Functional Aspects of Seminal Plasma in Bird Reproduction


Abstract
:1. Introduction
2. Bird Semen Biology
2.1. Semen Biology in the Male Tract
2.2. Males Accessory Tissues and Extra Seminal Fluids
2.3. The Sperm in the Female Tract
3. Seminal Plasma Composition
3.1. Inorganic Ions
3.2. Carbohydrates
3.3. Lipids
3.4. Hormones
3.5. Amino Acids
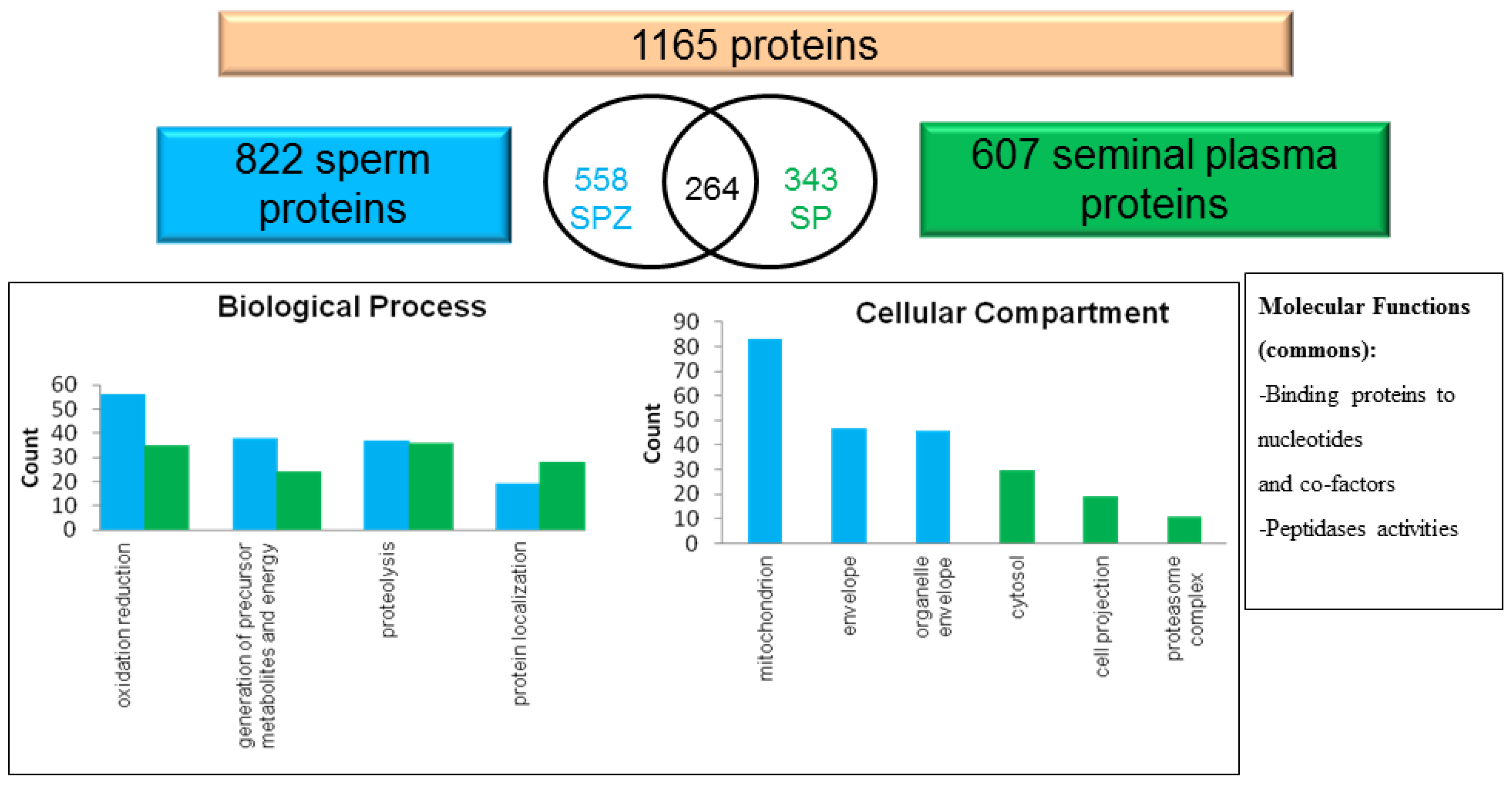
3.6. Proteins
4. Role of Seminal Proteins and Peptides on Sperm Function
4.1. Regulation of Membrane Stability and Fertilization Process
4.2. Sperm Motility
4.3. Antimicrobial Effect and Fertilizing Capacity
4.4. Antioxidant Enzymes
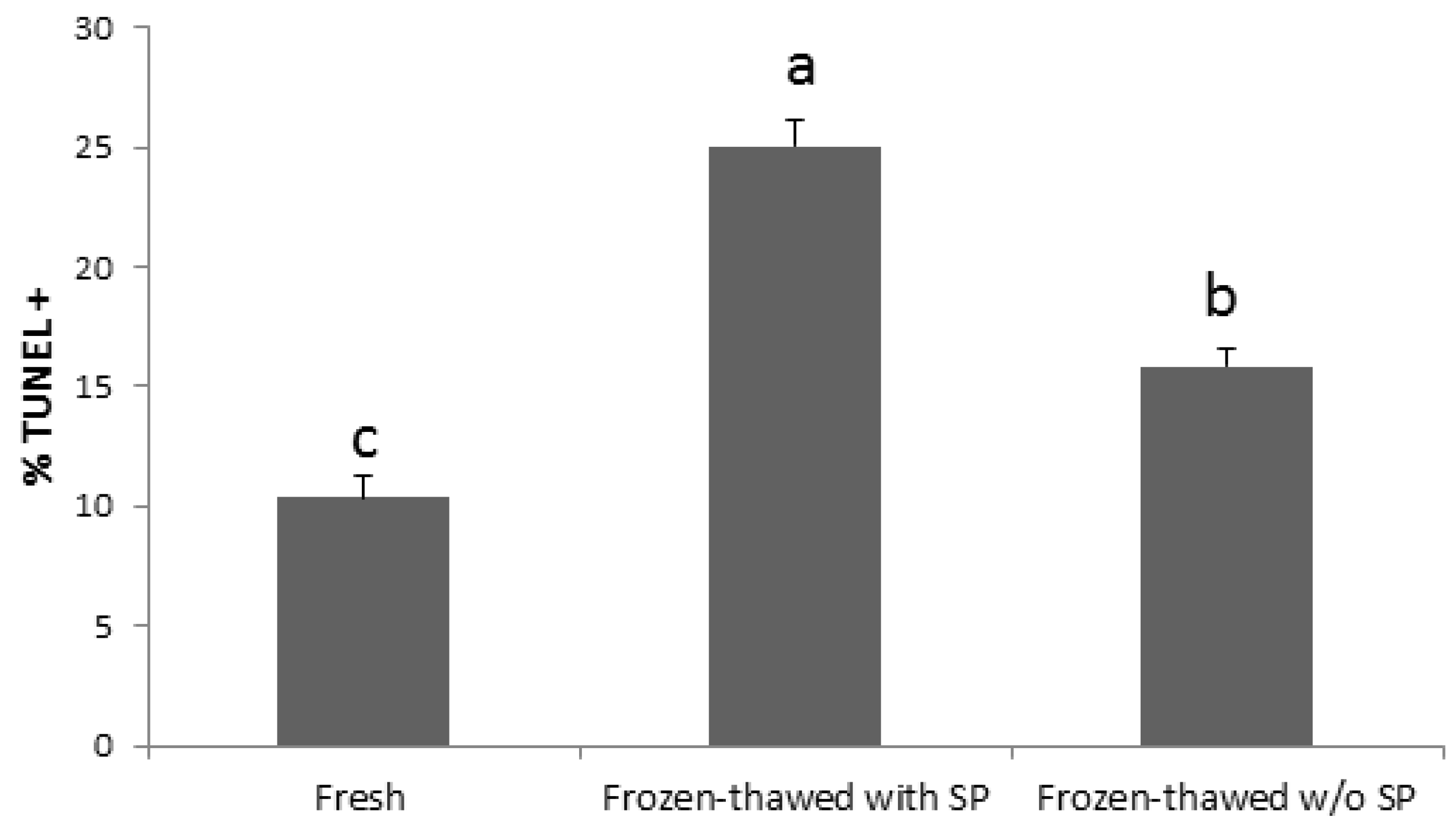
4.5. Exosomal Proteins and Sperm Maturation
4.6. Lipoproteins and Fertility Expression
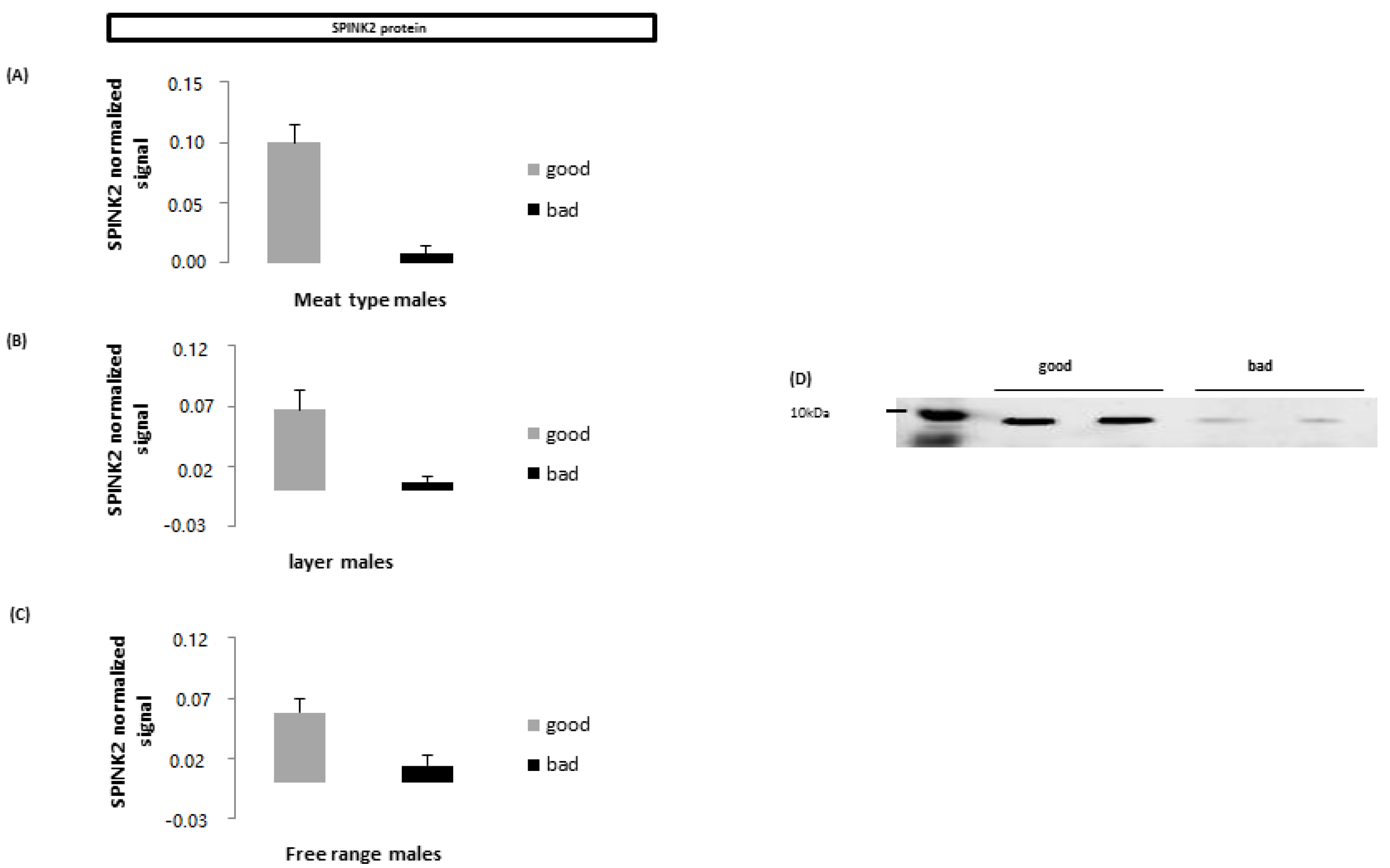
4.7. Specie-Specific Proteins
5. Implications of Seminal Plasma on Assisted Reproductive Technologies
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Druart, X.; de Graaf, S. Seminal plasma proteome and sperm fertility. Anim. Reprod. Sci. 2018, 194, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sridarhn, T.B.; Rao, K.A. Role of seminal plasma proteins in effective zygote formation-A success road to pregnancy. Protein Pept. Lett. 2019, 26, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Blesbois, E.; Brillard, J.P. Specific features of in vivo and in vitro sperm storage in birds. Animal 2007, 1, 1472–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago-Moreno, J.; Esteso, M.C.; Villaverde-Morcillo, S.; Toledano-Díaz, A.; Castaño, C.; Velázquez, R.; López-Sebastian, A.; López-Goya, A.; Gimeno Martínez, J. Recent advances in bird sperm morphometric analysis and its role in male gamete characterization and reproduction technologies. Asian J. Androl. 2016, 18, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Blesbois, E. Bird Reproduction Overview. In Encyclopedia of Reproduction; Skinner, M.K., Ed.; Academic Press: Cambridge, MA, USA, 2018; Volume 6, pp. 579–585. [Google Scholar]
- Finseth, F.R.; Iacovelli, S.R.; Harrison, R.G.; Adkins-Regan, E.K. A nonsemen copulatory fluid influences the outcome of sperm competition in Japanese quail. J. Evol. Biol. 2013, 26, 1875–1889. [Google Scholar] [CrossRef]
- Thélie, A.; Grasseau, I.; Grimaud-Jottreau, I.; Seigneurin, F.; Blesbois, E. Improvements of insemination conditions for successful fertilization in Japanese quail (Coturnix japonica). Theriogenology 2019, 139, 98–105. [Google Scholar] [CrossRef]
- Etches, R.J. The Male. In Reproduction in Poultry; Etches, R.J., Ed.; CAB International: Oxford, UK, 1996; pp. 208–233. [Google Scholar]
- Douard, V.; Hermier, D.; Blesbois, E. Changes in turkey semen lipids during liquid in vitro storage. Biol. Reprod. 2000, 63, 1450–1456. [Google Scholar] [CrossRef]
- Douard, V.; Gassama-Diene, A.; Hermier, D.; Blesbois, E. Activity of Phospholipases A and Lysophospholipase in turkey semen and oviducal fluid. Poult. Sci. 2004, 83, 1385–1393. [Google Scholar] [CrossRef]
- Atikuzzaman, A.; Sanz, L.; Pla, D.; Alvarez-Rodriguez, M.; Ruber, M.; Wright, D.; Calvete, J.J.; Rodriguez-Martinez, H. Selection for higher fertility reflects in the seminal fluid proteome of modern domestic chicken. Comp. Biochem. Physiol. Part D 2017, 21, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Slowinska, M.; Nynca, J.; Arnold, G.J.; Frohlich, T.; Jankowsky, J.; Kozlowski, K.; Mostek, A.; Ciereszko, A. Proteomic identification of turkey seminal plasma proteins. Poult. Sci. 2017, 96, 3422–3435. [Google Scholar] [CrossRef]
- Samour, J.H.; Baggott, G.K.; Williams, G.; Bailey, I.T.; Watson, P.F. Seminal plasma composition in budgerigars (Melopsittacus undulatus). Comp. Biochem. Physiol. A Comp. Physiol. 1986, 84, 735–738. [Google Scholar] [CrossRef]
- Borziak, K.; Alvarez-Fernandez, A.; Karr, T.L.; Pizzari, T.; Dorun, S. The Seminal fluid proteome of the polyandrous red jungle fowl offers insights into the molecular basis of fertility, reproductive ageing and domestication. Sci. Rep. 2016, 6, 35864. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Moreno, J.; Castaño, C.; Toledano-Díaz, A.; Esteso, M.C.; López-Sebastián, A.; Villaverde-Morcillo, S.; Dávila, S.G.; Gil, M.G.; Blesbois, E. Successful chilling of red-legged partridge (Alectoris rufa) sperm for use in artificial insemination. Poult. Sci. 2017, 96, 4068–4074. [Google Scholar] [CrossRef] [PubMed]
- Tawang, A. The Ability of Seminal Plasma to Protect Emu (Dromaius novaeholliandae) Sperm during in Vitro Storage. Ph.D. Thesis, Faculty of Science, School of Animal Biology, The University of Western Australia, Perth, Australia, 2017; p. 186. [Google Scholar]
- Rowe, M.; Whittington, E.; Borziak, K.; Ravinet, M.; Eroukhmanoff, F.; Sætre, G.-P.; Dorus, S. Molecular diversification of the seminal fluid proteome in a recently diverged passerine species pair. Mol. Biol. Evol. 2019, 37, 488–506. [Google Scholar] [CrossRef] [PubMed]
- Marzoni, M.; Castillo, A.; Sagona, S.; Rocchiccioli, S.; Romboli, I.; Felicioli, A. A proteomic approach to identify seminal plasma proteins in roosters (Gallus gallus domesticus). Anim. Reprod. Sci. 2013, 140, 216–223. [Google Scholar] [CrossRef]
- Labas, V.; Grasseau, I.; Cahier, K.; Gargaros, A.; Harichaux, G.; Teixeira-Gomes, A.P.; Alves, S.; Bourin, M.; Gérard, N.; Blesbois, E. Qualitative and quantitative proteomic approaches to phenotyping chicken semen. J. Proteom. 2015, 112, 313–335. [Google Scholar] [CrossRef]
- Soler, L.; Labas, V.; Thélie, A.; Grasseau, I.; Teixeira-Gomes, A.P.; Blesbois, E. Intact cell MALDI-TOF MS on sperm: Towards a molecular test for male fertility diagnosis. Mol. Cell. Proteom. 2016, 15, 1998–2010. [Google Scholar] [CrossRef] [Green Version]
- Slowinska, M.; Hejmej, A.; Bukowska, J.; Liszewska, E.; Bilinska, B.; Hliwa, P.; Kozlowski, K.; Jankowski, J.; Ciereszko, A. Expression and secretion of albumin in male turkey (Meleagris gallopavo) reproductive tract in relation to yellow semen syndrome. Poult. Sci. 2019, 98, 1872–1882. [Google Scholar] [CrossRef]
- Thélie, A.; Rehault-Godbert, S.; Poirier, J.C.; Labas, V.; Fouchecourt, S.; Blesbois, E. The seminal acrosin-inhibitor ClTI1/SPINK2 is a fertility-associated marker in the chicken. Mol. Reprod. Dev. 2019, 86, 762–775. [Google Scholar] [CrossRef]
- Li, Y.; Sun, Y.; Ni, A.; Shi, L.; Wang, P.; Isa, A.M.; Ge, P.; Jiang, L.; Fan, J.; Ma, H.; et al. Seminal plasma protein as an indicator of sperm dysfunction and low sperm motility in chickens. Mol. Cell Proteom. 2020, 19, 1035–1046. [Google Scholar] [CrossRef] [Green Version]
- Blesbois, E.; Hermier, D. Effects of high-density lipoproteins on storage at 4 degrees C of fowl spermatozoa. J. Reprod. Fertil. 1990, 90, 473–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blesbois, E.; de Reviers, M. Effect of different fractions of seminal plasma on the fertilizing ability of fowl spermatozoa stored in vitro. J. Reprod. Fertil. 1992, 95, 263–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iaffaldano, N.; Meluzzi, A. Effect of dialysis on quality characteristics of turkey semen during liquid storage. Theriogenology 2003, 60, 421–427. [Google Scholar] [CrossRef]
- Douard, V.; Hermier, D.; Magistrini, M.; Blesbois, E. Role of seminal plasma in damage to turkey spermatozoa during in vitro storage. Theriogenology 2005, 63, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Blesbois, E. Biological features of the avian male gamete and their application to biotechnology of conservation. J. Poult. Sci. 2012, 49, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Santiago-Moreno, J.; Castaño, C.; Toledano-Díaz, A.; Esteso, M.C.; López-Sebastián, A.; Gañán, N.; Hierro, M.J.; Marchal, F.; Campo, J.L.; Blesbois, E. Characterization of red-legged partridge (Alectoris rufa) sperm: Seasonal changes and influence of genetic purity. Poult. Sci. 2015, 94, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Villaverde-Morillo, S.; Soler, A.J.; Esteso, M.C.; Castaño, C.; Miñano Berna, A.; Gonzalez, F.; Santiago-Moreno, J. Immature and mature sperm morphometry in fresh and frozen-thawed falcon ejaculates. Theriogenology 2017, 98, 94–100. [Google Scholar] [CrossRef]
- Jones, R.C. Evolution of the vertebrate epididymis. J. Reprod. Fertil. Suppl. 1998, 53, 163–181. [Google Scholar]
- Long, J.A. Applied andrology in chickens and turkeys. In Animal Andrology. Theories and Applications; Chenoweth, P.J., Lorton, S.P., Eds.; CABI: Boston, MA, USA, 2014; pp. 197–225. [Google Scholar]
- Clulow, J.; Jones, R.C. Studies of fluid and spermatozoal transport in the extratesticular genital ducts of the Japanese quail. J. Anat. 1988, 157, 1–11. [Google Scholar]
- Howarth, B. Fertilizing ability of cock spermatozoa from the testis, epididymis and vas deferens following intramagnal insemination. Biol. Reprod. 1983, 28, 586–590. [Google Scholar] [CrossRef] [Green Version]
- Morris, S.A.; Howarth, B., Jr.; Crim, J.W.; Rodriguez de Cordoba, A.; Esponda, P.; Bedford, K.M. Specificity of sperm-binding Wolffian duct proteins in the rooster and their persistence on spermatozoa in the female host glands. J. Exp. Zool. 1987, 242, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Glover, T.D.; Nicander, L. Some aspects of structure and function in te mammalian epididymis. J. Reprod. Fertil. Suppl. 1971, 13, 39–50. [Google Scholar]
- Burrows, W.H.; Quinn, J.P. The collection of spermatozoa from the domestic fowl and turkey. Poult. Sci. 1937, 16, 19–24. [Google Scholar] [CrossRef]
- Fujihara, N. Accessory reproductive fluids and organs in male domestic birds. World’s Poult. Sci. 1992, 48, 29–56. [Google Scholar] [CrossRef]
- Biswas, A.; Ranganatha, O.S.; Mohan, J. The effect of different foam concentrations on sperm motility in japanese quail. Vet. Med. Int. 2010, 2010, 564921. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Sastry, K.V.H.; Pandey, N.K.; Singh, K.B.; Malecki, I.A.; Farooq, U.; Mohan, J.; Saxena, V.K.; Moudgal, R.P. The role of the male cloacal gland in reproductive success in Japanese quail (Coturnix japonica). Reprod. Fertil. Dev. 2012, 24, 405–409. [Google Scholar] [CrossRef]
- Tan, N.S. The training of drakes for semen collection. Ann. Zootech. 1980, 29, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Bakst, M.R. Oviductal sperm storage in poultry: A review. Reprod. Fertil. Dev. 1993, 5, 595–599. [Google Scholar] [CrossRef]
- Birkhead, T.R.; Pizarri, T. Sperm competition and fertilization success. In Biology of Breeding Poultry. Poultry Science Symposium Series; Hocking, P.M., Ed.; CAB International: Oxfordshire, UK, 2009; Volume 29, pp. 133–150. [Google Scholar]
- Santiago-Moreno, J.; Castaño, C.; Toledano-Díaz, A.; Esteso, M.C.; López-Sebastián, A.; Dávila, S.G.; Campo, J.L. Role of sperm velocity variables associated with poultry breed in ‘last male precedence’. Reprod. Dom. Anim. 2014, 49, 134–139. [Google Scholar] [CrossRef]
- Pizzari, T.; Worley, K.; Burke, T.; Froman, D.P. Sperm competition dynamics: Ejaculate fertilising efficiency changes differentially with time. BMC Evol. Biol. 2008, 8, 332. [Google Scholar] [CrossRef] [Green Version]
- Froman, D. Deduction of a model for sperm storage in the oviduct of the domestic fowl (Gallus domesticus). Biol. Reprod. 2003, 69, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Mann, T.; Lutwak-Mann, C. Biochemistry of seminal plasma and male accessory fluids;application to andrological problems. In Male Reproductive Function and Semen; Mann, T., Lutwak-Mann, C., Eds.; Springer: Berlin/Heidelberg, Germany; New York, UY, USA, 1981; pp. 269–336. [Google Scholar]
- Lake, P.E.; Wishart, G.J. Comparative physiology of turkey and fowl semen. In Reproductive Biology of Poultry; Cunningham, F.J., Lake, P.E., Hewit, D., Eds.; British Poultry Science: Harlow, UK, 1984; pp. 151–160. [Google Scholar]
- Karaka, A.G.; Parker, H.M.; Yeatman, J.B.; McDaniel, C.D. The effects of heat stress and sperm quality classification on broiler breeder male fertility and semen ion concentration. Br. J. Poult. Sci. 2002, 43, 621–628. [Google Scholar]
- Ahluwalia, B.S.; Graham, E.F. Free amino acid in the semen of the fowl and the turkey. J. Reprod. Fertil. 1966, 12, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Wishart, G.J. Maintenance of ATP concentration in and of fertilizing ability of fowl and turkey spermatozoa in vitro. J. Reprod. Fertil. 1982, 66, 457–462. [Google Scholar] [CrossRef] [Green Version]
- McLean, D.J.; Jones, L.G., Jr.; Froman, D.P. Reduced glucose transport in sperm from roosters (Gallus domesticus) with heritable subfertility. Biol. Reprod. 1997, 57, 791–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushiyama, A.; Priyadarshana, C.; Setiawan, R.; Miyazaki, H.; Ishikawa, N.; Tajima, A.; Asano, A. Membrane raft-mediated regulation of glucose signaling pathway leading to acrosome reaction in chicken sperm. Biol. Reprod. 2019, 100, 1482–1491. [Google Scholar] [CrossRef]
- Zaniboni, L.; Cerolini, S. Liquid storage of turkey semen: Changes in quality parameters, lipid composition and susceptibility to induced in vitro peroxidation in control, n-3 fatty acids and alpha-tocopherol rich spermatozoa. Anim. Reprod. Sci. 2009, 112, 51–65. [Google Scholar] [CrossRef]
- Kelso, K.A.; Cerolini, S.; Noble, R.C.; Sparks, N.H.; Speake, B.K. Lipid and antioxidant changes in semen of broiler fowl from 25 to 60 weeks of age. J. Reprod. Fertil. 1996, 106, 201–206. [Google Scholar] [CrossRef]
- Cerolini, S.; Surai, P.; Maldjian, A.; Gliozzi, T.; Noble, R. Lipid composition of semen in different fowl breeders. Poult. Avian Biol. Rev. 1997, 8, 141–148. [Google Scholar]
- Burt, C.T.; Ribolow, H. Glycerol phosphorylcholine (GPC) and serine ethanolamine phosphodiester (SEP): Evolutionary mirrored metabolites and their potential metabolic roles. Comp. Biochem. Physiol. Biochem. Mol. Biol. 1994, 108, 11–20. [Google Scholar] [CrossRef]
- Blesbois, E.; Lessire, M.; Grasseau, I.; Hallouis, J.M.; Hermier, D. Effect of dietary fat on the fatty acid composition and on fertilizing ability of fowl semen. Biol. Reprod. 1997, 56, 1216–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blesbois, E.; Douard, V.; Germain, M.; Boniface, P.; Pellet, F. Effects of n-3 polyunsaturated dietary supplementation on the reproductive capacity of male turkeys. Theriogenology 2004, 61, 537–549. [Google Scholar] [CrossRef]
- Cecil, H.; Bakst, M.R. Testosterone concentrations in blood and seminal plasma of turkeys classified as high or low semen producers. Poult. Sci. 1988, 67, 1461–1464. [Google Scholar] [CrossRef] [PubMed]
- Zeman, M.; Kosutzky, J.; Bobakova, E. Testosterone concentration in the seminal plasma of cocks. Br. Pout. Sci. 1986, 27, 261–266. [Google Scholar] [CrossRef]
- Lelono, A.; Riedstra, B.; Groothuis, T. Ejaculate testosterone levels affect maternal investment in red junglefowl (Gallus gallus gallus). Sci. Rep. 2019, 9, 12126. [Google Scholar] [CrossRef]
- Santiago-Moreno, J.; Bernal, B.; Castaño, C.; Toledano-Díaz, A.; Gil, G.; Woelders, H.; Blesbois, E. Seminal Plasma amino acid profile in different breeds of chicken: Role of seminal plasma on sperm cryoresistance. PLoS ONE 2019, 14, e0209910. [Google Scholar] [CrossRef] [Green Version]
- Lake, P.E.; McIndoe, W.M. The glutamic acid and creatine content of cock seminal plasma. Biochem. J. 1959, 71, 303–306. [Google Scholar] [CrossRef] [Green Version]
- El Jack, M.H.; Lake, P.E. The distribution of the principal inorganic ions in semen from the vas deferens of the domestic fowl and the content of carbon dioxide in the seminal plasma. Q. J. Exp. Physiol. Cogn. Med. Sci. 1969, 54, 332–340. [Google Scholar] [CrossRef] [Green Version]
- Froman, D.P.; Kirby, J.D. Sperm mobility: Phenotype in roosters (Gallus domesticus) determines by mitochondrial function. Biol. Reprod. 2005, 72, 562–567. [Google Scholar] [CrossRef] [Green Version]
- Al-Aghbari, A.; Engel, H.N., Jr.; Froman, D.P. Analyses of seminal plasma from roosters carrying the Sd (Sperm degeneration) allele. Biol. Reprod. 1992, 47, 1059–1063. [Google Scholar] [CrossRef]
- Lake, P.E.; Hatton, M. Free amino acids in the vas deferens, semen, transparent fluid and blood plasma of the domestic rooster Gallus domesticus. J. Reprod. Fertil. 1968, 15, 139–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, G.C., Jr.; Sweeney, M.J. Changes in the proteins concentration of chicken seminal plasma after rapid freeze-thaw. Cryobiology 1971, 7, 209–215. [Google Scholar] [CrossRef]
- Amen, M.H.M.; Al-Daraji, H.J. Effect of dietary zinc supplementation on some seminal plasma characteristics of broiler breeder males. Int. J. Poult. Sci. 2011, 10, 814–818. [Google Scholar]
- Thurston, R.J.; Hess, R.A.; Froman, D.P.; Biellier, H.V. Elevated seminal plasma protein: A characteristic of yellow turkey semen. Poult. Sci. 1982, 61, 1905–1911. [Google Scholar] [CrossRef]
- Blesbois, E.; Caffin, J.P. “Serum like” albumin of fowl seminal plasma and effects of albumin on fowl spermatozoa stored at 4 degrees C. Br. Poult. Sci. 1992, 33, 663–670. [Google Scholar] [CrossRef]
- Thurston, R.J.; Korn, N.; Froman, D.P.; Bodine, A.B. Proteolytic enzymes in seminal plasma of domestic turkey. Biol. Reprod. 1993, 48, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Kannan, Y. Glycosidases in fowl semen. J. Reprod. Fertil. 1974, 40, 227–228. [Google Scholar] [CrossRef] [Green Version]
- Droba, B.; Dzugan, M. Acid alpha-D-mannosidase from turkey seminal plasma. Folia Biol. Krakov 1993, 41, 49–54. [Google Scholar]
- Dumitru, I.F.; Dinischiotu, A. Cock seminal plasma acid phosphatase: Active site directed inactivation, crystallization, and in vitro denaturation—Renaturation studies. Int. J. Biochem. 1994, 26, 497–503. [Google Scholar] [CrossRef]
- Surai, P.F.; Blesbois, E.; Grasseau, I.; Chalah, T.; Brillard, J.P.; Wishart, G.J.; Cerolini, S.; Sparks, N.H.C. Fatty acid composition, glutathione peroxidase and superoxide dismutase activity and total antioxidant activity of avian semen. Comp. Biochem. Physiol. 1998, 120, 527–533. [Google Scholar] [CrossRef]
- Khan, R.U.; Rahman, Z.U.; Javed, I.; Muhammad, F. Effects of vitamins, probiotics and protein on semen traits in post-molt male broiler breeders. Anim. Reprod. Sci. 2012, 135, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Blesbois, E.; Mauger, I. Effects of ovalbumin on the motility and fertilising ability of fowl spermatozoa stored for 24H at 4 degrees C. Br. Poult. Sci. 1987, 28, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Elzanaty, S.; Erenpriess, J.; Becker, C. Seminal plasma albumin origin and relation to the male reproductive parameters. Andrologia 2007, 39, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Arroteia, K.F.; Barbieri, M.F.; Souza, G.H.; Tanaka, M.; Eberlin, N.; Hyslop, S.; Alvares, L.E.; Pereira, A. Albumin is synthesized in epididymis and aggregates in a high molecular mass glycoprotein complex involved in sperm-egg fertilization. PLoS ONE 2014, 9, e103566. [Google Scholar] [CrossRef] [Green Version]
- Girouard, J.; Frenette, G.; Sullivan, R. Compartmentalization of proteins in epididymosomes coordinates the association of epididymal proteins with different functionnal structures of bovine spermatozoa. Biol. Reprod. 2009, 80, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.C.; Morgan, C.P.; Leu, N.A.; Shetty, A.; Cisse, Y.; Nugent, B.M.; Morrison, K.E.; Jasarevic, E.; Huang, W.; Kanyuch, N.; et al. Reproductive tract extracellular vesicles are sufficient to transmit intergenerational stress and program neurodevelopment. Nat. Commun. 2020, 11, 1499. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Rodriguez, M.; Ntzouni, M.; Wright, D.; Khan, K.I.; Lopez-Bejar, M.; Martinez, C.A.; Rodriguez-Martinez, H. Chicken seminal fluid lacks CD9 and CD44-bearing extracellular vesicles. Reprod. Domest. Anim. 2020, 55, 293–300. [Google Scholar] [CrossRef]
- Ansari, M.S.; Rakha, B.A.; Akhter, S.; Blesbois, E.; Santiago-Moreno, J. Effect of cryopreservation on lipid peroxidation, antioxidant potential, chromatin integrity, and mitochondrial activity of Indian red jungle fowl (Gallus gallus murghi) semen. Biopreserv. Biobank. 2019, 17, 288–295. [Google Scholar] [CrossRef]
- Blesbois, E.; Lessire, M.; Hermier, D. Effect of cryopreservation and diet on lipids of fowl sperm and fertility. Poult. Avian Biol. Rev. 1997, 8, 149–154. [Google Scholar]
- Blesbois, E.; Mauger, I. Zinc content of fowl seminal plasma and its effects on spermatozoa after storage at 4 degrees C. Br. Poult. Sci. 1989, 39, 677–685. [Google Scholar] [CrossRef]
- Thananurak, P.; Chuaychy-noo, N.; Thélie, A.; Phasuk, Y.; Vongpralub, T.; Blesbois, E. Different concentrations of Cysteamine, Ergothioneine, and Serine modulate quality and fertilizing ability of cryopreserved chicken sperm. Poult. Sci. 2020, 99, 1185–1198. [Google Scholar] [CrossRef] [PubMed]
- Withers, L.A.; King, P.J. Proline: A novel cryoprotectant for the freeze preservation of cultured cells of Zea mays L. Plant Physiol. (Bethesda) 1979, 64, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, J.F.; Crowe, J.H. The mechanism of cryoprotection of protein by solutes. Cryobiology 1988, 25, 244–255. [Google Scholar] [CrossRef]
- Anchordoguy, T.; Carpenter, J.; Looms, S.; Crowe, J. Mechanisms of interaction of amino acids with phospholipids bilayers during freezing. Biochim. Biophys. Acta 1988, 946, 299–306. [Google Scholar] [CrossRef]
- Heber, U.; Tyankova, L.; Santarius, K.A. Stabilization and inactivation of biological membranes during freezing in the presence of amino acids. Biochim. Biophys. Acta 1971, 241, 578–592. [Google Scholar] [CrossRef]
- Partyka, A.; Rodak, O.; Bajzert, J.; Kochan, J.; Nizanski, W. The effect of L-Carnitine, Hypotaurine and taurine supplementation on the quality of cryopreserved chicken semen. BioMed. Res. Int. 2017, 2017, 7279341. [Google Scholar] [CrossRef] [Green Version]
- Bernal, B.; Iglesias-Cabeza, N.; Sánchez-Rivera, U.; Toledano-Díaz, A.; Castaño, C.; Pérez-Cerezales, S.; Gutiérrez-Adán, A.; López-Sebastián, A.; García-Casado, P.; Gil, M.G.; et al. Effect of supplementation of valine to chicken extender on sperm cryoresistance and post-thaw fertilization capacity. Poult. Sci. 2020. under review. [Google Scholar]
- Sanchez-Partida, L.; Maxwell, W.; Paleg, L.; Setchell, B. Proline and glycine betaine in cryoprotective diluents for ram spermatozoa. Reprod. Fertil. Dev. 1992, 4, 113–118. [Google Scholar] [CrossRef]
- Renard, P.; Grizard, G.; Griveau, J.F.; Sion, B.; Boucher, D.; Le Lannou, D. Improvement of motility and fertilization potential or post-thaw human sperm using glutamine. Cryobiology 1996, 33, 311–319. [Google Scholar] [CrossRef]
- Trimeche, A.; Yvon, J.; Vidament, M.; Palmer, E.; Magistrini, M. Effects of glutamine, proline, histidine and betaine on post-thaw motility of stallion spermatozoa. Theriogenology 1999, 52, 181–191. [Google Scholar] [CrossRef]
- Kundu, C.; Das, K.; Majumder, G. Effect of amino acids on goat cauda epididymal sperm cryopreservation using a chemically defined model system. Cryobiology 2001, 42, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Khiabania, A.B.; Moghaddamb, G.; Kiab, H.D. Effects of adding different levels of Glutamine to modified Beltsville extender on the survival of frozen rooster semen. Anim. Reprod. Sci. 2017, 184, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Ogretmen, F.; Inanan, B.E.; Kutluyer, F.; Kayım, M. Effect of semen extender supplementation with cysteine on post-thaw sperm quality, DNA damage, and fertilizing ability in the common carp (Cyprinus carpio). Theriogenology 2015, 83, 1548–1552. [Google Scholar] [CrossRef] [PubMed]
- Amidi, F.; Pazhohan, A.; Nashtaei, M.S.; Khodarahmian, M.; Nekoonam, S. The role of antioxidants in sperm freezing: A review. Cell Tissue Bank. 2016, 17, 745–756. [Google Scholar] [CrossRef]
- Aitken, R.J.; Buckingham, D.W.; Brindle, J.; Gomez, E.; Baker, H.W.; Irvine, D.S. Analysis of sperm movement in relation to the oxidative stress created by leukocytes in washed sperm preparations and seminal plasma. Hum. Reprod. 1995, 10, 2061–2071. [Google Scholar] [CrossRef]
- Bilodeau, J.F.; Chatterjee, S.; Sirard, M.A.; Gagnon, C. Levels of antioxidant defenses are decreased in bovine spermatozoa after a cycle of freezing and thawing. Mol. Reprod. Dev. 2000, 55, 282–288. [Google Scholar] [CrossRef]
- Millán de la Blanca, M.G.; Martínez-Nevado, E.; Castaño, C.; García, J.; Toledano-Díaz, A.; Esteso, M.C.; Bóveda, P.; Martínez-Fresneda, L.; López-Sebastián, A.; Santiago-Moreno, J. Sperm cryopreservation in American flamingo (Phoenicopterus ruber): Influence of cryoprotectants and seminal plasma removal. Animals 2020. under review. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Lipid | Chicken | Turkey |
| Total lipid (µg mL−1 seminal plasma) | 623.4 | 124 |
| Phospholipid (% of total lipids) | 31.3 | 50.7 |
| Free cholesterol (% of total lipids) | 18.0 | 31.1 |
| Triglycerides (% of total lipids) | 21.1 | 6.8 |
| Free fatty acid (% of total lipids) | 13.7 | 2.5 |
| Cholesterol ester (% of total lipids) | 15.9 | 12.3 |
| Phospholipids | ||
| Phosphatidyl choline (% of total phospholipids) | 15.5 | 4.6 |
| Phosphatidyl ethanolamine (% of total phospholipids) | 43.6 | 31.5 |
| Sphingomyelin (% of total phospholipids) | 13.4 | 20.3 |
| Lipid | Chicken | Turkey |
| Total lipid (µg mL−1 seminal plasma) | 623.4 | 124 |
| Phospholipid (% of total lipids) | 31.3 | 50.7 |
| Free cholesterol (% of total lipids) | 18.0 | 31.1 |
| Triglycerides (% of total lipids) | 21.1 | 6.8 |
| Free fatty acid (% of total lipids) | 13.7 | 2.5 |
| Cholesterol ester (% of total lipids) | 15.9 | 12.3 |
| Phospholipids | ||
| Phosphatidyl choline (% of total phospholipids) | 15.5 | 4.6 |
| Phosphatidyl ethanolamine (% of total phospholipids) | 43.6 | 31.5 |
| Sphingomyelin (% of total phospholipids) | 13.4 | 20.3 |
| Glu | Ala | Ser | Val | Gly | Thr | Pro | Asp | Leu | Arg | Ile | Phe | His | Cys | Met | Tyr | Lys | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | 44.9 | 1.3 | 1.2 | 0.9 | 0.9 | 0.7 | 0.6 | 0.5 | 0.5 | 0.5 | 0.3 | 0.3 | 0.3 | 0.2 | 0.2 | 0.2 | 0.2 |
| SD | 8.3 | 0.3 | 0.3 | 0.2 | 0.2 | 0.2 | 0.1 | 0.1 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| C:VB | * | * | * | * | |||||||||||||
| C:DNA | * | * | * | * | * | * | * | * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santiago-Moreno, J.; Blesbois, E. Functional Aspects of Seminal Plasma in Bird Reproduction. Int. J. Mol. Sci. 2020, 21, 5664. https://doi.org/10.3390/ijms21165664
Santiago-Moreno J, Blesbois E. Functional Aspects of Seminal Plasma in Bird Reproduction. International Journal of Molecular Sciences. 2020; 21(16):5664. https://doi.org/10.3390/ijms21165664
Chicago/Turabian StyleSantiago-Moreno, Julian, and Elisabeth Blesbois. 2020. "Functional Aspects of Seminal Plasma in Bird Reproduction" International Journal of Molecular Sciences 21, no. 16: 5664. https://doi.org/10.3390/ijms21165664
APA StyleSantiago-Moreno, J., & Blesbois, E. (2020). Functional Aspects of Seminal Plasma in Bird Reproduction. International Journal of Molecular Sciences, 21(16), 5664. https://doi.org/10.3390/ijms21165664