The Role of MicroRNAs in Epidermal Barrier

Abstract
:1. Introduction
2. Association of miRNAs with Keratinocyte Differentiation and Proliferation
2.1. Factors Regulating Keratinocyte Differentiation and Proliferation
2.1.1. Epidermal Calcium Gradients
2.1.2. p63
2.1.3. Notch Signaling
2.2. Keratinocyte Proliferation
2.2.1. Psoriasis
2.2.2. Wound Healing
2.2.3. Cutaneous Squamous Cell Carcinoma
2.2.4. HPV Infection
2.3. Structural Proteins of Corneocytes and Cornified Envelopes
2.3.1. Filaggrin, Involucrin, Loricrin and their Transcription Factors
2.3.2. S100 and its Transcription Factors
2.3.3. Others
3. Role of miRNAs in Cell-Cell Adhesion
3.1. Synthesis of Corneodesmosomes
3.2. Degradation of Corneodesmosomes
3.3. Others
4. Role of miRNAs in Skin Lipids
4.1. Skin Lipids
4.2. Epidermal Receptors Binding Skin Lipids
5. Conclusions
Funding
Conflicts of Interest
Abbreviations
| ADAM19 | a disintegrin and metalloproteinase domain 19 |
| AKT3 | AKT serine/threonine kinase 3 |
| ANG-1 | angiopoietin-1 |
| AP-1 | activator protein 1 |
| ASPP | apoptosis-stimulating protein of p53 |
| ATP2C1 | ATPase secretory pathway Ca2+ transporting 1 |
| AVEN | apoptosis and caspase activation inhibitor |
| BCL2L2 | Bcl-2-like 2 |
| BNIP3 | Bcl2 interacting protein 3 |
| CaR | calcium-sensing receptor |
| CCR7 | C-C motif chemokine receptor 7 |
| CDKN2B | cyclin-dependent kinase inhibitor 2B |
| C/EBPα/C/EBPβ | CCAAT/enhancer binding protein α/ CCAAT/enhancer binding protein β |
| CLDN1 | claudin-1 |
| cSCC | cutaneous squamous cell carcinoma |
| CTNNB1 | catenin beta 1 |
| DENND1B | DENN domain containing 1B |
| DEPDC1/DEPDC1B | DEP domain containing 1/DEP domain containing 1B |
| DLX5 | distal-less homeobox 5 |
| DND1 | DND microRNA-mediated repression inhibitor 1 |
| EGFR | epidermal growth factor receptor |
| EMP-1 | epithelial membrane protein 1 |
| FGFR2 | fibroblast growth factor receptor 2 |
| FIH1 | factor-inhibiting hypoxia-inducible factor 1 |
| FZD5 | frizzled class receptor 5 |
| GRHL2/ GRHL3 | grainyhead-like transcription factor 2/grainyhead-like transcription factor 3 |
| HAS3 | hyaluronan synthase 3 |
| HIF-1α | hypoxia-inducible factor1 subunit α |
| HPV | human papilloma virus |
| Hras | Harvey rat sarcoma viral oncogene homolog |
| hTERT | human telomerase reverse transcriptase |
| IDH1 | isocitrate dehydrogenase 1 |
| IGF-1R | insulin-like growth factor 1 receptor |
| IL-13Rα1 | interleukin 13 receptor subunit α1 |
| JNK | c-Jun N-terminal kinase |
| K1/K10 | keratin 1/keratin 10 |
| KLF4 | Kruppel-like factor 4 |
| KLKs | kallikrein-related peptidases |
| KLLN | killin, p53-regulated DNA replication inhibitor |
| KRAS | Kirsten rat sarcoma 2 viral oncogene homolog |
| LATS2 | large tumor suppressor kinase 2 |
| lncRNA | long noncoding RNA |
| LOX | lysyl oxidase |
| LXR | liver X receptor |
| miRNA | microRNA |
| MLK3 | mixed-lineage kinase 3 |
| MSH2 | MutS homolog 2 |
| Msi2 | Musashi RNA binding protein 2 |
| MYOT | myotilin |
| NAMPT | nicotinamide phosphoribosyltransferase |
| NKX3.1 | NK3 homeobox 1 |
| ΔNp63α | p63 isoforms truncated at the NH2-terminus lacking the TA domain |
| PAK4 | p21-activated kinase 4 |
| PAR2 | protease-activated receptor type 2 |
| PKC | protein kinase C |
| PLK2 | polo-like kinase 2 |
| PPAR | peroxisome proliferating activated receptor |
| PPP3R1 | protein phosphatase 3 regulatory subunit B, alpha |
| PPP6C | protein phosphatase 6 catalytic subunit |
| PTEN | phosphatase and tensin homolog |
| RASA1 | ras p21 protein activator 1 |
| RB | retinoblastoma 1 |
| RhoBTB1 | rho-related BTB domain containing 1 |
| RRM2 | ribonucleotide reductase regulatory subunit M2 |
| SFMBT1 | scm-like with four mbt domains 1 |
| Skp2 | S-phase kinase associated protein 2 |
| SMAD | Sma and Mad related |
| SNAI2 | snail family transcriptional repressor 2 |
| SOCE | store-operated calcium entry |
| SPRED1/SPRED2 | sprouty-related EVH1 domain containing 1/sprouty-related EVH1 domain containing 2 |
| SPRR1/SPRR2 | small proline-rich protein 1/small proline-rich protein 2 |
| SPRY2/SPRY4 | sprouty homolog 2/sprouty homolog 4 |
| STIM1 | stromal interaction molecule1 |
| STK40 | serine/threonine kinase 40 |
| TAp63 | p63 isoforms with the transactivation domain |
| TGF-β | transforming growth factor-β |
| TGIF1 | transforming growth factor-β-induced factor homeobox 1 |
| Th2 | type 2 T helper |
| TRL4 | toll-like receptor 4 |
| TSLP | thymic stromal lymphopoietin |
| VEGFA | vascular endothelial growth factor A |
| UV | ultraviolet |
References
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Andl, T.; Murchison, E.P.; Liu, F.; Zhang, Y.; Yunta-Gonzalez, M.; Tobias, J.W.; Andl, C.D.; Seykora, J.T.; Hannon, G.J.; Millar, S.E. The miRNA-processing enzyme dicer is essential for the morphogenesis and maintenance of hair follicles. Curr. Biol. 2006, 16, 1041–1049. [Google Scholar] [CrossRef] [Green Version]
- Yi, R.; Pasolli, H.A.; Landthaler, M.; Hafner, M.; Ojo, T.; Sheridan, R.; Sander, C.; O’Carrollm, D.; Stoffel, M.; Tuschl, T.; et al. DGCR8-dependent microRNA biogenesis is essential for skin development. Proc. Natl. Acad. Sci. USA 2009, 106, 498–502. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.R. MicroRNAs as novel players in skin development, homeostasis and disease. Br. J. Dermatol. 2012, 166, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, P.; Ceccoli, J. Advances in the application and impact of microRNAs as therapies for skin disease. BioDrugs 2017, 31, 423–438. [Google Scholar] [CrossRef] [Green Version]
- Singhvi, G.; Manchanda, P.; Krishna Rapalli, V.; Kumar Dubey, S.; Gupta, G.; Dua, K. MicroRNAs as biological regulators in skin disorders. Biomed. Pharmacother. 2018, 108, 996–1004. [Google Scholar] [CrossRef]
- Schmuth, M.; Blunder, S.; Dubrac, S.; Gruber, R.; Moosbrugger-Martinz, V. Epidermal barrier in hereditary ichthyoses, atopic dermatitis, and psoriasis. J. Dtsch. Dermatol. Ges. 2015, 13, 1119–1123. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.P.; Nguyen, G.H.; Jin, H.Z. MicroRNA-143 inhibits IL-13-induced dysregulation of the epidermal barrier-related proteins in skin keratinocytes via targeting to IL-13Rα1. Mol. Cell. Biochem. 2016, 416, 63–70. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Y.; Yuan, W.; Yao, L.; Wang, S.; Jia, Z.; Wu, P.; Li, L.; Wei, P.; Wang, X.; et al. MicroRNA-155–5p is a key regulator of allergic inflammation, modulating the epithelial barrier by targeting PKIα. Cell Death Dis. 2019, 10, 884. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Wang, M.; Li, L.; Zhang, L.; Chan, M.T.V.; Wu, W.K.K. MicroRNAs in atopic dermatitis: A systematic review. J. Cell. Mol. Med. 2020, 24, 5966–5972. [Google Scholar] [CrossRef] [PubMed]
- Elsholz, F.; Harteneck, C.; Muller, W.; Friedland, K. Calcium--a central regulator of keratinocyte differentiation in health and disease. Eur. J. Dermatol. 2014, 24, 650–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, B.C.; Lefort, K.; Mandinova, A.; Antonini, D.; Devgan, V.; Della Gatta, G.; Koster, M.I.; Zhang, Z.; Wang, J.; Tommasi di Vignano, A.; et al. Cross-regulation between Notch and p63 in keratinocyte commitment to differentiation. Genes Dev. 2006, 20, 1028–1042. [Google Scholar] [CrossRef] [Green Version]
- Koh, L.F.; Ng, B.K.; Bertrand, J.; Thierry, F. Transcriptional control of late differentiation in human keratinocytes by TAp63 and Notch. Exp. Dermatol. 2015, 24, 754–760. [Google Scholar] [CrossRef]
- Soares, E.; Zhou, H. Master regulatory role of p63 in epidermal development and disease. Cell. Mol. Life Sci. 2018, 75, 1179–1190. [Google Scholar] [CrossRef] [Green Version]
- Collu, G.M.; Hidalgo-Sastre, A.; Brennan, K. Wnt-Notch signalling crosstalk in development and disease. Cell. Mol. Life Sci. 2014, 71, 3553–3567. [Google Scholar] [CrossRef]
- Ma, X.; Li, L.; Jia, T.; Chen, M.; Liu, G.; Li, C.; Li, N.; Yang, D. miR-203a controls keratinocyte proliferation and differentiation via targeting the stemness-associated factor ΔNp63 and establishing a regulatory circuit with SNAI2. Biochem. Biophys. Res. Commun. 2017, 491, 241–249. [Google Scholar] [CrossRef]
- Chen, H.L.; Chiang, P.C.; Lo, C.H.; Lo, Y.H.; Hsu, D.K.; Chen, H.Y.; Liu, F.T. Galectin-7 regulates keratinocyte proliferation and differentiation through JNK-miR-203-p63 signaling. J. Investig. Dermatol. 2016, 136, 182–191. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J.; Lee, S.B.; Lee, H.B. Oleic acid enhances keratinocytes differentiation via the upregulation of miR-203 in human epidermal keratinocytes. J. Cosmet. Dermatol. 2019, 18, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.J.; Zhang, Z.; Feng, D.; Flagg, M.; O’Loughlin, E.; Wang, D.; Stokes, N.; Fuchs, E.; Yi, R. Rapid and widespread suppression of self-renewal by microRNA-203 during epidermal differentiation. Development 2013, 140, 1882–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chikh, A.; Matin, R.N.; Senatore, V.; Hufbauer, M.; Lavery, D.; Raimondi, C.; Ostano, P.; Mello-Grand, M.; Ghimenti, C.; Bahta, A.; et al. iASPP/p63 autoregulatory feedback loop is required for the homeostasis of stratified epithelia. EMBO J. 2011, 30, 4261–4273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amelio, I.; Lena, A.M.; Viticchiè, G.; Shalom-Feuerstein, R.; Terrinoni, A.; Dinsdale, D.; Russo, G.; Fortunato, C.; Bonanno, E.; Spagnoli, L.G.; et al. miR-24 triggers epidermal differentiation by controlling actin adhesion and cell migration. J. Cell Biol. 2012, 199, 347–363. [Google Scholar] [CrossRef]
- Barbollat-Boutrand, L.; Joly-Tonetti, N.; Dos Santos, M.; Metral, E.; Boher, A.; Masse, I.; Berthier-Vergnes, O.; Bertolino, P.; Damour, O.; Lamartine, J. MicroRNA-23b-3p regulates human keratinocyte differentiation through repression of TGIF1 and activation of the TGF-ß-SMAD2 signalling pathway. Exp. Dermatol. 2017, 26, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.L.; Zhang, T.; Wang, J.; Zhang, D.B.; Zhao, F.; Lin, X.W.; Wang, Z.; Shi, P.; Pang, X.N. MiR-378b promotes differentiation of keratinocytes through NKX3.1. PLoS ONE 2015, 10, e0136049. [Google Scholar] [CrossRef] [PubMed]
- Muther, C.; Jobeili, L.; Garion, M.; Heraud, S.; Thepot, A.; Damour, O.; Lamartine, J. An expression screen for aged-dependent microRNAs identifies miR-30a as a key regulator of aging features in human epidermis. Aging (Albany NY) 2017, 9, 2376–2396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, A.; Powell, A.K.; Sexton, D.W.; Parsons, J.L.; Reynolds, N.J.; Ross, K. microRNA-184 is induced by store-operated calcium entry and regulates early keratinocyte differentiation. J. Cell Physiol. 2020, 10, 6854–6861. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, J.; Rütze, M.; Walz, N.; Gallinat, S.; Wenck, H.; Deppert, W.; Grundhoff, A.; Knott, A. A comprehensive analysis of microRNA expression during human keratinocyte differentiation in vitro and in vivo. J. Investig. Dermatol. 2011, 131, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Neu, J.; Dziunycz, P.J.; Dzung, A.; Lefort, K.; Falke, M.; Denzler, R.; Freiberger, S.N.; Iotzova-Weiss, G.; Kuzmanov, A.; Levesque, M.P.; et al. miR-181a decelerates proliferation in cutaneous squamous cell Carcinoma by targeting the proto-oncogene KRAS. PLoS ONE 2017, 12, e0185028. [Google Scholar] [CrossRef] [Green Version]
- Manca, S.; Magrelli, A.; Cialfi, S.; Lefort, K.; Ambra, R.; Alimandi, M.; Biolcati, G.; Uccelletti, D.; Palleschi, C.; Screpanti, I.; et al. Oxidative stress activation of miR-125b is part of the molecular switch for Hailey-Hailey disease manifestation. Exp. Dermatol. 2011, 20, 932–937. [Google Scholar] [CrossRef]
- Kim, K.H.; Cho, E.G.; Yu, S.J.; Kang, H.; Kim, Y.J.; Kim, S.H.; Lee, T.R. ΔNp63 intronic miR-944 is implicated in the ΔNp63-mediated induction of epidermal differentiation. Nucleic Acids Res. 2015, 43, 7462–7479. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Sulpice, E.; Obeid, P.; Benzina, S.; Kermarrec, F.; Combe, S.; Gidrol, X. The miR-17 family links p63 protein to MAPK signaling to promote the onset of human keratinocyte differentiation. PLoS ONE 2012, 7, e45761. [Google Scholar] [CrossRef] [PubMed]
- Nagosa, S.; Leesch, F.; Putin, D.; Bhattacharya, S.; Altshuler, A.; Serror, L.; Amitai-Lange, A.; Nasser, W.; Aberdam, E.; Rouleau, M.; et al. microRNA-184 induces a commitment switch to epidermal differentiation. Stem Cell Rep. 2017, 9, 1991–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefort, K.; Brooks, Y.; Ostano, P.; Cario-André, M.; Calpini, V.; Guinea-Viniegra, J.; Albinger-Hegyi, A.; Hoetzenecker, W.; Kolfschoten, I.; Wagner, E.F.; et al. A miR-34a-SIRT6 axis in the squamous cell differentiation network. EMBO J. 2013, 32, 2248–2263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Z.; Liu, D.; Peng, Y.; Li, J.; Zhang, Z.; Ning, P. Differential microRNA expression profile comparison between epidermal stem cells and differentiated keratinocytes. Mol. Med. Rep. 2015, 11, 2285–2291. [Google Scholar] [CrossRef] [PubMed]
- Panatta, E.; Lena, A.M.; Mancini, M.; Affinati, M.; Smirnov, A.; Annicchiarico-Petruzzelli, M.; Piro, M.C.; Campione, E.; Bianchi, L.; Mazzanti, C.; et al. Kruppel-like factor 4 regulates keratinocyte senescence. Biochem. Biophys. Res. Commun. 2018, 499, 389–395. [Google Scholar] [CrossRef]
- Sonkoly, E.; Wei, T.; Pavez Loriè, E.; Suzuki, H.; Kato, M.; Törmä, H.; Ståhle, M.; Pivarcsi, A. Protein kinase C-dependent upregulation of miR-203 induces the differentiation of human keratinocytes. J. Investig. Dermatol. 2010, 130, 124–134. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Orfanidis, K.; Xu, N.; Janson, P.; Ståhle, M.; Pivarcsi, A.; Sonkoly, E. The expression of microRNA-203 during human skin morphogenesis. Exp. Dermatol. 2010, 19, 854–856. [Google Scholar] [CrossRef] [Green Version]
- Buschke, S.; Stark, H.J.; Cerezo, A.; Prätzel-Wunder, S.; Boehnke, K.; Kollar, J.; Langbein, L.; Heldin, C.H.; Boukamp, P. A decisive function of transforming growth factor-β/Smad signaling in tissue morphogenesis and differentiation of human HaCaT keratinocytes. Mol. Biol. Cell 2011, 22, 782–794. [Google Scholar] [CrossRef]
- Lee, S.E.; Lee, S.H. Skin Barrier and Calcium. Ann. Dermatol. 2018, 30, 265–275. [Google Scholar] [CrossRef]
- Moses, M.A.; George, A.L.; Sakakibara, N.; Mahmood, K.; Ponnamperuma, R.M.; King, K.E.; Weinberg, W.C. Molecular mechanisms of p63-mediated squamous cancer pathogenesis. Int. J. Mol. Sci. 2019, 20, 3590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, H.; Xiao, H. The Role of the ATP2C1 gene in Hailey-Hailey disease. Cell. Mol. Life Sci. 2017, 74, 3687–3696. [Google Scholar] [CrossRef] [PubMed]
- Antonini, D.; Russo, M.T.; De Rosa, L.; Gorrese, M.; Del Vecchio, L.; Missero, C. Transcriptional repression of miR-34 family contributes to p63-mediated cell cycle progression in epidermal cells. J. Investig. Dermatol. 2010, 130, 1249–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kypriotou, M.; Huber, M.; Hohl, D. The human epidermal differentiation complex: Cornified envelope precursors, S100 proteins and the ’fused genes’ family. Exp. Dermatol. 2012, 21, 643–649. [Google Scholar] [CrossRef]
- Pan, M.; Huang, Y.; Zhu, X.; Lin, X.; Luo, D. miR-125b-mediated regulation of cell proliferation through the Jagged-1/Notch signaling pathway by inhibiting BRD4 expression in psoriasis. Mol. Med. Rep. 2019, 19, 5227–5236. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.; Brodin, P.; Wei, T.; Meisgen, F.; Eidsmo, L.; Nagy, N.; Kemeny, L.; Mona Ståhle, M.; Sonkoly, E.; Pivarcsi, A. MiR-125b, a microRNA downregulated in psoriasis, modulates keratinocyte proliferation by targeting FGFR2. J. Investig. Dermatol. 2011, 131, 1521–1529. [Google Scholar] [CrossRef]
- Zheng, Y.; Cai, B.; Li, X.; Li, D.; Yin, G. MiR-125b-5p and miR-181b-5p inhibit keratinocyte proliferation in skin by targeting Akt3. Eur. J. Pharmacol. 2019, 862, 172659. [Google Scholar] [CrossRef]
- Feng, C.; Bai, M.; Yu, N.Z.; Wang, X.J.; Liu, Z. MicroRNA-181b negatively regulates the proliferation of human epidermal keratinocytes in psoriasis through targeting TLR4. J. Cell. Mol. Med. 2017, 21, 278–285. [Google Scholar] [CrossRef]
- Wang, R.; Zhao, Z.; Zheng, L.; Xing, X.; Ba, W.; Zhang, J.; Huang, M.; Zhu, W.; Liu, B.; Meng, X.; et al. MicroRNA-520a suppresses the proliferation and mitosis of HaCaT cells by inactivating protein kinase B. Exp. Ther. Med. 2017, 14, 6207–6212. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, X.; Wang, L.; Ma, W.; Sun, Q. miR-320b is down-regulated in psoriasis and modulates keratinocyte proliferation by targeting AKT3. Inflammation 2018, 41, 2160–2170. [Google Scholar] [CrossRef]
- Yu, X.; An, J.; Hua, Y.; Li, Z.; Yan, N.; Fan, W.; Su, C. MicroRNA-194 regulates keratinocyte proliferation and differentiation by targeting Grainyhead-like 2 in psoriasis. Pathol. Res. Pract. 2017, 213, 89–97. [Google Scholar] [CrossRef]
- Zhu, H.; Hou, L.; Liu, J.; Li, Z. MiR-217 is down-regulated in psoriasis and promotes keratinocyte differentiation via targeting GRHL2. Biochem. Biophys. Res. Commun. 2016, 471, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.J.; Chu, R.Q.; Ma, J.; Wang, Z.X.; Zhang, G.J.; Yang, X.F.; Zhi Song, Z.; Ma, Y.Y. MicroRNA138 regulates keratin 17 protein expression to affect HaCaT cell proliferation and apoptosis by targeting hTERT in psoriasis vulgaris. Biomed. Pharmacother. 2017, 85, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Su, J.; Li, F.; Chen, X.; Zhang, G. MiR-150 regulates human keratinocyte proliferation in hypoxic conditions through targeting HIF-1α and VEGFA: Implications for psoriasis treatment. PLoS ONE 2017, 12, e0175459. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.J.; Qiao, M.; Li, R.H.; Zhao, X.T.; Wang, X.Y.; Sun, Q. Downregulation of miR-145–5p contributes to hyperproliferation of keratinocytes and skin inflammation in psoriasis. Br. J. Dermatol. 2019, 180, 365–372. [Google Scholar] [CrossRef]
- Li, R.; Qiao, M.; Zhao, X.; Yan, J.; Wang, X.; Sun, Q. MiR-20a-3p regulates TGF-β1/Survivin pathway to affect keratinocytes proliferation and apoptosis by targeting SFMBT1 in vitro. Cell Signal. 2018, 49, 95–104. [Google Scholar] [CrossRef]
- Rongna, A.; Yu, P.; Hao, S.; Li, Y. MiR-876–5p Suppresses cell proliferation by targeting angiopoietin-1 in the psoriasis. Biomed. Pharmacother. 2018, 103, 1163–1169. [Google Scholar]
- Shen, H.; Tian, Y.; Yao, X.; Liu, W.; Zhang, Y.; Yang, Z. MiR-99a inhibits keratinocyte proliferation by targeting Frizzled-5 (FZD5) / FZD8 through β-catenin signaling in psoriasis. Pharmazie 2017, 72, 461–467. [Google Scholar]
- Tang, L.; He, S.; Zhu, Y.; Feng, B.; Su, Z.; Liu, B.; Xu, F.; Wang, X.; Liu, H.; Li, C.; et al. Downregulated miR-187 contributes to the keratinocytes hyperproliferation in psoriasis. J. Cell. Physiol. 2019, 234, 3661–3674. [Google Scholar] [CrossRef]
- Zhao, X.; Li, R.; Qiao, M.; Yan, J.; Sun, Q. MiR-548a-3p promotes keratinocyte proliferation targeting PPP3R1 after being induced by IL-22. Inflammation 2018, 41, 496–504. [Google Scholar] [CrossRef]
- Shen, H.; Zeng, B.; Wang, C.; Tang, X.; Wang, H.; Liu, W.; Yang, Z. MiR-330 inhibits IL-22-induced keratinocyte proliferation through targeting CTNNB1. Biomed. Pharmacother. 2017, 91, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yi, X.; Guo, S.; Shi, Q.; Wei, C.; Li, X.; Gao, L.; Wang, G.; Gao, T.; Wang, L.; et al. A single-nucleotide polymorphism of miR-146a and psoriasis: An association and functional study. J. Cell. Mol. Med. 2014, 18, 2225–2234. [Google Scholar] [CrossRef]
- Peta, E.; Cappellesso, R.; Masi, G.; Sinigaglia, A.; Trevisan, M.; Grassi, A.; Di Camillo, B.; Vassarotto, E.; Fassina, A.; Palù, G.; et al. Down-regulation of microRNA-146a is associated with high-risk human papillomavirus infection and epidermal growth factor receptor overexpression in penile squamous cell carcinoma. Hum. Pathol. 2017, 61, 33–40. [Google Scholar] [PubMed]
- Wu, P.; Cao, Y.; Zhao, R.; Wang, Y. miR-96–5p regulates wound healing by targeting BNIP3/FAK pathway. J. Cell. Biochem. 2019, 120, 12904–12911. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Fang, R.; Gong, Q.; Wang, J. miR-99b suppresses IGF-1R expression and contributes to inhibition of cell proliferation in human epidermal keratinocytes. Biomed. Pharmacother. 2015, 75, 159–164. [Google Scholar] [CrossRef]
- Marthaler, A.M.; Podgorska, M.; Feld, P.; Fingerle, A.; Knerr-Rupp, K.; Grässer, F.; Smola, H.; Roemer, K.; Ebert, E.; Kim, Y.J.; et al. Identification of C/EBPα as a novel target of the HPV8 E6 protein regulating miR-203 in human keratinocytes. PLoS Pathog. 2017, 13, e1006406. [Google Scholar] [CrossRef]
- Meisgen, F.; Xu, N.; Wei, T.; Janson, P.C.; Obad, S.; Broom, O.; Nagy, N.; Kauppinen, S.; Kemény, L.; Ståhle, M.; et al. MiR-21 Is up-regulated in psoriasis and suppresses T cell apoptosis. Exp. Dermatol. 2012, 21, 312–314. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.-Y.; Zhang, K.; Lu, W.J.; Xu, G.W.; Zhang, J.F.; Tang, Z.L. LncRNA MEG3 influences the proliferation and apoptosis of psoriasis epidermal cells by targeting miR-21/caspase-8. BMC Mol. Cell Biol. 2019, 20, 46. [Google Scholar] [CrossRef]
- Wang, S.Y.; Kim, H.; Kwak, G.; Jo, S.D.; Cho, D.; Yang, Y.; Kwon, I.C.; Jeong, J.H.; Kim, S.H. Development of microRNA-21 mimic nanocarriers for the treatment of cutaneous wounds. Theranostics 2020, 10, 3240–3253. [Google Scholar] [CrossRef]
- Yang, C.; Luo, L.; Bai, X.; Shen, K.; Liu, K.; Jing Wang, J.; Hu, D. Highly-expressed micoRNA-21 in adipose derived stem cell exosomes can enhance the migration and proliferation of the HaCaT cells by increasing the MMP-9 expression through the PI3K/AKT pathway. Arch. Biochem. Biophys. 2020, 681, 108259. [Google Scholar] [CrossRef]
- Bhandari, A.; Gordon, W.; Dizon, D.; Hopkin, A.S.; Gordon, E.; Yu, Z.; Andersen, B. The grainyhead transcription factor Grhl3/Get1 suppresses miR-21 expression and tumorigenesis in skin: Modulation of the miR-21 target MSH2 by RNA-binding protein DND1. Oncogene 2013, 32, 1497–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.; Xu, Z.; Lou, F.; Zhang, L.; Ke, F.; Bai, J.; Liu, Z.; Liu, J.; Wang, H.; Zhu, H.; et al. NF-κB-induced microRNA-31 promotes epidermal hyperplasia by repressing Protein Phosphatase 6 in psoriasis. Nat. Commun. 2015, 6, 7652. [Google Scholar] [CrossRef] [Green Version]
- Luan, L.; Shi, J.; Yu, Z.; Andl, T. The Major miR-31 target genes STK40 and LATS2 and their implications in the regulation of keratinocyte growth and hair differentiation. Exp. Dermatol. 2017, 26, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, X.I.; Wang, A.; Meisgen, F.; Pivarcsi, A.; Sonkoly, E.; Ståhle, M.; Landén, N.X. MicroRNA-31 promotes skin wound healing by enhancing keratinocyte proliferation and migration. J. Investig. Dermatol. 2015, 135, 1676–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Ma, X.; Su, Y.; Song, Y.; Tian, Y.; Yuan, S.; Zhang, X.; Yang, D.; Hao Zhang, H.; Shuai, J.; et al. MiR-31 mediates inflammatory signaling to promote re-epithelialization during skin wound healing. J. Investig. Dermatol. 2018, 138, 2253–2263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, N.; Zhou, Y.; Lian, X.; Tu, Y. MicroRNA-31 functions as an oncogenic microRNA in cutaneous squamous cell carcinoma cells by targeting RhoTBT1. Oncol. Lett. 2017, 13, 1078–1082. [Google Scholar] [CrossRef]
- Wang, C.; Zong, J.; Li, Y.; Wang, X.; Du, W.; Li, L. MiR-744–3p regulates keratinocyte proliferation and differentiation via targeting KLLN in psoriasis. Exp. Dermatol. 2019, 28, 283–291. [Google Scholar] [CrossRef]
- Zhang, W.; Yi, X.; An, Y.; Guo, S.; Li, S.; Song, P.; Chang, Y.; Zhang, S.; Gao, T.; Wang, G.; et al. MicroRNA-17–92 cluster promotes the proliferation and the chemokine production of keratinocytes: Implication for the pathogenesis of psoriasis. Cell Death Dis. 2018, 9, 567. [Google Scholar] [CrossRef]
- Xiong, Y.; Chen, H.; Liu, L.; Lu, L.; Wang, Z.; Tian, F.; Zhao, Y. microRNA-130a promotes human keratinocyte viability and migration and inhibits apoptosis through direct regulation of STK40-mediated NF-κB pathway and indirect regulation of SOX9-meditated JNK/MAPK pathway: A potential role in psoriasis. DNA Cell Biol. 2017, 36, 219–226. [Google Scholar] [CrossRef]
- Jiang, M.; Ma, W.; Gao, Y.; Jia, K.; Zhang, Y.; Liu, H.; Sun, Q. IL-22-induced miR-122–5p promotes keratinocyte proliferation by targeting Sprouty2. Exp. Dermatol. 2017, 26, 368–374. [Google Scholar] [CrossRef]
- Wang, R.; Wang, F.F.; Cao, H.W.; Yang, J.Y. MiR-223 regulates proliferation and apoptosis of IL-22-stimulated HaCat human keratinocyte cell lines via the PTEN/Akt pathway. Life Sci. 2019, 230, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhang, G. MicroRNA-17–3p promotes keratinocyte cells growth and metastasis via targeting MYOT and regulating Notch1/NF-κB pathways. Pharmazie 2017, 72, 543–549. [Google Scholar] [PubMed]
- Chang, L.; Liang, J.; Xia, X.; Chen, X. miRNA-126 enhances viability, colony formation, and migration of keratinocytes HaCaT cells by regulating PI3 K/AKT signaling pathway. Cell Biol. Int. 2019, 43, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.K.; Chen, W.; Park, N.-H. Regulation of epithelial cell proliferation, differentiation, and plasticity by Grainyhead-Like 2 during oral carcinogenesis. Crit. Rev. Oncog. 2018, 23, 201–217. [Google Scholar] [CrossRef]
- Peng, H.; Kaplan, N.; Hamanaka, R.B.; Katsnelson, J.; Blatt, H.; Yang, W.; Hao, L.; Bryar, P.J.; Johnson, R.S.; Getsios, S.; et al. microRNA-31/factor-inhibiting hypoxia-inducible factor 1 nexus regulates keratinocyte differentiation. Proc. Natl. Acad. Sci. USA 2012, 109, 14030–14034. [Google Scholar] [CrossRef] [Green Version]
- Riemondy, K.; Wang, X.; Torchia, E.C.; Roop, D.R.; Rui Yi, R. MicroRNA-203 represses selection and expansion of oncogenic Hras transformed tumor initiating cells. Elife 2015, 4, e07004. [Google Scholar] [CrossRef]
- Hitomi, K. Transglutaminases in skin epidermis. Eur. J. Dermatol. 2005, 15, 313–319. [Google Scholar]
- van der Velden, J.J.; van Geel, M.; Nellen, R.G.; Jonkman, M.F.; McGrath, J.A.; Nanda, A.; Sprecher, E.; van Steensel, M.A.; McLean, W.H.; Cassidy, A.J. Novel TGM5 mutations in acral peeling skin syndrome. Exp. Dermatol. 2015, 24, 285–289. [Google Scholar] [CrossRef]
- Karim, N.; Phinney, B.S.; Salemi, M.; Wu, P.W.; Naeem, M.; Rice, R.H. Human stratum corneum proteomics reveals cross-linking of a broad spectrum of proteins in cornified envelopes. Exp. Dermatol. 2019, 28, 618–622. [Google Scholar] [CrossRef] [Green Version]
- Takeda, M.; Nomura, T.; Sugiyama, T.; Miyauchi, T.; Suzuki, S.; Fujita, Y.; Shimizu, H. Compound heterozygous missense mutations p.Leu207Pro and p.Tyr544Cys in TGM1 cause a severe form of lamellar ichthyosis. J. Dermatol. 2018, 45, 1463–1467. [Google Scholar] [CrossRef]
- Li, X.; Wu, Y.; Xie, F.; Zhang, F.; Zhang, S.; Zhou, J.; Chen, D.; Liu, A. miR-339-5p negatively regulates loureirin A-induced hair follicle stem cell differentiation by targeting DLX5. Mol. Med. Rep. 2018, 18, 1279–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Ghatak, S.; El Masry, M.S.; Das, A.; Liu, Y.; Roy, S.; Lee, R.J.; Sen, C.K. Topical lyophilized targeted lipid nanoparticles in the restoration of skin barrier function following burn wound. Mol. Ther. 2018, 26, 2178–2188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.E.; Leung, D.Y.; Boguniewicz, M.; Howell, M.D. Loricrin and involucrin expression is down-regulated by Th2 cytokines through STAT-6. Clin. Immunol. 2008, 126, 332–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, M.D.; Kim, B.E.; Gao, P.; Grant, A.V.; Boguniewicz, M.; DeBenedetto, A.; Schneider, L.; Beck, L.A.; Barnes, K.C.; Leung, D.Y. Cytokine modulation of atopic dermatitis filaggrin skin expression. J. Allergy Clin. Immunol. 2009, 124, R7–R12. [Google Scholar] [CrossRef]
- Li, H.M.; Xiao, Y.J.; Min, Z.S.; Tan, C. Identification and interaction analysis of key genes and microRNAs in atopic dermatitis by bioinformatics analysis. Clin. Exp. Dermatol. 2019, 44, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Qiao, M.; Li, R.; Zhao, X.; Yan, J.; Sun, Q. Up-regulated lncRNA-MSX2P1 promotes the growth of IL-22-stimulated keratinocytes by inhibiting miR-6731–5p and activating S100A7. Exp. Cell Res. 2018, 363, 243–254. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xu, H.; Li, C.; Zhang, X.; Zhou, P.; Xiao, X.; Zhang, W.; Wu, Y.; Zeng, R.; Wang, B. Nicastrin/miR-30a-3p/RAB31 axis regulates keratinocyte differentiation by impairing EGFR signaling in familial acne inversa. J. Investig. Dermatol. 2019, 139, 124–134. [Google Scholar] [CrossRef] [Green Version]
- Kezic, S.; Jakasa, I. Filaggrin and skin barrier function. Curr. Probl. Dermatol. 2016, 49, 1–7. [Google Scholar]
- Elias, P.M.; Wakefield, J.S. Mechanisms of abnormal lamellar body secretion and the dysfunctional skin barrier in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2014, 134, 781–791. [Google Scholar] [CrossRef] [Green Version]
- Ghatak, S.; Chan, Y.C.; Khanna, S.; Banerjee, J.; Weist, J.; Roy, S.; Sen, C.K. Barrier function of the repaired skin is disrupted following arrest of Dicer in keratinocytes. Mol. Ther. 2015, 23, 1201–1210. [Google Scholar] [CrossRef] [Green Version]
- Ristori, E.; Lopez-Ramirez, M.A.; Narayanan, A.; Hill Teran, G.; Moro, A.; Calvo, C.F.; Thomas, J.L.; Nicoli, S. A Dicer-miR-107 interaction regulates biogenesis of specific miRNAs crucial for neurogenesis. Dev. Cell. 2015, 32, 546–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, Y.; Zhang, R.; Liu, C.; Zhou, L.; Wang, H.; Zhuang, W.; Huang, Y.; Hong, Z. miR-143 inhibits interleukin-13-induced inflammatory cytokine and mucus production in nasal epithelial cells from allergic rhinitis patients by targeting IL13Rα1. Biochem. Biophys. Res. Commun. 2015, 457, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Eckert, R.L.; Lee, K.C. S100A7 (Psoriasin): A story of mice and men. J. Investig. Dermatol. 2006, 126, 1442–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hämäläinen, L.; Kärkkäinen, E.; Takabe, P.; Rauhala, L.; Bart, G.; Kärnä, R.; Pasonen-Seppänen, S.; Oikari, S.; Tammi, M.I.; Tammi, R.H. Hyaluronan metabolism enhanced during epidermal differentiation is suppressed by vitamin C. Br. J. Dermatol. 2018, 179, 651–661. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.-Y. Molecular mechanism of epidermal barrier dysfunction as primary abnormalities. Int. J. Mol. Sci. 2020, 21, 1194. [Google Scholar] [CrossRef] [Green Version]
- Li, C.X.; Li, H.G.; Huang, L.T.; Kong, Y.W.; Chen, F.Y.; Liang, J.Y.; Yu, H.; Yao, Z.R. H19 lncRNA regulates keratinocyte differentiation by targeting miR-130b-3p. Cell Death Dis. 2017, 8, e3174. [Google Scholar] [CrossRef]
- Ahmed, M.I.; Alam, M.; Emelianov, V.U.; Poterlowicz, K.; Patel, A.; Sharov, A.A.; Mardaryev, A.N.; Botchkareva, N.V. MicroRNA-214 controls skin and hair follicle development by modulating the activity of the Wnt pathway. J. Cell Biol. 2014, 207, 549–567. [Google Scholar] [CrossRef] [Green Version]
- Cheong, K.A.; Lee, T.R.; Lee, A.Y. Complementary effect of hydroquinone and retinoic acid on corneocyte desquamation with their combination use. J. Dermatol. Sci. 2017, 87, 192–200. [Google Scholar] [CrossRef] [Green Version]
- Stefansson, K.; Brattsand, M.; Ny, A.; Glas, B.; Egelrud, T. Kallikrein-related peptidase 14 may be a major contributor to trypsin-like proteolytic activity in human stratum corneum. Biol. Chem. 2006, 387, 761–768. [Google Scholar] [CrossRef]
- Demerjian, M.; Hachem, J.P.; Tschachler, E.; Denecker, G.; Declercq, W.; Vandenabeele, P.; Mauro, T.; Hupe, M.; Crumrine, D.; Roelandt, T.; et al. Acute modulations in permeability barrier function regulate epidermal cornification: Role of caspase-14 and the protease-activated receptor type 2. Am. J. Pathol. 2008, 172, 86–97. [Google Scholar] [CrossRef] [Green Version]
- Hardman, M.J.; Liu, K.; Avilion, A.A.; Merritt, A.; Brennan, K.; Garrod, D.R.; Byrne, C. Desmosomal cadherin misexpression alters beta-catenin stability and epidermal differentiation. Mol. Cell. Biol. 2005, 25, 969–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wertz, P. Epidermal Lamellar Granules. Skin Pharmacol. Physiol. 2018, 31, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Ludovici, M.; Kozul, N.; Materazzi, S.; Risoluti, R.; Picardo, M.; Camera, E. Influence of the sebaceous gland density on the stratum corneum lipidome. Sci. Rep. 2018, 8, 1–12. [Google Scholar]
- Sajić, D.; Asiniwasis, R.; Skotnicki-Grant, S. A look at epidermal barrier function in atopic dermatitis: Physiologic lipid replacement and the role of ceramides. Skin Therapy Lett. 2012, 17, 6–9. [Google Scholar] [PubMed]
- Kihara, A. Synthesis and degradation pathways, functions, and pathology of ceramides and epidermal acylceramides. Prog. Lipid Res. 2016, 63, 50–69. [Google Scholar] [CrossRef]
- Takeichi, T.; Hirabayashi, T.; Miyasaka, Y.; Kawamoto, A.; Okuno, Y.; Taguchi, S.; Tanahashi, K.; Murase, C.; Takama, H.; Tanaka, K.; et al. SDR9C7 catalyzes critical dehydrogenation of acylceramides for skin barrier formation. J. Clin. Investig. 2020, 130, 890–903. [Google Scholar] [CrossRef]
- Schmuth, M.; Jiang, Y.J.; Dubrac, S.; Elias, P.M.; Feingold, K.R. Thematic review series: Skin lipids. Peroxisome proliferator-activated receptors and liver X receptors in epidermal biology. J. Lipid Res. 2008, 49, 499–509. [Google Scholar] [CrossRef] [Green Version]
- Ratovitski, E.A. Phospho-ΔNp63α regulates AQP3, ALOX12B, CASP14 and CLDN1 expression through transcription and microRNA modulation. FEBS Lett. 2013, 587, 3581–3586. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.R.; Samborski, A.; Bauersachs, S.; Zouboulis, C.C. Differentially regulated microRNAs during human sebaceous lipogenesis. J. Dermatol. Sci. 2013, 70, 88–93. [Google Scholar] [CrossRef]
- Degueurce, G.; D’Errico, I.; Pich, C.; Ibberson, M.; Schütz, F.; Montagner, A.; Sgandurra, M.; Mury, L.; Jafari, P.; Boda, A.; et al. Identification of a novel PPARβ/δ/miR-21–3p axis in UV-induced skin inflammation. EMBO Mol. Med. 2016, 8, 919–936. [Google Scholar] [CrossRef]
- Urgard, E.; Lorents, A.; Klaas, M.; Padari, K.; Viil, J.; Runnel, T.; Langel, K.; Kingo, K.; Tkaczyk, E.; Lange, L.Ü.; et al. Pre-administration of PepFect6-microRNA-146a nanocomplexes inhibits inflammatory responses in keratinocytes and in a mouse model of irritant contact dermatitis. J. Control. Release 2016, 235, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Ross, K. Towards topical microRNA-directed therapy for epidermal disorders. J. Control. Release 2018, 269, 136–147. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Regulatory Factor | miRNA | Target Molecule | Action Mechanism of miRNA | Reference |
|---|---|---|---|---|
| Calcium ↑ | miR-203 ↑ | SNAI2 and ΔNp63 | Activation of the PKC and AP-1 pathway | [18] |
| p63 | Upregulation of JNK by galectin-7 | [19] | ||
| Increased keratinocyte differentiation with involucrin expression by oleic acid | [20] | |||
| p63, Skp2 and Msi2 | Promotion of cell cycle exit in mouse skin | [21] | ||
| miR-574 ↑ | p63 | As direct targets of iASPP | [22] | |
| miR-720 ↑ | ||||
| miR-24 ↑ | PAK4 | Control of actin cable formation | [23] | |
| miR-23b-3p ↑ | TGIF1 | Interference in TGF-β/SMAD signaling | [24] | |
| miR-378b ↑ | NKX3.1 | [25] | ||
| miR-30a ↑ | LOX, IDH1, AVEN | Barrier function defects in aged epidermis | [26] | |
| miR-184 ↑ | Upregulation of cyclin E and p21 cyclin-dependent kinase inhibitor in a SOCE-dependent manner | [27] | ||
| miR-181a ↑ | cell differentiation under high calcium or UVA exposure | [28,29] | ||
| miR-125b ↑ | p63 | cell differentiation and proliferation in Hailey-Hailey disease | [30] | |
| Notch | ||||
| p63 ↑ | miR-944 ↑ | Upregulation of K1 and K10 by ERK inhibition and p53 upregulation | [31] | |
| p63 ↓ | miR-17/miR-20b /miR-106a ↓ | p21, RB, and JNK2 | Upregulation of K1 and K10 | [32] |
| Notch ↑ | miR-184 ↑ | K15 and FIH1 | Enhancing the Notch pathway | [33] |
| miRNAs targeted by p63 | miR-34a ↑ | SIRT6 | miR-34a and miR-34c as direct targets of p63 | [34,35] |
| miR-34a ↑ | KLK4 | Induction of a senescent phenotype in keratinocytes | [36] |
| miRNA | Target Molecule | Related Skin Diseases | Reference | |
|---|---|---|---|---|
| Change | Name | |||
| ↓ | miR-125b | FGFR2 | Psoriasis | [46] |
| AKT3 | [47] | |||
| miR-181b-5p | AKT3, TRL4 | [47,48] | ||
| miR-520a | AKT | [49] | ||
| miR-320b | AKT3 | [50] | ||
| miR-194 | GRHL2 | [51] | ||
| miR-217 | [52] | |||
| miR-138 | hTERT | [53] | ||
| miR-150 | HIF-1α, VEGFA | [54] | ||
| miR-145-5p | MLK3 | [55] | ||
| miR-20a-3p | SFMBT1 | [56] | ||
| miR-876-5p | ANG-1 | [57] | ||
| miR-99a | FZD5/FZD8 | [58] | ||
| miR-187 | CD276 | [59] | ||
| miR-548a-3p | PPP3R1 | [60] | ||
| miR-330 | CTNNB1 | [61] | ||
| miR-146a | EGFR | Psoriasis, cSCC | [62,63] | |
| miR-96-5p | BNP3 | Wound healing | [64] | |
| miR-181a | KRAS | cSCC | [29] | |
| miR-99b | IGF-1R | Condyloma acuminatum | [65] | |
| miR-203 | ΔNp63 | Epidermodysplasia verruciformis | [66] | |
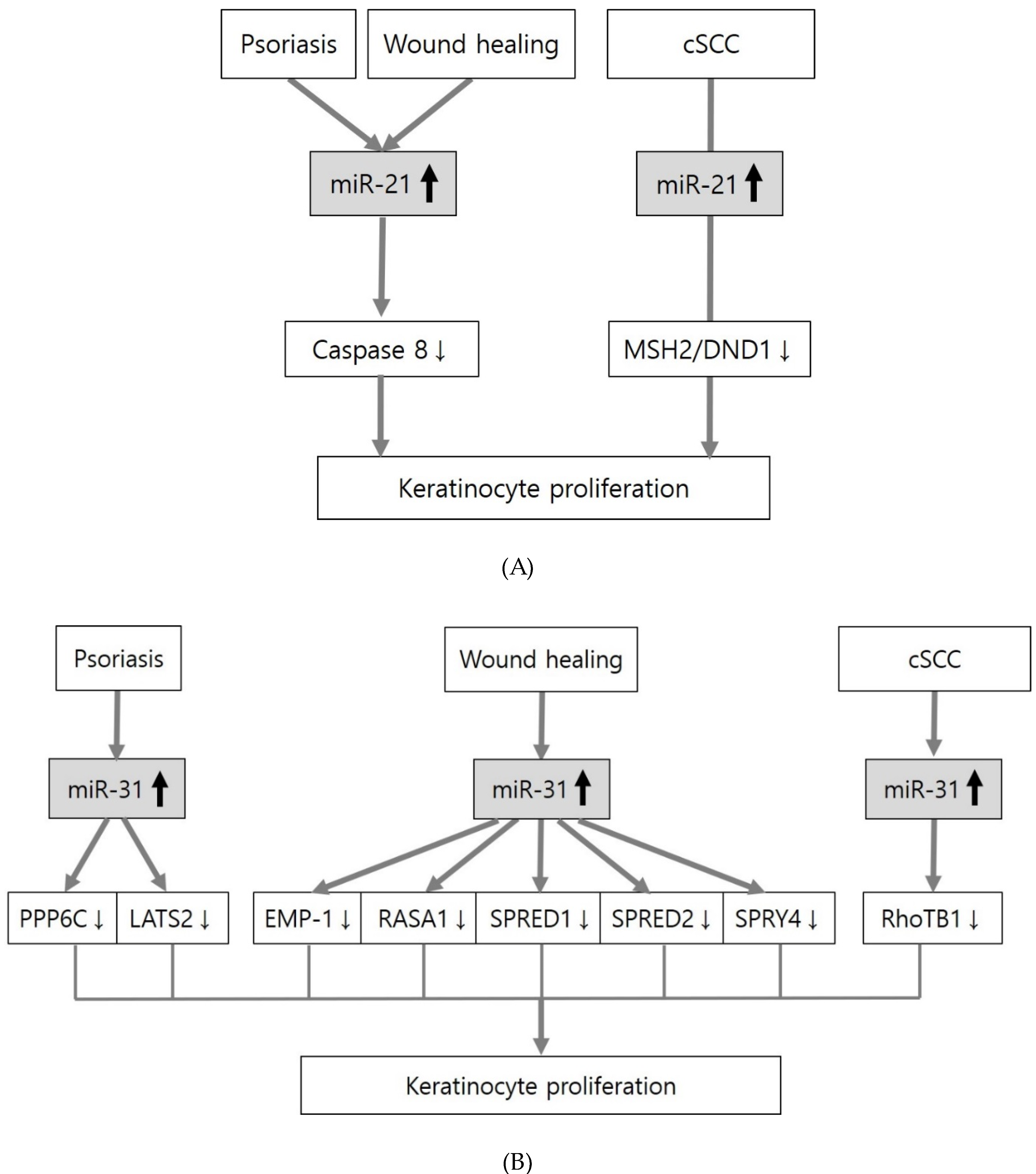
| ↑ | miR-21 | Caspase 8 | Psoriasis | [67,68] |
| Wound healing | [69,70] | |||
| MSH2 | cSCC | [71] | ||
| miR-31 | PPP6C | Psoriasis | [72] | |
| LATS2 | [73] | |||
| EMP-1 | Wound healing | [74] | ||
| RASA1 | [75] | |||
| SPRED1 | ||||
| SPRED2 | ||||
| SPRY4 | ||||
| RhoTB1 | cSCC | [76] | ||
| miR-744-3p | KLLN | Psoriasis | [77] | |
| miR-17-92 | CDKN2B | [78] | ||
| miR-130a | STK40 | [79] | ||
| miR-122-5p | SPRY2 | [80] | ||
| miR-223 | PTEN | [81] | ||
| miR-17-3p | MYOT | Wound healing | [82] | |
| miR-126 | PLK2 | [83] | ||
| miRNA | Target Molecule | Related Skin Disease | Action Mechanism of miRNA | Reference |
|---|---|---|---|---|
| miR-339-5p ↓ | DLX5 | Increased involucrin expression through Wnt/β-catenin signaling pathway activation | [91] | |
| miR-107 ↓ | Dicer | Increased filaggrin and loricrin expression | [92] | |
| miR-203 (C/EBPα/miR-203 pathway) ↓ | ΔNp63α | HPV8 infection | Downregulation of involucrin | [66] |
| miR-143 ↓ | IL-13Rα1 | Atopic dermatitis | Reduced filaggrin/ involucrin /loricrin expression through Th2-derived IL-13 activity stimulation | [12,93,94] |
| Let-7a-5p ↓ | RRM2, CCR7 | Barrier abnormalities | [95] | |
| miR-26a-5p ↓ | HAS3, DEPDC1B, DEPDC1, NAMPT, DENND1B, ADAM19 | [95] | ||
| miR-10a-5p ↑ | HAS3 | [12] | ||
| miR-29b ↑ | BCL2L2 | Barrier abnormalities with Increased IFN-γ-induced keratinocyte apoptosis | [12] | |
| miR-6731-5p ↓ | S100A7 | IL-22-stimulated keratinocyte proliferation | [96] | |
| miR-146a ↑ | EGFR | Terminal differentiation and proliferation inhibition in keratinocytes | [62] | |
| miR-30a-3p ↓ | Familial acne inversa | Abnormal keratinocyte differentiation by accelerated EGFR degradation | [97] |
| miRNA | Target Molecule | Action Mechanism of miRNA | Reference |
|---|---|---|---|
| miR-130b-3p ↓ | Desmoglein 1 ↑ | miR-130b-3p is inhibited by upregulated H19 in keratinocytes treated with calcium | [106] |
| miR-214 ↑ | β-catenin ↓ | β-catenin is linked to desmosomal cadherins, resulting in epidermal barrier dysfunction. | [107] |
| miRNA | Target Molecule | Related Skin Condition | Action Mechanism of miRNA | Reference |
|---|---|---|---|---|
| miR-185-5p ↑ | ALOX12B | Upregulated by p-ΔNp63, resulting in reduced ALOX12B activity. | [117] | |
| miR-203 ↑ | Upregulated by oleic acid, promoting keratinocyte differentiation | [20] | ||
| miR-203 ↑ | Upregulated by linoleic acid, stimulating keratinocyte differentiation | [118] | ||
| miR-574-3p ↑ | ||||
| miR-21-3p ↑ | SMAD7 | UV exposure | Upregulated in a PPARβ/δ-dependent manner Besides regulatory role in immune and inflammatory responses, miR-213p can downregulate caspase-14 expression. (--Arrangement was changed as a center alignment array) | [119] |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, A.-Y. The Role of MicroRNAs in Epidermal Barrier. Int. J. Mol. Sci. 2020, 21, 5781. https://doi.org/10.3390/ijms21165781
Lee A-Y. The Role of MicroRNAs in Epidermal Barrier. International Journal of Molecular Sciences. 2020; 21(16):5781. https://doi.org/10.3390/ijms21165781
Chicago/Turabian StyleLee, Ai-Young. 2020. "The Role of MicroRNAs in Epidermal Barrier" International Journal of Molecular Sciences 21, no. 16: 5781. https://doi.org/10.3390/ijms21165781
APA StyleLee, A. -Y. (2020). The Role of MicroRNAs in Epidermal Barrier. International Journal of Molecular Sciences, 21(16), 5781. https://doi.org/10.3390/ijms21165781