Indoleamine 2,3-Dioxygenase 2 Deficiency Exacerbates Imiquimod-Induced Psoriasis-Like Skin Inflammation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
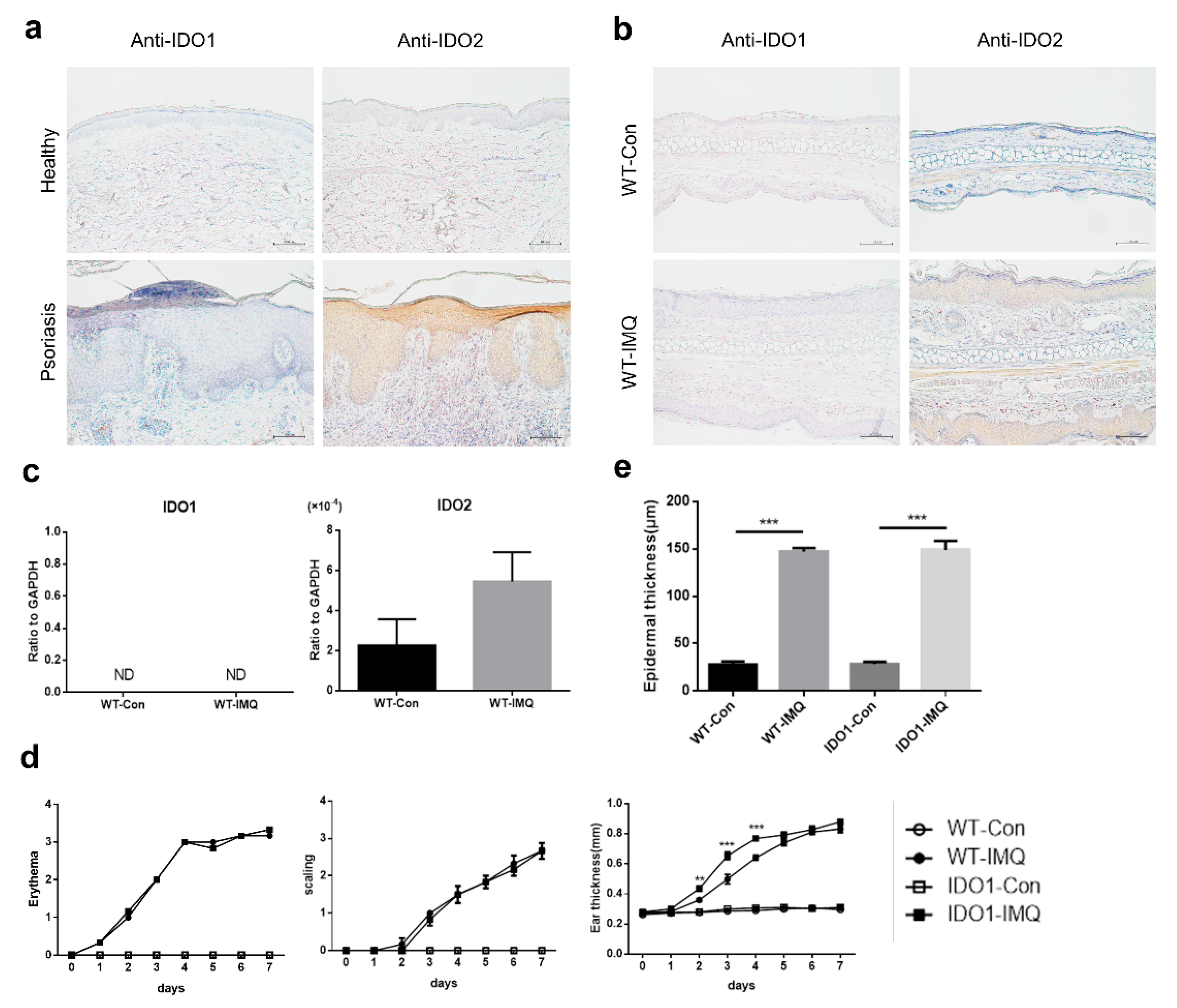
2.1. Psoriasiform Lesions in the Imiquimod-Induced Mouse Model of Psoriasis Are Associated with IDO2 But Not with IDO1
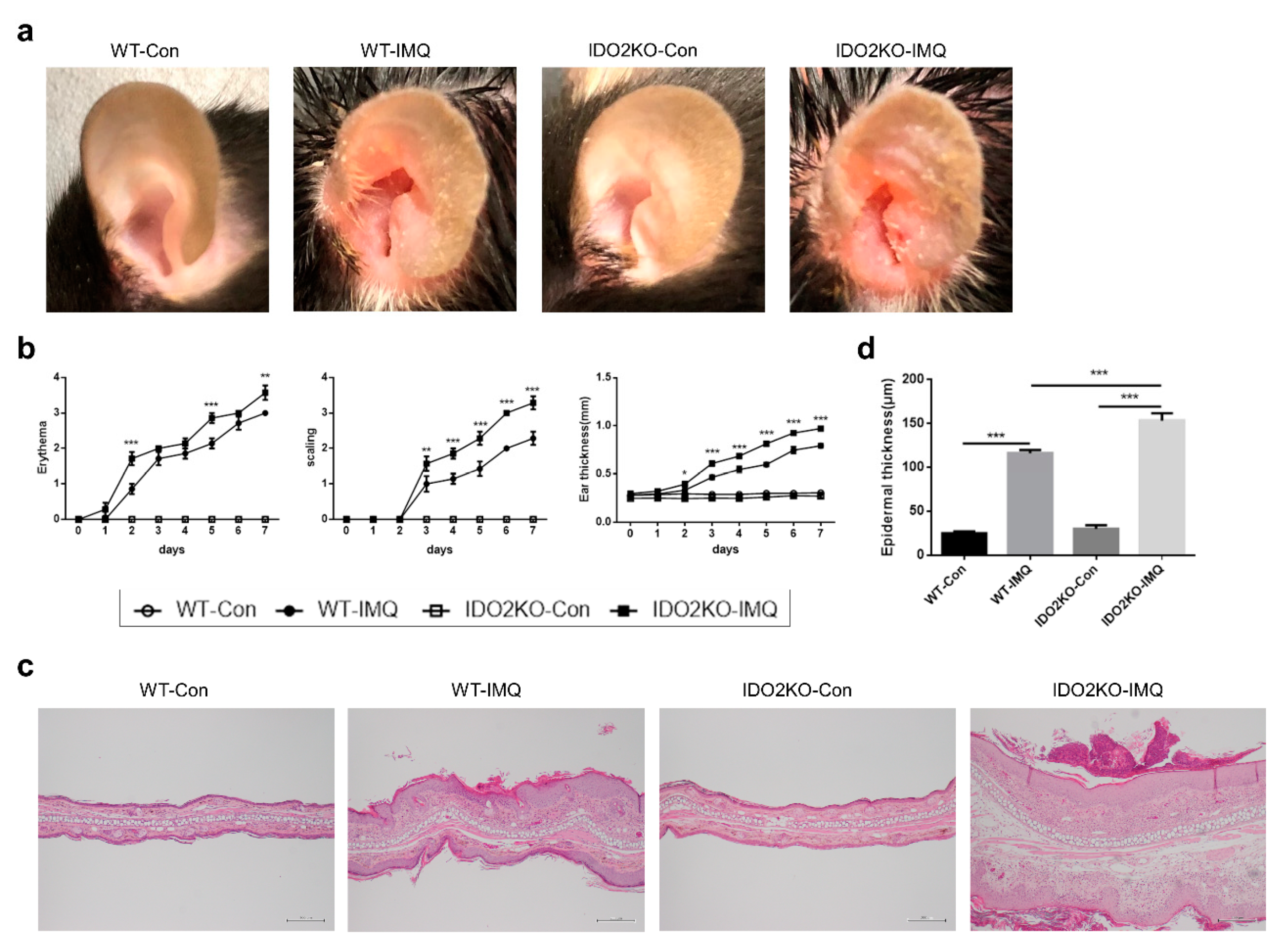
2.2. IMQ-Induced Psoriasis-Like Inflammation Is Significantly Increased in IDO2 KO Mice
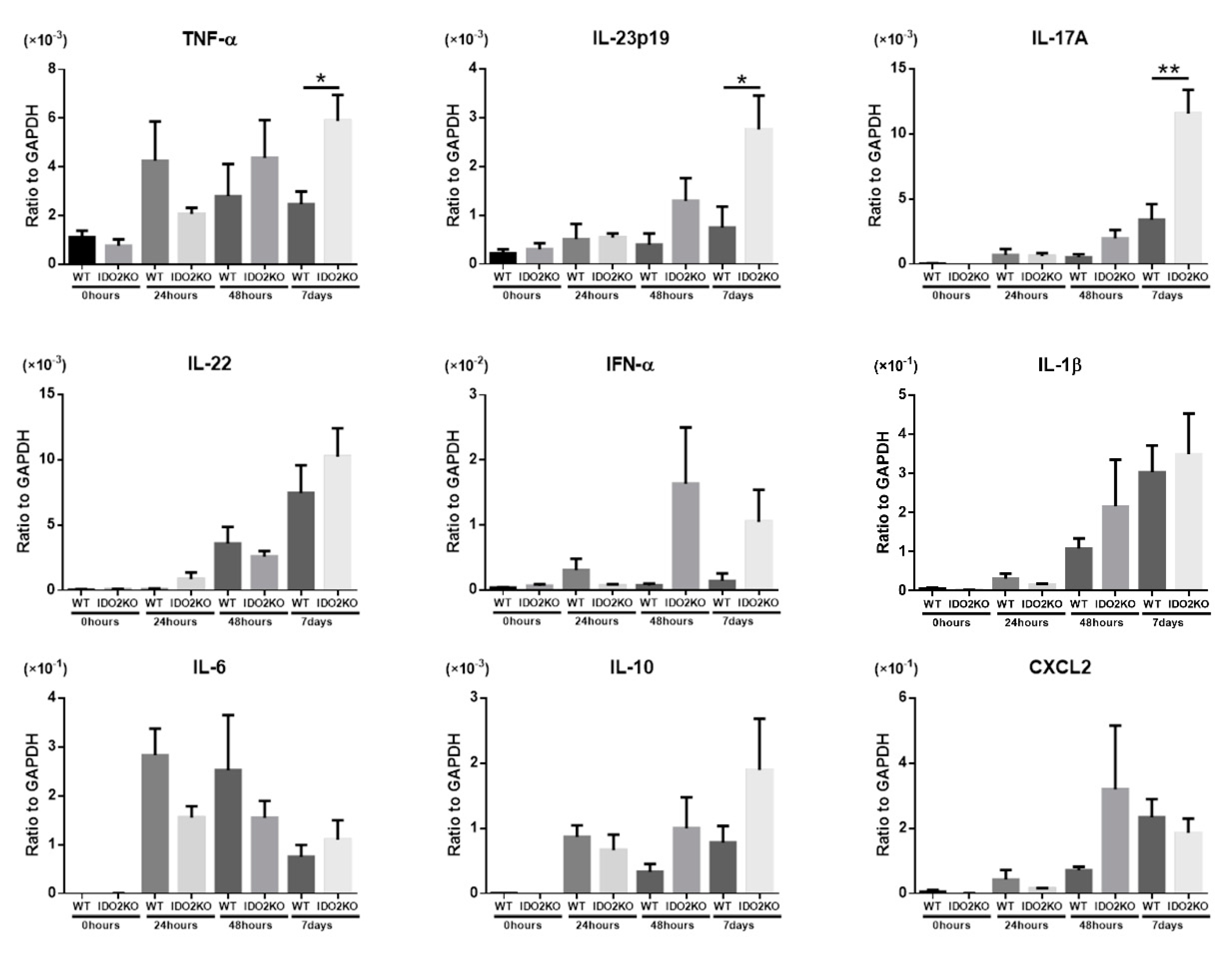
2.3. The Deletion of IDO2 Affects the mRNA Expression of Key Cytokines Involved in the Pathogenesis of Psoriasis
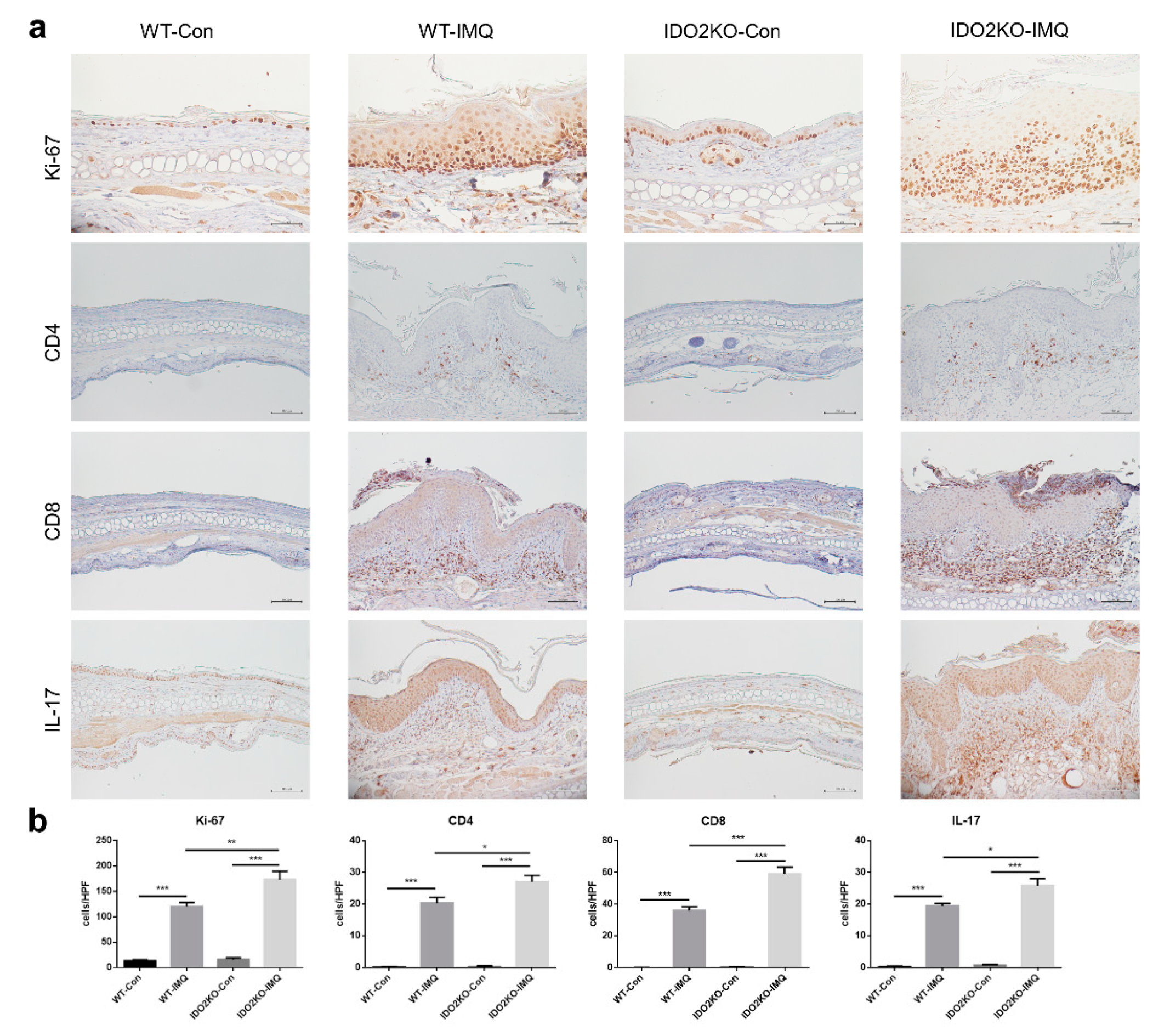
2.4. Keratinocyte Proliferation and Inflammatory Cell Infiltration Are Significantly Induced in IMQ-Treated IDO2 KO Mice
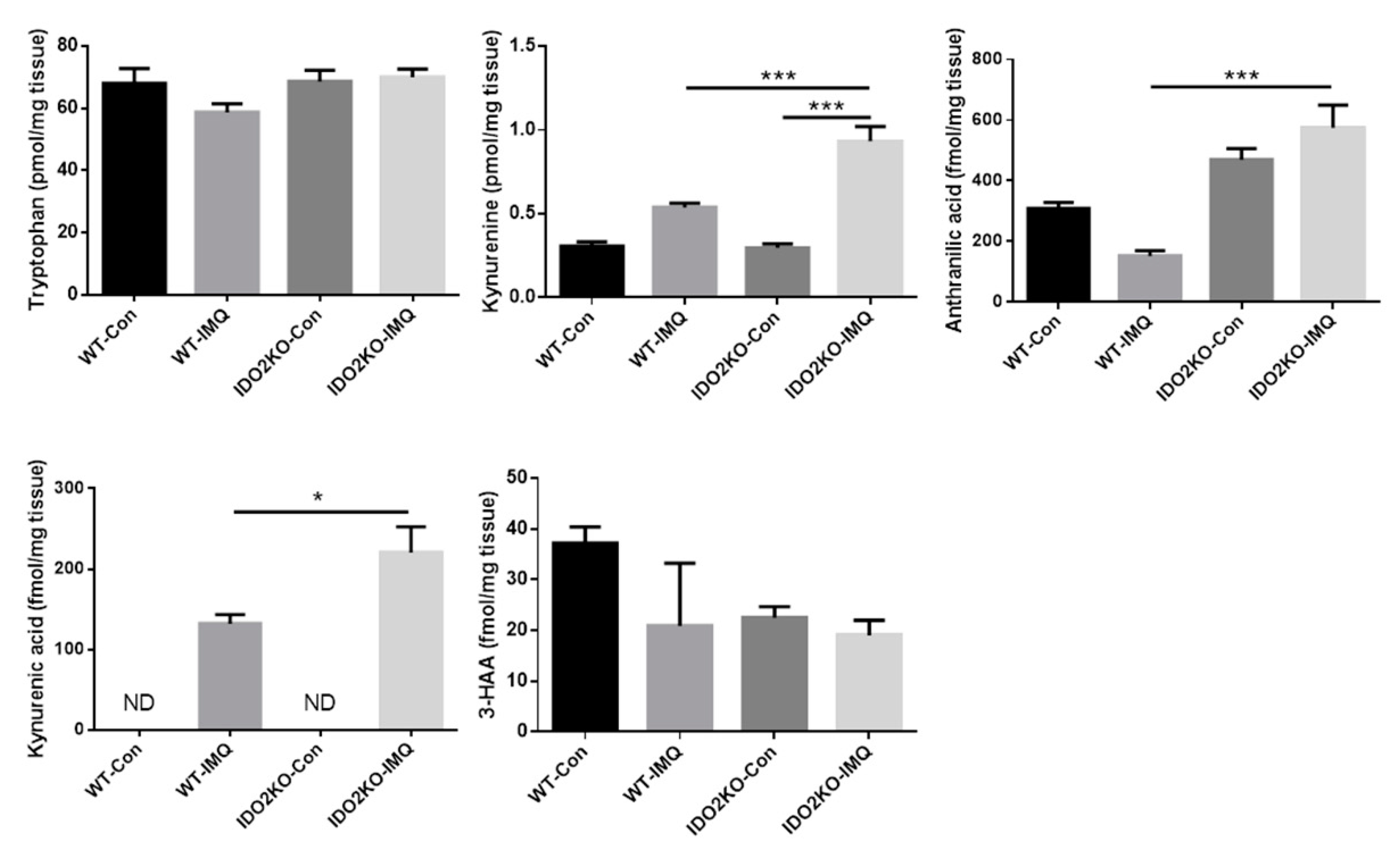
2.5. Exacerbation of IMQ-Induced Psoriasis-Like Skin Inflammation in IDO2 KO Mice Is Associated with IDO2 Itself rather than Tryptophan Metabolites
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. IMQ-Induced Psoriasis-Like Inflammation Models
4.3. Patients
4.4. RNA Extraction and Quantitative Real-Time PCR
4.5. Histology and Immunohistochemistry
4.6. Measurement of Tryptophan Metabolites
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Imiquimod | IMQ |
| Indoleamine 2,3-dioxygenase 1 | IDO1 |
| Indoleamine 2,3-dioxygenase 2 | IDO2 |
| 3-hydroxy anthranilic acid | 3-HAA |
| High performance liquid chromatography | HPLC |
| Psoriasis area and severity index | PASI |
References
- Wagner, E.F.; Schonthaler, H.B.; Guinea-Viniegra, J.; Tschachler, E. Psoriasis: What we have learned from mouse models. Nat. Rev. Rheumatol. 2010, 6, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Nestle, F.O.; Kaplan, D.H.; Barker, J. Psoriasis. N. Engl. J. Med. 2009, 361, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Kurschus, F.C.; Moos, S. IL-17 for therapy. J. Derm. Sci. 2017, 87, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Greaves, M.W.; Weinstein, G.D. Treatment of Psoriasis. N. Engl. J. Med. 1995, 332, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, C.E.; Barker, J.N. Pathogenesis and clinical features of psoriasis. Lancet (Lond. Engl.) 2007, 370, 263–271. [Google Scholar] [CrossRef]
- Van der Fits, L.; Mourits, S.; Voerman, J.S.; Kant, M.; Boon, L.; Laman, J.D.; Cornelissen, F.; Mus, A.M.; Florencia, E.; Prens, E.P.; et al. Imiquimod-induced psoriasis-like skin inflammation in mice is mediated via the IL-23/IL-17 axis. J. Immunol. 2009, 182, 5836–5845. [Google Scholar] [CrossRef]
- Gilliet, M.; Conrad, C.; Geiges, M.; Cozzio, A.; Thürlimann, W.; Burg, G.; Nestle, F.O.; Dummer, R. Psoriasis triggered by toll-like receptor 7 agonist imiquimod in the presence of dermal plasmacytoid dendritic cell precursors. Arch. Derm. 2004, 140, 1490–1495. [Google Scholar] [CrossRef] [Green Version]
- Geisse, J.; Caro, I.; Lindholm, J.; Golitz, L.; Stampone, P.; Owens, M. Imiquimod 5% cream for the treatment of superficial basal cell carcinoma: Results from two phase III, randomized, vehicle-controlled studies. J. Am. Acad. Derm. 2004, 50, 722–733. [Google Scholar] [CrossRef]
- Lebwohl, M.; Dinehart, S.; Whiting, D.; Lee, P.K.; Tawfik, N.; Jorizzo, J.; Lee, J.H.; Fox, T.L. Imiquimod 5% cream for the treatment of actinic keratosis: Results from two phase III, randomized, double-blind, parallel group, vehicle-controlled trials. J. Am. Acad. Derm. 2004, 50, 714–721. [Google Scholar] [CrossRef]
- Wu, J.K.; Siller, G.; Strutton, G. Psoriasis induced by topical imiquimod. Australas. J. Derm. 2004, 45, 47–50. [Google Scholar] [CrossRef]
- Fanti, P.A.; Dika, E.; Vaccari, S.; Miscial, C.; Varotti, C. Generalized psoriasis induced by topical treatment of actinic keratosis with imiquimod. Int. J. Derm. 2006, 45, 1464–1465. [Google Scholar] [CrossRef] [PubMed]
- Takikawa, O.; Yoshida, R.; Kido, R.; Hayaishi, O. Tryptophan degradation in mice initiated by indoleamine 2,3-dioxygenase. J. Biol. Chem. 1986, 261, 3648–3653. [Google Scholar] [PubMed]
- Mellor, A.L.; Baban, B.; Chandler, P.; Marshall, B.; Jhaver, K.; Hansen, A.; Koni, P.A.; Iwashima, M.; Munn, D.H. Cutting edge: Induced indoleamine 2,3 dioxygenase expression in dendritic cell subsets suppresses T cell clonal expansion. J. Immunol. 2003, 171, 1652–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellor, A.L.; Sivakumar, J.; Chandler, P.; Smith, K.; Molina, H.; Mao, D.; Munn, D.H. Prevention of T cell-driven complement activation and inflammation by tryptophan catabolism during pregnancy. Nat. Immunol. 2001, 2, 64–68. [Google Scholar] [CrossRef]
- Fallarino, F.; Grohmann, U.; Vacca, C.; Orabona, C.; Spreca, A.; Fioretti, M.C.; Puccetti, P. T Cell Apoptosis by Kynurenines. In Developments in Tryptophan and Serotonin Metabolism; Allegri, G., Costa, C.V.L., Ragazzi, E., Steinhart, H., Laresio, L., Eds.; Springer US: Boston, MA, USA, 2003; pp. 183–190. [Google Scholar]
- Kahler, D.J.; Mellor, A.L. T cell regulatory plasmacytoid dendritic cells expressing indoleamine 2,3 dioxygenase. Handb. Exp. Pharm. 2009, 188, 165–196. [Google Scholar]
- Llamas-Velasco, M.; Bonay, P.; Jose Concha-Garzon, M.; Corvo-Villen, L.; Vara, A.; Cibrian, D.; Sanguino-Pascual, A.; Sanchez-Madrid, F.; de la Fuente, H.; Dauden, E. Immune cells from patients with psoriasis are defective in inducing indoleamine 2,3-dioxygenase expression in response to inflammatory stimuli. Br. J. Derm. 2017, 176, 695–704. [Google Scholar] [CrossRef]
- Trabanelli, S. Indoleamine 2,3-dioxygenase in psoriasis: A defective mechanism. Br. J. Derm. 2017, 176, 570–572. [Google Scholar] [CrossRef]
- Ball, H.J.; Yuasa, H.J.; Austin, C.J.; Weiser, S.; Hunt, N.H. Indoleamine 2,3-dioxygenase-2; a new enzyme in the kynurenine pathway. Int. J. Biochem. Cell Biol. 2009, 41, 467–471. [Google Scholar] [CrossRef]
- Fatokun, A.A.; Hunt, N.H.; Ball, H.J. Indoleamine 2,3-dioxygenase 2 (IDO2) and the kynurenine pathway: Characteristics and potential roles in health and disease. Amino Acids. 2013, 45, 1319–1329. [Google Scholar] [CrossRef]
- Yuasa, H.J.; Takubo, M.; Takahashi, A.; Hasegawa, T.; Noma, H.; Suzuki, T. Evolution of Vertebrate Indoleamine 2,3-Dioxygenases. J. Mol. Evol. 2007, 65, 705. [Google Scholar] [CrossRef]
- Ball, H.J.; Sanchez-Perez, A.; Weiser, S.; Austin, C.J.; Astelbauer, F.; Miu, J.; McQuillan, J.A.; Stocker, R.; Jermiin, L.S.; Hunt, N.H. Characterization of an indoleamine 2,3-dioxygenase-like protein found in humans and mice. Gene 2007, 396, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Yuasa, H.J.; Hasegawa, T.; Nakamura, T.; Suzuki, T. Bacterial expression and characterization of molluscan IDO-like myoglobin. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 146, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, M.; Yamamoto, Y.; Kawasoe, M.; Arioka, Y.; Murakami, Y.; Hoshi, M.; Saito, K. Studies on tissue and cellular distribution of indoleamine 2,3-dioxygenase 2: The absence of IDO1 upregulates IDO2 expression in the epididymis. J. Histochem. Cytochem. 2012, 60, 854–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlo, L.M.F.; Pigott, E.; DuHadaway, J.B.; Grabler, S.; Metz, R.; Prendergast, G.C.; Mandik-Nayak, L. IDO2 is a critical mediator of autoantibody production and inflammatory pathogenesis in a mouse model of autoimmune arthritis. J. Immunol. 2014, 192, 2082–2090. [Google Scholar] [CrossRef] [Green Version]
- Merlo, L.M.F.; Grabler, S.; DuHadaway, J.B.; Pigott, E.; Manley, K.; Prendergast, G.C.; Laury-Kleintop, L.D.; Mandik-Nayak, L. Therapeutic antibody targeting of indoleamine-2,3-dioxygenase (IDO2) inhibits autoimmune arthritis. Clin. Immunol. 2017, 179, 8–16. [Google Scholar] [CrossRef]
- Metz, R.; Smith, C.; DuHadaway, J.B.; Chandler, P.; Baban, B.; Merlo, L.M.F.; Pigott, E.; Keough, M.P.; Rust, S.; Mellor, A.L.; et al. IDO2 is critical for IDO1-mediated T-cell regulation and exerts a non-redundant function in inflammation. Int. Immunol. 2014, 26, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Merlo, L.M.; DuHadaway, J.B.; Grabler, S.; Prendergast, G.C.; Muller, A.J.; Mandik-Nayak, L. IDO2 Modulates T Cell-Dependent Autoimmune Responses through a B Cell-Intrinsic Mechanism. J. Immunol. 2016, 196, 4487–4497. [Google Scholar] [CrossRef] [Green Version]
- Asadullah, K.; Sterry, W.; Stephanek, K.; Jasulaitis, D.; Leupold, M.; Audring, H.; Volk, H.D.; Döcke, W.D. IL-10 is a key cytokine in psoriasis. Proof of principle by IL-10 therapy: A new therapeutic approach. J. Clin. Invest. 1998, 101, 783–794. [Google Scholar] [CrossRef]
- Hawkes, J.E.; Yan, B.Y.; Chan, T.C.; Krueger, J.G. Discovery of the IL-23/IL-17 Signaling Pathway and the Treatment of Psoriasis. J. Immunol. 2018, 201, 1605–1613. [Google Scholar] [CrossRef]
- El Malki, K.; Karbach, S.H.; Huppert, J.; Zayoud, M.; Reissig, S.; Schuler, R.; Nikolaev, A.; Karram, K.; Munzel, T.; Kuhlmann, C.R.; et al. An alternative pathway of imiquimod-induced psoriasis-like skin inflammation in the absence of interleukin-17 receptor a signaling. J. Invest. Derm. 2013, 133, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Moos, S.; Mohebiany, A.N.; Waisman, A.; Kurschus, F.C. Imiquimod-Induced Psoriasis in Mice Depends on the IL-17 Signaling of Keratinocytes. J. Invest. Derm. 2019, 139, 1110–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezrich, J.D.; Fechner, J.H.; Zhang, X.; Johnson, B.P.; Burlingham, W.J.; Bradfield, C.A. An interaction between kynurenine and the aryl hydrocarbon receptor can generate regulatory T cells. J. Immunol. 2010, 185, 3190–3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julliard, W.; Fechner, J.H.; Mezrich, J.D. The aryl hydrocarbon receptor meets immunology: Friend or foe? A little of both. Front. Immunol. 2014, 5, 458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salimi Elizei, S.; Poormasjedi-Meibod, M.S.; Wang, X.; Kheirandish, M.; Ghahary, A. Kynurenic acid downregulates IL-17/1L-23 axis in vitro. Mol. Cell. Biochem. 2017, 431, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Flutter, B.; Nestle, F.O. TLRs to cytokines: Mechanistic insights from the imiquimod mouse model of psoriasis. Eur. J. Immunol. 2013, 43, 3138–3146. [Google Scholar] [CrossRef]
- Asp, L.; Johansson, A.S.; Mann, A.; Owe-Larsson, B.; Urbanska, E.M.; Kocki, T.; Kegel, M.; Engberg, G.; Lundkvist, G.B.; Karlsson, H. Effects of pro-inflammatory cytokines on expression of kynurenine pathway enzymes in human dermal fibroblasts. J. Inflamm. 2011, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Sheipouri, D.; Grant, R.; Bustamante, S.; Lovejoy, D.; Guillemin, G.; Braidy, N. Characterisation of the Kynurenine Pathway in Skin-Derived Fibroblasts and Keratinocytes. J. Cell. Biochem. 2015, 116, 903–922. [Google Scholar] [CrossRef]
- Scheler, M.; Wenzel, J.; Tüting, T.; Takikawa, O.; Bieber, T.; von Bubnoff, D. Indoleamine 2,3-Dioxygenase (IDO): The Antagonist of Type I Interferon-Driven Skin Inflammation? Am. J. Pathol. 2007, 171, 1936–1943. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Yamasuge, W.; Imai, S.; Kunisawa, K.; Hoshi, M.; Fujigaki, H.; Mouri, A.; Nabeshima, T.; Saito, K. Lipopolysaccharide shock reveals the immune function of indoleamine 2,3-dioxygenase 2 through the regulation of IL-6/stat3 signalling. Sci. Rep. 2018, 8, 15917. [Google Scholar] [CrossRef] [Green Version]
- Sano, S.; Chan, K.S.; Carbajal, S.; Clifford, J.; Peavey, M.; Kiguchi, K.; Itami, S.; Nickoloff, B.J.; DiGiovanni, J. Stat3 links activated keratinocytes and immunocytes required for development of psoriasis in a novel transgenic mouse model. Nat. Med. 2005, 11, 43–49. [Google Scholar] [CrossRef]
- Harris, T.J.; Grosso, J.F.; Yen, H.R.; Xin, H.; Kortylewski, M.; Albesiano, E.; Hipkiss, E.L.; Getnet, D.; Goldberg, M.V.; Maris, C.H.; et al. Cutting edge: An in vivo requirement for STAT3 signaling in TH17 development and TH17-dependent autoimmunity. J. Immunol. 2007, 179, 4313–4317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuasa, H.J.; Ball, H.J.; Ho, Y.F.; Austin, C.J.D.; Whittington, C.M.; Belov, K.; Maghzal, G.J.; Jermiin, L.S.; Hunt, N.H. Characterization and evolution of vertebrate indoleamine 2, 3-dioxygenases: IDOs from monotremes and marsupials. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol. 2009, 153, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Yuasa, H.J.; Ball, H.J.; Austin, C.J.D.; Hunt, N.H. 1-l-methyltryptophan is a more effective inhibitor of vertebrate IDO2 enzymes than 1-d-methyltryptophan. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol. 2010, 157, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Pantouris, G.; Serys, M.; Yuasa, H.J.; Ball, H.J.; Mowat, C.G. Human indoleamine 2,3-dioxygenase-2 has substrate specificity and inhibition characteristics distinct from those of indoleamine 2,3-dioxygenase-1. Amino Acids. 2014, 46, 2155–2163. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujii, K.; Yamamoto, Y.; Mizutani, Y.; Saito, K.; Seishima, M. Indoleamine 2,3-Dioxygenase 2 Deficiency Exacerbates Imiquimod-Induced Psoriasis-Like Skin Inflammation. Int. J. Mol. Sci. 2020, 21, 5515. https://doi.org/10.3390/ijms21155515
Fujii K, Yamamoto Y, Mizutani Y, Saito K, Seishima M. Indoleamine 2,3-Dioxygenase 2 Deficiency Exacerbates Imiquimod-Induced Psoriasis-Like Skin Inflammation. International Journal of Molecular Sciences. 2020; 21(15):5515. https://doi.org/10.3390/ijms21155515
Chicago/Turabian StyleFujii, Kento, Yasuko Yamamoto, Yoko Mizutani, Kuniaki Saito, and Mariko Seishima. 2020. "Indoleamine 2,3-Dioxygenase 2 Deficiency Exacerbates Imiquimod-Induced Psoriasis-Like Skin Inflammation" International Journal of Molecular Sciences 21, no. 15: 5515. https://doi.org/10.3390/ijms21155515
APA StyleFujii, K., Yamamoto, Y., Mizutani, Y., Saito, K., & Seishima, M. (2020). Indoleamine 2,3-Dioxygenase 2 Deficiency Exacerbates Imiquimod-Induced Psoriasis-Like Skin Inflammation. International Journal of Molecular Sciences, 21(15), 5515. https://doi.org/10.3390/ijms21155515