Lipoteichoic Acid Accelerates Bone Healing by Enhancing Osteoblast Differentiation and Inhibiting Osteoclast Activation in a Mouse Model of Femoral Defects

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
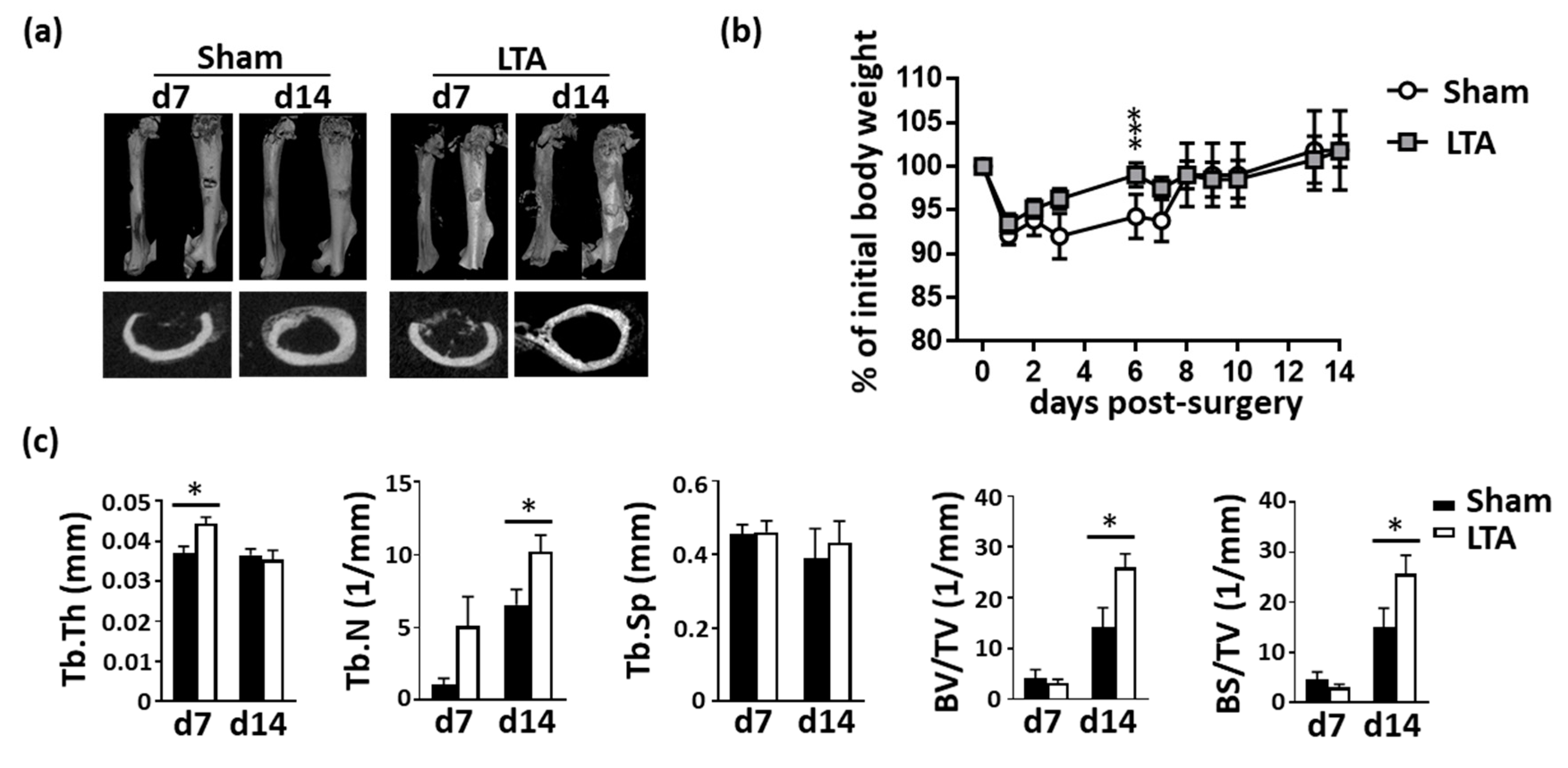
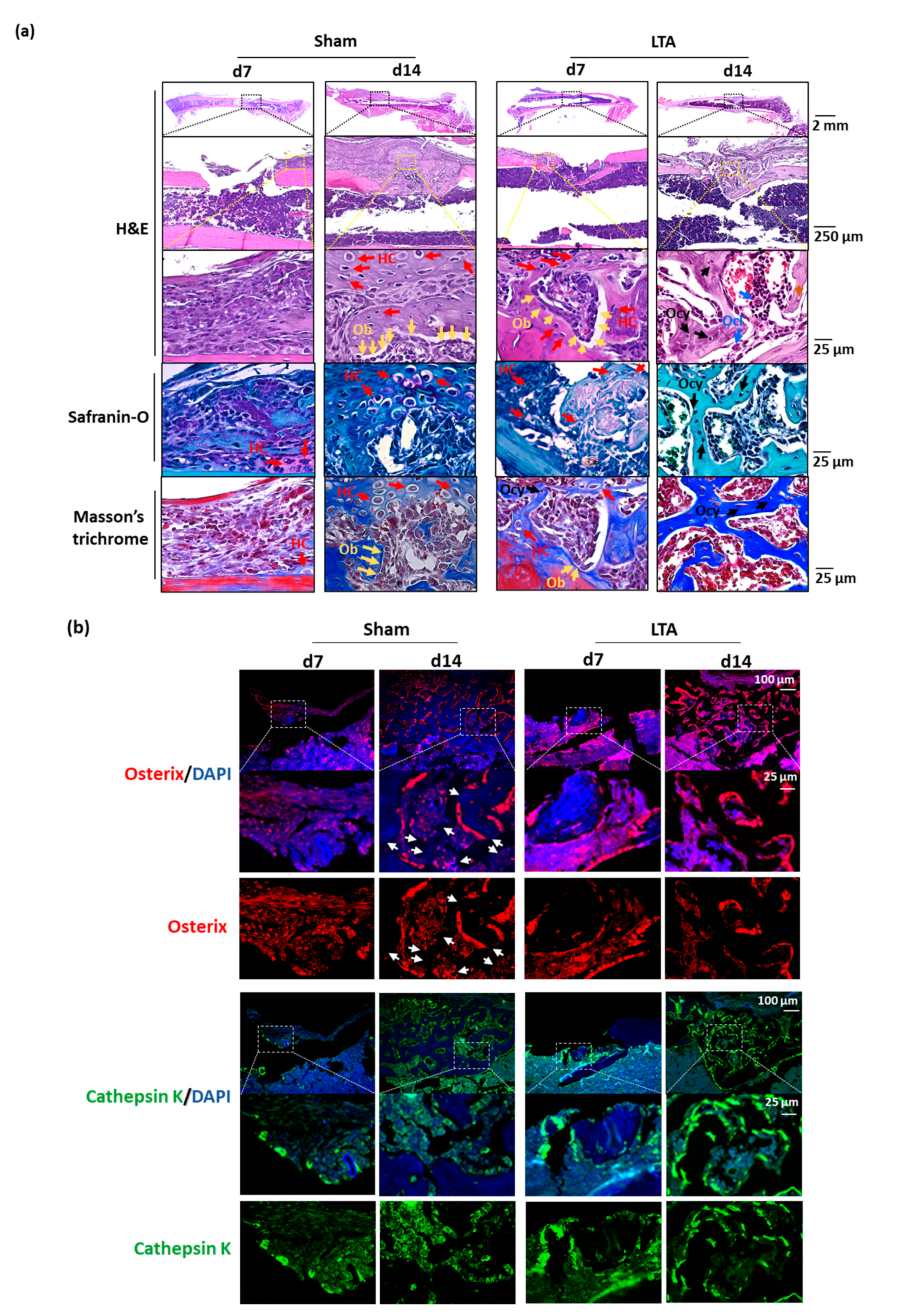
2.1. LTA Accelerated Bone Healing in Mice with Femoral Bone Defects
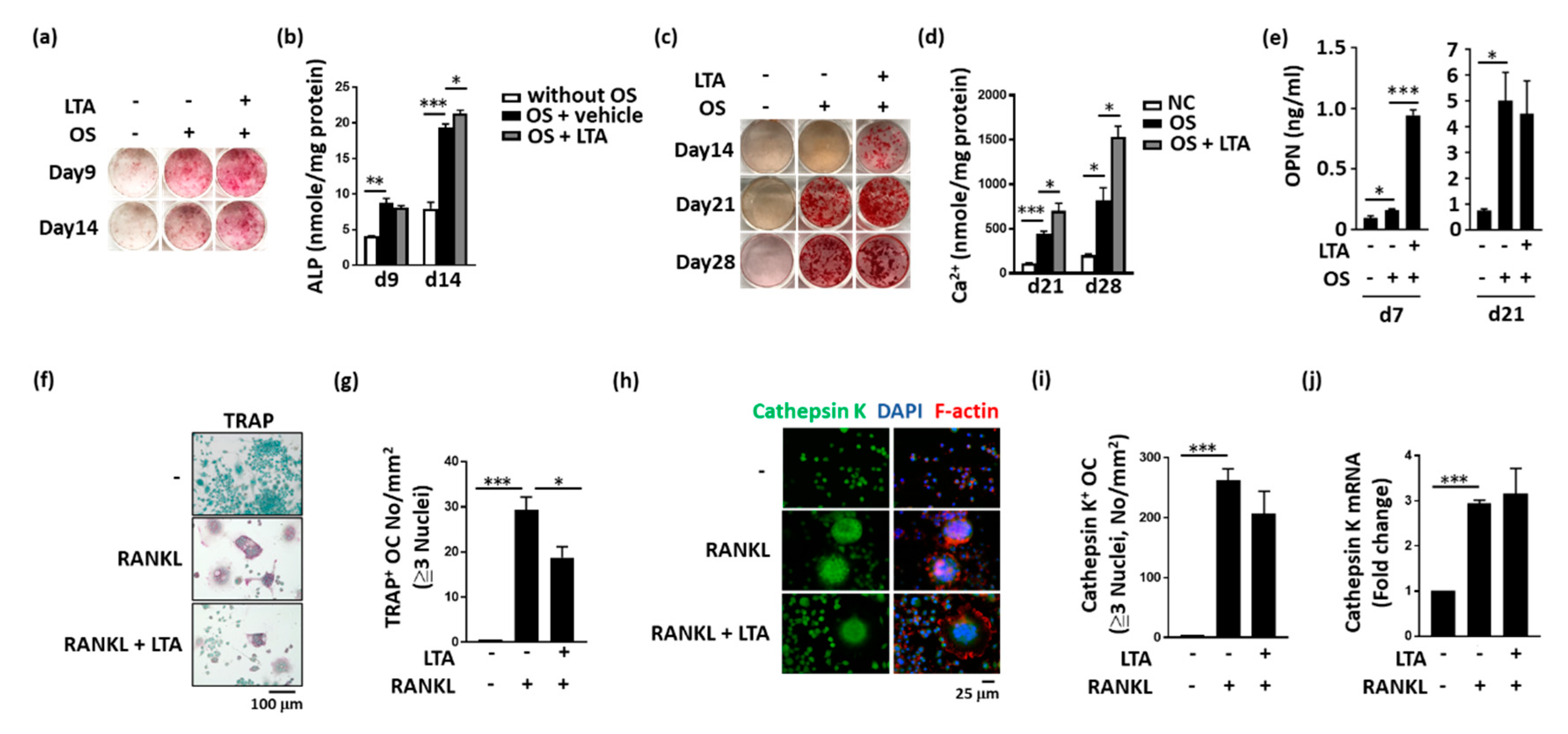
2.2. LTA Had Beneficial Effects on Bone Formation In Vitro
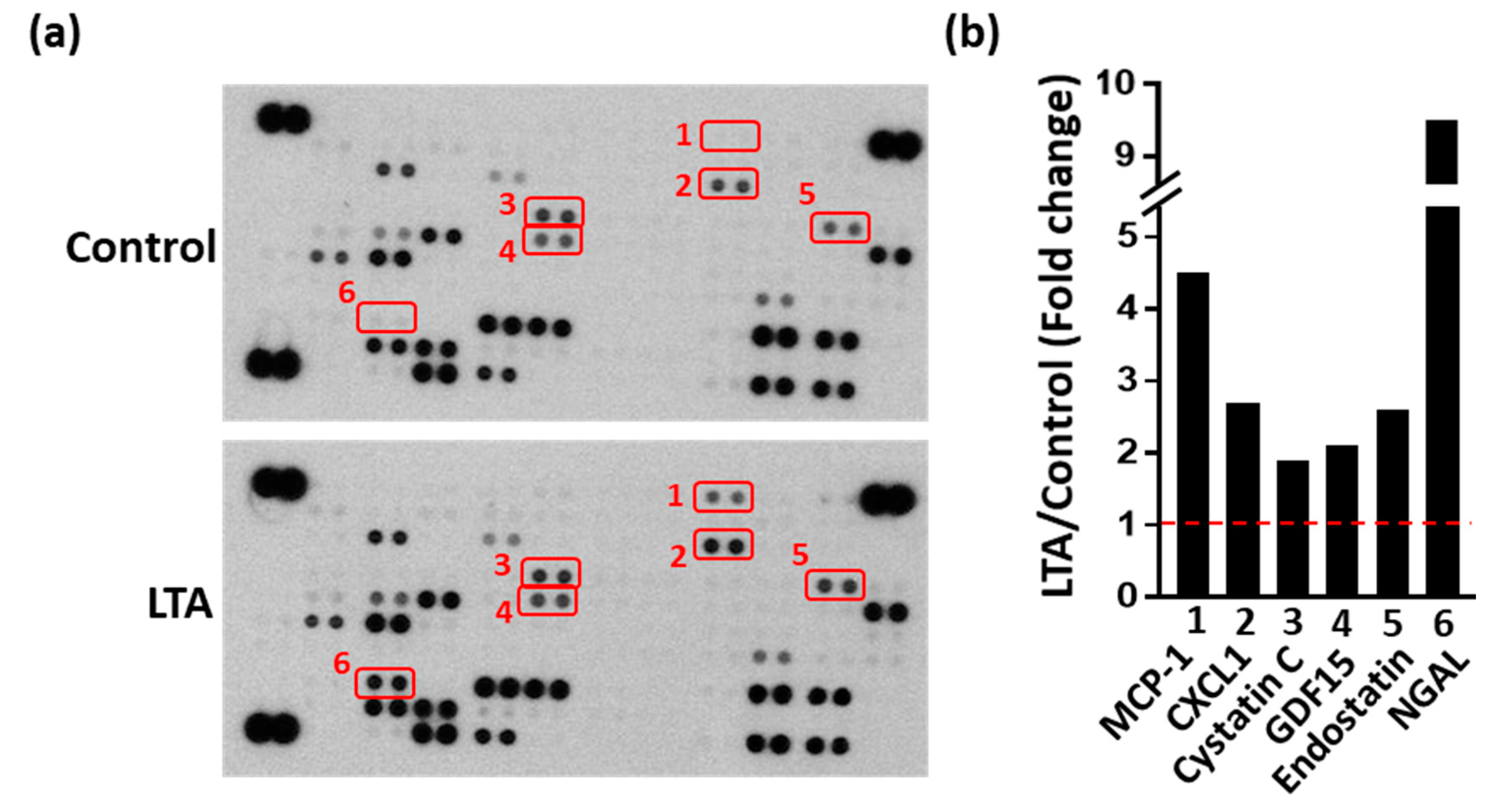
2.3. LTA Positively Regulated Bone Healing by Promoting Protein Secretion from Obs
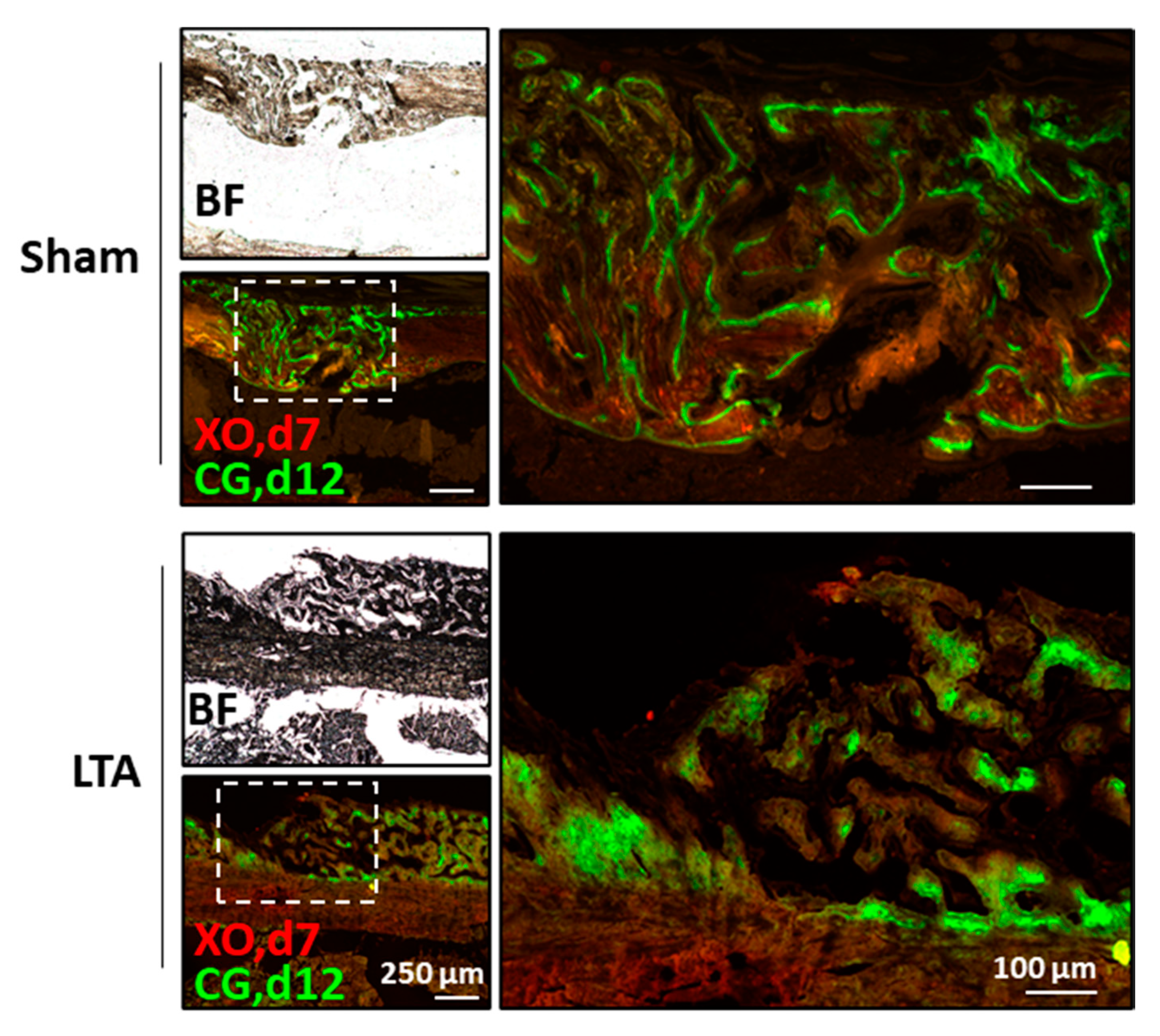
2.4. LTA Enhanced Dynamic Bone-Formation Rates In Vivo
3. Discussion
4. Materials and Methods
4.1. Experimental Animal Study
4.2. Micro-CT Bone Imaging
4.3. Histochemistry and Immunofluorescence Staining
4.4. Ocl and Ob Differentiation
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Protein Array
4.7. Fluorochrome Labeling to Measure Dynamic Bone-Formation Rates
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D | Three-dimensional |
| ALP | Alkaline phosphatase |
| ANOVA | Analysis of variance |
| BMM | Bone marrow-derived macrophage |
| BS | Bone surface |
| BV | Bone volume |
| CG | Calcein green |
| CT | Computed tomography |
| CXCL1 | Chemokine (C-X-C motif) ligand 1 |
| d | Day |
| ELISA | Enzyme-linked immunosorbent assay |
| FBS | Fetal bovine serum |
| GDF15 | Growth/differentiation factor 15 |
| GN | Gram-negative |
| GP | Gram-positive |
| H&E | Hematoxylin and eosin |
| HC | Hypertrophic chondrocyte |
| JNK | c-Jun N-terminal kinase |
| LPS | Lipopolysaccharide |
| LTA | Lipoteichoic acid |
| MAPK | Mitogen-activated protein kinase |
| MCP-1 | Monocyte chemoattractant protein 1 |
| NF-kB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NGAL | Neutrophil gelatinase-associated lipocalin |
| Ob | Osteoblast |
| Ocl | Osteoclast |
| Ocy | Mature osteocyte |
| OPN | Osteopontin |
| OS | Osteogenic factors |
| PC | Proliferation chondrocyte |
| PJI | Periprosthetic joint infection |
| RANKL | Receptor activator of the nuclear factor-kappa B ligand |
| Tb.N | Trabecular number |
| Tb.Sp | Trabecular spacing |
| Tb.Th | Trabecular thickness |
| TLR2 | Toll-like receptor 2 |
| TRAP | Tartrate-resistant acid phosphatase |
| TV | Tissue volume |
| XO | xylene orange |
References
- Schlickewei, C.W.; Kleinertz, H.; Thiesen, D.M.; Mader, K.; Priemel, M.; Frosch, K.H.; Keller, J. Current and future concepts for the treatment of impaired fracture healing. Int. J. Mol. Sci. 2019, 20, 5805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, W.M.; Smith, T.J.; Nicholson, J.A.; Molyneux, S.G.; White, T.O.; Clement, N.D.; Duckworth, A.D. The Radiographic Union Score for HUmeral fractures (RUSHU) predicts humeral shaft nonunion. Bone Joint J. 2019, 101, 1300–1306. [Google Scholar] [CrossRef] [PubMed]
- Wiss, D.A.; Garlich, J.M. Healing the index humeral shaft nonunion: Risk factors for development of a recalcitrant nonunion in 125 patients. J. Bone Joint Surg. Am. 2020, 102, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Chou, W.Y.; Cheng, J.H.; Wang, C.J.; Hsu, S.L.; Chen, J.H.; Huang, C.Y. Shockwave targeting on subchondral bone is more suitable than articular cartilage for knee osteoarthritis. Int. J. Med. Sci. 2019, 16, 156–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Przekora, A. Current trends in fabrication of biomaterials for bone and cartilage regeneration: Materials modifications and biophysical stimulations. Int. J. Mol. Sci. 2019, 20, 435. [Google Scholar] [CrossRef] [Green Version]
- Salem, H.S.; Axibal, D.P.; Wolcott, M.L.; Vidal, A.F.; McCarty, E.C.; Bravman, J.T.; Frank, R.M. Two-stage revision anterior cruciate ligament reconstruction: A systematic review of bone graft options for tunnel augmentation. Am. J. Sports Med. 2020, 48, 767–777. [Google Scholar] [CrossRef]
- El-Hawary, A.; Kandil, Y.R.; Ahmed, M.; Elgeidi, A.; El-Mowafi, H. Distraction subtalar arthrodesis for calcaneal malunion: Comparison of local versus iliac bone graft. Bone Joint J. 2019, 101, 596–602. [Google Scholar] [CrossRef]
- Sigrist, B.; Ferguson, S.; Boehm, E.; Jung, C.; Scheibel, M.; Moroder, P. The biomechanical effect of bone grafting and bone graft remodeling in patients with anterior shoulder instability. Am. J. Sports Med. 2020. [Google Scholar] [CrossRef]
- Roberts, S.J.; Ke, H.Z. Anabolic strategies to augment bone fracture healing. Curr. Osteoporos. Rep. 2018, 16, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Ahlfeld, T.; Doberenz, F.; Kilian, D.; Vater, C.; Korn, P.; Lauer, G.; Lode, A.; Gelinsky, M. Bioprinting of mineralized constructs utilizing multichannel plotting of a self-setting calcium phosphate cement and a cell-laden bioink. Biofabrication 2018, 10, 045002. [Google Scholar] [CrossRef]
- Hwang, K.S.; Choi, J.W.; Kim, J.H.; Chung, H.Y.; Jin, S.; Shim, J.H.; Yun, W.S.; Jeong, C.M.; Huh, J.B. Comparative efficacies of collagen-based 3D printed PCL/PLGA/beta-TCP composite block bone grafts and biphasic calcium phosphate bone substitute for bone regeneration. Materials 2017, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westhauser, F.; Essers, C.; Karadjian, M.; Reible, B.; Schmidmaier, G.; Hagmann, S.; Moghaddam, A. Supplementation with 45S5 bioactive glass reduces in vivo resorption of the beta-tricalcium-phosphate-based bone substitute material vitoss. Int. J. Mol. Sci. 2019, 20, 4253. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, Y.; Cao, Z.; Dou, C.; Bai, Y.; Liu, C.; Dong, S.; Fei, J. Staphylococcal lipoteichoic acid promotes osteogenic differentiation of mouse mesenchymal stem cells by increasing autophagic activity. Biochem. Biophys. Res. Commun. 2017, 485, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.F.; Chang, C.H.; Hu, C.C.; Wu, Y.Y.; Chang, Y.; Ueng, S.W.N. Periprosthetic joint infection caused by gram-positive versus gram-negative bacteria: Lipopolysaccharide, but not lipoteichoic acid, exerts adverse osteoclast-mediated effects on the bone. J. Clin. Med. 2019, 8, 1289. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Park, O.J.; Kim, J.; Baik, J.E.; Yun, C.H.; Han, S.H. Lipoteichoic acid of enterococcus faecalis inhibits the differentiation of macrophages into osteoclasts. J. Endod. 2016, 42, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Dickson, K.; Lehmann, C. Inflammatory response to different toxins in experimental sepsis models. Int. J. Mol. Sci. 2019, 20, 4341. [Google Scholar] [CrossRef] [Green Version]
- Croes, M.; Kruyt, M.C.; Boot, W.; Pouran, B.; Braham, M.V.; Pakpahan, S.A.; Weinans, H.; Vogely, H.C.; Fluit, A.C.; Dhert, W.J.; et al. The role of bacterial stimuli in inflammation-driven bone formation. Eur. Cell. Mater. 2019, 37, 402–419. [Google Scholar] [CrossRef]
- Posner, L.J.; Miligkos, T.; Gilles, J.A.; Carnes, D.L.; Taddeo, D.R.; Graves, D.T. Monocyte chemoattractant protein-1 induces monocyte recruitment that is associated with an increase in numbers of osteoblasts. Bone 1997, 21, 321–327. [Google Scholar] [CrossRef]
- Onan, D.; Allan, E.H.; Quinn, J.M.; Gooi, J.H.; Pompolo, S.; Sims, N.A.; Gillespie, M.T.; Martin, T.J. The chemokine Cxcl1 is a novel target gene of parathyroid hormone (PTH)/PTH-related protein in committed osteoblasts. Endocrinology 2009, 150, 2244–2253. [Google Scholar] [CrossRef] [Green Version]
- Danjo, A.; Yamaza, T.; Kido, M.A.; Shimohira, D.; Tsukuba, T.; Kagiya, T.; Yamashita, Y.; Nishijima, K.; Masuko, S.; Goto, M.; et al. Cystatin C stimulates the differentiation of mouse osteoblastic cells and bone formation. Biochem. Biophys. Res. Commun. 2007, 360, 199–204. [Google Scholar] [CrossRef]
- Johansson, L.; Grubb, A.; Abrahamson, M.; Kasprzykowski, F.; Kasprzykowska, R.; Grzonka, Z.; Lerner, U.H. A peptidyl derivative structurally based on the inhibitory center of cystatin C inhibits bone resorption in vitro. Bone 2000, 26, 451–459. [Google Scholar] [CrossRef]
- Lerner, U.H.; Johansson, L.; Ranjso, M.; Rosenquist, J.B.; Reinholt, F.P.; Grubb, A. Cystatin C, and inhibitor of bone resorption produced by osteoblasts. Acta Physiol. Scand. 1997, 161, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Vanhara, P.; Lincova, E.; Kozubik, A.; Jurdic, P.; Soucek, K.; Smarda, J. Growth/differentiation factor-15 inhibits differentiation into osteoclasts—A novel factor involved in control of osteoclast differentiation. Differentiation 2009, 78, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Gao, R.F.; Yuan, F.L.; Zhao, M.D. Recombinant Human endostatin suppresses mouse osteoclast formation by inhibiting the NF-kappaB and MAPKs signaling pathways. Front. Pharmacol. 2016, 7, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Ohk, B.; Kang, W.Y.; Seong, S.J.; Suk, K.; Lim, M.S.; Kim, S.Y.; Yoon, Y.R. Deficiency of lipocalin-2 promotes proliferation and differentiation of osteoclast precursors via regulation of c-Fms expression and nuclear factor-kappa B activation. J. Bone Metab. 2016, 23, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Yoon, H.J.; Yoon, K.A.; Gwon, M.R.; Jin Seong, S.; Suk, K.; Kim, S.Y.; Yoon, Y.R. Lipocalin-2 inhibits osteoclast formation by suppressing the proliferation and differentiation of osteoclast lineage cells. Exp. Cell Res. 2015, 334, 301–309. [Google Scholar] [CrossRef]
- Sipola, A.; Nelo, K.; Hautala, T.; Ilvesaro, J.; Tuukkanen, J. Endostatin inhibits VEGF-A induced osteoclastic bone resorption in vitro. BMC Musculoskelet. Disord. 2006, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Ryu, Y.H.; Yun, C.H.; Han, S.H. Impaired osteoclastogenesis by staphylococcal lipoteichoic acid through Toll-like receptor 2 with partial involvement of MyD88. J. Leukoc. Biol. 2009, 86, 823–831. [Google Scholar] [CrossRef]
- Wang, S.; Heng, B.C.; Qiu, S.; Deng, J.; Shun Pan Cheung, G.; Jin, L.; Zhao, B.; Zhang, C. Lipoteichoic acid of Enterococcus faecalis inhibits osteoclastogenesis via transcription factor RBP-J. Innate Immun. 2019, 25, 13–21. [Google Scholar] [CrossRef]
- Sinder, B.P.; Pettit, A.R.; McCauley, L.K. Macrophages: Their emerging roles in bone. J. Bone Miner. Res. 2015, 30, 2140–2149. [Google Scholar] [CrossRef] [Green Version]
- Alexander, K.A.; Chang, M.K.; Maylin, E.R.; Kohler, T.; Muller, R.; Wu, A.C.; Van Rooijen, N.; Sweet, M.J.; Hume, D.A.; Raggatt, L.J.; et al. Osteal macrophages promote in vivo intramembranous bone healing in a mouse tibial injury model. J. Bone Miner. Res. 2011, 26, 1517–1532. [Google Scholar] [CrossRef] [PubMed]
- Schlundt, C.; El Khassawna, T.; Serra, A.; Dienelt, A.; Wendler, S.; Schell, H.; van Rooijen, N.; Radbruch, A.; Lucius, R.; Hartmann, S.; et al. Macrophages in bone fracture healing: Their essential role in endochondral ossification. Bone 2018, 106, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Hozain, S.; Cottrell, J. CDllb+ targeted depletion of macrophages negatively affects bone fracture healing. Bone 2020, 138, 115479. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gareta, E.; Hua, J.; Blunn, G.W. Osseointegration of acellular and cellularized osteoconductive scaffolds: Is tissue engineering using mesenchymal stem cells necessary for implant fixation? J. Biomed. Mater. Res. Part A 2015, 103, 1067–1076. [Google Scholar] [CrossRef]
- Zhou, W.; Han, C.; Song, Y.; Yan, X.; Li, D.; Chai, Z.; Feng, Z.; Dong, Y.; Li, L.; Chen, F.; et al. The performance of bone marrow mesenchymal stem cell-implant complexes prepared by cell sheet engineering techniques. Biomaterials 2010, 31, 3212–3221. [Google Scholar] [CrossRef]
- Morath, S.; von Aulock, S.; Hartung, T. Structure/function relationships of lipoteichoic acids. J. Endotoxin Res. 2005, 11, 348–356. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.; Hsiao, Y.M.; Hu, C.C.; Chang, C.H.; Li, C.Y.; Ueng, S.W.N.; Chen, M.F. Synovial fluid interleukin-16 contributes to osteoclast activation and bone loss through the JNK/NFATc1 signaling cascade in patients with periprosthetic joint infection. Int. J. Mol. Sci. 2020, 21, 2904. [Google Scholar] [CrossRef] [Green Version]
- van Gaalen, S.M.; Kruyt, M.C.; Geuze, R.E.; de Bruijn, J.D.; Alblas, J.; Dhert, W.J. Use of fluorochrome labels in in vivo bone tissue engineering research. Tissue Eng. Part B Rev. 2010, 16, 209–217. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, C.-C.; Chang, C.-H.; Hsiao, Y.-m.; Chang, Y.; Wu, Y.-Y.; Ueng, S.W.N.; Chen, M.-F. Lipoteichoic Acid Accelerates Bone Healing by Enhancing Osteoblast Differentiation and Inhibiting Osteoclast Activation in a Mouse Model of Femoral Defects. Int. J. Mol. Sci. 2020, 21, 5550. https://doi.org/10.3390/ijms21155550
Hu C-C, Chang C-H, Hsiao Y-m, Chang Y, Wu Y-Y, Ueng SWN, Chen M-F. Lipoteichoic Acid Accelerates Bone Healing by Enhancing Osteoblast Differentiation and Inhibiting Osteoclast Activation in a Mouse Model of Femoral Defects. International Journal of Molecular Sciences. 2020; 21(15):5550. https://doi.org/10.3390/ijms21155550
Chicago/Turabian StyleHu, Chih-Chien, Chih-Hsiang Chang, Yi-min Hsiao, Yuhan Chang, Ying-Yu Wu, Steve W. N. Ueng, and Mei-Feng Chen. 2020. "Lipoteichoic Acid Accelerates Bone Healing by Enhancing Osteoblast Differentiation and Inhibiting Osteoclast Activation in a Mouse Model of Femoral Defects" International Journal of Molecular Sciences 21, no. 15: 5550. https://doi.org/10.3390/ijms21155550
APA StyleHu, C. -C., Chang, C. -H., Hsiao, Y. -m., Chang, Y., Wu, Y. -Y., Ueng, S. W. N., & Chen, M. -F. (2020). Lipoteichoic Acid Accelerates Bone Healing by Enhancing Osteoblast Differentiation and Inhibiting Osteoclast Activation in a Mouse Model of Femoral Defects. International Journal of Molecular Sciences, 21(15), 5550. https://doi.org/10.3390/ijms21155550