Anti-Epileptic Effects of FABP3 Ligand MF1 through the Benzodiazepine Recognition Site of the GABAA Receptor


 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. MF1 Promotes GABAA Receptor Currents through the Benzodiazepine Recognition Site
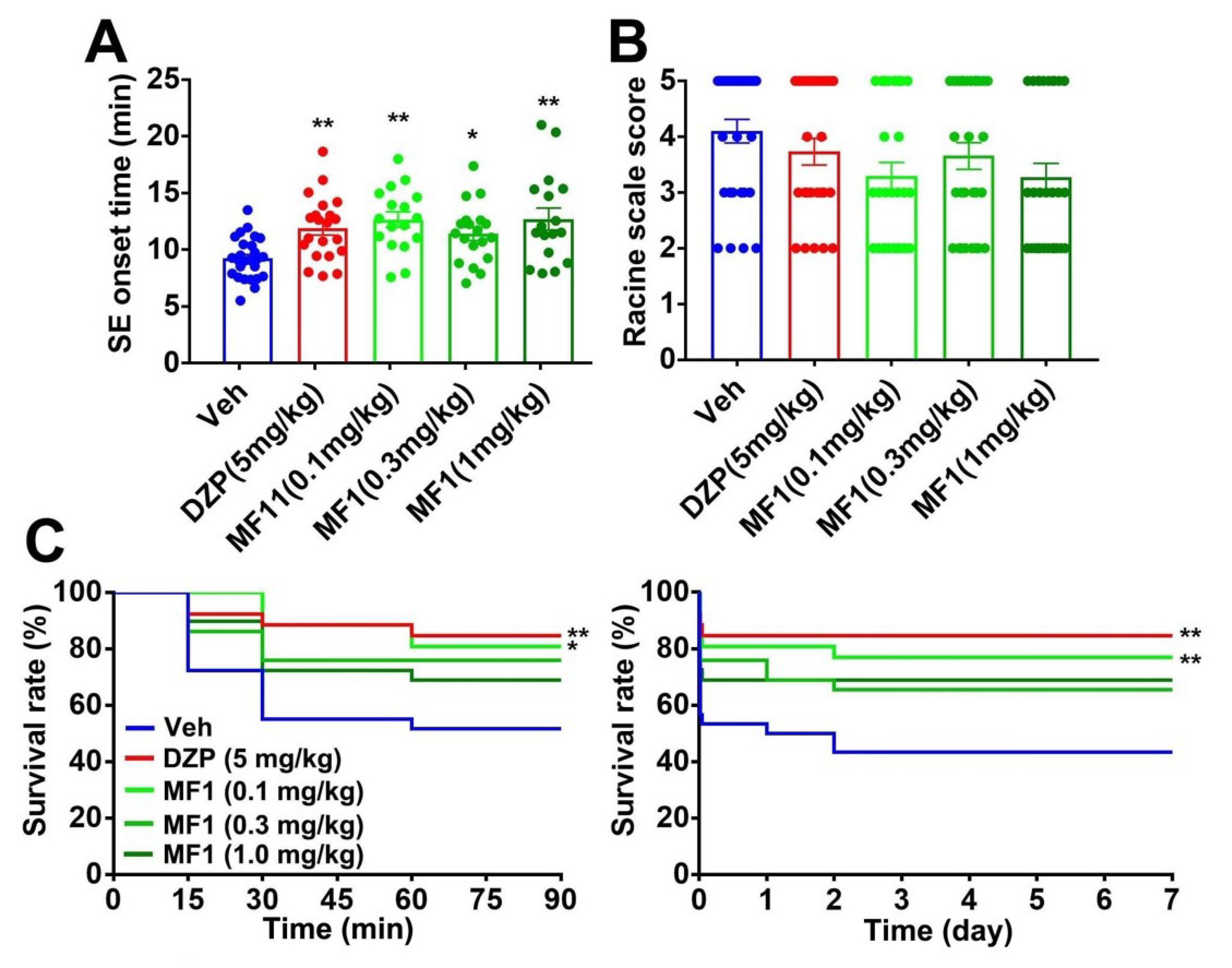
2.2. Acute MF1 Administration Attenuates SE and Mortality in PILO-Treated Mice
2.3. Acute MF1 Administration Does Not Improve Epileptic Seizures in PTZ-Treated Mice
2.4. Chronic MF1 Administration Suppresses SE and Mortality in PILO-Treated Mice
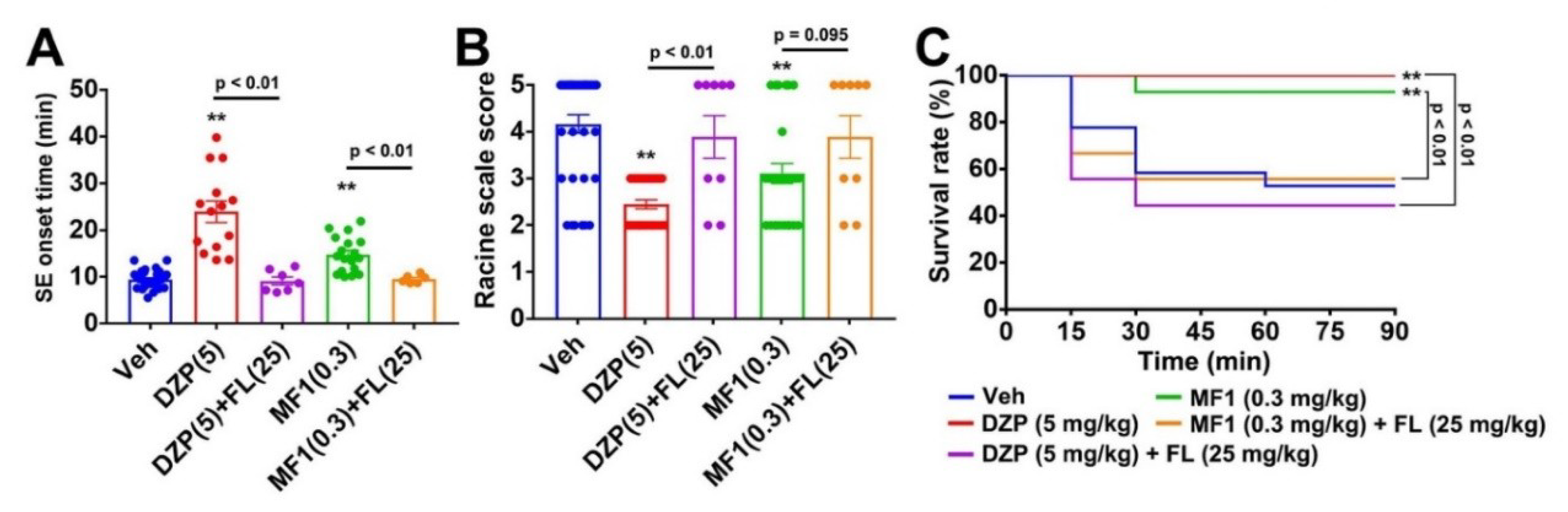
2.5. Flumazenil Blocks Anti-Epileptic Effect of MF1 in PILO-Treated Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemicals
4.3. Cell Transfection
4.4. Whole Cell Patch-Clamp Recording
4.5. Evaluation of Epileptic Behaviors
4.5.1. PILO-Induced SE Model
4.5.2. PTZ-Induced Seizure
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| ANOVA | analysis of variance |
| CMC | carboxymethylcellulose |
| DMSO | dimethyl sulfoxide |
| DZP | diazepam |
| FABP | fatty acid binding protein |
| GABA | γ-aminobutyrate |
| GABAA | GABA type-A |
| GAD | glutamic acid decarboxylase |
| GFP | green fluorescent protein |
| MF1 | 4-(2-(1-(2-chlorophenyl)-5-phenyl-1H-pyrazol-3-yl)phenoxy) butanoic acid |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| PD | Parkinson’s disease |
| PFF | preformed fibril |
| PILO | pilocarpine |
| PTZ | pentylenetetrazol |
| SE | status epilepticus |
| SEM | standard error of the mean |
References
- Whiting, P.J.; Bonnert, T.P.; McKernan, R.M.; Farrar, S.; Le Bourdellès, B.; Heavens, R.P.; Smith, D.W.; Hewson, L.; Rigby, M.R.; Sirinathsinghji, D.J.; et al. Molecular and functional diversity of the expanding GABA-A receptor gene family. Ann. N. Y. Acad. Sci. 1999, 868, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.J.; Silver, R.A. Shunting inhibition modulates neuronal gain during synaptic excitation. Neuron 2003, 38, 433–445. [Google Scholar] [CrossRef] [Green Version]
- Crunelli, V.; Lőrincz, M.L.; McCafferty, C.; Lambert, R.C.; Leresche, N.; Di Giovanni, G.; David, F. Clinical and experimental insight into pathophysiology, comorbidity and therapy of absence seizures. Brain 2020, 21, awaa072. [Google Scholar] [CrossRef] [PubMed]
- Rogawski, M.A.; Löscher, W.; Rho, J.M. Mechanisms of Action of Antiseizure Drugs and the Ketogenic Diet. Cold Spring Harb. Perspect. Med. 2016, 6, a022780. [Google Scholar] [CrossRef] [PubMed]
- Błaszczyk, J.W. Parkinson’s Disease and Neurodegeneration: GABA-Collapse Hypothesis. Front. Neurosci. 2016, 10, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, D.; Wen, L.; Wu, Z.; Shen, Y. GABAergic dysfunction in excitatory and inhibitory (E/I) imbalance drives the pathogenesis of Alzheimer’s disease. Alzheimers Dement. 2020, 1–18. [Google Scholar] [CrossRef]
- Fung, L.K.; Flores, R.E.; Gu, M.; Sun, K.L.; James, D.; Schuck, R.K.; Jo, B.; Park, J.H.; Lee, B.C.; Jung, J.H.; et al. Thalamic and prefrontal GABA concentrations but not GABA(A) receptor densities are altered in high-functioning adults with autism spectrum disorder. Mol. Psychiatry 2020. [Google Scholar] [CrossRef]
- Faulkner, M.A.; Singh, S.P. Neurogenetic disorders and treatment of associated seizures. Pharmacotherapy 2013, 33, 330–343. [Google Scholar] [CrossRef]
- Giorgi, F.S.; Saccaro, L.F.; Busceti, C.L.; Biagioni, F.; Fornai, F. Epilepsy and Alzheimer’s Disease: Potential mechanisms for an association. Brain Res. Bull. 2020, 160, 107–120. [Google Scholar] [CrossRef]
- Löscher, W. Animal Models of Seizures and Epilepsy: Past, Present, and Future Role for the Discovery of Antiseizure Drugs. Neurochem. Res. 2017, 42, 1873–1888. [Google Scholar] [CrossRef]
- Czapiński, P.; Blaszczyk, B.; Czuczwar, S.J. Mechanisms of action of antiepileptic drugs. Curr. Top. Med. Chem. 2005, 5, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Bormann, J. The ‘ABC’ of GABA Receptors. Trends Pharmacol. Sci. 2000, 21, 16–19. [Google Scholar] [CrossRef]
- Rudolph, U.; Möhler, H. GABAA receptor subtypes: Therapeutic potential in Down syndrome, affective disorders, schizophrenia, and autism. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 483–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sassoè-Pognetto, M.; Fritschy, J.M. Mini-review: Gephyrin, a Major Postsynaptic Protein of GABAergic Synapses. Eur. J. Neurosci. 2000, 12, 2205–2210. [Google Scholar] [CrossRef] [PubMed]
- Glykys, J.; Mody, I. Activation of GABAA Receptors: Views from Outside the Synaptic Cleft. Neuron 2007, 56, 763–770. [Google Scholar] [CrossRef] [Green Version]
- Coe, N.R.; Bernlohr, D.A. Physiological properties and functions of intracellular fatty acid-binding proteins. Biochim. Biophys. Acta 1998, 1391, 287–306. [Google Scholar] [CrossRef]
- Owada, Y.; Yoshimoto, T.; Kondo, H. Spatio-temporally differential expression of genes for three members of fatty acid binding proteins in developing and mature rat brains. J. Chem. Neuroanat. 1996, 12, 113–122. [Google Scholar] [CrossRef]
- Owada, Y. Fatty acid binding protein: Localization and functional significance in the brain. Tohoku J. Exp. Med. 2008, 214, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Sepe, F.N.; Chiasserini, D.; Parnetti, L. Role of FABP3 as biomarker in Alzheimer’s disease and synucleinopathies. Future Neurol. 2018, 13, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Chiasserini, D.; Biscetti, L.; Eusebi, P.; Salvadori, N.; Frattini, G.; Simoni, S.; De Roeck, N.; Tambasco, N.; Stoops, E.; Vanderstichele, H.; et al. Differential role of CSF fatty acid binding protein 3, α-synuclein, and Alzheimer’s disease core biomarkers in Lewy body disorders and Alzheimer’s dementia. Alzheimers Res. Ther. 2017, 9, 52. [Google Scholar] [CrossRef]
- Bjerke, M.; Kern, S.; Blennow, K.; Zetterberg, H.; Waern, M.; Börjesson-Hanson, A.; Östling, S.; Kern, J.; Skoog, I. Cerebrospinal Fluid Fatty Acid-Binding Protein 3 is Related to Dementia Development in a Population-Based Sample of Older Adult Women Followed for 8 Years. J. Alzheimers Dis. 2016, 49, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Shioda, N.; Yabuki, Y.; Kobayashi, Y.; Onozato, M.; Owada, Y.; Fukunaga, K. FABP3 protein promotes α-synuclein oligomerization associated with 1-methyl-1,2,3,6-tetrahydropiridine-induced neurotoxicity. J. Biol. Chem. 2014, 289, 18957–18965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yabuki, Y.; Matsuo, K.; Kawahata, I.; Fukui, N.; Mizobata, T.; Kawata, Y.; Owada, Y.; Shioda, N.; Fukunaga, K. Fatty Acid Binding Protein 3 Enhances the Spreading and Toxicity of α-Synuclein in Mouse Brain. Int. J. Mol. Sci. 2020, 21, 2230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawahata, I.; Bousset, L.; Melki, R.; Fukunaga, K. Fatty Acid-Binding Protein 3 is Critical for α-Synuclein Uptake and MPP+-Induced Mitochondrial Dysfunction in Cultured Dopaminergic Neurons. Int. J. Mol. Sci. 2019, 20, 5358. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.; Shinoda, Y.; Yamamoto, T.; Miyachi, H.; Fukunaga, K. Development of FABP3 ligands that inhibit arachidonic acid-induced α-synuclein oligomerization. Brain Res. 2019, 1707, 190–197. [Google Scholar] [CrossRef]
- Matsuo, K.; Cheng, A.; Yabuki, Y.; Takahata, I.; Miyachi, H.; Fukunaga, K. Inhibition of MPTP-induced α-synuclein oligomerization by fatty acid-binding protein 3 ligand in MPTP-treated mice. Neuropharmacology 2019, 150, 164–174. [Google Scholar] [CrossRef]
- Liu, Y.M.; Fan, H.R.; Ding, J.; Huang, C.; Deng, S.; Zhu, T.; Xu, T.L.; Ge, W.H.; Li, W.G.; Li, F. Curcumol allosterically modulates GABA(A) receptors in a manner distinct from benzodiazepines. Sci. Rep. 2017, 7, 46654. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.P.; Ferreira, P.B.; De Sousa, D.P.; Jordan, J.; Freitas, R.M. Anticonvulsant effect of phytol in a pilocarpine model in mice. Neurosci. Lett. 2012, 523, 115–118. [Google Scholar] [CrossRef] [Green Version]
- Khoshnoud, M.J.; Tanideh, N.; Namdarian, S. Anticonvulsant activity of atorvastatin against seizure induced by pentylenetetrazole and maximal electroshock in mice. Trends Pharm. Sci. 2015, 1, 44–47. [Google Scholar]
- Vonderlin, N.; Fischer, F.; Zitron, E.; Seyler, C.; Scherer, D.; Thomas, D.; Katus, H.A.; Scholz, E.P. Inhibition of cardiac Kv1.5 potassium current by the anesthetic midazolam: Mode of action. Drug Des. Dev. Ther. 2014, 8, 2263–2271. [Google Scholar] [CrossRef] [Green Version]
- So, E.C.; Wu, K.C.; Kao, F.C.; Wu, S.N. Effects of midazolam on ion currents and membrane potential in differentiated motor neuron-like NSC-34 and NG108-15 cells. Eur. J. Pharmacol. 2014, 724, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Smolders, I.; Khan, G.M.; Manil, J.; Ebinger, G.; Michotte, Y. NMDA receptor-mediated pilocarpine-induced seizures: Characterization in freely moving rats by microdialysis. Br. J. Pharmacol. 1997, 121, 1171–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millan, M.H.; Chapman, A.G.; Meldrum, B.S. Extracellular amino acid levels in hippocampus during pilocarpine-induced seizures. Epilepsy Res. 1993, 14, 139–148. [Google Scholar] [CrossRef]
- Macdonald, R.L.; Kapur, J. Acute cellular alterations in the hippocampus after status epilepticus. Epilepsia 1999, 40, S9–S20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramanjaneyulu, R.; Ticku, M.K. Interactions of pentamethylenetetrazole and tetrazole analogues with the picrotoxinin site of the benzodiazepine-GABA receptor-ionophore complex. Eur. J. Pharmacol. 1984, 98, 337–345. [Google Scholar] [CrossRef]
- Hansen, S.L.; Sperling, B.B.; Sánchez, C. Anticonvulsant and antiepileptogenic effects of GABAA receptor ligands in pentylenetetrazole-kindled mice. Pharmacology 1984, 98, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues de Oliveira, F.; Eleuterio Rodrigues, K.; Hamoy, M.; Sarquis, Í.R.; Otake Hamoy, A.; Crespo Lopez, M.E.; Maciel Ferreira, I.; Macchi, B.M.; Luiz Martins do Nascimento, J. Fatty Acid Amides Synthesized from Andiroba Oil (Carapa guianensis Aublet.) Exhibit Anticonvulsant Action with Modulation on GABA-A Receptor in Mice: A Putative Therapeutic Option. Pharmaceuticals 2020, 13, 43. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Kida, H.; Kagawa, Y.; Yasumoto, Y.; Miyazaki, H.; Islam, A.; Ogata, M.; Yanagawa, Y.; Mitsushima, D.; Fukunaga, K.; et al. FABP3 in the Anterior Cingulate Cortex Modulates the Methylation Status of the Glutamic Acid Decarboxylase67 Promoter Region. J. Neurosci. 2018, 38, 10411–10423. [Google Scholar] [CrossRef] [Green Version]
- Yabuki, Y.; Takahata, I.; Matsuo, K.; Owada, Y.; Fukunaga, K. Ramelteon Improves Post-traumatic Stress Disorder-Like Behaviors Exhibited by Fatty Acid-Binding Protein 3 Null Mice. Mol. Neurobiol. 2018, 55, 3577–3591. [Google Scholar] [CrossRef]
- Riss, J.; Cloyd, J.; Gates, J.; Collins, S. Benzodiazepines in epilepsy: Pharmacology and pharmacokinetics. Acta Neurol. Scand. 2008, 118, 69–86. [Google Scholar] [CrossRef]
- Parsonage, M.J.; Norris, J.W. Use of diazepam in the treatment of severe convulsive status epilepticus. Br. Med. J. 1967, 3, 85–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beagle, A.J.; Darwish, S.M.; Ranasinghe, K.G.; La, A.L.; Karageorgiou, E.; Vossel, K.A. Relative Incidence of Seizures and Myoclonus in Alzheimer’s Disease, Dementia with Lewy Bodies, and Frontotemporal Dementia. J. Alzheimers Dis. 2017, 60, 211–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, M.; Horiuchi, E.; Kanazawa, I. Zonisamide has beneficial effects on Parkinson’s disease patients. Neurosci. Res. 2001, 41, 397–399. [Google Scholar] [CrossRef]
- Beniyama, Y.; Matsuno, K.; Miyachi, H. Structure-guided design, synthesis and in vitro evaluation of a series of pyrazole-based fatty acid binding protein (FABP) 3 ligands. Bioorg. Med. Chem. Lett. 2013, 23, 1662–1666. [Google Scholar] [CrossRef] [PubMed]
- Migita, K.; Yamada, J.; Nikaido, Y.; Shi, X.; Kaneko, S.; Hirose, S.; Ueno, S. Properties of a novel GABAA receptor γ2 subunit mutation associated with seizures. J. Pharmacol. Sci. 2013, 121, 84–87. [Google Scholar] [CrossRef] [Green Version]
- Yabuki, Y.; Matsuo, K.; Izumi, H.; Haga, H.; Yoshida, T.; Wakamori, M.; Kakei, A.; Sakimura, K.; Fukuda, T.; Fukunaga, K. Pharmacological properties of SAK3, a novel T-type voltage-gated Ca2+ channel enhancer. Neuropharmacology 2017, 117, 1–13. [Google Scholar] [CrossRef]
- Racine, R.J. Modification of Seizure Activity by Electrical Stimulation. II. Motor Seizure. Electroencephalogr. Clin. Neurophysiol. 1972, 32, 281–294. [Google Scholar] [CrossRef]
- Lowenstein, D.H.; Alldredge, B.K. Status Epilepticus. N. Engl. J. Med. 1998, 338, 970–976. [Google Scholar] [CrossRef]
- Shibley, H.; Smith, B.N. Pilocarpine-induced status epilepticus results in mossy fiber sprouting and spontaneous seizures in C57BL/6 and CD-1 mice. Epilepsy Res. 2002, 49, 109–120. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | GTCs Onset Time (Min) | Mortality (%) | Seizure Scale |
|---|---|---|---|
| Vehicle | 1.7 ± 0.53 | 50 (5/10) | 4 ± 0 |
| DZP (5 mg/kg, i.p.) | 3.3 ± 0.70 ** | 0 (0/10) | 3.7 ± 0.15 |
| MF1 (0.1 mg/kg, p.o.) | 0.92 ± 0.15 | 40 (4/10) | 3.8 ± 0.20 |
| MF1 (0.3 mg/kg, p.o.) | 1.2 ± 0.22 | 50 (5/10) | 3.8 ± 0.20 |
| MF1 (1.0 mg/kg, p.o.) | 1.2 ± 0.28 | 70 (7/10) | 3.8 ± 0.20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yabuki, Y.; Liu, J.; Kawahata, I.; Izumi, H.; Shinoda, Y.; Koga, K.; Ueno, S.; Shioda, N.; Fukunaga, K. Anti-Epileptic Effects of FABP3 Ligand MF1 through the Benzodiazepine Recognition Site of the GABAA Receptor. Int. J. Mol. Sci. 2020, 21, 5525. https://doi.org/10.3390/ijms21155525
Yabuki Y, Liu J, Kawahata I, Izumi H, Shinoda Y, Koga K, Ueno S, Shioda N, Fukunaga K. Anti-Epileptic Effects of FABP3 Ligand MF1 through the Benzodiazepine Recognition Site of the GABAA Receptor. International Journal of Molecular Sciences. 2020; 21(15):5525. https://doi.org/10.3390/ijms21155525
Chicago/Turabian StyleYabuki, Yasushi, Jiaqi Liu, Ichiro Kawahata, Hisanao Izumi, Yasuharu Shinoda, Kohei Koga, Shinya Ueno, Norifumi Shioda, and Kohji Fukunaga. 2020. "Anti-Epileptic Effects of FABP3 Ligand MF1 through the Benzodiazepine Recognition Site of the GABAA Receptor" International Journal of Molecular Sciences 21, no. 15: 5525. https://doi.org/10.3390/ijms21155525
APA StyleYabuki, Y., Liu, J., Kawahata, I., Izumi, H., Shinoda, Y., Koga, K., Ueno, S., Shioda, N., & Fukunaga, K. (2020). Anti-Epileptic Effects of FABP3 Ligand MF1 through the Benzodiazepine Recognition Site of the GABAA Receptor. International Journal of Molecular Sciences, 21(15), 5525. https://doi.org/10.3390/ijms21155525