao, X. Extracellular heat shock protein 70 inhibits tumour necrosis factor-α induced proinflammatory mediator production in fibroblast-like synoviocytes. Arthritis Res. Ther. 2008, 10, R41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferat-Osorio, E.; Sánchez-Anaya, A.; Gutiérrez-Mendoza, M.; Boscó-Gárate, I.; Wong-Baeza, I.; Pastelin-Palacios, R.; Pedraza-Alva, G.; Bonifaz, L.C.; Cortés-Reynosa, P.; Pérez-Salazar, E.; et al. shock protein 70 down-regulates the production of toll-like receptor-induced pro-inflammatory cytokines by a heat shock factor-1/constitutive heat shock element-binding factor-dependent mechanism. J. Inflamm. (Lond.) 2014, 11, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stokes, J.; Singh, U.P.; Gunn, K.S.; Acharya, A.; Manne, U.; Mishra, M. Targeting Hsp70: A possible therapy for cancer. Cancer Lett. 2016, 374, 156–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.E. The HSP70 family and cancer. Carcinogenesis 2013, 34, 1181–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baehrecke, E.H. How death shapes life during development. Nat. Rev. Mol. Cell Biol. 2002, 3, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Specht, H.M.; Ahrens, N.; Blankenstein, C.; Duell, T.; Fietkau, R.; Gaipl, U.S.; Günther, C.; Gunther, S.; Habl, G.; Hautmann, H.; et al. Heat Shock Protein 70 (Hsp70) Peptide Activated Natural Killer (NK) Cells for the Treatment of Patients with Non-Small Cell Lung Cancer (NSCLC) after Radiochemotherapy (RCTx)—From Preclinical Studies to a Clinical Phase II Trial. Front. Immunol. 2015, 6, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albakova, Z.; Armeev, G.A.; Kanevskiy, L.M.; Kovalenko, E.I.; Sapozhnikov, A.M. HSP70 Multi-Functionality in Cancer. Cells 2020, 9, 587. [Google Scholar] [CrossRef] [Green Version]
- Costa, T.E.; Raghavendra, N.M.; Penido, C. Natural heat shock protein 90 inhibitors in cancer and inflammation. Eur. J. Med. Chem. 2020, 189, 112063. [Google Scholar] [CrossRef]
- Collins, C.B.; Aherne, C.M.; Yeckes, A.; Pound, K.; Eltzschig, H.K.; Jedlicka, P.; Zoeten, E.F.D. Inhibition of N-terminal ATPase on HSP90 attenuates colitis through enhanced Treg function. Mucosal. Immunol. 2013, 6, 960–971. [Google Scholar] [CrossRef] [Green Version]
- Tukaj, S.; Tiburzy, B.; Manz, R.; Marques, A.D.C.; Orosz, A.; Ludwig, R.J.; Zillikens, D.; Kasperkiewicz, M. Immunomodulatory effects of heat shock protein 90 inhibition on humoral immune responses. Exp. Dermatol. 2014, 23, 585–590. [Google Scholar] [CrossRef]
- Tukaj, S.; Bieber, K.; Kleszczyński, K.; Witte, M.; Cames, R.; Kalies, K.; Zillikens, D.; Ludwig, R.J.; Fischer, T.W.; Kasperkiewicz, M. Topically Applied Hsp90 Blocker 17AAG Inhibits Autoantibody-Mediated Blister-Inducing Cutaneous Inflammation. J. Investig. Dermatol. 2017, 137, 341–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hightower, L.E.; Guidon, P.T. Selective release from cultured mammalian cells of heat-shock (stress) proteins that resemble glia-axon transfer proteins. J. Cell. Physiol. 1989, 138, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Zou, G.; Huang, Y.; Xu, G.; Xu, F.; He, J.; Zhu, H.; Yu, P. Serum levels of HSP70 and other DAMP proteins can aid in patient diagnosis after traumatic injury. Cell Stress Chaperones 2016, 21, 677–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murshid, A.; Theriault, J.; Gong, J.; Calderwood, S.K. Molecular Chaperone Receptors. Methods Mol. Biol. 2018, 1709, 331–344. [Google Scholar] [PubMed]
- Broere, F.; van der Zee, R.; van Eden, W. Heat shock proteins are no DAMPs, rather DAMPERs. Nat. Rev. Immunol. 2011, 11, 565. [Google Scholar] [CrossRef] [Green Version]
- Rigo, M.M.; Borges, T.J.; Lang, B.J.; Murshid, A.; Nitika; Wolfgeher, D.; Calderwood, S.K.; Truman, A.W.; Bonorino, C. Host expression system modulates recombinant Hsp70 activity through post-translational modifications. FEBS J. 2020. [Google Scholar] [CrossRef]
- Jensen, P.E. Recent advances in antigen processing and presentation. Nat. Immunol. 2007, 8, 1041–1048. [Google Scholar] [CrossRef]
- Paludan, C. Endogenous MHC Class II Processing of a Viral Nuclear Antigen After Autophagy. Science 2005, 307, 593–596. [Google Scholar] [CrossRef]
- Stocki, P.; Morris, N.J.; Preisinger, C.; Wang, X.N.; Kolch, W.; Multhoff, G.; Dickinson, A.M. Identification of potential HLA class I and class II epitope precursors associated with heat shock protein 70 (HSPA). Cell Stress Chaperones 2010, 15, 729–741. [Google Scholar] [CrossRef] [Green Version]
- Stocki, P.; Wang, X.N.; Morris, N.J.; Dickinson, A.M. HSP70 Natively and Specifically Associates with an N-terminal Dermcidin-derived Peptide That Contains an HLA-A*03 Antigenic Epitope. J. Biol. Chem. 2011, 286, 12803–12811. [Google Scholar] [CrossRef] [Green Version]
- Deffit, S.N.; Blum, J.S. A central role for HSC70 in regulating antigen trafficking and MHC class II presentation. Mol. Immunol. 2015, 68, 85–88. [Google Scholar] [CrossRef] [Green Version]
- Pockley, A.G.; Shepherd, J.; Corton, J.M. Detection of heat shock protein 70 (HSP70) and anti-HSP70 antibodies in the serum of normal individuals. Immunol. Investig. 1998, 27, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Tukaj, S.; Kotlarz, A.; Jozwik, A.; Smolenska, Z.; Bryl, E.; Witkowski, J.M.; Lipinska, B. Hsp40 proteins modulate humoral and cellular immune response in rheumatoid arthritis patients. Cell Stress Chaperones 2010, 15, 555–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tukaj, S.; Kotlarz, A.; Jóźwik, A.; Smoleńska, Z.; Bryl, E.; Witkowski, J.M.; Lipińska, B. Cytokines of the Th1 and Th2 type in sera of rheumatoid arthritis patients; correlations with anti-Hsp40 immune response and diagnostic markers. Acta. Biochim. Pol. 2010, 57, 327–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najafizadeh, S.R.; Ghazizadeh, Z.; Nargesi, A.A.; Mahdavi, M.; Abtahi, S.; Mirmiranpour, H.; Nakhjavani, M. Analysis of serum heat shock protein 70 (HSPA1A) concentrations for diagnosis and disease activity monitoring in patients with rheumatoid arthritis. Cell Stress Chaperones 2015, 20, 537–543. [Google Scholar] [CrossRef]
- Ulmansky, R.; Cohen, C.J.; Szafer, F.; Moallem, E.; Fridlender, Z.G.; Kashi, Y.; Naparstek, Y. Resistance to adjuvant arthritis is due to protective antibodies against heat shock protein surface epitopes and the induction of IL-10 secretion. J. Immunol. 2002, 168, 6463–6469. [Google Scholar] [CrossRef]
- Satpute, S.R.; Rajaiah, R.; Polumuri, S.K.; Moudgil, K.D. Tolerization with Hsp65 induces protection against adjuvant-induced arthritis by modulating the antigen-directed interferon-gamma, interleukin-17, and antibody responses. Arthritis Rheum. 2009, 60, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Ulmansky, R.; Landstein, D.; Moallem, E.; Loeb, V.; Levin, A.; Meyuhas, R.; Katzavian, G.; Yair, S.; Naparstek, Y. A humanized monoclonal antibody against heat shock protein 60 suppresses murine arthritis and colitis and skews the cytokine balance toward an anti-inflammatory response. J. Immunol. 2015, 194, 5103–5109. [Google Scholar] [CrossRef] [Green Version]
- Pohar, J.; Simon, Q.; Fillatreau, S. Antigen-Specificity in the Thymic Development and Peripheral Activity of CD4 FOXP3 T Regulatory Cells. Front. Immunol. 2018, 9, 1701. [Google Scholar] [CrossRef] [Green Version]
- Adamopoulou, E.; Tenzer, S.; Hillen, N.; Klug, P.; Rota, I.A.; Tietz, S.; Gebhardt, M.; Stevanovic, S.; Schild, H.; Tolosa, E.; et al. Exploring the MHC-peptide matrix of central tolerance in the human thymus. Nat. Commun. 2013, 4, 2039. [Google Scholar] [CrossRef]
- van Eden, W.; Jansen, M.A.A.; Ludwig, I.; van Kooten, P.; van der Zee, R.; Broere, F. The Enigma of Heat Shock Proteins in Immune Tolerance. Front. Immunol. 2017, 8, 1599. [Google Scholar] [CrossRef] [PubMed]
- Wendling, U.; Paul, L.; van der Zee, R.; Prakken, B.; Singh, M.; van Eden, W. A Conserved Mycobacterial Heat Shock Protein (hsp) 70 Sequence Prevents Adjuvant Arthritis upon Nasal Administration and Induces IL-10-Producing T Cells That Cross-React with the Mammalian Self-hsp70 Homologue. J. Immunol. 2000, 164, 2711–2717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Eden, W.; van Herwijnen, M.; Wagenaar, J.; van Kooten, P.; Broere, F.; van der Zee, R. Stress proteins are used by the immune system for cognate interactions with anti-inflammatory regulatory T cells. FEBS Lett. 2013, 587, 1951–1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wachstein, J.; Tischer, S.; Figueiredo, C.; Limbourg, A.; Falk, C.; Immenschuh, S.; Blasczyk, R.; Eiz-Vesper, B. HSP70 Enhances Immunosuppressive Function of CD4+ CD25+ FoxP3+ T Regulatory Cells and Cytotoxicity in CD4+ CD25− T Cells. PLoS ONE 2012, 7, e51747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Eden, W.; Jansen, M.A.A.; Wolf, A.C.M.D.; Ludwig, I.S.; Leufkens, P.; Broere, F. The Immunomodulatory Potential of tolDCs Loaded with Heat Shock Proteins. Front. Immunol. 2017, 8, 1690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stocki, P.; Wang, X.N.; Dickinson, A.M. Inducible Heat Shock Protein 70 Reduces T Cell Responses and Stimulatory Capacity of Monocyte-derived Dendritic Cells. J. Biol. Chem. 2012, 287, 12387–12394. [Google Scholar] [CrossRef] [Green Version]
- Spiering, R.; Jansen, M.A.A.; Wood, M.J.; Fath, A.A.; Eltherington, O.; Anderson, A.E.; Pratt, A.G.; van Eden, W.; Isaacs, J.D.; Broere, F.; et al. Targeting of tolerogenic dendritic cells to heat-shock proteins in inflammatory arthritis. J. Transl. Med. 2019, 17, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Ferretti, C.; Shi, F.D.; Cohen, I.R.; Quintana, F.J.; La Cava, A. DNA Vaccination With Hsp70 Protects Against Systemic Lupus Erythematosus in (NZB × NZW)F1 Mice. Arthritis Rheumatol. 2020, 72, 997–1002. [Google Scholar] [CrossRef]
- Quintana, F.J.; Carmi, P.; Mor, F.; Cohen, I.R. Inhibition of adjuvant-induced arthritis by DNA vaccination with the 70-kd or the 90-kd human heat-shock protein: Immune cross-regulation with the 60-kd heat-shock protein. Arthritis Rheum. 2004, 50, 712–720. [Google Scholar] [CrossRef]
- Prakken, B.J.; Samodal, R.; Le, T.D.; Giannoni, F.; Yung, G.P.; Scavulli, J.; Amox, D.; Roord, S.; Kleer, I.D.; Bonnin, D.; et al. Epitope-specific immunotherapy induces immune deviation of proinflammatory T cells in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2004, 101, 4228–4233. [Google Scholar] [CrossRef] [Green Version]
- Koffeman, E.C.; Genovese, M.; Amox, D.; Keogh, E.; Santana, E.; Matteson, E.L.; Kavanaugh, A.; Molitor, J.A.; Schiff, M.H.; Posever, J.O.; et al. Epitope-specific immunotherapy of rheumatoid arthritis: Clinical responsiveness occurs with immune deviation and relies on the expression of a cluster of molecules associated with T cell tolerance in a double-blind, placebo-controlled, pilot phase II trial. Arthritis Rheum. 2009, 60, 3207–3216. [Google Scholar] [CrossRef] [PubMed]
- Corrales, O.; Hernández, L.; Prada, D.; Gómez, J.; Reyes, Y.; López, A.M.; González, L.J.; Horta, M.D.C.D. CIGB-814, an altered peptide ligand derived from human heat-shock protein 60, decreases anti-cyclic citrullinated peptides antibodies in patients with rheumatoid arthritis. Clin. Rheumatol. 2018, 38, 955–960. [Google Scholar] [CrossRef]
- Lazar, L.; Ofan, R.; Weintrob, N.; Avron, A.; Tamir, M.; Elias, D.; Phillip, M.; Josefsberg, Z. Heat-shock protein peptide DiaPep277 treatment in children with newly diagnosed type 1 diabetes: A randomised, double-blind phase II study. Diabetes Metab. Res. Rev. 2007, 23, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Huurman, V.A.L.; Meide, P.E.V.D.; Duinkerken, G.; Willemen, S.; Cohen, I.R.; Elias, D.; Roep, B.O. Immunological efficacy of heat shock protein 60 peptide DiaPep277TM therapy in clinical type I diabetes. Clin. Exp. Immunol. 2008, 152, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Kirkham, B.; Chaabo, K.; Hall, C.; Garrood, T.; Mant, T.; Allen, E.; Vincent, A.; Vasconcelos, J.C.; Prevost, A.T.; Panayi, G.S.; et al. Safety and patient response as indicated by biomarker changes to binding immunoglobulin protein in the phase I/IIA RAGULA clinical trial in rheumatoid arthritis. Rheumatology (Oxford) 2016, 55, 1993–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tukaj, S. Heat Shock Protein 70 as a Double Agent Acting Inside and Outside the Cell: Insights into Autoimmunity. Int. J. Mol. Sci. 2020, 21, 5298. https://doi.org/10.3390/ijms21155298
Tukaj S. Heat Shock Protein 70 as a Double Agent Acting Inside and Outside the Cell: Insights into Autoimmunity. International Journal of Molecular Sciences. 2020; 21(15):5298. https://doi.org/10.3390/ijms21155298
Chicago/Turabian StyleTukaj, Stefan. 2020. "Heat Shock Protein 70 as a Double Agent Acting Inside and Outside the Cell: Insights into Autoimmunity" International Journal of Molecular Sciences 21, no. 15: 5298. https://doi.org/10.3390/ijms21155298
APA StyleTukaj, S. (2020). Heat Shock Protein 70 as a Double Agent Acting Inside and Outside the Cell: Insights into Autoimmunity. International Journal of Molecular Sciences, 21(15), 5298. https://doi.org/10.3390/ijms21155298