Upregulation of the TRPA1 Ion Channel in the Gastric Mucosa after Iodoacetamide-Induced Gastritis in Rats: A Potential New Therapeutic Target

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Macroscopic Evaluation of Rat Gastric Mucosa
2.2. Microscopic Alterations in the Inflamed Rat Gastric Mucosa
2.3. Weight Change and Water Consumption
2.4. GSH Concentration of the Rat Stomach Tissue
2.5. Quantification of TRPA1 and TRPV1 Immunopositivity
2.6. Trpa1, Trpv1 Relative Gene Expression Changes in the Inflamed Rat Stomach
2.7. IAA-Induced Alterations in Mice
3. Discussion
4. Materials and Methods
4.1. Animals and Ethics
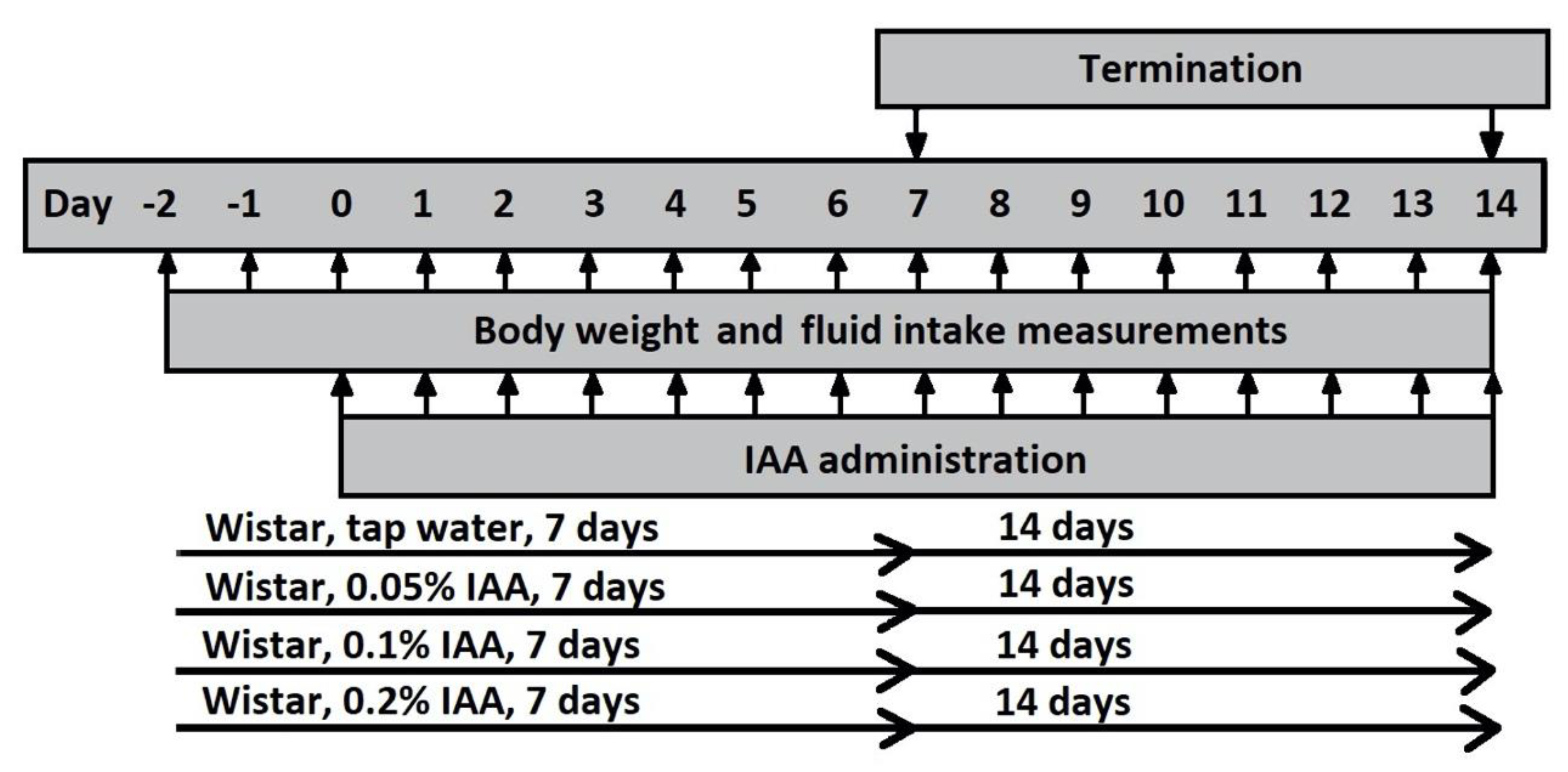
4.2. Experimental Protocol
4.3. Study in Rats
4.4. Study in Mice
4.5. Macroscopic and Microscopic Evaluations of IAA-Induced Gastric Lesions
4.6. GSH Measurement
4.7. TRPV1 and TRPA1 Immunohistochemistry and Scoring
4.8. Determination of Trpv1 and Trpa1 Relative Gene Expression
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Szabo, I.L.; Cseko, K.; Czimmer, J.; Mozsik, G. Diagnosis of Gastritis—Review from Early Pathological Evaluation to Present Day Management; Mozsik, G., Ed.; InTech Open Access Publisher: Rijeka, Yugoslavia, 2013; pp. 3–20. [Google Scholar]
- Lu, C.L.; Chang, S.S.; Wang, S.S.; Chang, F.Y.; Lee, S.D. Silent peptic ulcer disease: Frequency, factors leading to “silence,” and implications regarding the pathogenesis of visceral symptoms. Gastrointest. Endosc. 2004, 60, 34–38. [Google Scholar] [CrossRef]
- Appelman, H.D. Gastritis: Terminology, Etiology, and Clinicopathological Correlations: Another Biased View. Hum. Pathol. 1994, 25, 1006–1019. [Google Scholar] [CrossRef] [Green Version]
- McColl, K.E.L. Helicobacter pylori-Negative Nonsteroidal Anti-Inflammatory Drug-Negative Ulcer. Gastroenterol. Clin. N. Am. 2009, 38, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Kavitt, R.T.; Lipowska, A.M.; Anyane-Yeboa, A.; Gralnek, I.M. Diagnosis and Treatment of Peptic Ulcer Disease. Am. J. Med. 2019, 132, 447–456. [Google Scholar] [CrossRef]
- Szolcsanyi, J.; Barthó, L. Impaired defense mechanism to peptic ulcer in the capsaicin-desensitized rat. In Gastrointestinal Defense Mechanism; Mózsik, G., Hanninen, O., Jávor, T., Eds.; Akadémiai Kiadó-Pergamon Press: Budapest, Hungary, 1981; pp. 39–51. [Google Scholar]
- Abdel-Salam, O.M.E.; Debreceni, A.; Mózsik, G.; Szolcsányi, J. Capsaicin-sensitive afferent sensory nerves in modulating gastric mucosal defense against noxious agents. J. Physiol. Paris 1999, 93, 443–454. [Google Scholar] [CrossRef]
- Szolcsányi, J.; Barthó, L. Capsaicin-sensitive afferents and their role in gastroprotection: An update. J. Physiol. Paris 2001, 95, 181–188. [Google Scholar] [CrossRef]
- Geppetti, P.; Nassini, R.; Materazzi, S.; Benemei, S. The concept of neurogenic inflammation. BJU Int. 2008, 101, 2–6. [Google Scholar] [CrossRef]
- Talavera, K.; Startek, J.B.; Alvarez-Collazo, J.; Boonen, B.; Alpizar, Y.A.; Sanchez, A.; Naert, R.; Nilius, B. Mammalian Transient Receptor Potential TRPA1 Channels: From Structure to Disease. Physiol. Rev. 2020, 100, 725–803. [Google Scholar] [CrossRef]
- Giorgi, S.; Nikolaeva-Koleva, M.; Alarcón-Alarcón, D.; Butrón, L.; González-Rodríguez, S. Is TRPA1 burning down TRPV1 as druggable target for the treatment of chronic pain? Int. J. Mol. Sci. 2019, 20, 2906. [Google Scholar] [CrossRef] [Green Version]
- Caterina, M.J.; Park, U. TRPV1: A Polymodal Sensor in the Nociceptor Terminal. Curr. Top. Membr. 2006, 57, 113–150. [Google Scholar] [CrossRef]
- Viana, F. TRPA1 channels: Molecular sentinels of cellular stress and tissue damage. J. Physiol. 2016, 594, 4151–4169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szolcsányi, J.; Helyes, Z.; Oroszi, G.; Németh, J.; Pintér, E. Release of somatostatin and its role in the mediation of the anti-inflammatory effect induced by antidromic stimulation of sensory fibres of rat sciatic nerve. Br. J. Pharmacol. 1998, 123, 936–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szolcsányi, J. Forty years in capsaicin research for sensory pharmacology and physiology. Neuropeptides 2004, 38, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Sgouros, S.N.; Bergele, C.; Viazis, N.; Avgerinos, A. Somatostatin and its analogues in peptic ulcer bleeding: Facts and pathophysiological aspects. Dig. Liver Dis. 2006, 38, 143–148. [Google Scholar] [CrossRef]
- Faussone-Pellegrini, M.S.; Taddei, A.; Bizzoco, E.; Lazzeri, M.; Vannucchi, M.G.; Bechi, P. Distribution of the vanilloid (capsaicin) receptor type 1 in the human stomach. Histochem. Cell Biol. 2005, 124, 61–68. [Google Scholar] [CrossRef]
- Kun, J.; Szitter, I.; Kemény, Á.; Perkecz, A.; Kereskai, L.; Pohóczky, K.; Vincze, Á.; Gódi, S.; Szabó, I.; Szolcsányi, J.; et al. Upregulation of the transient receptor potential ankyrin 1 ion channel in the inflamed human and mouse colon and its protective roles. PLoS ONE 2014, 9, e108164. [Google Scholar] [CrossRef]
- Poole, D.P.; Pelayo, J.C.; Cattaruzza, F.; Kuo, Y.; Gai, G.; Chiu, J.V.; Bron, R.; Furness, J.B.; Grady, E.F.; Bunnett, N.W. Transient receptor potential ankyrin 1 is expressed by inhibitory motoneurons of the mouse intestine. Gastroenterology 2011, 141, 565–575. [Google Scholar] [CrossRef]
- Kaji, I.; Yasuoka, Y.; Karaki, S.; Kuwahara, A. Activation of TRPA1 by luminal stimuli induces EP 4-mediated anion secretion in human and rat colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 690–701. [Google Scholar] [CrossRef] [Green Version]
- Bertin, S.; Aoki-Nonaka, Y.; De Jong, P.R.; Nohara, L.L.; Xu, H.; Stanwood, S.R.; Srikanth, S.; Lee, J.; To, K.; Abramson, L.; et al. The ion channel TRPV1 regulates the activation and proinflammatory properties of CD4+ T cells. Nat. Immunol. 2014, 15, 1055–1063. [Google Scholar] [CrossRef]
- Bertin, S.; Aoki-Nonaka, Y.; Lee, J.; de Jong, P.R.; Kim, P.; Han, T.; Yu, T.; To, K.; Takahashi, N.; Boland, B.S.; et al. The TRPA1 ion channel is expressed in CD4+ T cells and restrains T cell-mediated colitis through inhibition of TRPV1. Gut 2017, 66, 1584–1596. [Google Scholar] [CrossRef]
- Csekő, K.; Beckers, B.; Keszthelyi, D.; Helyes, Z. Role of TRPV1 and TRPA1 Ion Channels in Inflammatory Bowel Diseases: Potential Therapeutic Targets? Pharmaceuticals 2019, 12, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utsumi, D.; Matsumoto, K.; Tsukahara, T.; Amagase, K.; Tominaga, M.; Kato, S. Transient receptor potential vanilloid 1 and transient receptor potential ankyrin 1 contribute to the progression of colonic inflammation in dextran sulfate sodium-induced colitis in mice: Links to calcitonin gene-related peptide and substance P. J. Pharmacol. Sci. 2018, 136, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Massa, F.; Sibaev, A.; Marsicano, G.; Blaudzun, H.; Storr, M.; Lutz, B. Vanilloid receptor (TRPV1)-deficient mice show increased susceptibility to dinitrobenzene sulfonic acid induced colitis. J. Mol. Med. 2006, 84, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Mózsik, G.; Szolcsányi, J.; Rácz, I. Gastroprotection induced by capsaicin in healthy human subjects. World J. Gastroenterol. 2005, 11, 5180–5184. [Google Scholar] [CrossRef] [PubMed]
- Mózsik, G. Capsaicin as New orally Applicable Gastroprotective and Therapeutic Drug Alone or in Combination with Nonsteroidal Anti-Inflammatory Drugs in Healthy Human Subjects and in Patients; Abdel-Salam, O.M.E., Ed.; Springer: Basel, Switzerland, 2014; Volume 68, ISBN 9783034808279. [Google Scholar]
- Yu, X.; Yu, M.; Liu, Y.; Yu, S. TRP channel functions in the gastrointestinal tract. Semin. Immunopathol. 2015, 38, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Szabo, S.; Trier, J.S.; Brown, A.; Schnoor, J. Sulfhydryl blockers induce severe inflammatory gastritis in the rat. Gastroenterology 1984, 86, 1271. [Google Scholar]
- Szabo, S.; Trier, J.S.; Frankel, P.W. Sulfhydryl compounds may mediate gastric cytoprotection. Science 1981, 214, 200–202. [Google Scholar] [CrossRef]
- Pihan, G.; Majzoubi, D.; Haudenschild, C.; Trier, J.S.; Szabo, S. Early microcirculatory stasis in acute gastric mucosal injury in the rat and prevention by 16,16-dimethyl prostaglandin E2 or sodium thiosulfate. Gastroenterology 1986, 91, 1415–1426. [Google Scholar] [CrossRef]
- Szabo, S.; Brown, A. Prevention of ethanol-induced vascular injury and gastric mucosal lesions by sucralfate and its components: Possible role of endogenous sulfhydryls. Proc. Soc. Expl. Biol. Med. 1987, 185, 493–497. [Google Scholar] [CrossRef]
- Lee, S.E.; Song, H.J.; Park, S.Y.; Nam, Y.; Min, C.H.; Lee, D.Y.; Jeong, J.Y.; Ha, H.S.; Kim, H.-J.; Whang, W.K.; et al. Effect of ECQ on iodoacetamide-induced chronic gastritis in rats. Korean J. Physiol. Pharmacol. 2013, 17, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Karmeli, F.; Okon, E.; Rachmilewitz, D. Sulphydryl blocker induced gastric damage is ameliorated by scavenging of free radicals. Gut 1996, 38, 826–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastoris, O.; Verri, M.; Boschi, F.; Kastsiuchenka, O.; Balestra, B.; Pace, F.; Tonini, M.; Natale, G. Effects of esomeprazole on glutathione levels and mitochondrial oxidative phosphorylation in the gastric mucosa of rats treated with indomethacin. Naunyn. Schmiedebergs. Arch. Pharmacol. 2008, 378, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Michaelsen, J.T.; Dehnert, S.; Giustarini, D.; Beckmann, B.; Tsikas, D. HPLC analysis of human erythrocytic glutathione forms using OPA and N-acetyl-cysteine ethyl ester: Evidence for nitrite-induced GSH oxidation to GSSG. J. Chromatogr. B 2009, 877, 3405–3417. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.M.; Dringen, R. Differential effects of iodoacetamide and iodoacetate on glycolysis and glutathione metabolism of cultured astrocytes. Front. Neuroenergetics 2009, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, D.B.; Acosta, D.; Bruckner, J.V. Role of glutathione depletion in the cytotoxicity of acetaminophen in a primary culture system of rat hepatocytes. Toxicology 1985, 37, 127–146. [Google Scholar] [CrossRef]
- Palmen, N.G.M.; Evelo, C.T.A. Glutathione depletion in human erythrocytes and rat liver: A study on the interplay between bioactivation and inactivation functions of liver and blood. Toxicol. Vitr. 1996, 10, 273–281. [Google Scholar] [CrossRef]
- Elseweidy, M.M.; Younis, N.N.; Amin, R.S.; Abdallah, F.R.; Fathy, A.M.; Yousif, Z.A. Effect of some natural products either alone or in combination on gastritis induced in experimental rats. Dig. Dis. Sci. 2008, 53, 1774–1784. [Google Scholar] [CrossRef]
- Rahman, I.; Antonicelli, F.; MacNee, W. Molecular mechanism of the regulation of glutathione synthesis by tumour necrosis factor-a and dexamethasone in human alveolar epithelial cells. J. Biol. Chem. 1999, 274, 5088–5096. [Google Scholar] [CrossRef] [Green Version]
- Yasin, R.; Leese, C.L. The production of chronic gastritis and ulceration in the glandular stomach of rats by iodoacetamide (IAM). Eur. J. Cancer 1970, 6, 425–432. [Google Scholar] [CrossRef]
- Dial, E.J.; Hall, L.R.; Romero, J.J.; Lichtenberger, L.M. Rats with gastritis have increased sensitivity to the gastrin stimulatory effects of luminal ammonia. Gastroenterology 1996, 110, 801–808. [Google Scholar] [CrossRef]
- Barnett, K.; Bell, C.J.; McKnight, W.; Dicay, M.; Sharkey, K.A.; Wallace, J.L. Role of cyclooxygenase-2 in modulating gastric acid secretion in the normal and inflamed rat stomach. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, 1292–1297. [Google Scholar] [CrossRef] [PubMed]
- Lalich, J.J. Iodoacetamide induced gastric ulcers in rats. Proc. Soc. Expl. Biol. Med. 1962, 109, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, N.; Bielefeldt, K.; Sengupta, J.N.; Gebhart, G.F. Models of gastric hyperalgesia in the rat. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, 666–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larauche, M.; Anton, P.M.; Peiro, G.; Eutamène, H.; Buéno, L.; Fioramonti, J. Role of capsaicin-sensitive afferent nerves in different models of gastric inflammation in rats. Auton. Neurosci. Basic Clin. 2004, 110, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, L.J.; Dubin, A.E.; Evans, M.J.; Marr, F.; Schultz, P.G.; Cravatt, B.F.; Patapoutian, A. Noxious compounds activate TRPA1 ion channels through covalent modification of cysteines. Nature 2007, 445, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Hinman, A.; Chuang, H.H.; Bautista, D.M.; Julius, D. TRP channel activation by reversible covalent modification. Proc. Natl. Acad. Sci. USA 2006, 103, 19564–19568. [Google Scholar] [CrossRef] [Green Version]
- Ibarra, Y.; Blair, N.T. Benzoquinone reveals a cysteine-dependent desensitization mechanism of TRPA1. Mol. Pharmacol. 2013, 83, 1120–1132. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Jia, J.; Xie, C.; Wu, Y.; Tu, W. Transient Receptor Potential Ankyrin 1 and Substance P Mediate the Development of Gastric Mucosal Lesions in a Water Immersion Restraint Stress Rat Model. Digestion 2018, 97, 228–239. [Google Scholar] [CrossRef]
- Dömötör, A.; Kereskay, L.; Szekeres, G.; Hunyady, B.; Szolcsányi, J.; Mózsik, G. Participation of capsaicin-sensitive afferent nerves in the gastric mucosa of patients with Helicobacter pylori-positive or-negative chronic gastritis. Dig. Dis. Sci. 2007, 52, 411–417. [Google Scholar] [CrossRef]
- Fernandes, E.S.; Fernandes, M.A.; Keeble, J.E. The functions of TRPA1 and TRPV1: Moving away from sensory nerves. Br. J. Pharmacol. 2012, 166, 510–521. [Google Scholar] [CrossRef] [Green Version]
- Ruparel, N.B.; Patwardhan, A.M.; Akopian, A.N.; Hargeaves, K.M. Homologous and Heterologous Desensitization of Capsaicin and Mustard Oil Responses Utilize Different Cellular Pathways in Nociceptors. Pain 2008, 135, 271–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielefeldt, K.; Ozaki, N.; Gebhart, G.F. Mild gastritis alters voltage-sensitive sodium currents in gastric sensory neurons in rats. Gastroenterology 2002, 122, 752–761. [Google Scholar] [CrossRef] [PubMed]
- Piqueras, L.; Corpa, J.M.; Martínez, J.; Martínez, V. Gastric hypersecretion associated to iodoacetamide-induced mild gastritis in mice. Naunyn Schmiedeberg’s Arch. Pharmacol. 2003, 367, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Holzer, P.; Wultsch, T.; Edelsbrunner, M.; Mitrovic, M.; Shahbazian, A.; Painsipp, E.; Bock, E.; Pabst, M.A. Increase in gastric acid-induced afferent input to the brainstem in mice with gastritis. Neuroscience 2007, 145, 1108–1119. [Google Scholar] [CrossRef] [Green Version]
- Wultsch, T.; Painsipp, E.; Shahbazian, A.; Mitrovic, M.; Edelsbrunner, M.; Waldmann, R.; Lazdunski, M.; Holzer, P. Deletion of the acid-sensing ion channel ASIC3 prevents gastritis-induced acid hyperresponsiveness of the stomach-brainstem axis. Pain 2015, 134, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Painsipp, E.; Wultsch, T.; Shahbazian, A.; Edelsbrunner, M.; Kreissl, C.; Schirbel, A.; Bock, E.; Pabst, M.A.; Thoeringer, C.K.; Huber, H.P.; et al. Experimental gastritis in mice enhances anxiety in a gender- related manner. Neuroscience 2007, 150, 522–536. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Du, S.; Kong, C.; Zhang, Z.; Mokhtar, A. Intrathecal administration of TRPA1 antagonists attenuate cyclophosphamide-induced cystitis in rats with hyper-reflexia micturition. BMC Urol. 2016, 16. [Google Scholar] [CrossRef] [Green Version]
- Price, T.J.; Louria, M.D.; Candelario-Soto, D.; Dussor, G.O.; Jeske, N.A.; Patwardhan, A.M.; Diogenes, A.; Trott, A.A.; Hargreaves, K.M.; Flores, C.M. Treatment of trigeminal ganglion neurons in vitro with NGF, GDNF or BDNF: Effects on neuronal survival, neurochemical properties and TRPV1-mediated neuropeptide secretion. BMC Neurosci. 2005, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kemény, Á.; Kodji, X.; Horváth, S.; Komlódi, R.; Szőke, É.; Sándor, Z.; Perkecz, A.; Gyömörei, C.; Sétáló, G.; Kelemen, B.; et al. TRPA1 acts in a protective manner in imiquimod-induced psoriasiform dermatitis in mice. J. Investig. Dermatol. 2018, 138, 1774–1784. [Google Scholar] [CrossRef] [Green Version]
- Bohonyi, N.; Pohóczky, K.; Szalontai, B.; Perkecz, A.; Kovács, K.; Kajtár, B.; Orbán, L.; Varga, T.; Szegedi, S.; Bódis, J.; et al. Local upregulation of transient receptor potential ankyrin I and transient receptor potential vanilloid I ion channels in rectosigmoid deep infiltrating endometriosis. Mol. Pain 2017, 13, 1–13. [Google Scholar] [CrossRef]
- Kun, J.; Helyes, Z.; Perkecz, A.; Bán, Á.; Polgár, B.; Szolcsányi, J.; Pintér, E. Effect of Surgical and Chemical Sensory Denervation on Non-neural Expression of the Transient Receptor Potential Vanilloid 1 (TRPV1) Receptors in the Rat. J. Mol. Neurosci. 2012, 48, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Potolicchio, I.; Santambrogio, L.; Strominger, J.L. Molecular interaction and enzymatic activity of macrophage migration inhibitory factor with immunorelevant peptides. J. Biol. Chem. 2003, 278, 30889–30895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuschwander-Teri, B.A.; Roll, F.J. Glutathione measurement by HPLC separation and fluorometric detection of the glutathione Orthophthalaldehyde adduct. Anal. Biochem. 1989, 179, 236–241. [Google Scholar] [CrossRef]
- Parmentier, C.; Leroy, P.; Wellman, M.; Nicolas, A. Determination of cellular thiols and glutathione-related enzyme activities: Versatility of high-performance liquid chromatography-spectrofluorimetric detection. J. Chromatogr. B 1998, 719, 37–46. [Google Scholar] [CrossRef]
- Bollenbach, A.; Tsikas, D. Measurement of the tripeptides glutathione and ophthalmic acid by gas chromatography-mass spectrometry. Anal. Biochem. 2020. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | IAA Conc. | Duration | Weight Loss | Water Intake | Macroscopic and Microscopic Picture |
|---|---|---|---|---|---|
| CD1 | 0.1% | 14 days | ~17% | ↓ | negative |
| CD1 | 0.1% | 7 days | ~14% | ↓ | negative |
| CD1 | 0.3% | 7 days | ~28% | ↓ | negative |
| CD1 | 0.5% | 7 days | ~42% | ↓↓ | negative |
| C57Bl/6J | 0.3% | 7 days | ~14% | ↓ | negative |
| C57Bl/6J | 0.3% + 2% sucrose | 7 days | ~21% | ↓↓ | negative |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Csekő, K.; Pécsi, D.; Kajtár, B.; Hegedűs, I.; Bollenbach, A.; Tsikas, D.; Szabó, I.L.; Szabó, S.; Helyes, Z. Upregulation of the TRPA1 Ion Channel in the Gastric Mucosa after Iodoacetamide-Induced Gastritis in Rats: A Potential New Therapeutic Target. Int. J. Mol. Sci. 2020, 21, 5591. https://doi.org/10.3390/ijms21165591
Csekő K, Pécsi D, Kajtár B, Hegedűs I, Bollenbach A, Tsikas D, Szabó IL, Szabó S, Helyes Z. Upregulation of the TRPA1 Ion Channel in the Gastric Mucosa after Iodoacetamide-Induced Gastritis in Rats: A Potential New Therapeutic Target. International Journal of Molecular Sciences. 2020; 21(16):5591. https://doi.org/10.3390/ijms21165591
Chicago/Turabian StyleCsekő, Kata, Dániel Pécsi, Béla Kajtár, Ivett Hegedűs, Alexander Bollenbach, Dimitrios Tsikas, Imre László Szabó, Sándor Szabó, and Zsuzsanna Helyes. 2020. "Upregulation of the TRPA1 Ion Channel in the Gastric Mucosa after Iodoacetamide-Induced Gastritis in Rats: A Potential New Therapeutic Target" International Journal of Molecular Sciences 21, no. 16: 5591. https://doi.org/10.3390/ijms21165591
APA StyleCsekő, K., Pécsi, D., Kajtár, B., Hegedűs, I., Bollenbach, A., Tsikas, D., Szabó, I. L., Szabó, S., & Helyes, Z. (2020). Upregulation of the TRPA1 Ion Channel in the Gastric Mucosa after Iodoacetamide-Induced Gastritis in Rats: A Potential New Therapeutic Target. International Journal of Molecular Sciences, 21(16), 5591. https://doi.org/10.3390/ijms21165591