Osteoclast Multinucleation: Review of Current Literature


Abstract
:1. Introduction
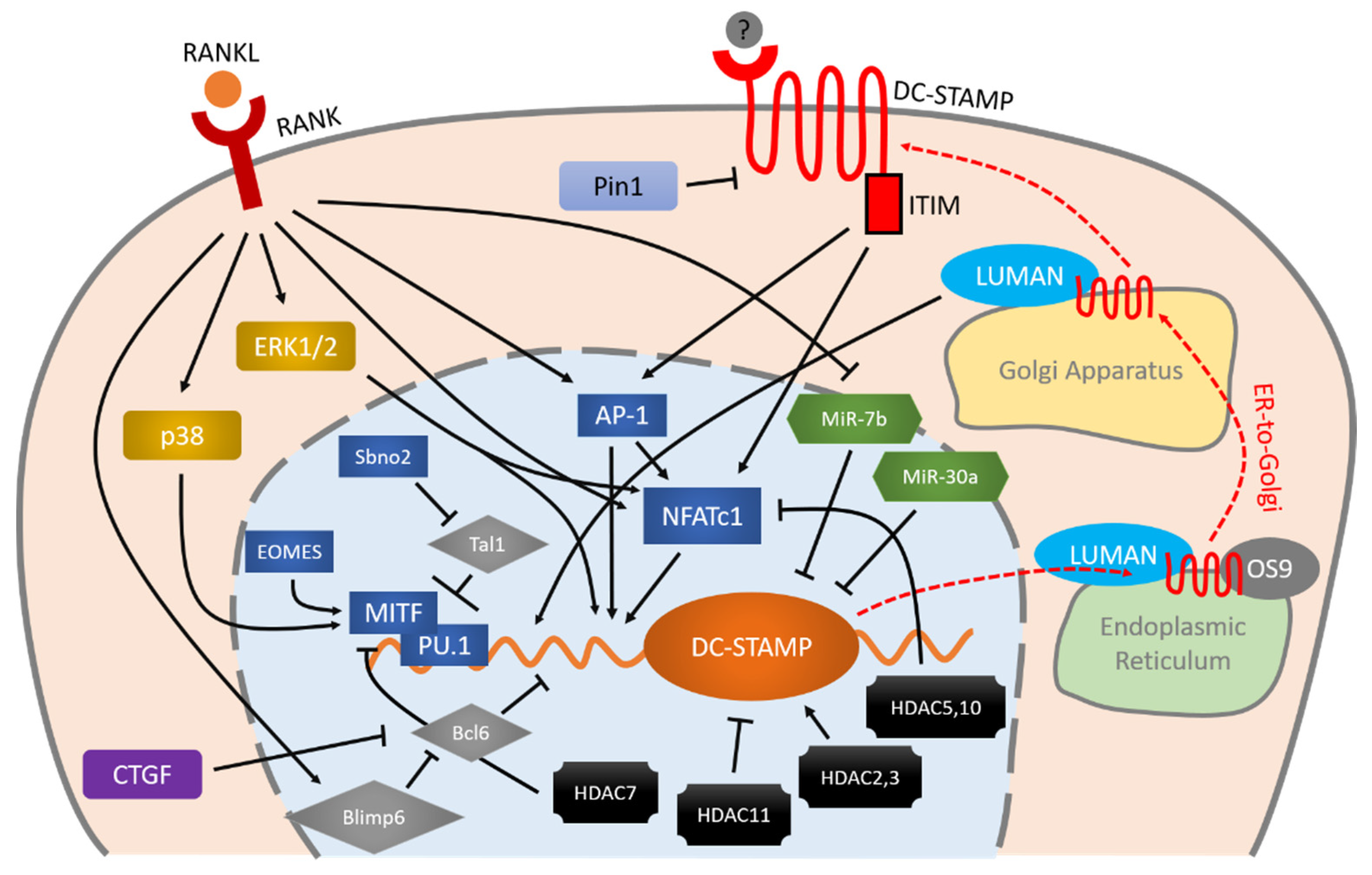
2. DC-STAMP
2.1. DC-STAMP Is a Transmembrane Protein
2.2. DC-STAMP Works in a Receptor-Ligand Machinery
2.3. The Expression and Cellular Distribution of DC-STAMP Change under RANKL Signaling
2.4. C-Fos and NFATc1 Modulate the Gene Expression of DC-STAMP
2.5. Extracellular Signal-Regulated Kinases (ERKs) Regulate DC-STAMP
2.6. Bcl6 Is a Transcriptional Repressor of DC-STAMP
2.7. MITF-PU.1 Complex and Tal1 Work Reciprocally to Regulate DC-STAMP Expression
2.8. Several HDACs Are Involved in DC-STAMP Regulation
2.9. Proteins Interact with DC-STAMP
2.9.1. Pin1 Directly Suppresses DC-STAMP
2.9.2. Luman, DC-STAMP and OS-9 Work as a Complex with Regard to Intracellular Localization and Activities
2.10. Micro RNAs Directly Suppress the mRNA of DC-STAMP
2.11. DC-STAMP May Play a Role in Paget’s Disease of Bone
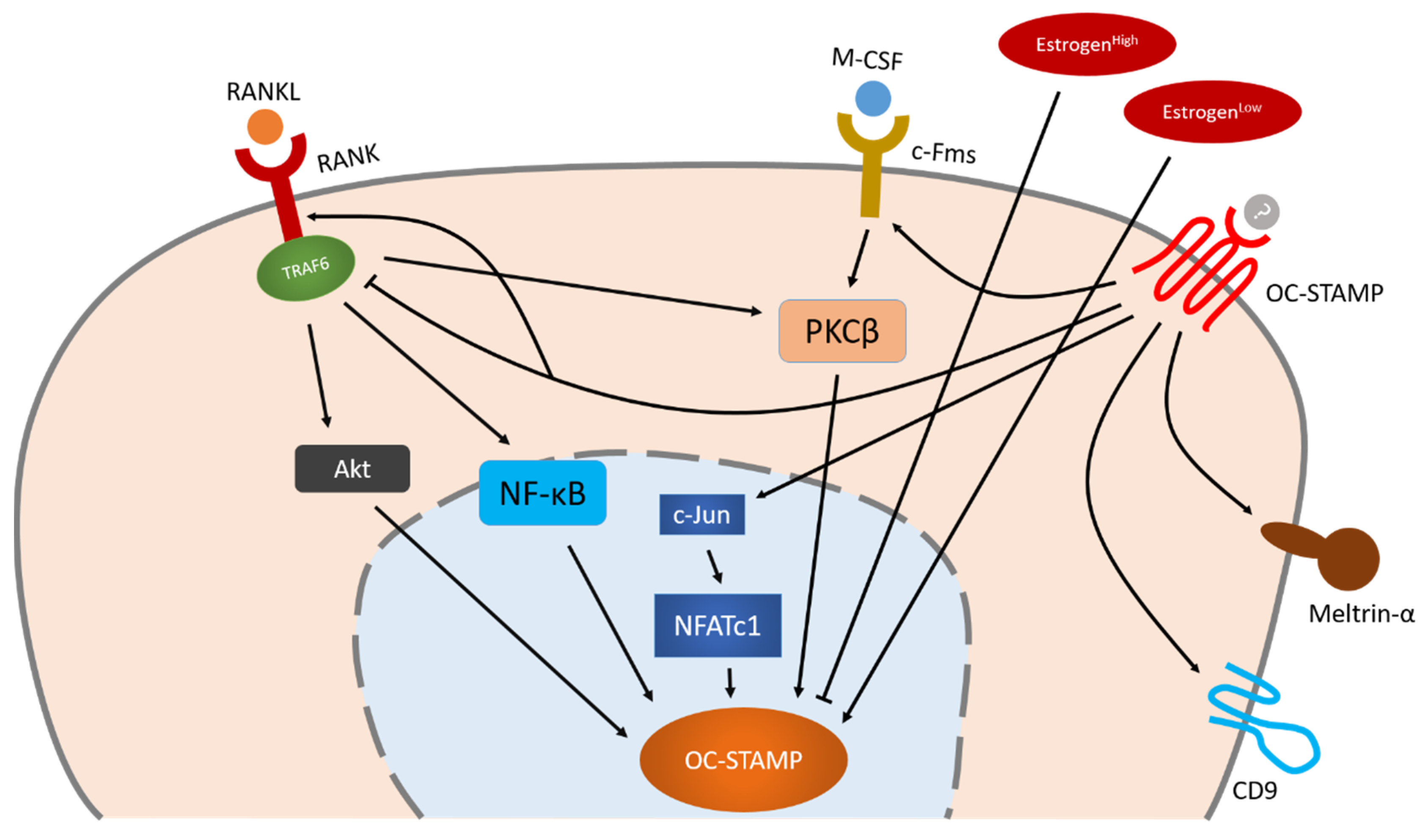
3. OC-STAMP
3.1. Protein Structure and Cellular Distribution of OC-STAMP Are Similar but not Identical to Those of DC-STAMP
3.2. Osteoclast Multinucleation Is Completely Inhibited in OC-STAMP-Deficient Mice, Which nevertheless Show Normal Growth and Skeletal Systems
3.3. Regulation of OC-STAMP Expression during Osteoclastogenesis
3.4. OC-STAMP and DC-STAMP Work Cooperatively
4. ATP6v0d2
4.1. ATP6v0d2 Is not Only a Proton Pump in Osteoclasts
4.2. How Is ATP6v0d2 Regulated and Involved in Osteoclast Multinucleation?
4.3. P-ATPase α1 and NHE10 also Regulate Osteoclast Multinucleation
5. CD47-MFR
6. Cadherins
7. Syncytins
8. ITAM-Bearing Immunoreceptors
8.1. OSCAR-FcRγ
8.2. DAP12
8.3. Siglec-15
8.4. Siglec-15/DAP Association
8.5. Treatment of Osteoporosis Targeting Siglec-15
9. NETRIN-1, Flrt2 and Unc5b axis
10. Transglutaminases
11. Purinergic Receptors
12. TRAIL
13. Recent Pre-Clinical Studies

{kind=link}
{kind=link}
| Author and Year of Publication | Compound Used | In-Vitro Cell Types | In-Vivo Model | Affected Molecules |
|---|---|---|---|---|
| Sun 2020 [183] | Pregnenolone | Mouse bone marrow macrophages | LPS-induced calvarial osteolysis model of mice | MAPK, NF-κB, c-Fos, NFATc1, Cathepsin K, Trap, DC-STAMP, ATP6v0d2 |
| Choi 2019 [184] | Pisidium coreanum | Mouse bone marrow macrophages | OVX mice treated with Pisidium coreanum | NFATc1, p65, integrin αvβ3, DC-STAMP, OC-STAMP, Atp6v0d2, FAK, CD44, and MFR |
| Takagi 2017 [187] | Sulforaphane | Mouse bone marrow cells RAW264.7 | - | OSCAR, NFATc1, Trap, Cathepsin K, DC-STAMP, OC-STAMP |
| Choi 2017 [188] | Barley seedling extracts | Mouse bone marrow cells | - | IκB, c-Fos, NFATc1, DC-STAMP |
| Zeng 2016 [189] | Aconine | RAW264.7 | - | NF-κB, NFATc1, DC-STAMP |
| Chai 2017 [190] | Melphalan (increased multinucleation) | Mouse bone marrow cells RAW264.7 | Mice treated with melphalan | Mitf, DC-STAMP, OC-STAMP (all upregulated) |
| Arioka 2017 [191] | Inorganic phosphate | RAW-D cells | - | c-Fos, NFATc1, DC-STAMP |
| Tsukamoto 2016 [192] | Elcatonin | Mouse bone marrow cells | Mice tail suspension model (microgravity) treated with elcatonin | NFATc1, cathepsin K, ATP6v0d2 |
| Dou 2016 [193] | Cyanidin (low dose promotes osteoclastogenesis while high dose inhibits it) | Mouse bone marrow monocytes RAW264.7 | - | c-Fos, NFATc1, Dual effects on Mitf, CD9, CD47, ATP6v0d2, DC-STAMP, OC-STAMP, OSCAR |
| Kuriya 2020 [194] | Tea extract (epigallocatechin gallate) | RAW264.7 | - | DC-STAMP |
| Kanzaki 2017 [195] | PGDHC (promotes osteoclast multinucleation) | RAW264.7 Mouse bone marrow cells and peritoneal macrophages | Mice given calvarial injections of PGDHC | Myh9, Rac1, DC-STAMP (upregulated) |
| Ausk 2017 [195] | Botulinum toxin A causing muscle paralysis | Mouse bone marrow cells | Mice injected with botulinum toxin A in right calf muscle | TNF-α, IL-1, DC-STAMP, OC-STAMP |
| Fujita 2012 [197] | Vitamin E (α-tocopherol, increased osteoclast multinucleation) | Mouse bone marrow cells from Ttpa–/– and WT mice | WT mice treated with α-tocopherol | p38, Mitf, DC-STAMP (all upregulated) |
14. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TNF-α | Tumor necrosis factor α |
| OVX | Ovariectomy |
| MEF2 | Myocyte enhancer factor 2 |
| Adrm1 | Adhesion-regulating molecule 1 protein |
| METTL3 | N6-adenosine-methyltransferase 70 kDa subunit |
| YTHDF2 | YTH-Domain Family Member 2 |
| MFR/SHPS-1/SIRPα | Macrophage fusion receptor/Src homology 2 domain-containing protein tyrosine phosphatase substrate-1/Signal-regulatory protein alpha |
| SHP-1 | Src homology region 2-containing protein tyrosine phosphatase-1 |
| Myh9 | Non-muscle myosin IIA |
| Mac-1 | Integrin macrophage antigen 1 |
| ASCT2 | System alanine-serine-cysteine transporter 2 |
| DAP12 | DNAX associated protein 12kD size |
| FcRγ | FcεR1 gamma chain |
| TREM2 | Triggering receptor expressed on myeloid cells 2 |
| Siglec-15 | Sialic acid-binding Ig-like lectin 15 |
| SYK | Spleen tyrosine kinase |
| Unc5b | Uncoordinated-5b receptor |
| PIR-B | Paired immunoglobulin-like receptor |
| Flrt2 | Fibronectin leucine-rich transmembrane protein 2 |
| TG | Transglutaminase |
| FGBC | Foreign giant body cell |
| SNP | Single nucleotide polymorphism |
| MEP50 | Methylosome protein 50 |
| PRMT5 | Protein arginine methyltransferase 5 |
| TRAIL | Human TNF-related apoptosis-inducing ligand |
References
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Jansen, I.D.C.; Vermeer, J.A.F.; Bloemen, V.; Stap, J.; Everts, V. Osteoclast fusion and fission. Calcif. Tissue Int. 2012, 90, 515–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takegahara, N.; Kim, N.; Mizuno, H.; Sakaue, S.A.; Miyawaki, A.; Tomura, M.; Kanagawa, O.; Ishii, M.; Choi, Y. Involvement of receptor activator of nuclear factor-kappaB ligand (RANKL)-induced incomplete cytokinesis in the polyploidization of osteoclasts. J. Biol. Chem. 2016, 291, 3439–3454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, T. The dendritic cell-specific transmembrane protein DC-STAMP is essential for osteoclast fusion and osteoclast bone-resorbing activity. Mod. Rheumatol. 2006, 16, 341–342. [Google Scholar] [CrossRef]
- Nabavi, N.; Khandani, A.; Camirand, A.; Harrison, R.E. Effects of microgravity on osteoclast bone resorption and osteoblast cytoskeletal organization and adhesion. Bone 2011, 49, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, S.L. The osteoclast and its unique cytoskeleton. Ann. N. Y. Acad. Sci. 2011, 1240, 14–17. [Google Scholar] [CrossRef]
- Palagano, E.; Menale, C.; Sobacchi, C.; Villa, A. Genetics of Osteopetrosis. Curr. Osteoporos. Rep. 2018, 16, 13–25. [Google Scholar] [CrossRef]
- Rachner, T.D.; Khosla, S.; Hofbauer, L.C. Osteoporosis: Now and the future. Lancet 2011, 377, 1276–1287. [Google Scholar] [CrossRef] [Green Version]
- Tateiwa, D.; Yoshikawa, H.; Kaito, T. Cartilage and Bone Destruction in Arthritis: Pathogenesis and Treatment Strategy: A Literature Review. Cells 2019, 8, 818. [Google Scholar] [CrossRef] [Green Version]
- Ukon, Y.; Makino, T.; Kodama, J.; Tsukazaki, H.; Tateiwa, D.; Yoshikawa, H.; Kaito, T. Molecular-Based Treatment Strategies for Osteoporosis: A Literature Review. Int. J. Mol. Sci. 2019, 20, 2557. [Google Scholar] [CrossRef] [Green Version]
- Furuya, M.; Kikuta, J.; Fujimori, S.; Seno, S.; Maeda, H.; Shirazaki, M.; Uenaka, M.; Mizuno, H.; Iwamoto, Y.; Morimoto, A.; et al. Direct cell-cell contact between mature osteoblasts and osteoclasts dynamically controls their functions in vivo. Nat. Commun. 2018, 9, 300. [Google Scholar] [CrossRef] [PubMed]
- Dou, C.; Ding, N.; Luo, F.; Hou, T.; Cao, Z.; Bai, Y.; Liu, C.; Xu, J.; Dong, S. Drug Delivery: Graphene-Based MicroRNA Transfection Blocks Preosteoclast Fusion to Increase Bone Formation and Vascularization (Adv. Sci. 2/2018). Adv. Sci. 2018, 5, 1700578. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.H.; Schwarz, E.; Li, D.; Xu, Y.; Sheu, T.-R.; Li, J.; de Mesy, B.K.L.; Feng, C.; Wang, B.; Wang, J.-C.; et al. Dendritic cell-specific transmembrane protein (DC-STAMP) regulates osteoclast differentiation via the Ca(2+)/NFATc1 axis. J. Cell. Physiol. 2017, 232, 2538–2549. [Google Scholar] [CrossRef]
- Witwicka, H.; Hwang, S.-Y.; Reyes-Gutiérrez, P.; Jia, H.; Odgren, P.E.; Donahue, L.R.; Birnbaum, M.J.; Odgren, P.R. Studies of OC-STAMP in Osteoclast Fusion: A New Knockout Mouse Model, Rescue of Cell Fusion, and Transmembrane Topology. PLoS ONE 2015, 10, e0128275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, H.; Suzuki, T.; Miyauchi, Y.; Iwasaki, R.; Kobayashi, T.; Sato, Y.; Miyamoto, K.; Hoshi, H.; Hashimoto, K.; Yoshida, S.; et al. Osteoclast stimulatory transmembrane protein and dendritic cell-specific transmembrane protein cooperatively modulate cell-cell fusion to form osteoclasts and foreign body giant cells. J. Bone Miner. Res. 2012, 27, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Birnbaum, M.J.; Mackay, C.A.; Mason-Savas, A.; Thompson, B.; Odgren, P.R. Osteoclast stimulatory transmembrane protein (OC-STAMP), a novel protein induced by RANKL that promotes osteoclast differentiation. J. Cell. Physiol. 2008, 215, 497–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagi, M.; Miyamoto, T.; Sawatani, Y.; Iwamoto, K.; Hosogane, N.; Fujita, N.; Morita, K.; Ninomiya, K.; Suzuki, T.; Miyamoto, K.; et al. DC-STAMP is essential for cell–cell fusion in osteoclasts and foreign body giant cells. J. Exp. Med. 2005, 202, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Hartgers, F.C.; Vissers, J.L.; Looman, M.W.; Van Zoelen, C.; Huffine, C.; Figdor, C.G.; Adema, G.J. DC-STAMP, a novel multimembrane-spanning molecule preferentially expressed by dendritic cells. Eur. J. Immunol. 2000, 30, 3585–3590. [Google Scholar] [CrossRef]
- Eleveld-Trancikova, D.; Triantis, V.; Moulin, V.; Looman, M.W.G.; Wijers, M.; Fransen, J.; Lemckert, A.A.C.; Havenga, M.J.E.; Figdor, C.G.; Janssen, R.A.J.; et al. The dendritic cell-derived protein DC-STAMP is highly conserved and localizes to the endoplasmic reticulum. J. Leukoc. Boil. 2004, 77, 337–343. [Google Scholar] [CrossRef]
- Sawatani, Y.; Miyamoto, T.; Nagai, S.; Maruya, M.; Imai, J.; Miyamoto, K.; Fujita, N.; Ninomiya, K.; Suzuki, T.; Iwasaki, R.; et al. The role of DC-STAMP in maintenance of immune tolerance through regulation of dendritic cell function. Int. Immunol. 2008, 20, 1259–1268. [Google Scholar] [CrossRef]
- Kukita, T.; Wada, N.; Kukita, A.; Kakimoto, T.; Sandra, F.; Toh, K.; Nagata, K.; Iijima, T.; Horiuchi, M.; Matsusaki, H.; et al. RANKL-induced DC-STAMP Is Essential for Osteoclastogenesis. J. Exp. Med. 2004, 200, 941–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, R.; Ninomiya, K.; Miyamoto, K.; Suzuki, T.; Sato, Y.; Kawana, H.; Nakagawa, T.; Suda, T.; Miyamoto, T. Cell fusion in osteoclasts plays a critical role in controlling bone mass and osteoblastic activity. Biochem. Biophys. Res. Commun. 2008, 377, 899–904. [Google Scholar] [CrossRef] [PubMed]
- Hartgers, F.C.; Looman, M.W.; van der Woning, B.; Merkx, G.F.; Figdor, C.G.; Adema, G.J. Genomic organization, chromosomal localization, and 5’ upstream region of the human DC-STAMP gene. Immunogenetics 2001, 53, 145–149. [Google Scholar] [CrossRef]
- Staege, H.; Brauchlin, A.; Schoedon, G.; Schaffner, A. Two novel genes FIND and LIND differentially expressed in deactivated and Listeria-infected human macrophages. Immunogenetics 2001, 53, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Foord, S.M.; Marks, B.; Stolz, M.; Bufflier, E.; Fraser, N.J.; Lee, M.G. The Structure of the Prostaglandin EP4 Receptor Gene and Related Pseudogenes. Genome 1996, 35, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Mummidi, S.; Ahuja, S.S.; McDaniel, B.L.; Ahuja, S.K. The human CC chemokine receptor 5 (CCR5) gene. Multiple transcripts with 5’-end heterogeneity, dual promoter usage, and evidence for polymorphisms within the regulatory regions and noncoding exons. J. Biol. Chem. 1997, 272, 30662–30671. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Day, C.J.; Morrison, N.A. MCP-1 Is Induced by Receptor Activator of Nuclear Factor-κB Ligand, Promotes Human Osteoclast Fusion, and Rescues Granulocyte Macrophage Colony-stimulating Factor Suppression of Osteoclast Formation. J. Boil. Chem. 2005, 280, 16163–16169. [Google Scholar] [CrossRef] [Green Version]
- Nishida, T.; Emura, K.; Kubota, S.; Lyons, K.M.; Takigawa, M. CCN family 2/connective tissue growth factor (CCN2/CTGF) promotes osteoclastogenesis via induction of and interaction with dendritic cell-specific transmembrane protein (DC-STAMP). J. Bone Miner. Res. 2011, 26, 351–363. [Google Scholar] [CrossRef]
- Jansen, B.J.; Eleveld-Trancikova, D.; Sanecka, A.; Van Hout-Kuijer, M.; Hendriks, I.A.; Looman, M.G.; Leusen, J.H.W.; Adema, G.J. OS9 interacts with DC-STAMP and modulates its intracellular localization in response to TLR ligation. Mol. Immunol. 2009, 46, 505–515. [Google Scholar] [CrossRef]
- Kanemoto, S.; Kobayashi, Y.; Yamashita, T.; Miyamoto, T.; Cui, M.; Asada, R.; Cui, X.; Hino, K.; Kaneko, M.; Takai, T.; et al. Luman is involved in osteoclastogenesis through the regulation of DC-STAMP expression, stability and localization. J. Cell Sci. 2015, 128, 4353–4365. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Lee, S.-H.; Kim, J.H.; Choi, Y.; Kim, N. NFATc1 induces osteoclast fusion via up-regulation of Atp6v0d2 and the dendritic cell-specific transmembrane protein (DC-STAMP). Mol. Endocrinol. 2007, 22, 176–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagi, M.; Ninomiya, K.; Fujita, N.; Suzuki, T.; Iwasaki, R.; Morita, K.; Hosogane, N.; Matsuo, K.; Toyama, Y.; Suda, T.; et al. Induction of DC-STAMP by Alternative Activation and Downstream Signaling Mechanisms. J. Bone Miner. Res. 2007, 22, 992–1001. [Google Scholar] [CrossRef] [PubMed]
- Mensah, K.A.; Ritchlin, C.T.; Schwarz, E.M. RANKL induces heterogeneous DC-STAMP(lo) and DC-STAMP(hi) osteoclast precursors of which the DC-STAMP(lo) precursors are the master fusogens. J. Cell Physiol. 2010, 223, 76–83. [Google Scholar] [PubMed] [Green Version]
- Chiu, Y.-H.; Mensah, K.A.; Schwarz, E.M.; Ju, Y.; Takahata, M.; Feng, C.; McMahon, L.A.; Hicks, D.G.; Panepento, B.; Keng, P.C.; et al. Regulation of human osteoclast development by dendritic cell-specific transmembrane protein (DC-STAMP). J. Bone Miner. Res. 2011, 27, 79–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, K.; Galson, D.L.; Zhao, C.; Peng, L.; LaPlace, C.; Wang, K.Z.Q.; Bachler, M.A.; Amano, H.; Aburatani, H.; Ishikawa, H.; et al. Nuclear Factor of Activated T-cells (NFAT) Rescues Osteoclastogenesis in Precursors Lacking c-Fos. J. Boil. Chem. 2004, 279, 26475–26480. [Google Scholar] [CrossRef] [Green Version]
- Asagiri, M.; Sato, K.; Usami, T.; Ochi, S.; Nishina, H.; Yoshida, H.; Morita, I.; Wagner, E.F.; Mak, T.W.; Serfling, E.; et al. Autoamplification of NFATc1 expression determines its essential role in bone homeostasis. J. Exp. Med. 2005, 202, 1261–1269. [Google Scholar] [CrossRef] [Green Version]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.-I.; et al. Induction and Activation of the Transcription Factor NFATc1 (NFAT2) Integrate RANKL Signaling in Terminal Differentiation of Osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Roskoski, R., Jr. ERK1/2 MAP kinases: Structure, function, and regulation. Pharmacol. Res. 2012, 66, 105–143. [Google Scholar] [CrossRef]
- He, Y.; Staser, K.; Rhodes, S.D.; Liu, Y.; Wu, X.; Park, S.-J.; Yuan, J.; Yang, X.; Li, X.; Jiang, L.; et al. Erk1 Positively Regulates Osteoclast Differentiation and Bone Resorptive Activity. PLoS ONE 2011, 6, e24780. [Google Scholar] [CrossRef]
- Oh, J.H.; Lee, J.Y.; Park, J.H.; No, J.H.; Lee, N.K. Obatoclax regulates the proliferation and fusion of osteoclast precursors through the inhibition of ERK activation by RANKL. Mol. Cells 2015, 38, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Li, S.; Feng, R.; Ma, H.; Sabeh, F.; Roodman, G.D.; Wang, J.; Robinson, S.T.; Guo, X.E.; Lund, T.; et al. Multiple myeloma-derived MMP-13 mediates osteoclast fusogenesis and osteolytic disease. J. Clin. Investig. 2016, 126, 1759–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- 1Kim, H.J.; Lee, Y. Endogenous Collagenases Regulate Osteoclast Fusion. Biomolecules 2020, 10, 705. [Google Scholar]
- Matsumoto, M.; Kogawa, M.; Wada, S.; Takayanagi, H.; Tsujimoto, M.; Katayama, S.; Hisatake, K.; Nogi, Y. Essential Role of p38 Mitogen-activated Protein Kinase in Cathepsin K Gene Expression during Osteoclastogenesis through Association of NFATc1 and PU.1. J. Boil. Chem. 2004, 279, 45969–45979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyauchi, Y.; Ninomiya, K.; Miyamoto, H.; Sakamoto, A.; Iwasaki, R.; Hoshi, H.; Miyamoto, K.; Hao, W.; Yoshida, S.; Morioka, H.; et al. The Blimp1-Bcl6 axis is critical to regulate osteoclast differentiation and bone homeostasis. J. Exp. Med. 2010, 207, 751–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ihn, H.J.; Lee, T.; Lee, D.; Bae, J.-S.; Kim, S.-H.; Jang, I.H.; Bae, Y.C.; Shin, H.-I.; Park, E.K. Inhibitory Effect of KP-A038 on Osteoclastogenesis and Inflammatory Bone Loss Is Associated With Downregulation of Blimp1. Front. Pharmacol. 2019, 10, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Yoo, J.H.; Lee, J.-H.; Lee, Y.; Bae, M.-K.; Kim, Y.-D.; Kim, H.J. Connective tissue growth factor (CTGF) regulates the fusion of osteoclast precursors by inhibiting Bcl6 in periodontitis. Int. J. Med. Sci. 2020, 17, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Courtial, N.; Smink, J.J.; Kuvardina, O.N.; Leutz, A.; Göthert, J.R.; Lausen, J. Tal1 regulates osteoclast differentiation through suppression of the master regulator of cell fusion DC-STAMP. FASEB J. 2011, 26, 523–532. [Google Scholar] [CrossRef]
- Luchin, A.; Suchting, S.; Merson, T.; Rosol, T.J.; Hume, D.A.; Cassady, A.; Ostrowski, M.C. Genetic and Physical Interactions betweenMicrophthalmiaTranscription Factor and PU.1 Are Necessary for Osteoclast Gene Expression and Differentiation. J. Boil. Chem. 2001, 276, 36703–36710. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.M.; Bronisz, A.; Hu, R.; Patel, K.; Mansky, K.C.; Sif, S.; Ostrowski, M.C. MITF and PU.1 Recruit p38 MAPK and NFATc1 to Target Genes during Osteoclast Differentiation. J. Boil. Chem. 2007, 282, 15921–15929. [Google Scholar] [CrossRef] [Green Version]
- So, H.; Rho, J.; Jeong, D.; Park, R.; Fisher, D.E.; Ostrowski, M.C.; Choi, Y.; Kim, N. Microphthalmia Transcription Factor and PU.1 Synergistically Induce the Leukocyte Receptor Osteoclast-associated Receptor Gene Expression. J. Boil. Chem. 2003, 278, 24209–24216. [Google Scholar] [CrossRef] [Green Version]
- Carey, H.A.; Hildreth, B.E.; Geisler, J.A.; Nickel, M.C.; Cabrera, J.; Ghosh, S.; Jiang, Y.; Yan, J.; Lee, J.; Makam, S.; et al. Enhancer variants reveal a conserved transcription factor network governed by PU.1 during osteoclast differentiation. Bone Res. 2018, 6, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, X.; Chen, X.; Yu, X. MicroRNAs in Osteoclastogenesis and Function: Potential Therapeutic Targets for Osteoporosis. Int. J. Mol. Sci. 2016, 17, 349. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Rao, L.; Jia, H.; Chen, J.; Lu, X.; Yang, G.; Li, Q.; Lee, K.K.H.; Yang, L. Baicalin positively regulates osteoclast function by activating MAPK/Mitf signalling. J. Cell. Mol. Med. 2017, 21, 1361–1372. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhang, J.; Shao, H.; Liu, J.; Jin, M.; Chen, J.; Huang, Y. miRNA-340 inhibits osteoclast differentiation via repression of MITF. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [PubMed]
- Bloor, A.J.; Sánchez, M.-J.; Green, A.R.; Göttgens, B. The Role of the Stem Cell Leukemia (SCL) Gene in Hematopoietic and Endothelial Lineage Specification. J. Hematotherapy 2002, 11, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Dou, C.; Zhang, C.; Kang, F.; Yang, X.; Jiang, H.; Bai, Y.; Xiang, J.; Xu, J.-Z.; Dong, S. MiR-7b directly targets DC-STAMP causing suppression of NFATc1 and c-Fos signaling during osteoclast fusion and differentiation. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2014, 1839, 1084–1096. [Google Scholar] [CrossRef]
- Maruyama, K.; Uematsu, S.; Kondo, T.; Takeuchi, O.; Martino, M.M.; Kawasaki, T.; Akira, S. Strawberry notch homologue 2 regulates osteoclast fusion by enhancing the expression of DC-STAMP. J. Exp. Med. 2013, 210, 1947–1960. [Google Scholar] [CrossRef]
- Carey, H.A.E.; Hildreth, B.; Samuvel, D.J.; Thies, K.A.; Rosol, T.J.; Toribio, R.E.; Charles, J.F.; Ostrowski, M.C.; Sharma, S.M. Eomes partners with PU.1 and MITF to Regulate Transcription Factors Critical for osteoclast differentiation. iScience 2019, 11, 238–245. [Google Scholar] [CrossRef] [Green Version]
- Faulkner, B.; Astleford, K.; Mansky, K.C. Regulation of Osteoclast Differentiation and Skeletal Maintenance by Histone Deacetylases. Molecules 2019, 24, 1355. [Google Scholar] [CrossRef] [Green Version]
- Dou, C.; Li, N.; Ding, N.; Liu, C.; Yang, X.; Kang, F.; Cao, Z.; Quan, H.; Hou, T.; Xu, J.; et al. HDAC2 regulates FoxO1 during RANKL-induced osteoclastogenesis. Am. J. Physiol. Physiol. 2016, 310, C780–C787. [Google Scholar] [CrossRef]
- Bartell, S.M.; Kim, H.-N.; Ambrogini, E.; Han, L.; Iyer, S.; Ucer, S.S.; Rabinovitch, P.; Jilka, R.L.; Weinstein, R.S.; Zhao, H.; et al. FoxO proteins restrain osteoclastogenesis and bone resorption by attenuating H2O2 accumulation. Nat. Commun. 2014, 5, 3773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, L.; Kaiser, B.; Romsa, A.; Schwarz, T.; Gopalakrishnan, R.; Jensen, E.D.; Mansky, K.C. HDAC3 and HDAC7 Have Opposite Effects on Osteoclast Differentiation. J. Boil. Chem. 2011, 286, 12056–12065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blixt, N.C.; Faulkner, B.K.; Astleford, K.; Lelich, R.; Schering, J.; Spencer, E.; Gopalakrishnan, R.; Jensen, E.D.; Mansky, K.C. Class II and IV HDACs function as inhibitors of osteoclast differentiation. PLoS ONE 2017, 12, e0185441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, R.; Bae, H.-S.; Yoon, W.-J.; Woo, K.M.; Baek, J.-H.; Kim, H.-H.; Uchida, T.; Ryoo, H.-M. Pin1 Regulates Osteoclast Fusion Through Suppression of the Master Regulator of Cell Fusion DC-STAMP. J. Cell. Physiol. 2014, 229, 2166–2174. [Google Scholar] [CrossRef]
- Cho, E.; Lee, J.-K.; Lee, J.-Y.; Chen, Z.; Ahn, S.-H.; Kim, N.D.; Kook, M.-S.; Min, S.H.; Ahn, B.-J.; Lee, T.-H. BCPA {N,N’-1,4-Butanediylbis[3 -(2-chlorophenyl)acrylamide]} inhibits osteoclast differentiation through increased retention of peptidyl-prolyl cis-trans isomerase never in mitosis a-interacting 1. Int. J. Mol. Sci. 2018, 19, 3436. [Google Scholar] [CrossRef] [Green Version]
- Eleveld-Trancikova, D.; Sanecka, A.; Van Hout-Kuijer, M.A.; Looman, M.W.; Hendriks, I.A.; Jansen, B.J.; Adema, G.J. DC-STAMP interacts with ER-resident transcription factor LUMAN which becomes activated during DC maturation. Mol. Immunol. 2010, 47, 1963–1973. [Google Scholar] [CrossRef]
- Yin, Y.; Tang, L.; Chen, J.; Lu, X. MiR-30a attenuates osteoclastogenesis via targeting DC-STAMP-c-Fos-NFATc1 signaling. Am. J. Transl. Res. 2017, 9, 5743–5753. [Google Scholar]
- The Genetic Determinants of Paget’s Disease (GDPD) Consortium; Albagha, O.M.E.; Wani, S.; Visconti, M.R.; Alonso, N.; Goodman, K.; Brandi, M.L.; Cundy, T.; Chung, P.Y.J.; Dargie, R.; et al. Genome-wide association identifies three new susceptibility loci for Paget’s disease of bone. Nat. Genet. 2011, 43, 685–689. [Google Scholar]
- Beauregard, M.; Gagnon, E.; Guay-Belanger, S.; Morissette, J.; Brown, J.; Michou, L. Identification of rare genetic variants in novel loci associated with Paget’s disease of bone. Qual. Life Res. 2013, 133, 755–768. [Google Scholar] [CrossRef]
- Laurier, E.; Amiable, N.; Gagnon, E.; Brown, J.; Michou, L. Effect of a rare genetic variant of TM7SF4 gene on osteoclasts of patients with Paget’s disease of bone. BMC Med. Genet. 2017, 18, 133. [Google Scholar] [CrossRef] [Green Version]
- Mullin, B.; Zhu, K.; Brown, S.J.; Mullin, S.; Tickner, J.; Pavlos, N.; Dudbridge, F.; Xu, J.; Walsh, J.P.; Wilson, S.G. Genetic regulatory mechanisms in human osteoclasts suggest a role for the STMP1 and DCSTAMP genes in Paget’s disease of bone. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultana, M.; Pavlos, N.; Ward, L.; Walsh, J.; Rea, S.L. Targeted sequencing of DCSTAMP in familial Paget’s disease of bone. Bone Rep. 2019, 10, 100198. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Park, M.; Baek, S.-H.; Kim, H.J.; Kim, S.H. Molecules and signaling pathways involved in the expression of OC-STAMP during osteoclastogenesis. Amino Acids 2010, 40, 1447–1459. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Hashimi, S.M.; Bakr, M.M.; Forwood, M.R.; Morrison, N.A. Foreign body giant cells and osteoclasts are TRAP positive, have podosome-belts and both require OC-STAMP for cell fusion. J. Cell. Biochem. 2013, 114, 1772–1778. [Google Scholar] [CrossRef]
- Hwang, Y.S.; Ma, G.T.; Park, K.K. Lysophosphatidic acid stimulates osteoclast fusion through OC-STAMP and P2 × 7 receptor signaling. J. Bone Miner. Metab. 2014, 32, 110–122. [Google Scholar] [CrossRef]
- Abe, E.; Mocharla, H.; Yamate, T.; Taguchi, Y.; Manolagas, S.C. Meltrin-α, a Fusion Protein Involved in Multinucleated Giant Cell and Osteoclast Formation. Calcif. Tissue Int. 1999, 64, 508–515. [Google Scholar] [CrossRef]
- Ishii, T.; Ruiz-Torruella, M.; Ikeda, A.; Shindo, S.; Movila, A.; Mawardi, H.; Albassam, A.; Kayal, R.A.; Al-Dharrab, A.A.; Egashira, K.; et al. OC-STAMP promotes osteoclast fusion for pathogenic bone resorption in periodontitis via up-regulation of permissive fusogen CD9. FASEB J. 2018, 32, 4016–4030. [Google Scholar] [CrossRef] [Green Version]
- Forgac, M. Vacuolar ATPases: Rotary proton pumps in physiology and pathophysiology. Nat. Rev. Mol. Cell Boil. 2007, 8, 917–929. [Google Scholar] [CrossRef]
- Holliday, L.S. Vacuolar H+-ATPases (V-ATPases) as therapeutic targets: A brief review and recent developments. Biotarget 2017, 1, 18. [Google Scholar] [CrossRef]
- Duan, X.; Yang, S.; Zhang, L.; Yang, T. V-ATPases and osteoclasts: Ambiguous future of V-ATPases inhibitors in osteoporosis. Theranostics 2018, 8. [Google Scholar] [CrossRef]
- Toei, M.; Saum, R.; Forgac, M. Regulation and Isoform Function of the V-ATPases. Biochemistry 2010, 49, 4715–4723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, A.; Cheng, T.S.; Pavlos, N.; Lin, Z.; Dai, K.; Zheng, M. V-ATPases in osteoclasts: Structure, function and potential inhibitors of bone resorption. Int. J. Biochem. Cell Boil. 2012, 44, 1422–1435. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Liu, N.; Xie, X.; Bi, G.; Ba, H.; Li, L.; Zhang, J.; Deng, X.; Yao, Y.; Tang, Z.; et al. The macrophage-specific V-ATPase subunit ATP6V0D2 restricts inflammasome activation and bacterial infection by facilitating autophagosome-lysosome fusion. Autophagy 2019, 15, 960–975. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Rho, J.; Jeong, D.; Sul, J.-Y.; Kim, T.; Kim, N.; Kang, J.-S.; Miyamoto, T.; Suda, T.; Lee, S.-K.; et al. v-ATPase V0 subunit d2–deficient mice exhibit impaired osteoclast fusion and increased bone formation. Nat. Med. 2006, 12, 1403–1409. [Google Scholar] [CrossRef]
- Kim, T.; Ha, H.; Kim, N.; Park, E.-S.; Rho, J.; Kim, E.C.; Lorenzo, J.; Choi, Y.; Lee, S.H. ATP6v0d2 deficiency increases bone mass, but does not influence ovariectomy-induced bone loss. Biochem. Biophys. Res. Commun. 2010, 403, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Cheng, T.S.; Steer, J.H.; Joyce, D.A.; Pavlos, N.; Leong, C.; Kular, J.; Liu, J.; Feng, X.; Zheng, M.H.; et al. Myocyte Enhancer Factor 2 and Microphthalmia-associated Transcription Factor Cooperate with NFATc1 to Transactivate the V-ATPase d2 Promoter during RANKL-induced Osteoclastogenesis. J. Boil. Chem. 2009, 284, 14667–14676. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.; Ha, H.-I.; Kim, N.; Yi, O.; Lee, S.H.; Choi, Y. Adrm1 interacts with Atp6v0d2 and regulates osteoclast differentiation. Biochem. Biophys. Res. Commun. 2009, 390, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Cai, L.; Meng, R.; Feng, Z.; Xu, Q. METTL3 Modulates Osteoclast Differentiation and Function by Controlling RNA Stability and Nuclear Export. Int. J. Mol. Sci. 2020, 21, 1660. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Zhao, Y.; He, J.; Zhang, Y.; Xi, H.; Liu, M.; Ma, J.; Wu, L. YTHDF2 destabilizes m6A-containing RNA through direct recruitment of the CCR4–NOT deadenylase complex. Nat. Commun. 2016, 7, 12626. [Google Scholar] [CrossRef]
- Makihira, S.; Nikawa, H.; Kajiya, M.; Kawai, T.; Mine, Y.; Kosaka, E.; Silva, M.; Tobiume, K.; Terada, Y. Blocking of sodium and potassium ion-dependent adenosine triphosphatase-α1 with ouabain and vanadate suppresses cell–cell fusion during RANKL-mediated osteoclastogenesis. Eur. J. Pharmacol. 2011, 670, 409–418. [Google Scholar] [CrossRef]
- Mine, Y.; Shuto, T.; Nikawa, H.; Kawai, T.; Ohara, M.; Kawahara, K.; Ohta, K.; Kukita, T.; Terada, Y.; Makihira, S. Inhibition of RANKL-dependent cellular fusion in pre-osteoclasts by amiloride and a NHE10-specific monoclonal antibody. Cell Boil. Int. 2015, 39, 696–709. [Google Scholar] [CrossRef] [PubMed]
- Saginario, C.; Sterling, H.; Beckers, C.; Kobayashi, R.; Solimena, M.; Ullu, E.; Vignery, A. MFR, a Putative Receptor Mediating the Fusion of Macrophages. Mol. Cell. Boil. 1998, 18, 6213–6223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saginario, C.; Qian, H.Y.; Vignery, A. Identification of an inducible surface molecule specific to fusing macrophages. Proc. Natl. Acad. Sci. USA 1995, 92, 12210–12214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Sterling, H.; Chen, Y.; Saginario, C.; Brown, E.J.; Frazier, W.A.; Lindberg, F.P.; Vignery, A. CD47, a Ligand for the Macrophage Fusion Receptor, Participates in Macrophage Multinucleation. J. Boil. Chem. 2000, 275, 37984–37992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundberg, P.; Koskinen, C.; Baldock, P.A.; Löthgren, H.; Stenberg, A.; Lerner, U.H.; Oldenborg, P.-A. Osteoclast formation is strongly reduced both in vivo and in vitro in the absence of CD47/SIRPalpha-interaction. Biochem. Biophys. Res. Commun. 2007, 352, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Koskinen, C.; Persson, E.; Baldock, P.; Stenberg, Å.; Boström, I.; Matozaki, T.; Oldenborg, P.-A.; Lundberg, P. Lack of CD47 Impairs Bone Cell Differentiation and Results in an Osteopenic Phenotype in Vivo due to Impaired Signal Regulatory Protein α (SIRPα) Signaling*. J. Boil. Chem. 2013, 288, 29333–29344. [Google Scholar] [CrossRef] [Green Version]
- Maile, L.A.; DeMambro, V.E.; Wai, C.; Aday, A.W.; Capps, B.E.; Beamer, W.G.; Rosen, C.J.; Clemmons, D.R. An essential role for the association of CD47 to SHPS-1 in skeletal remodeling. J. Bone Miner. Res. 2011, 26, 2068–2081. [Google Scholar] [CrossRef]
- Lee, B.S. Myosins in Osteoclast Formation and Function. Biomolecules 2018, 8, 157. [Google Scholar] [CrossRef] [Green Version]
- McMichael, B.K.; Wysolmerski, R.B.; Lee, B.S. Regulated Proteolysis of Nonmuscle Myosin IIA Stimulates Osteoclast Fusion. J. Boil. Chem. 2009, 284, 12266–12275. [Google Scholar] [CrossRef] [Green Version]
- Hobolt-Pedersen, A.-S.; Delaissé, J.M.; Søe, K. Osteoclast Fusion is Based on Heterogeneity Between Fusion Partners. Calcif. Tissue Int. 2014, 95, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Møller, A.M.J.; Delaissé, J.M.; Søe, K. Osteoclast Fusion: Time-Lapse Reveals Involvement of CD47 and Syncytin-1 at Different Stages of Nuclearity. J. Cell. Physiol. 2016, 232, 1396–1403. [Google Scholar] [CrossRef] [PubMed]
- Koduru, S.; Sun, B.-H.; Walker, J.M.; Zhu, M.; Simpson, C.; Dhodapkar, M.; Insogna, K.L. The contribution of cross-talk between the cell-surface proteins CD36 and CD47-TSP-1 in osteoclast formation and function. J. Boil. Chem. 2018, 293, 15055–15069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amend, S.R.; Uluckan, O.; Hurchla, M.; Leib, D.; Novack, D.V.; Silva, M.J.; Frazier, W.; Weilbaecher, K.N. Thrombospondin-1 Regulates Bone Homeostasis Through Effects on Bone Matrix Integrity and Nitric Oxide Signaling in Osteoclasts. J. Bone Miner. Res. 2014, 30, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podolnikova, N.P.; Hlaváčková, M.; Wu, Y.; Yakubenko, V.P.; Faust, J.; Balabiyev, A.; Wang, X.; Ugarova, T.P. Interaction between the integrin Mac-1 and signal regulatory protein α (SIRPα) mediates fusion in heterologous cells. J. Boil. Chem. 2019, 294, 7833–7849. [Google Scholar] [CrossRef]
- Mbalaviele, G.; Nishimura, R.; Myoi, A.; Niewolna, M.; Reddy, S.V.; Chen, D.; Feng, J.; Roodman, D.; Mundy, G.R.; Yoneda, T. Cadherin-6 Mediates the Heterotypic Interactions between the Hemopoietic Osteoclast Cell Lineage and Stromal Cells in a Murine Model of Osteoclast Differentiation. J. Cell Boil. 1998, 141, 1467–1476. [Google Scholar] [CrossRef] [Green Version]
- Fiorino, C.; Harrison, R.E. E-cadherin is important for cell differentiation during osteoclastogenesis. Bone 2016, 86, 106–118. [Google Scholar] [CrossRef]
- Quan, J.; Du, Q.; Hou, Y.; Wang, Z.; Zhang, J. Utilization of E-cadherin by monocytes from tumour cells plays key roles in the progression of bone invasion by oral squamous cell carcinoma. Oncol. Rep. 2017, 38, 850–858. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, H.; Nakashima, T.; Hayashi, M.; Izawa, N.; Yasui, T.; Aburatani, H.; Tanaka, S.; Takayanagi, H. Global epigenomic analysis indicates protocadherin-7 activates osteoclastogenesis by promoting cell–cell fusion. Biochem. Biophys. Res. Commun. 2014, 455, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Yasui, T.; Hirose, J.; Tsutsumi, S.; Nakamura, K.; Aburatani, H.; Tanaka, S. Epigenetic regulation of osteoclast differentiation: Possible involvement of Jmjd3 in the histone demethylation of Nfatc1. J. Bone Miner. Res. 2011, 26, 2665–2671. [Google Scholar] [CrossRef]
- Lavialle, C.; Cornelis, G.; Dupressoir, A.; Esnault, C.; Heidmann, O.; Vernochet, C.; Heidmann, T. Paleovirology of ‘syncytins’, retroviralenvgenes exapted for a role in placentation. Philos. Trans. R. Soc. B Boil. Sci. 2013, 368. [Google Scholar] [CrossRef] [Green Version]
- Mi, S.; Lee, X.; Li, X.-P.; Veldman, G.M.; Finnerty, H.; Racie, L.; LaVallie, E.; Tang, X.-Y.; Edouard, P.; Howes, S.; et al. Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis. Nature 2000, 403, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Gong, R.; Peng, X.; Kang, S.; Feng, H.; Huang, J.; Zhang, W.; Lin, D.; Tien, P.; Xiao, G. Structural characterization of the fusion core in syncytin, envelope protein of human endogenous retrovirus family W. Biochem. Biophys. Res. Commun. 2005, 331, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Soe, K.; Andersen, T.; Hobolt-Pedersen, A.-S.; Bjerregaard, B.; Larsson, L.-I.; Delaisse, J.-M. Involvement of human endogenous retroviral syncytin-1 in human osteoclast fusion. Bone 2011, 48, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Soe, K.; Hobolt-Pedersen, A.-S.; Delaisse, J.-M. The elementary fusion modalities of osteoclasts. Bone 2015, 73, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Leikina, E.; Melikov, K.; Gebert, C.; Kram, V.; Young, M.F.; Uygur, B.; Chernomordik, L.V. Cell-surface phosphatidylserine regulates osteoclast precursor fusion. J. Boil. Chem. 2017, 293, 254–270. [Google Scholar] [CrossRef] [Green Version]
- Irie, A.; Yamamoto, K.; Miki, Y.; Murakami, M. Phosphatidylethanolamine dynamics are required for osteoclast fusion. Sci. Rep. 2017, 7, 46715. [Google Scholar] [CrossRef] [Green Version]
- Coudert, A.; Redelsperger, F.; Chabbi-Achengli, Y.; Vernochet, C.; Marty, C.; Decrouy, X.; Heidmann, T.; De Vernejoul, M.-C.; Dupressoir, A. Role of the captured retroviral envelope syncytin-B gene in the fusion of osteoclast and giant cell precursors and in bone resorption, analyzed ex vivo and in vivo in syncytin-B knockout mice. Bone Rep. 2019, 11, 100214. [Google Scholar] [CrossRef]
- Humphrey, M.B.; Nakamura, M.C. A Comprehensive Review of Immunoreceptor Regulation of Osteoclasts. Clin. Rev. Allergy Immunol. 2015, 51, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Barrow, A.D.; Raynal, N.; Andersen, T.; Slatter, D.A.; Bihan, M.; Pugh, N.; Cella, M.; Kim, T.; Rho, J.; Negishi-Koga, T.; et al. OSCAR is a collagen receptor that costimulates osteoclastogenesis in DAP12-deficient humans and mice. J. Clin. Investig. 2011, 121, 3505–3516. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, M.B.; Daws, M.R.; Spusta, S.C.; Niemi, E.C.A.; Torchia, J.; Lanier, L.L.; E. Seaman, W.; Nakamura, M.C. TREM2, a DAP12-Associated Receptor, Regulates Osteoclast Differentiation and Function. J. Bone Miner. Res. 2005, 21, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Ishida-Kitagawa, N.; Tanaka, K.; Bao, X.; Kimura, T.; Miura, T.; Kitaoka, Y.; Hayashi, K.; Sato, M.; Maruoka, M.; Ogawa, T.; et al. Siglec-15 Protein Regulates Formation of Functional Osteoclasts in Concert with DNAX-activating Protein of 12 kDa (DAP12). J. Boil. Chem. 2012, 287, 17493–17502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, N.; Takami, M.; Rho, J.; Josien, R.; Choi, Y. A Novel Member of the Leukocyte Receptor Complex Regulates Osteoclast Differentiation. J. Exp. Med. 2002, 195, 201–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merck, E.; Gaillard, C.; Gorman, D.M.; Montero-Julian, F.; Durand, I.; Zurawski, S.M.; Menetrier-Caux, C.; Carra, G.; Lebecque, S.; Trinchieri, G.; et al. OSCAR is an FcRgamma-associated receptor that is expressed by myeloid cells and is involved in antigen presentation and activation of human dendritic cells. Blood 2004, 104, 1386–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Søe, K.; Andersen, T.; Hinge, M.; Rolighed, L.; Marcussen, N.; Delaissé, J.M. Coordination of Fusion and Trafficking of Pre-osteoclasts at the Marrow–Bone Interface. Calcif. Tissue Int. 2019, 105, 430–445. [Google Scholar] [CrossRef]
- Helming, L.; Tomasello, E.; Kyriakides, T.R.; Martínez, F.O.; Takai, T.; Gordon, S.; Vivier, E. Essential Role of DAP12 Signaling in Macrophage Programming into a Fusion-Competent State. Sci. Signal. 2008, 1, ra11. [Google Scholar] [CrossRef] [Green Version]
- Tomasello, E.; Olcese, L.; Vély, F.; Geourgeon, C.; Bléry, M.; Moqrich, A.; Gautheret, D.; Djabali, M.; Mattei, M.; Vivier, E. Gene structure, expression pattern, and biological activity of mouse killer cell activating receptor-associated protein (KARAP)/DAP-12. J. Biol. Chem. 1998, 273, 34115–34119. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, M.B.; Ogasawara, K.; Yao, W.; Spusta, S.C.; Daws, M.R.; Lane, N.E.; Lanier, L.L.; Nakamura, M.C. The Signaling Adapter Protein DAP12 Regulates Multinucleation During Osteoclast Development. J. Bone Miner. Res. 2003, 19, 224–234. [Google Scholar] [CrossRef]
- Negishi-Koga, T.; Inui, M.; Inoue, K.; Kim, S.; Suematsu, A.; Kobayashi, E.; Iwata, T.; Ohnishi, H.; Matozaki, T.; Kodama, T.; et al. Costimulatory signals mediated by the ITAM motif cooperate with RANKL for bone homeostasis. Nature 2004, 428, 758–763. [Google Scholar] [CrossRef]
- Angata, T.; Tabuchi, Y.; Nakamura, K.; Nakamura, M. Siglec-15: An immune system Siglec conserved throughout vertebrate evolution. Glycobiology 2007, 17, 838–846. [Google Scholar] [CrossRef]
- Hiruma, Y.; Hirai, T.; Tsuda, E. Siglec-15, a member of the sialic acid-binding lectin, is a novel regulator for osteoclast differentiation. Biochem. Biophys. Res. Commun. 2011, 409, 424–429. [Google Scholar] [CrossRef]
- Kameda, Y.; Takahata, M.; Komatsu, M.; Mikuni, S.; Hatakeyama, S.; Shimizu, T.; Angata, T.; Kinjo, M.; Minami, A.; Iwasaki, N. Siglec-15 Regulates Osteoclast Differentiation by Modulating RANKL-Induced Phosphatidylinositol 3-Kinase/Akt and Erk Pathways in Association With Signaling Adaptor DAP12. J. Bone Miner. Res. 2013, 28, 2463–2475. [Google Scholar] [CrossRef] [PubMed]
- Hiruma, Y.; Tsuda, E.; Maeda, N.; Okada, A.; Kabasawa, N.; Miyamoto, M.; Hattori, H.; Fukuda, C. Impaired osteoclast differentiation and function and mild osteopetrosis development in Siglec-15-deficient mice. Bone 2013, 53, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Chen, Y.-J.; Fan, C.-Y.; Tang, C.-J.; Low, P.-Y.; Ventura, A.; Lin, C.-C.; Angata, T.; Chen, Y.-H.; Chen, Y.-J. Identification of Siglec Ligands Using a Proximity Labeling Method. J. Proteome Res. 2017, 16, 3929–3941. [Google Scholar] [CrossRef]
- Zou, W.; Kitaura, H.; Reeve, J.; Long, F.; Tybulewicz, V.L.J.; Shattil, S.J.; Ginsberg, M.H.; Ross, F.P.; Teitelbaum, S.L. Syk, c-Src, the alphavbeta3 integrin, and ITAM immunoreceptors, in concert, regulate osteoclastic bone resorption. J. Cell Biol. 2007, 176, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Kameda, Y.; Takahata, M.; Mikuni, S.; Shimizu, T.; Hamano, H.; Angata, T.; Hatakeyama, S.; Kinjo, M.; Iwasaki, N. Siglec-15 is a potential therapeutic target for postmenopausal osteoporosis. Bone 2015, 71, 217–226. [Google Scholar] [CrossRef]
- Nakamura, I.; Takahashi, N.; Sasaki, T.; Tanaka, S.; Udagawa, N.; Murakami, H.; Kimura, K.; Kabuyama, Y.; Kurokawa, T.; Suda, T.; et al. Wortmannin, a specific inhibitor of phosphatidylinositol-3 kinase, blocks osteoclastic bone resorption. FEBS Lett. 1995, 361, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, T.; Nagase, Y.; Iwasawa, M.; Yasui, T.; Masuda, H.; Kadono, Y.; Nakamura, K.; Tanaka, S. Distinguishing the proapoptotic and antiresorptive functions of risedronate in murine osteoclasts: Role of the Akt pathway and the ERK/Bim axis. Arthritis Rheum. 2011, 63, 3908–3917. [Google Scholar] [CrossRef]
- Song, R.; Liu, X.; Zhu, J.; Zhang, J.; Gao, Q.; Zhao, H.; Sheng, A.; Yuan, Y.; Gu, J.; Zou, H.; et al. New roles of filopodia and podosomes in the differentiation and fusion process of osteoclasts. Genet. Mol. Res. 2014, 13, 4776–4787. [Google Scholar] [CrossRef]
- Takito, J.; Nakamura, M.; Yoda, M.; Tohmonda, T.; Uchikawa, S.; Horiuchi, K.; Toyama, Y.; Chiba, K. The transient appearance of zipper-like actin superstructures during the fusion of osteoclasts. J. Cell Sci. 2012, 125, 662–672. [Google Scholar] [CrossRef] [Green Version]
- Takito, J.; Otsuka, H.; Inoue, S.; Kawashima, T.; Nakamura, M. Symmetrical retrograde actin flow in the actin fusion structure is involved in osteoclast fusion. Boil. Open 2017, 6, 1104–1114. [Google Scholar] [CrossRef] [Green Version]
- Stuible, M.; Moraitis, A.; Fortin, A.; Saragosa, S.; Kalbakji, A.; Filion, M.; Tremblay, G.B. Mechanism and Function of Monoclonal Antibodies Targeting Siglec-15 for Therapeutic Inhibition of Osteoclastic Bone Resorption. J. Boil. Chem. 2014, 289, 6498–6512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, D.; Takahata, M.; Ota, M.; Fukuda, C.; Tsuda, E.; Shimizu, T.; Okada, A.; Hiruma, Y.; Hamano, H.; Hiratsuka, S.; et al. Siglec-15-targeting therapy increases bone mass in rats without impairing skeletal growth. Bone 2018, 116, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Sato, D.; Takahata, M.; Ota, M.; Fukuda, C.; Hasegawa, T.; Yamamoto, T.; Amizuka, N.; Tsuda, E.; Okada, A.; Hiruma, Y.; et al. Siglec-15-targeting therapy protects against glucocorticoid-induced osteoporosis of growing skeleton in juvenile rats. Bone 2020, 135, 115331. [Google Scholar] [CrossRef] [PubMed]
- Chung, D.-H.; Humphrey, M.B.; Nakamura, M.C.; Ginzinger, D.G.; Seaman, W.E.; Daws, M.R. CMRF-35-like molecule-1, a novel mouse myeloid receptor, can inhibit osteoclast formation. J. Immunol. 2003, 171, 6541–6548. [Google Scholar] [CrossRef] [Green Version]
- Van Beek, E.M.; De Vries, T.J.; Mulder, L.; Schoenmaker, T.; Hoeben, K.A.; Matozaki, T.; Langenbach, G.E.J.; Kraal, G.; Everts, V.; Berg, T.K.V.D. Inhibitory regulation of osteoclast bone resorption by signal regulatory protein α. FASEB J. 2009, 23, 4081–4090. [Google Scholar] [CrossRef] [Green Version]
- Aoki, K.; Didomenico, E.; Sims, N.A.; Mukhopadhyay, K.; Neff, L.; Houghton, A.; Amling, M.; Levy, J.B.; Horne, W.C.; Baron, R. The tyrosine phosphatase SHP-1 is a negative regulator of osteoclastogenesis and osteoclast resorbing activity: Increased resorption and osteopenia in me(v)/me(v) mutant mice. Bone 1999, 25, 261–267. [Google Scholar] [CrossRef]
- Faccio, R.; Teitelbaum, S.L.; Fujikawa, K.; Chappel, J.; Zallone, A.; Tybulewicz, V.L.J.; Ross, F.P.; Swat, W. Vav3 regulates osteoclast function and bone mass. Nat. Med. 2005, 11, 284–290. [Google Scholar] [CrossRef]
- Maruyama, K.; Kawasaki, T.; Hamaguchi, M.; Hashimoto, M.; Furu, M.; Ito, H.; Fujii, T.; Takemura, N.; Karuppuchamy, T.; Kondo, T.; et al. Bone-protective Functions of Netrin 1 Protein. J. Boil. Chem. 2016, 291, 23854–23868. [Google Scholar] [CrossRef] [Green Version]
- Shirakawa, J.; Takegahara, N.; Kim, H.; Lee, S.H.; Sato, K.; Yamagishi, S.; Choi, Y. Flrt2 is involved in fine-tuning of osteoclast multinucleation. BMB Rep. 2019, 52, 514–519. [Google Scholar] [CrossRef] [Green Version]
- Griffin, M.; Casadio, R.; Bergamini, C.M. Transglutaminases: Nature’s biological glues. Biochem. J. 2002, 368, 377–396. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Shinki, T.; Takito, J.; Jin, C.H.; Suda, T. Transglutaminase is involved in the fusion of mouse alveolar macrophages induced by 1α,25-dihydroxyvitamin D3. Exp. Cell Res. 1991, 192, 165–172. [Google Scholar] [CrossRef]
- Raghu, H.; Cruz, C.; Rewerts, C.L.; Frederick, M.D.; Thornton, S.; Mullins, E.; Schoenecker, J.G.; Degen, J.L.; Flick, M.J. Transglutaminase factor XIII promotes arthritis through mechanisms linked to inflammation and bone erosion. Blood 2015, 125, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Mousa, A.; Cui, C.; Song, A.; Myneni, V.D.; Sun, H.; Li, J.J.; Murshed, M.; Melino, G.; Kaartinen, M.T. Transglutaminases factor XIII-A and TG2 regulate resorption, adipogenesis and plasma fibronectin homeostasis in bone and bone marrow. Cell Death Differ. 2017, 24, 844–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.-S.; Kim, H.; Jeong, E.M.; Kim, H.J.; Lee, Z.H.; Kim, I.-G.; Kim, H. Transglutaminase 2 regulates osteoclast differentiation via a Blimp1-dependent pathway. Sci. Rep. 2017, 7, 10626. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Kaartinen, M.T. Assessment of expression and specific activities of transglutaminases TG1, TG2, and FXIII-A during osteoclastogenesis. Anal. Biochem. 2020, 591, 113512. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Kaartinen, M.T. Transglutaminase activity regulates differentiation, migration and fusion of osteoclasts via affecting actin dynamics. J. Cell. Physiol. 2018, 233, 7497–7513. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Yao, P.; Zhao, H. P2X7, a critical regulator and potential target for bone and joint diseases. J. Cell. Physiol. 2018, 234, 2095–2103. [Google Scholar] [CrossRef]
- Falzoni, S.; Chiozz, P.; Ferrari, D.; Buell, G.; Di Virgilio, F. P2 × 7 Receptor and polykarion formation. Mol. Biol. Cell. 2000, 11, 3169–3176. [Google Scholar] [CrossRef] [Green Version]
- Gartland, A.; Buckley, K.A.; Hipskind, R.A.; Perry, M.J.; Tobias, J.H.; Buell, G.; Bowler, W.B.; Gallagher, J.A. Multinucleated osteoclast formation in vivo and in vitro by P2 × 7 receptor-deficient mice. Crit. Rev. Eukaryot Gene Expr. 2003, 13, 243–253. [Google Scholar]
- Agrawal, A.; Buckley, K.A.; Bowers, K.; Furber, M.; Gallagher, J.A.; Gartland, A. The effects of P2 × 7 receptor antagonists on the formation and function of human osteoclasts in vitro. Purinergic Signal. 2010, 6, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Gartland, A.; Buckley, K.A.; Bowler, W.B.; Gallagher, J.A. Blockade of the pore-forming P2 × 7 receptor inhibits formation of multinucleated human osteoclasts in vitro. Calcif. Tissue Int. 2003, 73, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Hiken, J.F.; Steinberg, T.H. ATP downregulates P2 × 7 and inhibits osteoclast formation in RAW cells. Am. J. Physiol. Cell Physiol. 2004, 287, C403–C412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegatti, P.; Falzoni, S.; Donvito, G.; Irma, L.; Virgilio, F.D. P2 × 7 receptor drives osteoclast fusion by increasing the extracellular adenosine concentration. FASEB J. 2011, 25, 1264–1274. [Google Scholar] [CrossRef]
- Knowles, H.J. The Adenosine A2B Receptor Drives Osteoclast-Mediated Bone Resorption in Hypoxic Microenvironments. Cells 2019, 8, 624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aeschlimann, D.; Knäuper, V. P2 × 7 receptor-mediated TG2 externalization: A link to inflammatory arthritis? Amino Acids 2017, 49, 453–460. [Google Scholar]
- Adamczyk, M.; Griffiths, R.; Dewitt, S.; Knäuper, V.; Aeschlimann, D. P2 × 7 receptor activation regulates rapid unconventional export of transglutaminase-2. J. Cell Sci. 2015, 128, 4615–4628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syberg, S.; Petersen, S.; Jensen, J.-E.B.; Gartland, A.; Teilmann, J.; Chessell, I.; Steinberg, T.H.; Schwarz, P.; Jørgensen, N.R. Genetic background strongly influences the bone phenotype of P2 × 7 receptor knockout mice. J. Osteoporos. 2012, 2012, 391097. [Google Scholar] [CrossRef] [Green Version]
- Ke, H.Z.; Qi, H.; Weidema, A.F.; Zhang, Q.; Panupinthu, N.; Crawford, D.T.; Grasser, W.A.; Paralkar, V.M.; Li, M.; Audoly, L.P.; et al. Deletion of the P2 × 7 nucleotide receptor reveals its regulatory roles in bone formation and resorption. Mol. Endocrinol. 2003, 17, 1356–1367. [Google Scholar] [CrossRef] [Green Version]
- Gartland, A.; Skarratt, K.K.; Hocking, L.J.; Parsons, C.; Stokes, L.; Jørgensen, N.R.; Fraser, W.D.; Reid, D.M.; Gallagher, J.A.; Wiley, J.S.; et al. Polymorphisms in the P2 × 7 receptor gene are associated with low lumbar spine bone mineral density and accelerated bone loss in post-menopausal women. Eur. J. Hum. Genet. 2012, 20, 559–564. [Google Scholar] [CrossRef]
- Ohlendorff, S.D.; Tofteng, C.L.; Jensen, J.-E.B.; Petersen, S.; Civitelli, R.; Fenger, M.; Abrahamsen, B.; Hermann, A.P.; Eiken, P.; Jørgensen, N.R.; et al. Single nucleotide polymorphisms in the P2 × 7 gene are associated to fracture risk and to effect of estrogen treatment. Pharmacogenet. Genom. 2007, 17. [Google Scholar] [CrossRef]
- Li, J.; Liu, D.; Ke, H.Z.; Duncan, R.L.; Turner, C.H. The P2 × 7 nucleotide receptor mediates skeletal mechanotransduction. J. Biol. Chem. 2005, 280, 42952–42959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, W.Y.; Fritton, J.C.; Morgan, S.A.; Seref, F.Z.; Basta, -P.J.; Thi, M.M.; Suadicani, S.O.; Spray, D.C.; Majeska, R.J.; Schaffler, M.B.; et al. Pannexin-1 and P2 × 7-receptor are required for apoptotic osteocytes in fatigued bone to trigger RANKL production in neighboring bystander osteocytes. J. Bone Miner. Res. 2016, 31, 890–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gartland, A.; Hipskind, R.A.; Gallagher, J.A.; Bowler, W.B. Expression of a P2 × 7 receptor by a subpopulation of human osteoblasts. J. Bone Miner. Res. 2001, 16, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Walsh, M.C.; Yu, J.; Laskoski, P.; Takigawa, K.; Takegahara, N.; Choi, Y. Methylosome protein 50 associates with the purinergic receptor P2 × 5 and is involved in osteoclast maturation. FEBS Lett. 2020, 594, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kajikawa, T.; Walsh, M.C.; Takegahara, N.; Jeong, Y.H.; Hajishengallis, G.; Choi, Y. The purinergic receptor P2 × 5 contributes to bone loss in experimental periodontitis. BMB Rep. 2018, 51, 468–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Walsh, M.C.; Takegahara, N.; Middleton, S.A.; Shin, H.-I.; Kim, J.; Choi, Y. The purinergic receptor P2 × 5 regulates inflammasome activity and hyper-multinucleation of murine osteoclasts. Sci. Rep. 2017, 7, 196. [Google Scholar] [CrossRef] [Green Version]
- Yen, M.-L.; Hsu, P.-N.; Liao, H.-J.; Lee, B.-H.; Tsai, H.-F. TRAF-6 Dependent Signaling Pathway Is Essential for TNF-Related Apoptosis-Inducing Ligand (TRAIL) Induces Osteoclast Differentiation. PLoS ONE 2012, 7, e38048. [Google Scholar] [CrossRef]
- Yen, M.-L.; Tsai, H.-F.; Wu, Y.-Y.; Hwa, H.-L.; Lee, B.-H.; Hsu, P.-N. TNF-related apoptosis-inducing ligand (TRAIL) induces osteoclast differentiation from monocyte/macrophage lineage precursor cells. Mol. Immunol. 2008, 45, 2205–2213. [Google Scholar] [CrossRef]
- Chatani, M.; Mantoku, A.; Takeyama, K.; Abduweli, D.; Sugamori, Y.; Aoki, K.; Ohya, K.; Suzuki, H.; Uchida, S.; Sakimura, T.; et al. Microgravity promotes osteoclast activity in medaka fish reared at the international space station. Sci. Rep. 2015, 5, 14172. [Google Scholar] [CrossRef] [Green Version]
- Shanmugarajan, S.; Zhang, Y.; Moreno-Villanueva, M.; Clanton, R.; Rohde, L.H.; Ramesh, G.T.; Sibonga, J.; Wu, H. Combined Effects of Simulated Microgravity and Radiation Exposure on Osteoclast Cell Fusion. Int. J. Mol. Sci. 2017, 18, 2443. [Google Scholar] [CrossRef] [Green Version]
- Sambandam, Y.; Baird, K.L.; Stroebel, M.; Kowal, E.; Balasubramanian, S.; Reddy, S.V. Microgravity Induction of TRAIL Expression in Preosteoclast Cells Enhances Osteoclast Differentiation. Sci. Rep. 2016, 6, 25143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, H.-J.; Tsai, H.-F.; Wu, C.-S.; Chyuan, I.-T.; Hsu, P.-N. TRAIL inhibits RANK signaling and suppresses osteoclast activation via inhibiting lipid raft assembly and TRAF6 recruitment. Cell Death Dis. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhang, C.; Guo, H.; Chen, J.; Tao, Y.; Wang, F.; Lin, X.; Liu, Q.; Su, L.; Qin, A. Pregnenolone Inhibits Osteoclast Differentiation and Protects Against Lipopolysaccharide-Induced Inflammatory Bone Destruction and Ovariectomy-Induced Bone Loss. Front. Pharmacol. 2020, 11, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, M.H.; Lee, K.; Kim, M.Y.; Shin, H.-I.; Jeong, D. Pisidium coreanum Inhibits Multinucleated Osteoclast Formation and Prevents Estrogen-Deficient Osteoporosis. Int. J. Mol. Sci. 2019, 20, 6076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, P.; Hu, X.; Powers, J.; Nay, N.; Chang, E.; Kwon, J.; Wong, S.W.; Han, L.; Wu, T.-H.; Lee, D.-J.; et al. CDDO-Me, Sulforaphane and tBHQ attenuate the RANKL-induced osteoclast differentiation via activating the NRF2-mediated antioxidant response. Biochem. Biophys. Res. Commun. 2019, 511, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Takagi, T.; Inoue, H.; Takahashi, N.; Katsumata-Tsuboi, R.; Uehara, M. Sulforaphene attenuates multinucleation of pre-osteoclasts by suppressing expression of cell–cell fusion-associated genes DC-STAMP, OC-STAMP, and Atp6v0d2. Biosci. Biotechnol. Biochem. 2017, 81, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Takagi, T.; Inoue, H.; Takahashi, N.; Katsumata-Tsuboi, R.; Uehara, M. Sulforaphane inhibits osteoclast differentiation by suppressing the cell-cell fusion molecules DC-STAMP and OC-STAMP. Biochem. Biophys. Res. Commun. 2017, 483, 718–724. [Google Scholar] [CrossRef]
- Choi, S.-W.; Kim, S.-H.; Lee, K.-S.; Kang, H.J.; Lee, M.J.; Park, K.-I.; Lee, J.H.; Park, K.D.; Seo, W.D. Barley Seedling Extracts Inhibit RANKL-Induced Differentiation, Fusion, and Maturation of Osteoclasts in the Early-to-Late Stages of Osteoclastogenesis. Evid. Based Complement. Altern. Med. 2017, 2017, 1–12. [Google Scholar] [CrossRef]
- Zeng, X.; He, L.; Wang, S.; Wang, K.; Zhang, Y.; Tao, L.; Li, X.; Liu, S. Aconine inhibits RANKL-induced osteoclast differentiation in RAW264.7 cells by suppressing NF-kappaB and NFATc1 activation and DC-STAMP expression. Acta Pharmacol. Sin. 2016, 37, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Chai, R.C.; McDonald, M.M.; Terry, R.L.; Kovačić, N.; Down, J.M.; Pettitt, J.A.; Mohanty, S.T.; Shah, S.; Haffari, G.; Xu, J.; et al. Melphalan modifies the bone microenvironment by enhancing osteoclast formation. Oncotarget 2017, 8, 68047–68058. [Google Scholar] [CrossRef] [Green Version]
- Arioka, M.; Takahashi-Yanaga, F.; Tatsumoto, N.; Sasaguri, T. Inorganic phosphate-induced impairment of osteoclast cell-cell fusion by the inhibition of AP-1-mediated DC-STAMP expression. Biochem. Biophys. Res. Commun. 2017, 493, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, M.; Menuki, K.; Murai, T.; Hatakeyama, A.; Takada, S.; Furukawa, K.; Sakai, A. Elcatonin prevents bone loss caused by skeletal unloading by inhibiting preosteoclast fusion through the unloading-induced high expression of calcitonin receptors in bone marrow cells. Bone 2016, 85, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Dou, C.; Li, J.; Kang, F.; Cao, Z.; Yang, X.; Jiang, H.; Yang, B.; Xiang, J.; Xu, J.; Dong, S. Dual Effect of Cyanidin on RANKL-Induced Differentiation and Fusion of Osteoclasts. J. Cell. Physiol. 2015, 231, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Kuriya, K.; Nishio, M.; Matsuda, T.; Umekawa, H. Tea extract increases cell fusion via regulation of cell surface DC-STAMP. Biochem. Biophys. Rep. 2020, 22, 100759. [Google Scholar] [CrossRef] [PubMed]
- Kanzaki, H.; Movila, A.; Kayal, R.; Napimoga, M.H.; Egashira, K.; Dewhirst, F.E.; Sasaki, H.; Howait, M.; Al-Dharrab, A.; Mira, A.; et al. Phosphoglycerol dihydroceramide, a distinctive ceramide produced by Porphyromonas gingivalis, promotes RANKL-induced osteoclastogenesis by acting on non-muscle myosin II-A (Myh9), an osteoclast cell fusion regulatory factor. Biochim. Biophys. Acta (BBA) Mol. Cell Boil. Lipids 2017, 1862, 452–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ausk, B.J.; Worton, L.E.; Smigiel, K.S.; Kwon, R.Y.; Bain, S.D.; Srinivasan, S.; Gardiner, E.M.; Gross, T.S. Muscle paralysis induces bone marrow inflammation and predisposition to formation of giant osteoclasts. Am. J. Physiol. Physiol. 2017, 313, C533–C540. [Google Scholar] [CrossRef] [Green Version]
- Fujita, K.; Iwasaki, M.; Ochi, H.; Fukuda, T.; Ma, C.; Miyamoto, T.; Takitani, K.; Negishi-Koga, T.; Sunamura, S.; Kodama, T.; et al. Vitamin E decreases bone mass by stimulating osteoclast fusion. Nat. Med. 2012, 18, 589–594. [Google Scholar] [CrossRef]
- Ikegami, H.; Kawawa, R.; Ichi, I.; Ishikawa, T.; Koike, T.; Aoki, Y.; Fujiwara, Y. Excessive Vitamin E Intake Does Not Cause Bone Loss in Male or Ovariectomized Female Mice Fed Normal or High-Fat Diets. J. Nutr. 2017, 147, 1932–1937. [Google Scholar] [CrossRef] [Green Version]
- Kasai, S.; Ito, A.; Shindo, K.; Toyoshi, T.; Bando, M. High-Dose α-Tocopherol Supplementation Does Not Induce Bone Loss in Normal Rats. PLoS ONE 2015, 10, e0132059. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, X. Associations between serum vitamin E concentration and bone mineral density in the US elderly population. Osteoporos. Int. 2016, 28, 1245–1253. [Google Scholar] [CrossRef]
- Ilesanmi-Oyelere, B.L.; Brough, L.; Heath, A.-L.; Roy, N.C.; Kruger, M.C. The Relationship between Nutrient Patterns and Bone Mineral Density in Postmenopausal Women. Nutrients 2019, 11, 1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odai, T.; Terauchi, M.; Hirose, A.; Kato, K.; Miyasaka, N. Bone Mineral Density in Premenopausal Women Is Associated with the Dietary Intake of α-Tocopherol: A Cross-Sectional Study. Nutrients 2019, 11, 2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.C.; Duthie, G.G.; Aucott, L.S.; Macdonald, H.M. Vitamin E homologues α- and γ-tocopherol are not associated with bone turnover markers or bone mineral density in peri-menopausal and post-menopausal women. Osteoporos. Int. 2016, 27, 2281–2290. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Mohamad, N.V.; Ibrahim, N.; Chin, K.; Shuid, A.N.; Ima-Nirwana, S. The Molecular Mechanism of Vitamin E as a Bone-Protecting Agent: A Review on Current Evidence. Int. J. Mol. Sci. 2019, 20, 1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levaot, N.; Ottolenghi, A.; Mann, M.; Ram, G.G.; Kam, Z.; Geiger, B. Osteoclast fusion is initiated by a small subset of RANKL-stimulated monocyte progenitors, which can fuse to RANKL-unstimulated progenitors. Bone 2015, 79, 21–28. [Google Scholar] [CrossRef]
- Verma, S.K.; Leikina, E.; Melikov, K.; Chernomordik, L.V. Late stages of the synchronized macrophage fusion in osteoclast formation depend on dynamin. Biochem. J. 2014, 464, 293–300. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kodama, J.; Kaito, T. Osteoclast Multinucleation: Review of Current Literature. Int. J. Mol. Sci. 2020, 21, 5685. https://doi.org/10.3390/ijms21165685
Kodama J, Kaito T. Osteoclast Multinucleation: Review of Current Literature. International Journal of Molecular Sciences. 2020; 21(16):5685. https://doi.org/10.3390/ijms21165685
Chicago/Turabian StyleKodama, Joe, and Takashi Kaito. 2020. "Osteoclast Multinucleation: Review of Current Literature" International Journal of Molecular Sciences 21, no. 16: 5685. https://doi.org/10.3390/ijms21165685
APA StyleKodama, J., & Kaito, T. (2020). Osteoclast Multinucleation: Review of Current Literature. International Journal of Molecular Sciences, 21(16), 5685. https://doi.org/10.3390/ijms21165685